
Synergistic Effect in Neurological Recovery via Anti-Apoptotic Akt Signaling in Umbilical Cord Blood and Erythropoietin Combination Therapy for Neonatal Hypoxic-Ischemic Brain Injury

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
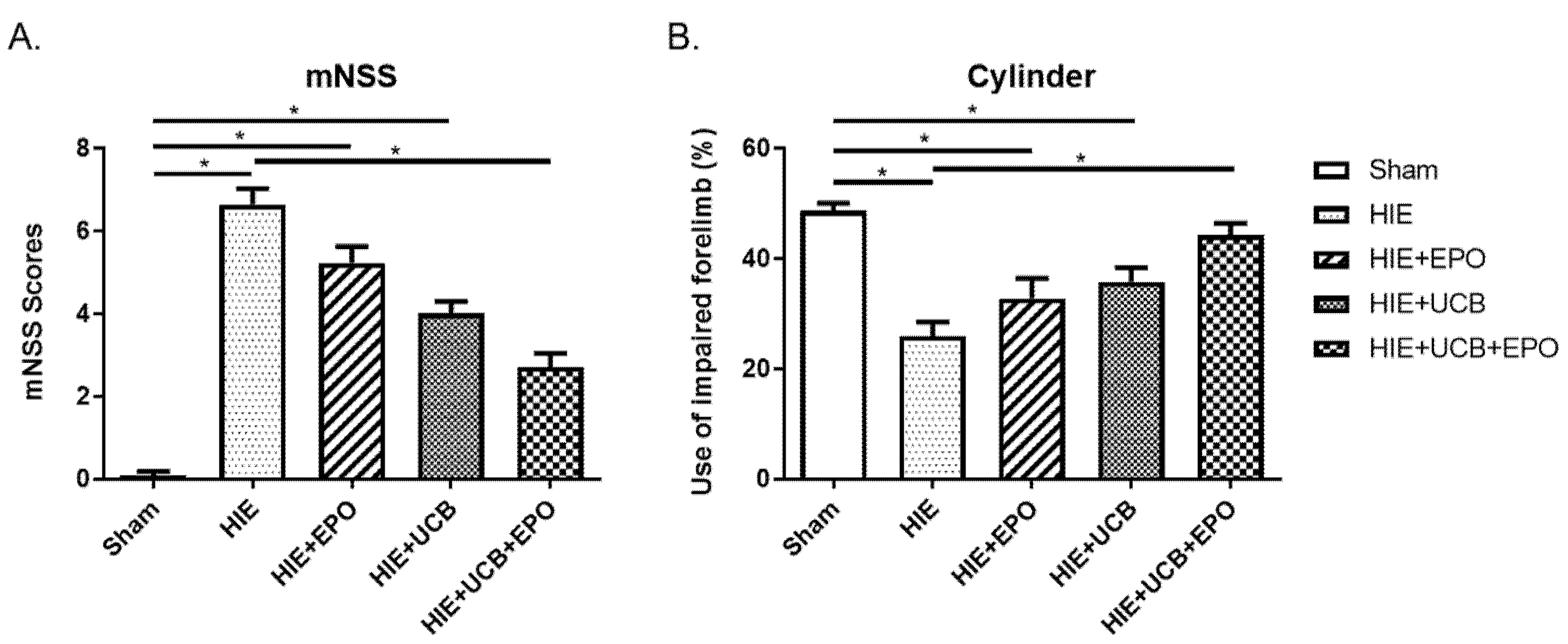
2.1. Neurobehavioral Outcome by rhEPO or hUCB or hUCB+rhEPO Combination Treatment in HIE
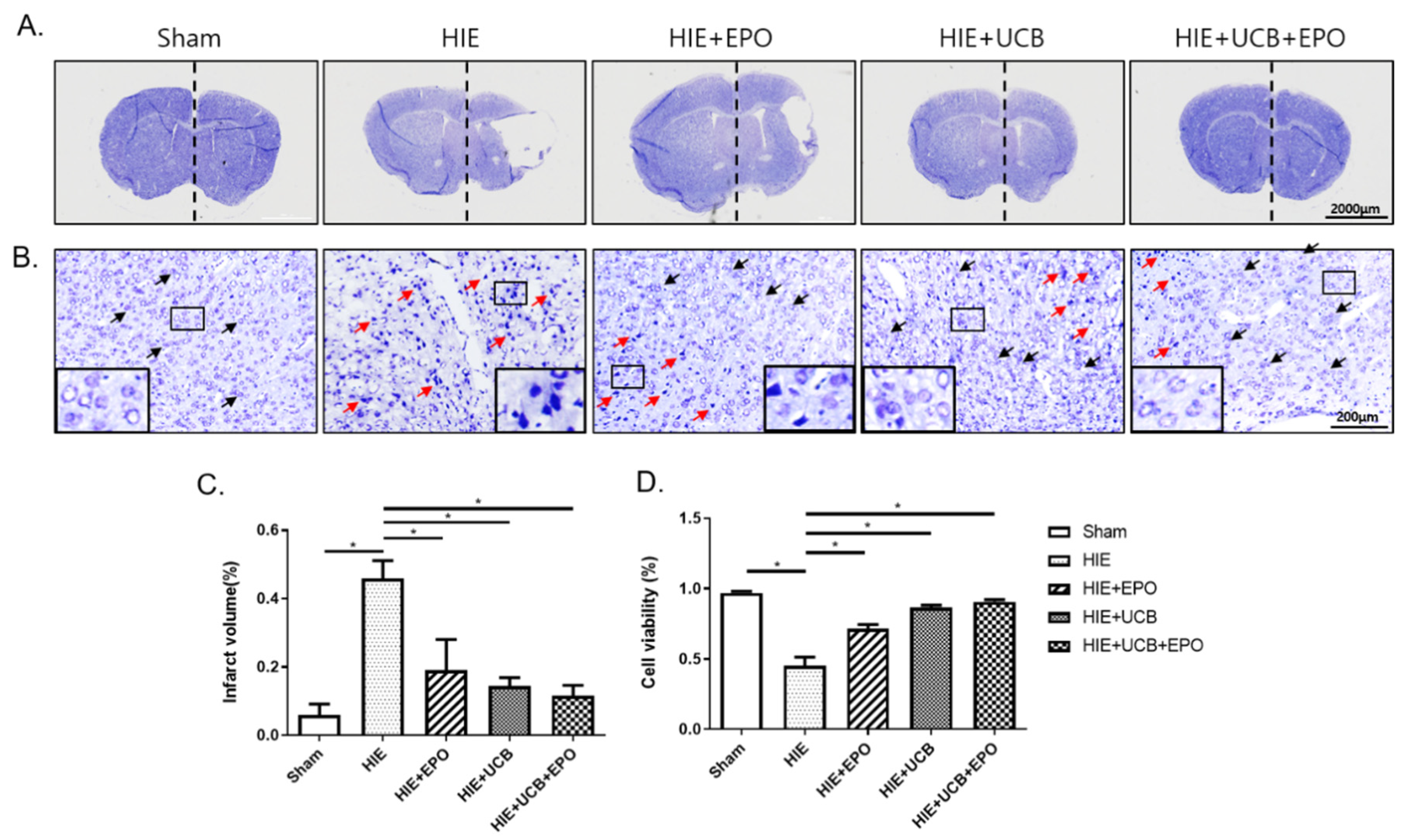
2.2. Neuroprotective Effect by rhEPO or hUCB or hUCB+rhEPO Combination Treatment in HIE
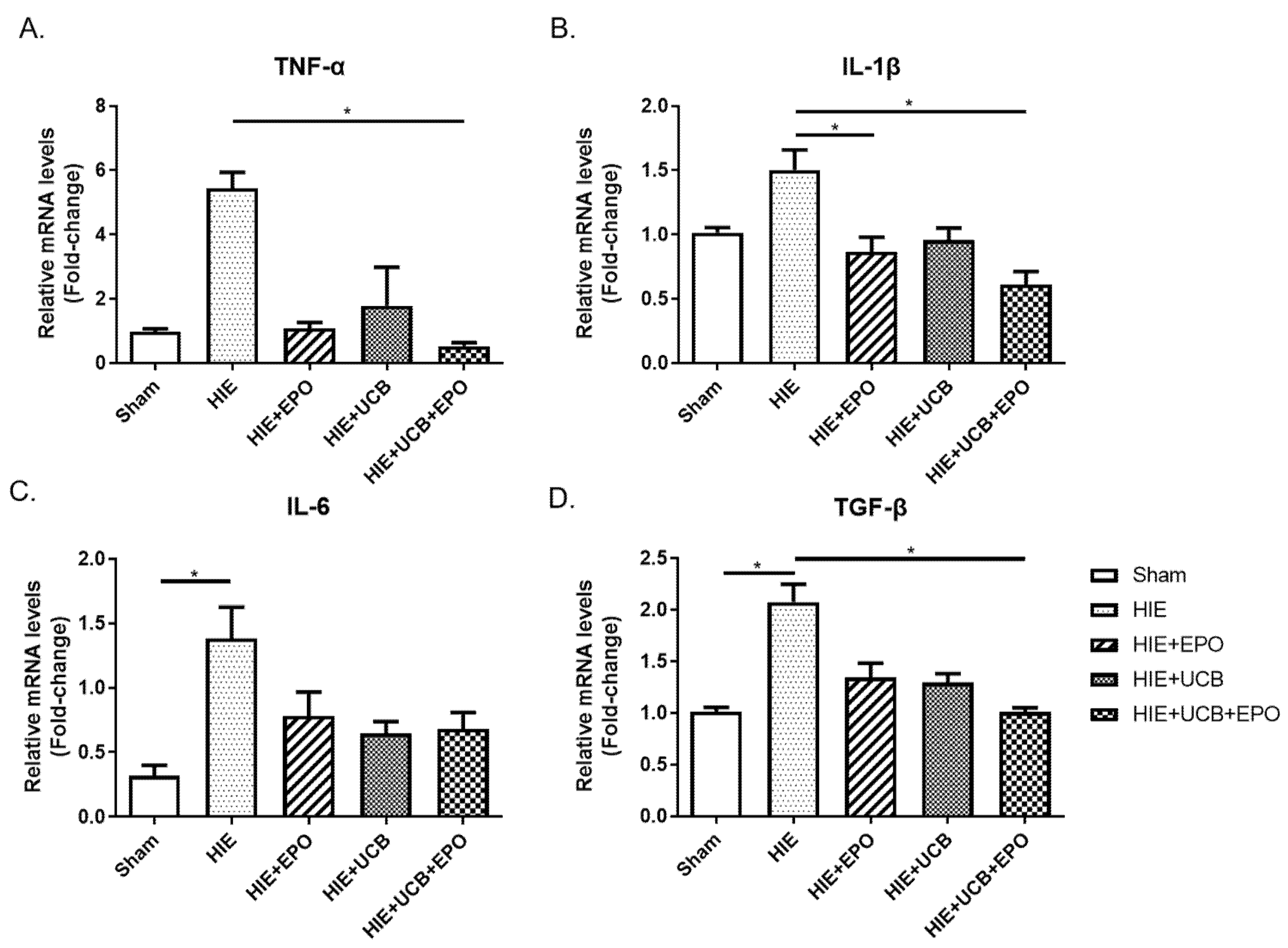
2.3. Reduction of Pro-Inflammatory Gene Expression by rhEPO or hUCB or hUCB+rhEPO Combination Treatment in HIE
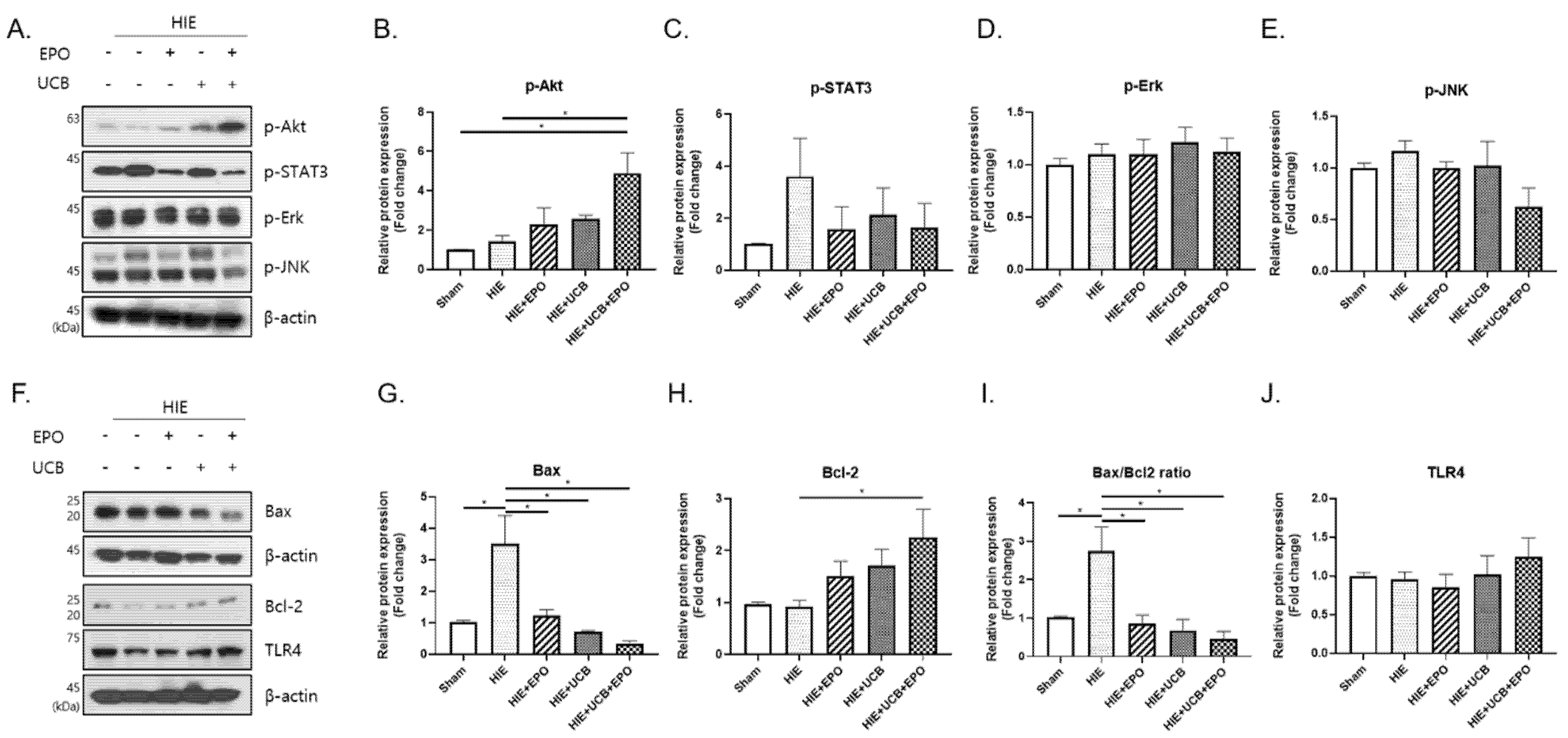
2.4. Up-Regulation of Anti-Apoptotic Effect by hUCB+rhEPO Combination Treatment in HIE
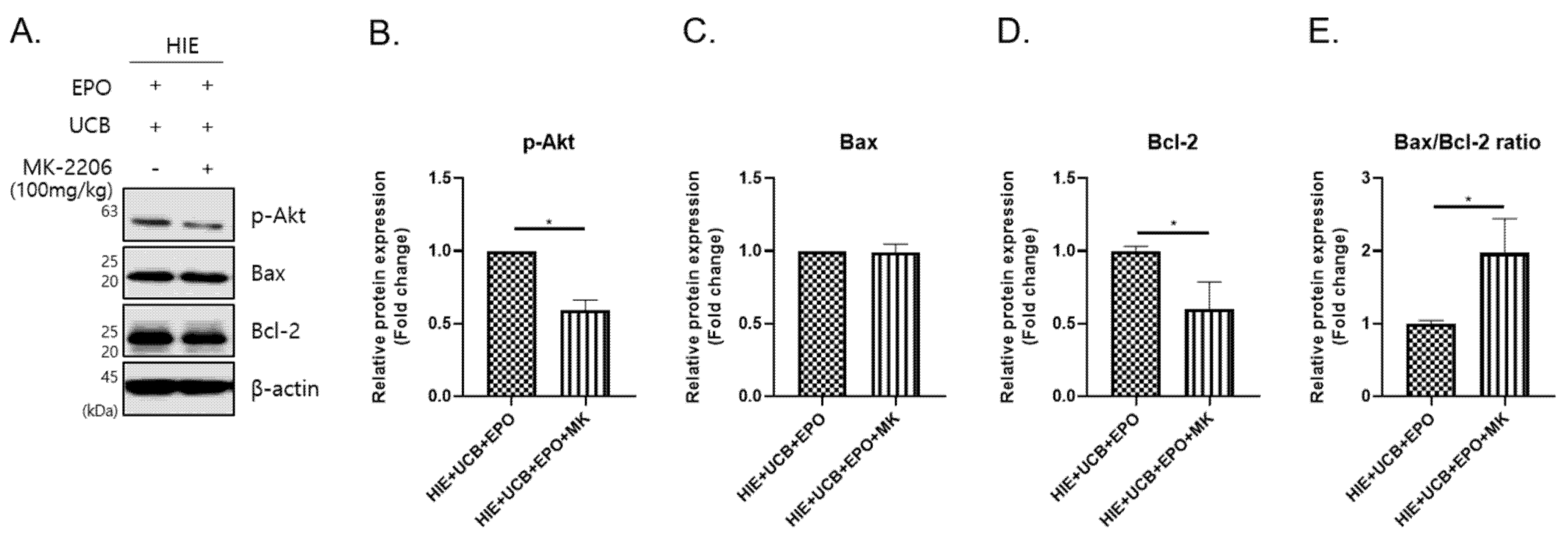
2.5. The MK-2206 Inhibits the Anti-Apoptotic Effect by Akt Phosphorylation in the hUCB+rhEPO Combination Treatment Group
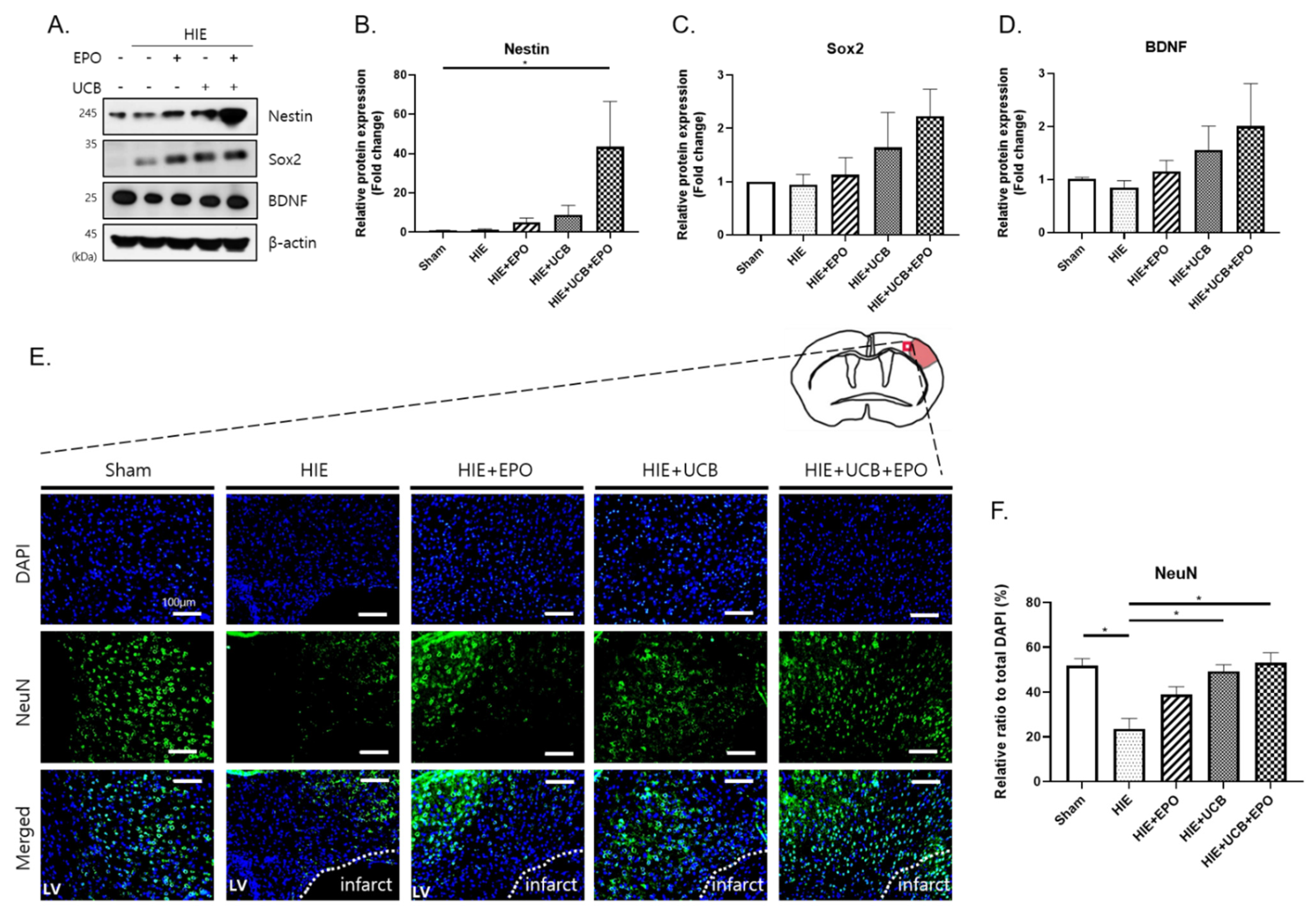
2.6. Enhancement of Neuroregenerative Capability by rhEPO or hUCB or hUCB+rhEPO Combination Treatment in HIE
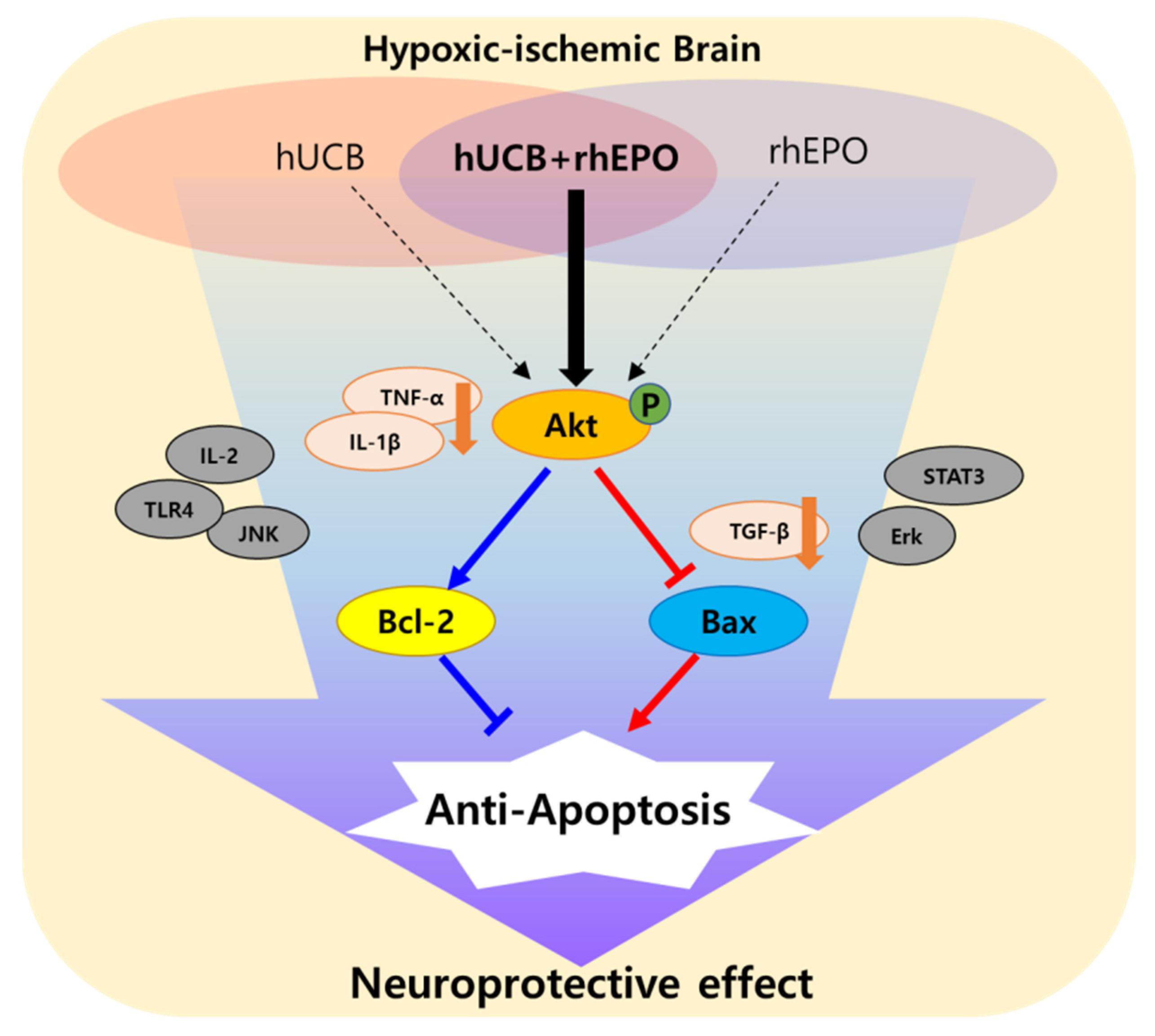
3. Discussion
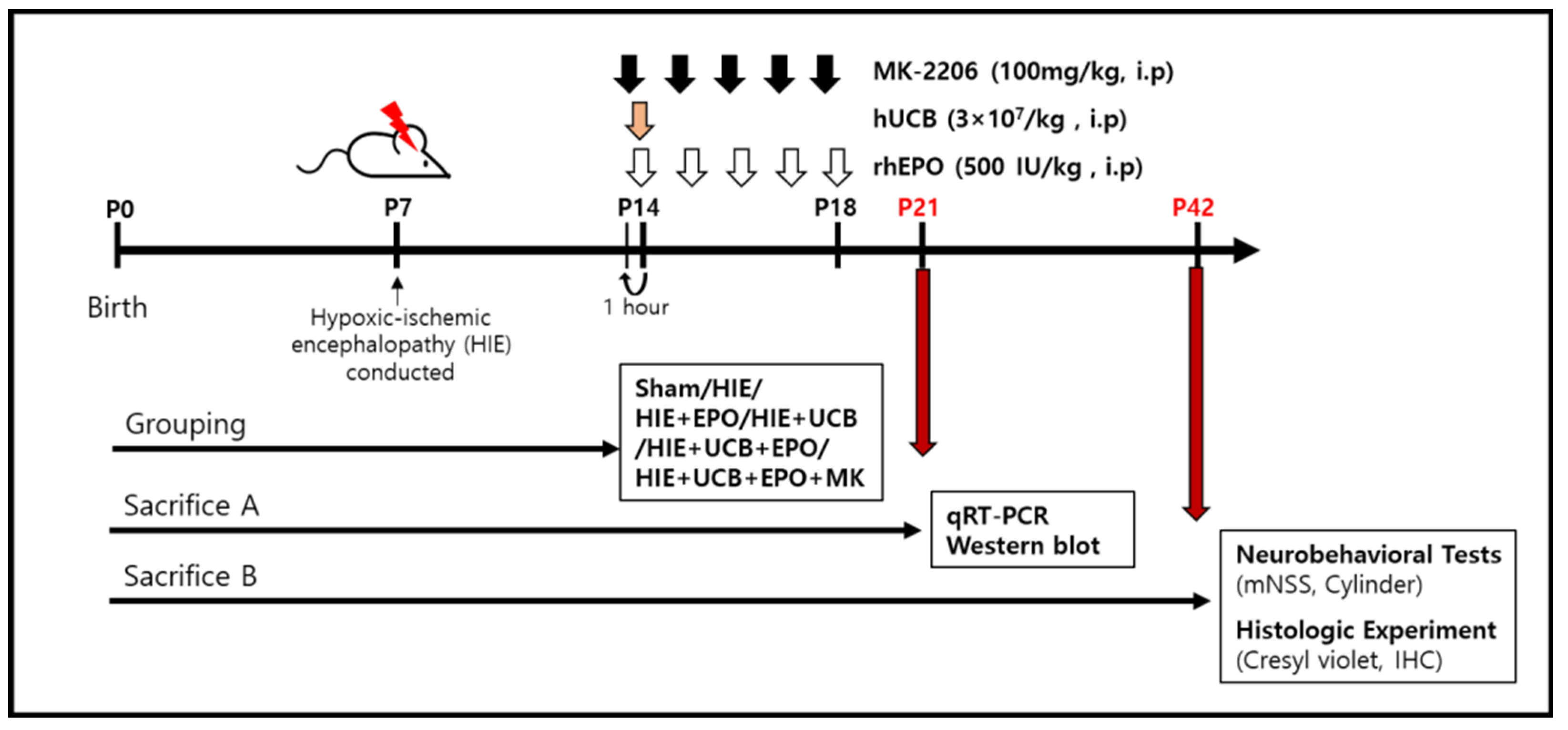
4. Materials and Methods
4.1. Animal
4.2. In Vivo Ischemia Model: Hypoxic–Ischemic Brain Injury (HIE)
4.3. Human Umbilical Cord Blood Cells
4.4. Administration Condition
4.5. Behavior Test
4.6. Histologic Experiment
4.7. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.8. Western Blotting
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferreira, E.R.; Hristova, M. Plasticity in the Neonatal Brain following Hypoxic-Ischaemic Injury. Neural Plast. 2016, 2016, 4901014. [Google Scholar] [CrossRef] [Green Version]
- Fatemi, A.; Wilson, M.A.; Johnston, M.V. Hypoxic-Ischemic Encephalopathy in the Term Infant. Clin. Perinatol. 2009, 36, 835–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perlman, J.M. Pathogenesis of hypoxic-ischemic brain injury. J. Perinatol. 2007, 27, S39–S46. [Google Scholar] [CrossRef] [Green Version]
- Krigger, K.W. Cerebral palsy: An overview. Am. Fam. Physician 2006, 73, 91–100. [Google Scholar]
