
Molecular and Functional Analysis of U-box E3 Ubiquitin Ligase Gene Family in Rice (Oryza sativa)


1
Department of Crop Science, College of Agriculture and Life & Environment Sciences, Chungbuk National University, Cheongju 28644, Korea
2
Division of Horticultural Biotechnology, Hankyong National University, Anseong 17579, Korea
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(21), 12088; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222112088
Submission received: 25 September 2021
/
Revised: 21 October 2021
/
Accepted: 5 November 2021
/
Published: 8 November 2021
(This article belongs to the Special Issue Crop Genome Editing)
Abstract
:Proteins encoded by U-box type ubiquitin ligase (PUB) genes in rice are known to play an important role in plant responses to abiotic and biotic stresses. Functional analysis has revealed a detailed molecular mechanism involving PUB proteins in relation to abiotic and biotic stresses. In this study, characteristics of 77 OsPUB genes in rice were identified. Systematic and comprehensive analyses of the OsPUB gene family were then performed, including analysis of conserved domains, phylogenetic relationships, gene structure, chromosome location, cis-acting elements, and expression patterns. Through transcriptome analysis, we confirmed that 16 OsPUB genes show similar expression patterns in drought stress and blast infection response pathways. Numerous cis-acting elements were found in promoter sequences of 16 OsPUB genes, indicating that the OsPUB genes might be involved in complex regulatory networks to control hormones, stress responses, and cellular development. We performed qRT-PCR on 16 OsPUB genes under drought stress and blast infection to further identify the reliability of transcriptome and cis-element analysis data. It was confirmed that the expression pattern was similar to RNA-sequencing analysis results. The transcription of OsPUB under various stress conditions indicates that the PUB gene might have various functions in the responses of rice to abiotic and biotic stresses. Taken together, these results indicate that the genome-wide analysis of OsPUB genes can provide a solid basis for the functional analysis of U-box E3 ubiquitin ligase genes. The molecular information of the U-box E3 ubiquitin ligase gene family in rice, including gene expression patterns and cis-acting regulatory elements, could be useful for future crop breeding programs by genome editing.
Keywords:
ubiquitination; E3 ligase; OsPUB; genome-wide; transcriptome; gene expression; cis-element; rice1. Introduction
Protein degradation regulated by the ubiquitin-proteasome system plays an important role in plants. This system is used to selectively degrade proteins through a process of poly-ubiquitination in eukaryotes [1,2]. In plants, adaptation in response to stress, signal transduction, DNA repair, differentiation, and immune response can be achieved through ubiquitination with the resulting degradation of components specific to these mechanisms [3,4]. Ubiquitination of a target protein is carried out involving a three-step (E1 → E2 → E3) conjugation cascade that detects specific ubiquitin signals. First, ubiquitins (Ub) are activated by a Ub-activating enzyme (E1). Ub is then transferred to a Ub-conjugating enzyme (E2). Finally, Ub is attached to the substrate protein by a specific Ub ligase enzyme (E3). Ub-conjugated proteins then become visible to the 26S proteasome, which is responsible for the protein degradation process [5,6].
E3 ubiquitin ligases are key factors in determining substrate specificity. They are classified into three main subfamilies: HECT (Homologous to E6-associated protein C-Terminus), RING (Real Interesting New Gene), and U-box according to the subunit construction and mechanism [7]. HECT proteins are single polypeptides that can form ubiquitin and thioester intermediates before target ubiquitination [8]. RING and U-box proteins are structurally related single polypeptides that use zinc chelation and hydrogen bonds and salt bridges to transfer ubiquitin from the E2-ubiquitin intermediate to the substrate, respectively [9,10].
U-box proteins are formed by a signature U-box domain containing 70 amino acids and a modified RING finger domain, which lacks characteristic zinc-chelating cysteine and histidine residues of the latter [11,12,13]. U-box proteins in plants (PUB) are classified into 13 groups based on the presence and structure of their domains. Most PUB genes contain armadillo repeats (ARM) that can mediate substrate interactions. Other protein domains include the U-box N-terminal domain (UND), Ser/Thr kinase domain (kinase), WD40 repeat (WD40), MIF4G domain (MIF4G), tetratricopeptide repeat (TPR), and peptidyl-prolyl isomerase (PPIase) [14,15,16]. Deletions of the PUB gene domain and point mutations of conserved amino acids within this domain can affect E3 ligase activity [17].
Plants use the ubiquitin/proteasome system (UPS) to promote changes in cellular protein content needed for continuous growth, development, and survival in a constantly changing environment. The ubiquitination of regulatory proteins such as transcription factors might be promoted or inhibited in response to environmental stimuli, leading to increased degradation or stabilization of these proteins, changes in gene expression patterns, and necessary cellular responses [18,19]. Targeted control of protein abundance can induce plants to respond appropriately by regulating signaling events, thereby reducing damage caused by external stress factors. A plant’s ability to effectively endure environmental stress is transformed when the UPS is interrupted. Mutations in regulatory particles of the 26S proteasome can reduce abiotic stress tolerance [20,21].