- Aisen, M.L.; Kerkovich, D.; Mast, J.; Mulroy, S.; Wren, T.A.; Kay, R.M.; Rethlefsen, S.A. Cerebral palsy: Clinical care and neurological rehabilitation. Lancet Neurol. 2011, 10, 844–852. [Google Scholar] [CrossRef]
- Peliowski-Davidovich, A.; Canadian Paediatric Society, F.; Newborn, C. Hypothermia for new-borns with hypoxic ischemic encephalopathy. Paediatr. Child Health 2012, 17, 41–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eggenberger, S.; Boucard, C.; Schoeberlein, A.; Guzman, R.; Limacher, A.; Surbek, D.; Mueller, M. Stem cell treatment and cerebral palsy: Systemic review and meta-analysis. World J. Stem Cells 2019, 11, 891–903. [Google Scholar] [CrossRef]
- Nair, J.; Kumar, V.H.S. Current and Emerging Therapies in the Management of Hypoxic Ischemic Encephalopathy in Neo-nates. Children 2018, 5, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizk, M.; Aziz, J.; Shorr, R.; Allan, D.S. Cell-Based Therapy Using Umbilical Cord Blood for Novel Indications in Regenerative Therapy and Immune Modulation: An Updated Systematic Scoping Review of the Literature. Biol. Blood Marrow Transplant. 2017, 23, 1607–1613. [Google Scholar] [CrossRef] [Green Version]
- McDonald, C.A.; Penny, T.R.; Paton, M.C.B.; Sutherland, A.E.; Nekkanti, L.; Yawno, T.; Cas-tillo-Melendez, M.; Fahey, M.C.; Jones, N.M.; Jenkin, G.; et al. Effects of umbilical cord blood cells, and subtypes, to reduce neuroinflammation following perinatal hypoxic-ischemic brain injury. J. Neuroinflamm. 2018, 15, 47. [Google Scholar] [CrossRef]
- Broxmeyer, H. Biology of cord blood cells and future prospects for enhanced clinical benefit. Cytotherapy 2005, 7, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Min, K.; Song, J.; Kang, J.Y.; Ko, J.; Ryu, J.S.; Kang, M.S.; Jang, S.J.; Kim, S.H.; Oh, D.; Kim, M.K.; et al. Umbilical Cord Blood Therapy Potentiated with Erythropoietin for Children with Cerebral Palsy: A Double-blind, Randomized, Placebo-Controlled Trial. Stem Cells 2013, 31, 581–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, M.; Min, K.; Jang, J.; Kim, S.C.; Kang, M.S.; Jang, S.J.; Lee, J.Y.; Kim, S.H.; Kim, M.K.; An, S.A.; et al. Involvement of Immune Responses in the Efficacy of Cord Blood Cell Thera-py for Cerebral Palsy. Stem Cells Dev. 2015, 24, 2259–2268. [Google Scholar] [CrossRef] [PubMed]
- Penny, T.R.; Sutherland, A.E.; Mihelakis, J.G.; Paton, M.; Pham, Y.; Lee, J.; Jones, N.M.; Jenkin, G.; Fahey, M.C.; Miller, S.L.; et al. Human Umbilical Cord Therapy Improves Long-Term Behavioral Outcomes Following Neonatal Hypoxic Ischemic Brain Injury. Front. Physiol. 2019, 10, 283. [Google Scholar] [CrossRef]
- Min, K.; Suh, M.R.; Cho, K.H.; Park, W.; Kang, M.S.; Jang, S.J.; Kim, S.H.; Rhie, S.; Choi, J.I.; Kim, H.-J.; et al. Potentiation of cord blood cell therapy with erythropoietin for children with CP: A 2 × 2 factorial randomized placebo-controlled trial. Stem Cell Res. Ther. 2020, 11, 509. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Park, J.-S. Usage of Human Mesenchymal Stem Cells in Cell-based Therapy: Advantages and Disadvantages. Dev. Reprod. 2017, 21, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schäbitz, W.-R.; Steigleder, T.; Cooper-Kuhn, C.M.; Schwab, S.; Sommer, C.; Schneider, A.; Kuhn, H.-G. Intravenous Brain-Derived Neurotrophic Factor Enhances Poststroke Sensorimotor Recovery and Stimulates Neurogenesis. Stroke 2007, 38, 2165–2172. [Google Scholar] [CrossRef] [Green Version]
- van Velthoven, C.T.; Braccioli, L.; Willemen, H.L.; Kavelaars, A.; Heijnen, C.J. Therapeutic po-tential of genetically modi-fied mesenchymal stem cells after neonatal hypoxic-ischemic brain damage. Mol. Ther. 2014, 22, 645–654. [Google Scholar] [CrossRef] [Green Version]
- Sirén, A.-L.; Fratelli, M.; Brines, M.; Goemans, C.; Casagrande, S.; Lewczuk, P.; Keenan, S.; Gleiter, C.; Pasquali, C.; Capobianco, A.; et al. Erythropoietin prevents neuronal apoptosis after cerebral ischemia and metabolic stress. Proc. Natl. Acad. Sci. USA 2001, 98, 4044–4049. [Google Scholar] [CrossRef] [Green Version]