In Arabidopsis, RPN10 mutants are weakly resistant to high-salinity conditions, heat, and UV treatment [22]. Similarly, RPN1a mutants are sensitive to salinity and heat stress [23]. RPN12a and RPT2a mutants are also weakly resistant to heat stress [24]. Mutants of RPT2a or RPT5a render plants weakly resistant to the zinc deficiency condition [25]. The results of these mutation analyses indicate the importance of the 26S proteasome’s function in plant responses to adverse growth conditions. Studies on the importance of UPS functions in abiotic stress tolerance results from the analysis of ubiquitin and ubiquitin enzymes assay, and the expression of ubiquitin genes responds to stress [26,27]. Transgenic plants overexpressing mono- or poly-ubiquitin genes show strong tolerance to several abiotic stresses, including cold, salinity, and drought [28,29]. In addition, changes in ubiquitin protein expression can alter the response of plants to abiotic stresses. Expression levels of many E2 proteins are differentially regulated by abiotic stress. Expression levels of 14 of 39 E2 protein genes in rice have been shown to be upregulated or downregulated in response to drought and salt stresses [30]. The overexpression of GmUBC2 from soybean, VrUBC1 from mung bean, and AhUBC2 from dehydrated peanut plants can increase the tolerance of these plants to drought stress [31,32,33]. In addition, the increased expression of tobacco-derived NtUBC1 can promote cadmium resistance [34].
In rice, 77 U-box E3 ligase genes are known, of which only six genes have been reported functionally. That is, many rice U-box E3 genes have not been studied yet. Phenotypic interference due to functional duplication might be one of the reasons. Therefore, the objective of this study was to obtain information on the environmental response characteristics of individual PUB genes related to abiotic and biotic stresses tolerance in rice for breeding programs. Systematic characterization of the U-box E3 ubiquitin ligase gene family in rice was performed, including expression profiles, and cis-acting regulatory element structures of PUB genes related to stresses were analyzed. The results of this study will facilitate future crop breeding programs by genome editing of OsPUB genes.
2. Results
2.1. Characteristics and Chromosomal Distribution of U-box-Type E3 Ubiquitin Ligase Gene Family in Rice
According to recent reports on the U-box-type E3 ubiquitin ligase gene family in rice, 77 estimated PUB proteins were identified through a whole genome analysis algorithm and divided into eight classes according to domain configuration [35]. We adopted genomic and protein sequences of OsPUB genes from Gramene, RAP-DA, and NCBI. Identified rice protein sequences were then analyzed using ProtParam and the compute pI/Mw tool from ExPasy to further confirm physical parameters. Detailed characteristics of the OsPUB gene family are listed in Supplementary Table S1, including gene name, number of nucleotides, number of amino acids, molecular weight, theoretical isoelectric point, instability index, aliphatic index, grand average of hydropathicity (GRAVY), and chromosomal positions. These 77 OsPUB proteins have diverse molecular sizes in gene base pairs and amino acid lengths, ranging from 432 bp (OsPUB30) to 7675 bp (OsPUB27) with an average of 3598 bp in gene base pairs and from 212 a.a. (OsPUB68) to 1392 a.a. (OsPUB71) with an average of 605.13 amino acids. OsPUB68 had the smallest molecular weight (24.15 kDa) and OsPUB71 had the largest molecular weight (148.25 kDa). Their average molecular weight was 36 kDa. Theoretical isoelectric points (pIs) of these OsPUB proteins varied from 5.07 (OsPUB10) to 9.04 (OsPUB69) with an average of 6.82. Their aliphatic indices ranged from 69.29 (OsPUB26) to 109.68 (OsPUB31) with an average of 92.31. Among these 77 OsPUB proteins, 54 were hydrophobic proteins with a value of GRAVY less than 0. The remaining 23 genes were identified as hydrophilic proteins with a value of 0 or more.
Chromosomal locations of OsPUB genes were identified by extracting chromosomal data and mapping using the MapGene2Chromosome V2 program. The 77 OsPUB genes were distributed in 10 of 12 chromosomes of Oryza sativa, showing that each gene was unevenly distributed nonrandomly on the chromosome (Figure 1). Fifteen (the largest number) OsPUB genes were mapped on chromosome 2, while no OsPUB gene was mapped on chromosome 7 or 11. Fifteen out of 28 Class 2 OsPUB genes were distributed mainly on Chr 2, Chr 6, and Chr 8. The rest of the OsPUB genes were located on six chromosomes. Ten out of 16 Class 3 genes were mapped mainly on Chr 2 and Chr 4. Nine OsPUB genes in Class 4 were positioned on Chr 2 and Chr 10. The remaining seven OsPUB genes in Class 4 were distributed on Chr 3, 6, and 9. Genes belonging to Class 1, Class 5, Class 6, Class 7, and Class 8 were sporadically distributed on eight chromosomes.