- Rangarajan, V.; Juul, S.E. Erythropoietin: Emerging Role of Erythropoietin in Neonatal Neuroprotection. Pediatr. Neurol. 2014, 51, 481–488. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Hodges, V.M.; Dunlop, E.; Percy, M.J.; Maxwell, A.P.; El-Tanani, M.; Lappin, T.R. Erythropoietin-Induced Activation of the JAK2/STAT5, PI3K/Akt, and Ras/ERK Pathways Promotes Malignant Cell Behavior in a Modified Breast Cancer Cell Line. Mol. Cancer Res. 2010, 8, 615–626. [Google Scholar] [CrossRef] [Green Version]
- Vairano, M.; Dello Russo, C.; Pozzoli, G.; Battaglia, A.; Scambia, G.; Tringali, G.; Aloe-Spiriti, M.A.; Preziosi, P.; Navarra, P. Erythropoietin exerts anti-apoptotic effects on rat microglial cells in vitro. Eur. J. Neurosci. 2002, 16, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.; Choi, J.; Kim, M. Combining Human Umbilical Cord Blood Cells With Erythropoietin Enhances Angiogene-sis/Neurogenesis and Behavioral Recovery After Stroke. Front. Neurol. 2019, 10, 357. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yao, L.; Yang, J.; Wang, Z.; Du, G. PI3K/Akt and HIF 1 signaling pathway in hypox-ia ischemia. Mol. Med. Rep. 2018, 18, 3547–3554. [Google Scholar]
- Zhang, H.; Gao, J.; Wang, M.; Yu, X.; Lv, X.; Deng, H.; Fan, X.; Chen, K. Effects of Scalp Electroacupuncture on the Pi3K/Akt Signalling Pathway and Apoptosis of Hippocampal Neurons in a Rat Model of Cerebral Palsy. Acupunct. Med. 2018, 36, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Rowe, D.D.; Leonardo, C.C.; Recio, J.A.; Collier, L.A.; Willing, A.E.; Pennypacker, K.R. Human Umbilical Cord Blood Cells Protect Oligodendrocytes from Brain Ischemia through Akt Signal Transduction. J. Biol. Chem. 2012, 287, 4177–4187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, K.H.; Choi, J.I.; Kim, J.-O.; Jung, J.E.; Kim, D.-W.; Kim, M. Therapeutic mechanism of cord blood mononuclear cells via the IL-8-mediated angiogenic pathway in neonatal hypoxic-ischaemic brain injury. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Chen, J.; Wang, L.; Lu, M.; Chopp, M. Treatment of stroke in rat with intracarotid administration of marrow stromal cells. Neurology 2001, 56, 1666–1672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, S.H.; Yu, J.H.; Park, E.S.; Lee, J.E.; Kim, H.O.; Park, K.I.; Kim, G.W.; Park, C.I.; Cho, S.R. Induction of striatal neu-rogenesis enhances functional recovery in an adult animal model of neonatal hypoxic-ischemic brain injury. Neuroscience 2010, 169, 259–268. [Google Scholar] [CrossRef]
- Meisel, C.; Schwab, J.M.; Prass, K.; Meisel, A.; Dirnagl, U. Central nervous system injury-induced immune deficiency syn-drome. Nat. Rev. Neurosci. 2005, 6, 775–786. [Google Scholar] [CrossRef]
- Fleiss, B.; Gressens, P. Tertiary mechanisms of brain damage: A new hope for treatment of cerebral palsy? Lancet Neurol. 2012, 11, 556–566. [Google Scholar] [CrossRef]
- Hanna, S.E.; Rosenbaum, P.L.; Pt, D.J.B.; Palisano, R.J.; Walter, S.D.; Avery, L.; Russell, D.J. Stability and decline in gross motor function among children and youth with cerebral palsy aged 2 to 21 years. Dev. Med. Child Neurol. 2009, 51, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Galieva, L.R.; Mukhamedshina, Y.O.; Arkhipova, S.S.; Rizvanov, A. Human Umbilical Cord Blood Cell Transplantation in Neuroregenerative Strategies. Front. Pharmacol. 2017, 8, 628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, C.; Fahey, M.C.; Jenkin, G.; Miller, S. Umbilical cord blood cells for treatment of cerebral palsy; timing and treatment options. Pediatr. Res. 2017, 83, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Gorio, A.; Gökmen, N.; Erbayraktar, S.; Yilmaz, O.; Madaschi, L.; Cichetti, C.; Di Giulio, A.M.; Vardar, E.; Cerami, A.; Brines, M. Recombinant human erythropoietin counteracts secondary injury and markedly enhances neurological recovery from experimental spinal cord trauma. Proc. Natl. Acad. Sci. USA 2002, 99, 9450–9455. [Google Scholar] [CrossRef] [Green Version]
- Iwai, M.; Cao, G.; Yin, W.; Stetler, R.A.; Liu, J.; Chen, J. Erythropoietin promotes neuronal re-placement through revascular-ization and neurogenesis after neonatal hypoxia/ischemia in rats. Stroke 2007, 38, 2795–2803. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.S.; Song, J.; Min, K.; Choi, Y.S.; Kim, S.M.; Cho, S.R.; Kim, M. Short-term effects of erythropoietin on neurodevelopment in infants with cerebral palsy: A pilot study. Brain Dev. 2014, 36, 764–769. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Guan, Y.M.; Huang, H.L.; Wang, Q.S. Human umbilical cord blood mesenchymal stem cell transplantation sup-presses inflammatory responses and neuronal apoptosis during early stage of focal cerebral ischemia in rabbits. Acta Pharmacol. Sin. 2014, 35, 585–591. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.W.; Li, F.; Zheng, Z.T.; Li, Y.D.; Chen, T.H.; Gao, W.W.; Chen, J.L.; Zhang, J.N. Erythropoietin regulates im-mune/inflammatory reaction and improves neurological function out-comes in traumatic brain injury. Brain Behav. 2017, 7, e00827. [Google Scholar] [CrossRef]
- Yin, W.; Signore, A.P.; Iwai, M.; Cao, G.; Gao, Y.; Johnnides, M.J.; Hickey, R.W.; Chen, J. Pre-conditioning suppresses inflammation in neonatal hypoxic ischemia via Akt activation. Stroke 2007, 38, 1017–1024. [Google Scholar] [CrossRef] [Green Version]
- Gibson, M.E.; Han, B.H.; Choi, J.; Knudson, C.M.; Korsmeyer, S.J.; Parsadanian, M.; Holtzman, D.M. BAX Contributes to Apoptotic-Like Death Following Neonatal Hypoxia-Ischemia: Evidence for Distinct Apoptosis Pathways. Mol. Med. 2001, 7, 644–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jian, Z.; Liu, R.; Zhu, X.; Smerin, D.; Zhong, Y.; Gu, L.; Fang, W.; Xiong, X. The Involvement and Therapy Target of Immune Cells After Ischemic Stroke. Front. Immunol. 2019, 10, 2167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, X.; Hua, F.; Sandhu, H.K.; Chao, D.; Balboni, G.; Salvadori, S.; He, X.; Xia, Y. Effect of δ-opioid receptor activation on BDNF-TrkB vs. TNF-α in the mouse cortex exposed to prolonged hypoxia. Int. J. Mol. Sci. 2013, 14, 15959–15976. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Concepcion, K.; Meng, X.; Zhang, L. Brain-immune interactions in perinatal hypoxic-ischemic brain injury. Prog. Neurobiol. 2017, 159, 50–68. [Google Scholar] [CrossRef] [PubMed]
- van der Kooij, M.A.; Groenendaal, F.; Kavelaars, A.; Heijnen, C.J.; van Bel, F. Neuroprotective properties and mechanisms of erythropoietin in in vitro and in vivo experimental models for hypoxia/ischemia. Brain Res. Rev. 2008, 59, 22–33. [Google Scholar] [CrossRef]