2.2. Phylogenic Analysis and Gene Structure Analysis of OsPUB Genes
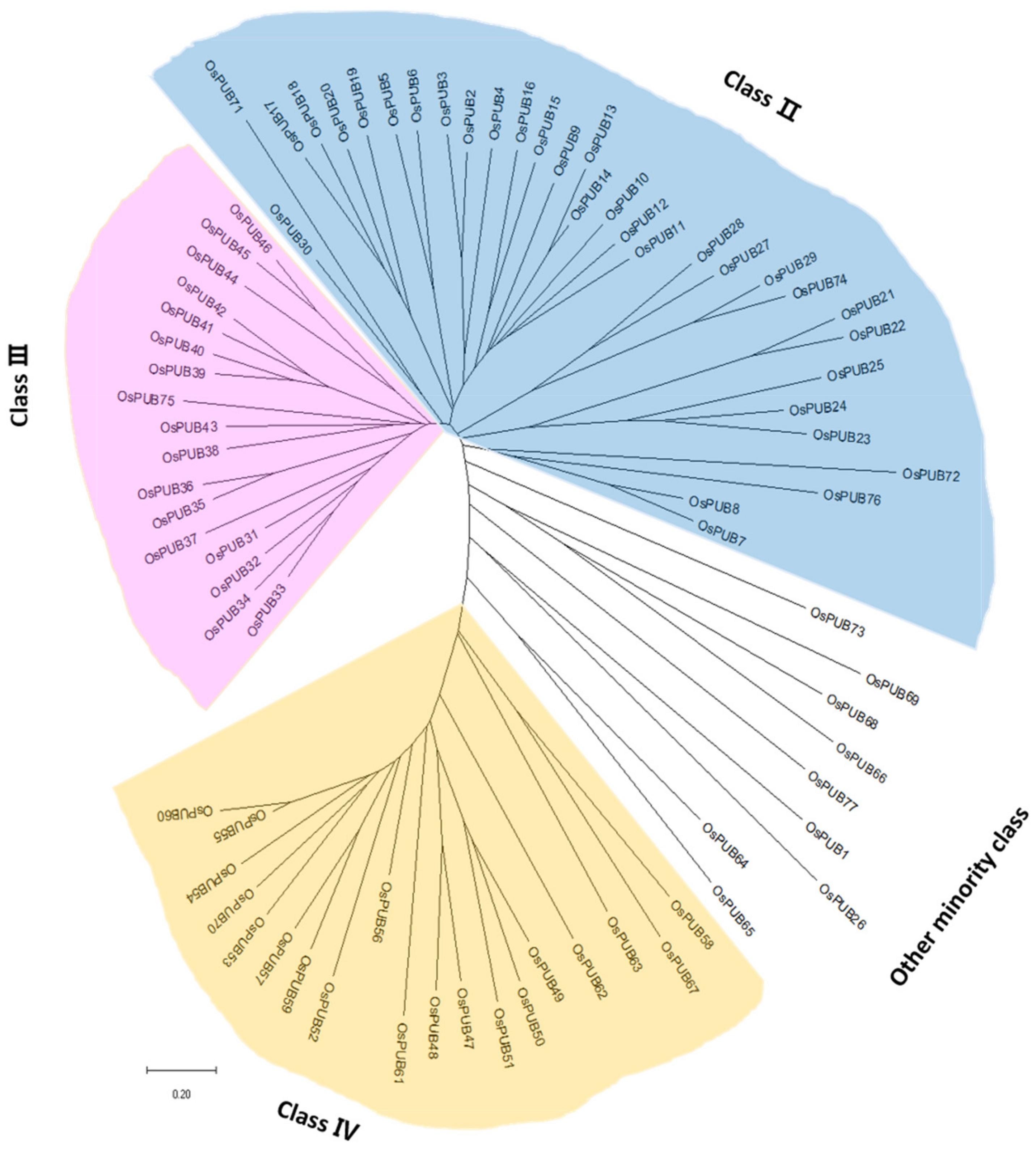
To better identify the phylogenetic relationship between PUB genes in rice and to evaluate the evolutionary history of the protein family, we performed multiple sequence alignment using ClustalW. Based on full-length sequences, a phylogenetic tree was constructed with MEGA 7.0 using the Neighbor-Joining method (Figure 2). As shown in the phylogenetic tree, all OsPUB genes were classified into three major subfamilies and one minor subfamily according to their structures or functions, consistent with previous studies on rice and Arabidopsis [35,36].
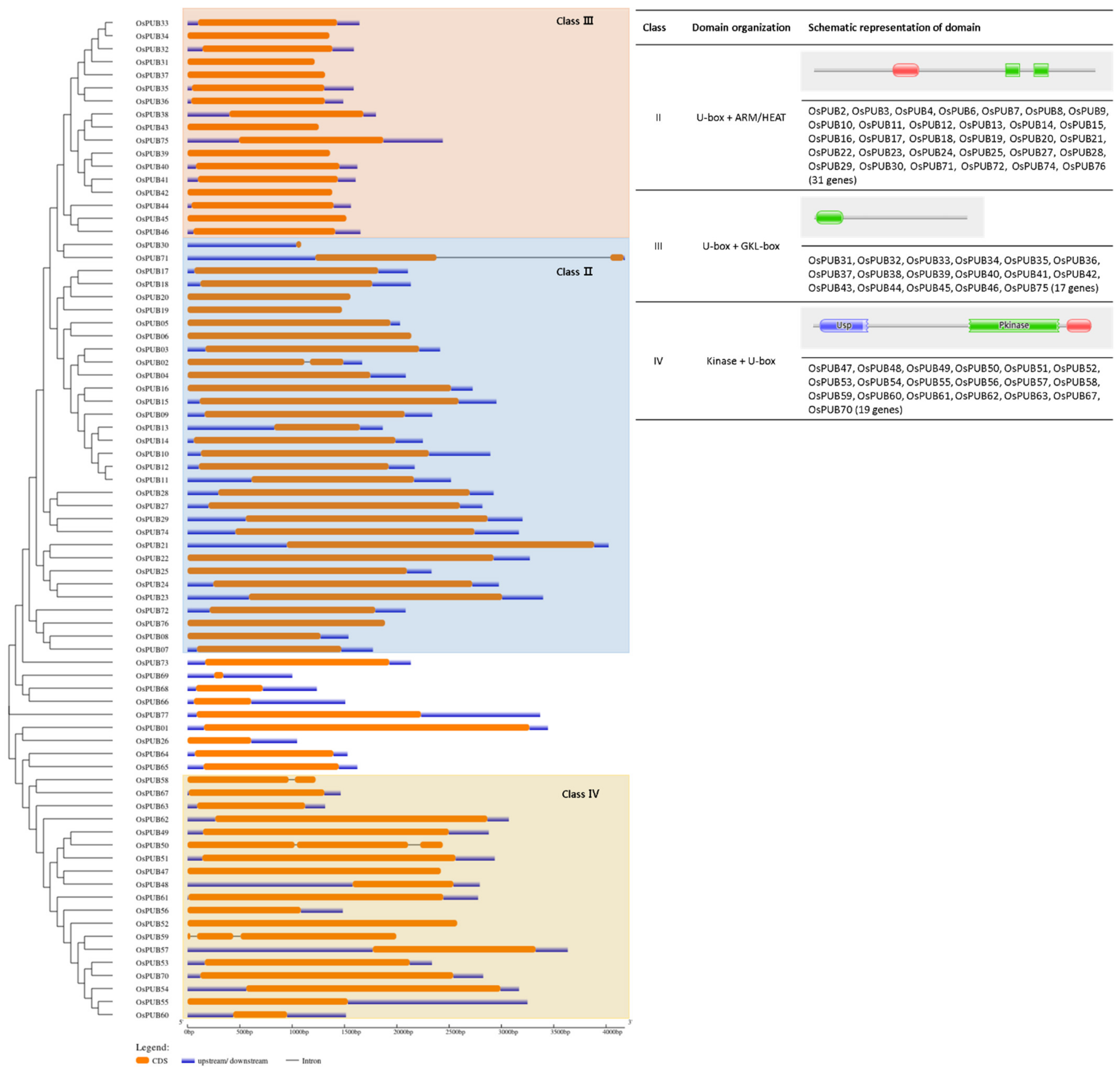
To understand exon/intron structures of individual OsPUB genes, their corresponding genomic DNA sequences were aligned using GSDS (Gene Structure Display Server). An unrooted tree categorized OsPUB genes into three major groups and one minor group with well-supporting bootstrap values, indicating that the classification was reliable (Figure 2 and Figure 3). In the Class 2 group, schematic structures revealed that most paralogs shared a similar exon/intron structure with one exon, while OsPUB2 and OsPUB71 had two exons. Class 2, the largest class, consisted of 32 genes with armadillo (ARM)/HEAT repeats. The ARM repeat is a double repeat sequence motif approximately 40 amino acids long. It was first identified in Drosophila melanogaster segment polarity gene armadillo [37]. It has been shown to be involved in protein–protein interactions [38]. The HEAT repeat derived its name from four different eukaryotic proteins, huntingtin, elongation factor 3, PR65/A subunit of protein phosphatase A, and TOR (target of rapamycin) [39]. Seventeen OsPUB genes in Class 3 shared a similar gene structure. Class 3, the second largest