- Li, L.; Qu, Y.; Mao, M.; Xiong, Y.; Mu, D. The involvement of phosphoinositid 3-kinase/Akt pathway in the activation of hypoxia-inducible factor-1α in the developing rat brain after hypoxia–ischemia. Brain Res. 2008, 1197, 152–158. [Google Scholar] [CrossRef]
- Osuka, K.; Watanabe, Y.; Usuda, N.; Nakazawa, A.; Tokuda, M.; Yoshida, J. Modification of Endothelial NO Synthase Through Protein Phosphorylation After Forebrain Cerebral Ischemia/Reperfusion. Stroke 2004, 35, 2582–2586. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.-Z.; Ren, Y.; Jiang, Y.; Shen, S.-Y.; Ding, J.; Hua, F. GluR1 protects hypoxic ischemic brain damage via activating Akt signaling pathway in neonatal rats. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 8857–8865. [Google Scholar]
- Min, K.; Song, J.; Lee, J.H.; Kang, M.S.; Jang, S.J.; Kim, S.H.; Kim, M. Allogenic umbilical cord blood therapy combined with erythropoietin for patients with severe traumatic brain injury: Three case reports. Restor. Neurol. Neurosci. 2013, 31, 397–410. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tests | Points |
|---|---|
| Raising rat by tail (normal = 0; maximum = 3) | |
| Flexion of forelimb | 1 |
| Flexion of hindlimb | 1 |
| Head moved > 10 to vertical axis within 30 s | 1 |
| Walking rat on floor (normal = 0; maximum = 3) | |
| Normal walk | 0 |
| Inability to walk straight | 1 |
| Circling toward the paretic side | 2 |
| Falls down to paretic side | 3 |
| Beam balance test (normal = 0; maximum = 6) | |
| Balances with steady posture (>60 s) | 0 |
| Grasps side of the beam | 1 |
| Hugs beam and 1 limb falls down from beam | 2 |
| Hugs beam and 2 limbs falls down from beam, or spins on beam (>60 s) | 3 |
| Attempts to balance on beam but falls off (>40 s) | 4 |
| Attempts to balance on beam but falls off (>20 s) | 5 |
| Falls off, no attempt to balance or hang on the beam (<20 s) | 6 |
| Sensory tests (normal = 0; maximum = 2) | |