class of U-box proteins in rice, has a conserved domain containing ~100 amino acid residues located close to the C-terminus of the protein according to sequence alignment. In addition to a high proportion of leucine residues, a high percentage of homology and several highly conserved residues were detected [40]. This domain was named the GKL domain because it was rich in conserved glycine (G), lysine (K)/arginine (R), and leucine (L). Sequence alignments of Arabidopsis proteins of the same class were also found to exhibit very similar patterns. Arabidopsis thaliana U-box proteins and Nicotiana benthamiana proteins of this class were shown to be essential for plant defense and disease resistance [41]. Class 4 consisting of 19 U-box proteins has a kinase domain in the N-terminal region of the protein. Considering the involvement of phosphorylation in most cell processes, we speculate that members of this class of OsPUB genes might play a broad role in cells, if not all.
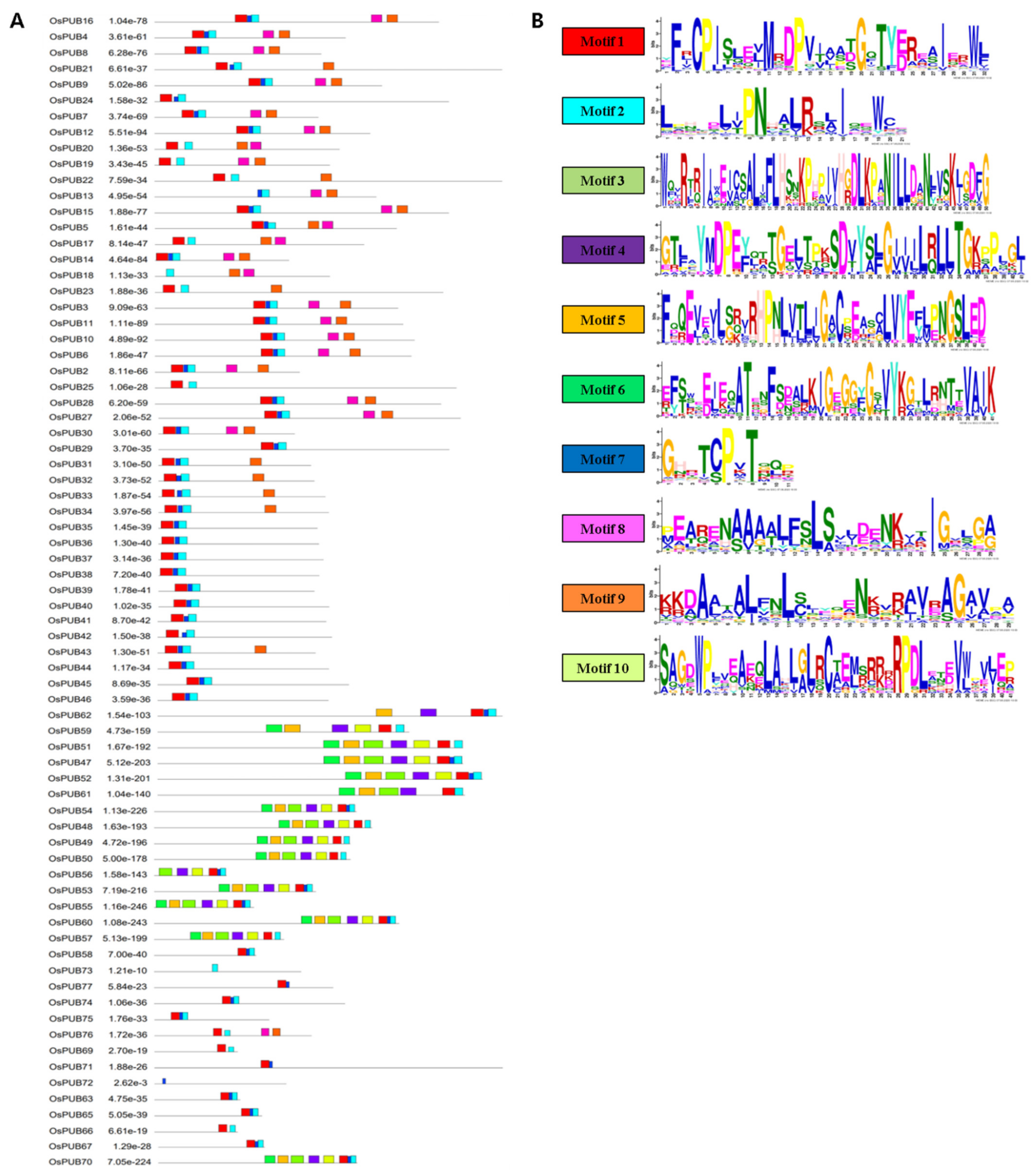
Conserved protein motifs of OsPUB genes were identified using MEME, a publicly available online tool. We identified 10 motifs with an amino acid sequence length that varied from 6 to 50. The motif sequence logo is presented in Figure 4. All OsPUB proteins except OsPUB1, OsPUB26, OsPUB64, and OsPUB69 contained motifs. The type and number of motifs were almost distinct for each family. Most member proteins in each family shared a common motif. A highly significant motif, motif 1 (YF[R]CPIS[L]E[V]MRDPVI[AA]TGQTYERE[A]I[E]RWL), showed regular expression. It is present in all protein sequences except OsPUB13, OsPUB18, and OsPUB73. Members of class 4 shared eight of 10 motifs, excluding motifs 8 and 9. Members of class 2 shared only five motifs. Members of class 3 shared only motifs 1, 2, and 7. OsPUB72 and OsPUB73 genes only shared a single motif.
2.3. Gene Expression Patterns of Rice U-box Gene Family Members under Abiotic and Biotic Stresses
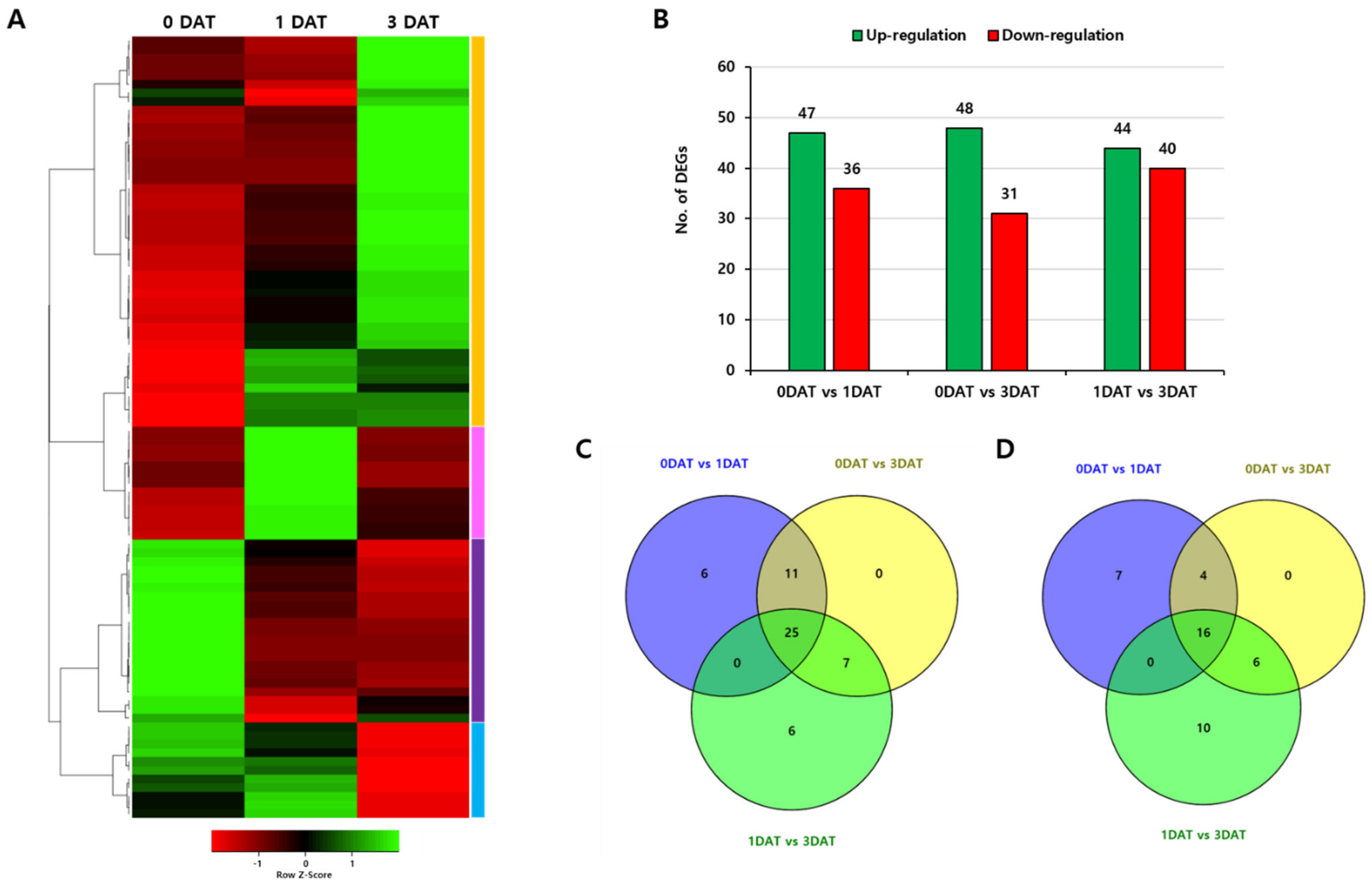
To investigate expression changes of OsPUB genes under abiotic stress after treatment, we collected leaves at 0, 1, and 3 days after treatment (DAT) with drought stress and performed RNA-sequencing analysis. High-throughput sequencing produced a total of 167.3 million short reads, of which 98.82% (165.3 million) passed the quality control thresholds. To calculate the expression level and profile, clean reads were mapped onto the Oryza sativa reference genome using Trimmomatic v0.38. Thereafter, the read count of the transcript expression level was calculated using a StringTie v1.3.4d program. A comparative analysis was performed between samples (0 DAT, 1 DAT, and 3 DAT) based on the read count of StringTie calculated at the transcript level. In DESeq (V2.0), read count is normalized through the size factor and dispersion, and DEG is performed through the calculated log2 fold change value and FDR after normalization. DEGs were reported in log2FC with a corresponding p-value for each gene. Three samples were divided into three comparison groups. DEG analysis was then performed. In all three samples, the number of transcripts having expression values was 36,726. Values were selected according to statistical significance and differences in expression levels. As a result of a Heatmap for 77 OsPUB genes, it was confirmed that each gene showed differential expression. There were four expression pattern groups. For experimental samples comparing 14-day-old seedlings not treated with up- or downregulation to controls, 25 genes were found to be commonly upregulated and 16 were all downregulated under the given conditions above (Figure 5).