| Placing test (visual and tactile test, try 5 times and have not moved more than 3 times) | 1 |
| Proprioceptive test (deep sensation, pushing the paw against the table edge to stimulate limb muscles, try 5 times and have not moved more than 3 times) | 1 |
| Maximum Points 14 | 14 |
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| TNF-α | TACTGAACTTCGGGGTGATCGGTCC | CAGCCTTGTCCCTTGAAGGAACC |
| IL-1 β | AAGGAGAACCAAGCAACGACAAAA | TGGGGAACTCTGCAGACTCAAACT |
| TGF-β | TGACGTCACTGGAGTTGTACGG | GGTTCATGTCATGGATGGTGC |
| IL-6 | TACCACTTCACAAGTCGGAGGC | CTGCAAGTGCATCATCGTTGTTC |
| IL-2 | GCACCCACTTCAAGCTCCA | AAATTTGAAGGTGAGCATCCTG |
| IL-10 | ATGCTCCTAGAGCTGCGGACT | CCTGCATTAAGGAGTCGGTTAG |
| TrkB | CCGGCTTAAAGTTTGTGGCTTAC | GGATCAGGTCAGACAAGTCAAG |
| Nkx2.1 | GATGGTACGGCGCCAACCCAG | ACTCATATTCATGCCGCTCGC |
| β-actin | AGAGGGAAATCGTGCGTGAC | CAATAGTGATGACCTGGCCGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.I.; Choi, J.-W.; Shim, K.-H.; Choung, J.S.; Kim, H.-J.; Sim, H.R.; Suh, M.R.; Jung, J.E.; Kim, M. Synergistic Effect in Neurological Recovery via Anti-Apoptotic Akt Signaling in Umbilical Cord Blood and Erythropoietin Combination Therapy for Neonatal Hypoxic-Ischemic Brain Injury. Int. J. Mol. Sci. 2021, 22, 11995. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111995
Choi JI, Choi J-W, Shim K-H, Choung JS, Kim H-J, Sim HR, Suh MR, Jung JE, Kim M. Synergistic Effect in Neurological Recovery via Anti-Apoptotic Akt Signaling in Umbilical Cord Blood and Erythropoietin Combination Therapy for Neonatal Hypoxic-Ischemic Brain Injury. International Journal of Molecular Sciences. 2021; 22(21):11995. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111995
Chicago/Turabian StyleChoi, Jee In, Joo-Wan Choi, Kyu-Ho Shim, Jin Seung Choung, Hyun-Jin Kim, Hye Ryeong Sim, Mi Ri Suh, Joo Eun Jung, and MinYoung Kim. 2021. "Synergistic Effect in Neurological Recovery via Anti-Apoptotic Akt Signaling in Umbilical Cord Blood and Erythropoietin Combination Therapy for Neonatal Hypoxic-Ischemic Brain Injury" International Journal of Molecular Sciences 22, no. 21: 11995. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111995