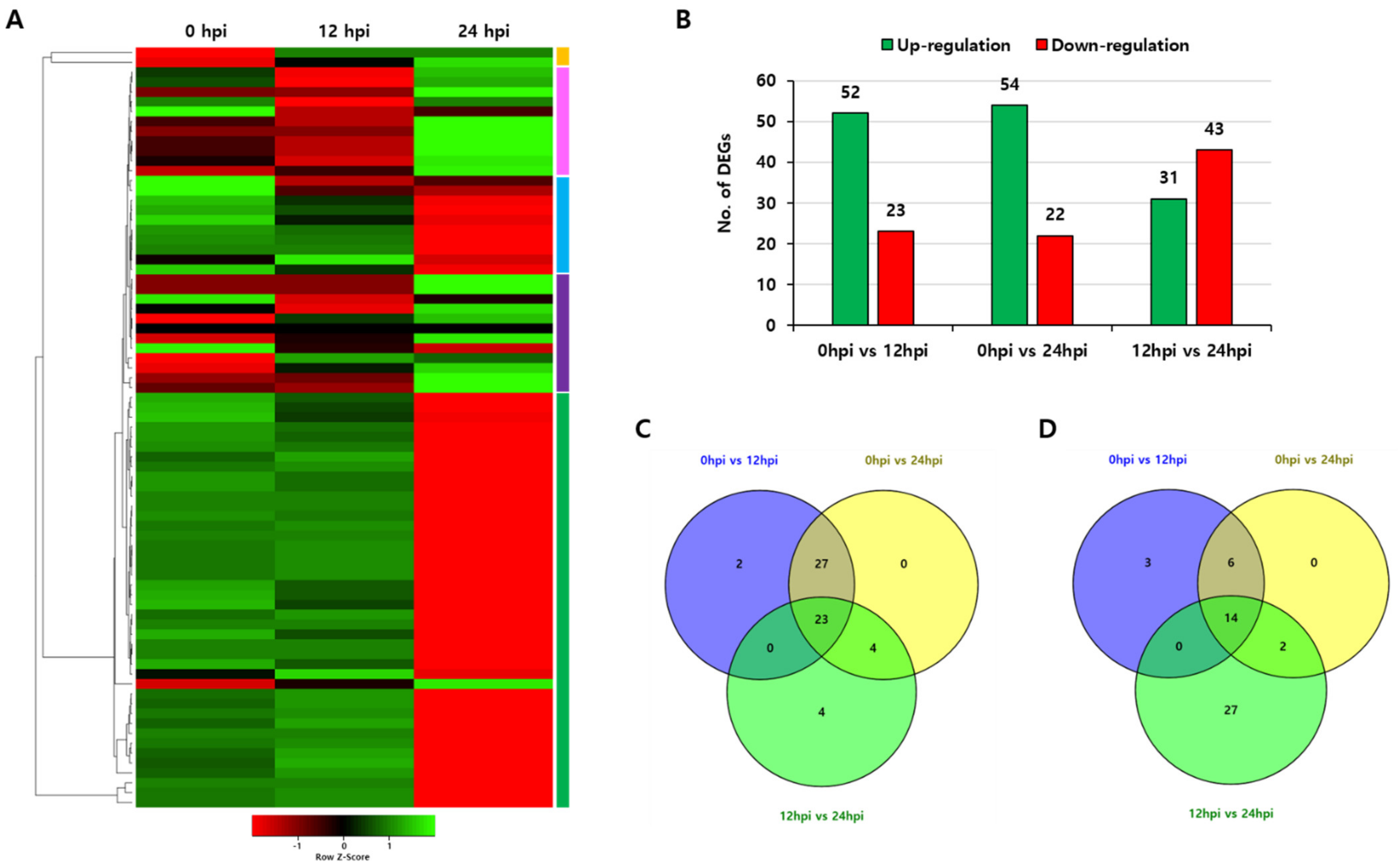
To investigate the putative role of the OsPUB gene in response to treatment with biotic stress, the specific expression pattern in rice during blast infection caused by fungus Magnaporthe oryzae was analyzed. Transcript accumulation patterns were investigated for each period (0 hpi, 12 hpi, and 24 hpi) of RNA-sequencing data of Oryza sativa L. ssp. indica after rice blast infection in GEO DataSets. Expression patterns at 0 hpi, 12 hpi, and 24 hpi were analyzed for 77 OsPUB genes out of 24,933 DEGs that were co-expressed at these three time points among all DEGs. Heatmap analysis for expression levels of all these genes showed that their expression profiles were quite diverse in response to rice blast infection. Five expression pattern groups were found. When the experimental sample was compared to the control, 23 and 14 genes were found to be commonly upregulated and downregulated, respectively, under the above conditions (Figure 6).
To further explore the role of OsPUB in regulating responses of rice to abiotic and biotic stresses, genes showing common expression patterns were identified. Results are shown in Table 1. Seven genes were identified to be upregulated and nine genes were found to be downregulated in common under both drought stress and rice blast infection. To identify protein structures of OsPUB genes showing common expression patterns, protein sequences were searched against the Simple Modular Architecture Research Tool (SMART) database to reveal distinct structures of these genes. Seven OsPUB genes commonly upregulated were found to have only a U-box domain in their protein structures. It was confirmed that the other 16 genes had a tetratrico peptide repeat region (TPR) and a protein kinase region in common. TPR is a structural motif present in a wide range of proteins. It mediates protein–protein interactions and the assembly of multi-protein complexes [42,43]. Protein kinases play a role in a multitude of cellular processes, including division, proliferation, apoptosis, and differentiation. Phosphorylation usually results in a functional change in the target protein by changing enzyme activity, cellular location, or association with other proteins [44,45].
2.4. Analysis of Cis-Acting Regulatory Elements in the Promoter of U-box E3 Ubiquitin Ligase Gene Family in Rice
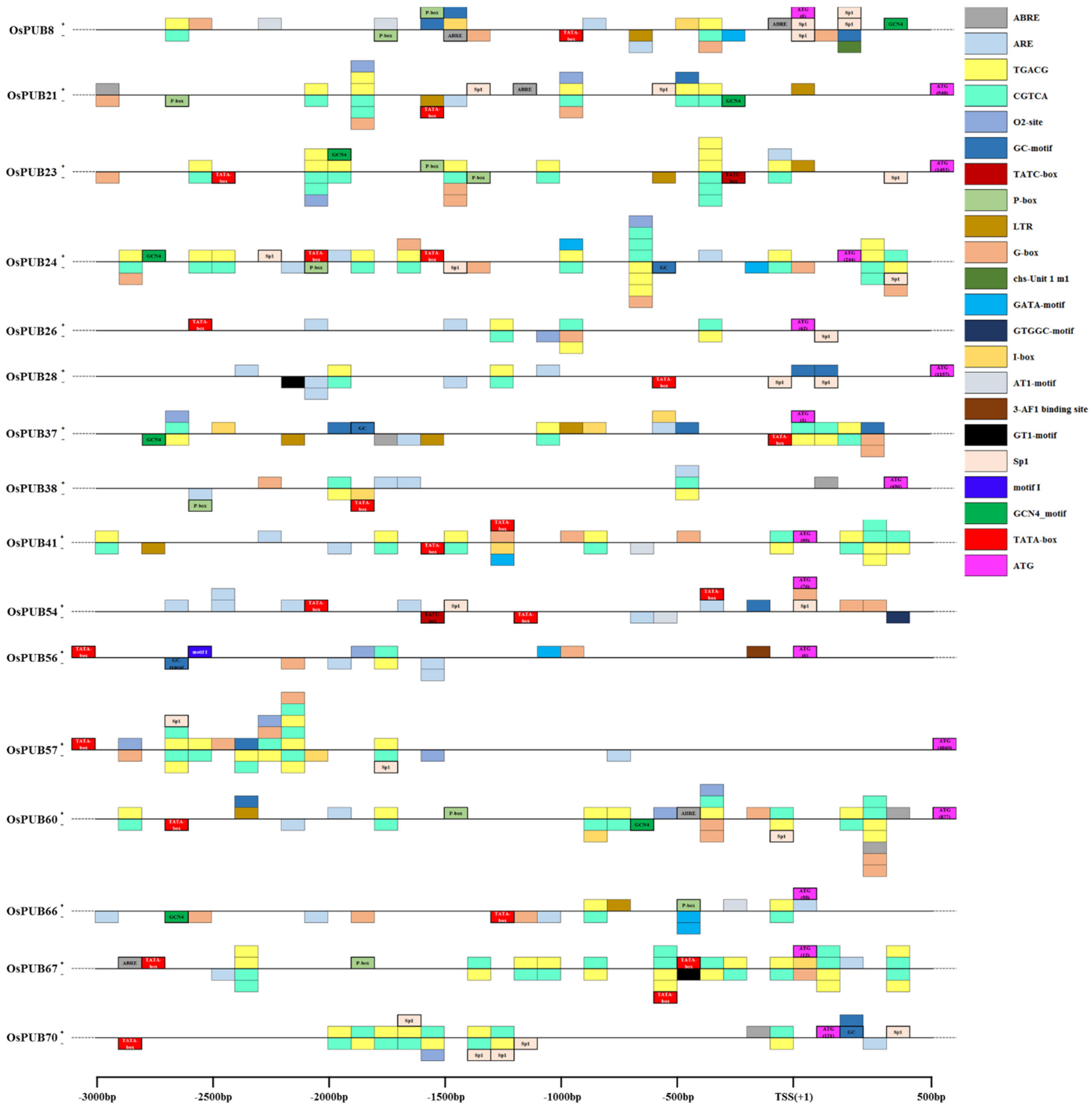
To further explain the regulatory mechanism of the OsPUB gene in response to abiotic or biotic stress, we searched 3000 bp regions upstream of 16 OsPUB genes in the PlantCARE database as the expression level of genes was regulated by cis-regulating elements present in the promoter domain (Figure 7). Eighteen types of cis-acting regulatory elements were detected in five crops (O. sativa, Z. mays, H. vulgare, T. aestivum, and S. tuberosum). Lengths of these cis-elements varied from 5 bp to 14 bp. Cis-elements are grouped into different functional categories, as shown in Table 2. Five hormone-related elements (TATC-box, ABRE, CGTCA-motif, TGACG-motif, and P-box), seven stress-related elements (LTR, G-box, GT1-motif, Sp1, GATA-motif, I-box, and AT1-motif), and four cellular development-related elements (GCN4-motif, O2-box, GC-motif, and ARE) were classified into promoters of OsPUB genes. All 16 OsPUB genes contained 5 to 14 cis-elements related to stress or hormonal response or cellular development. ABRE (abscisic acid response) was detected in seven rice OsPUB genes. CGTCA-motif (MeJA-responsiveness) and TGACG-motif (MeJA-responsiveness) were detected in all 16 rice OsPUB genes. Different cis-acting regulatory elements related to different stress responses such as light and low temperature were observed in each of the five crops. The G-box element is known to be involved in light, ABA, MeJA, ethylene induction, and seed-specific expression [46]. It was found to be present in 14 of the 16 PUB genes except for OsPUB28 and OsPUB70. Motifs involved in cellular development are relatively fewer in number compared to those involved in other responses. GCN4-motif is involved in endosperm expression. O2-box present in zein metabolism and circadian motifs plays a role in circadian control.
2.5. Gene Expression Levels of OsPUB Genes under Abiotic and Biotic Stresses
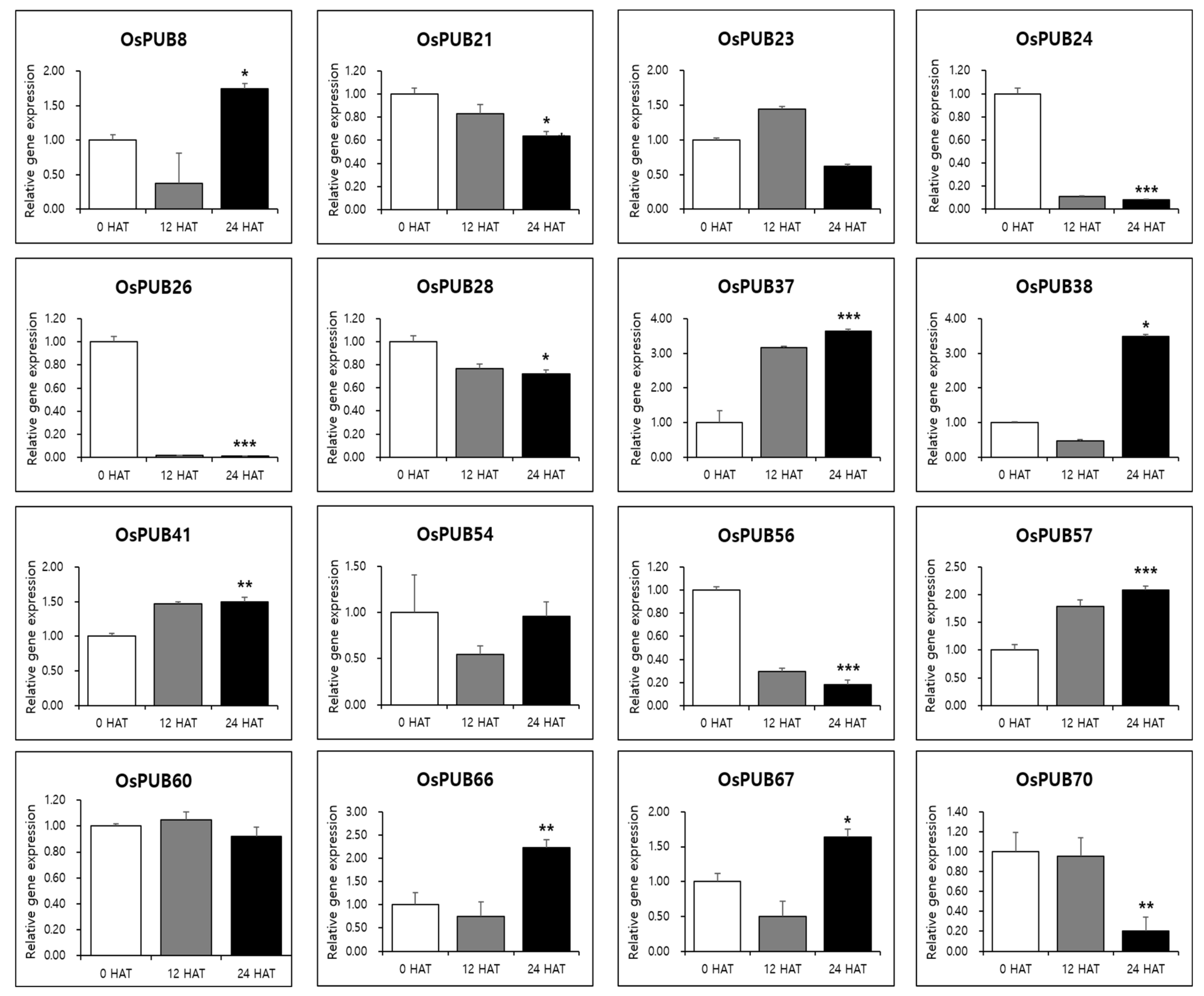
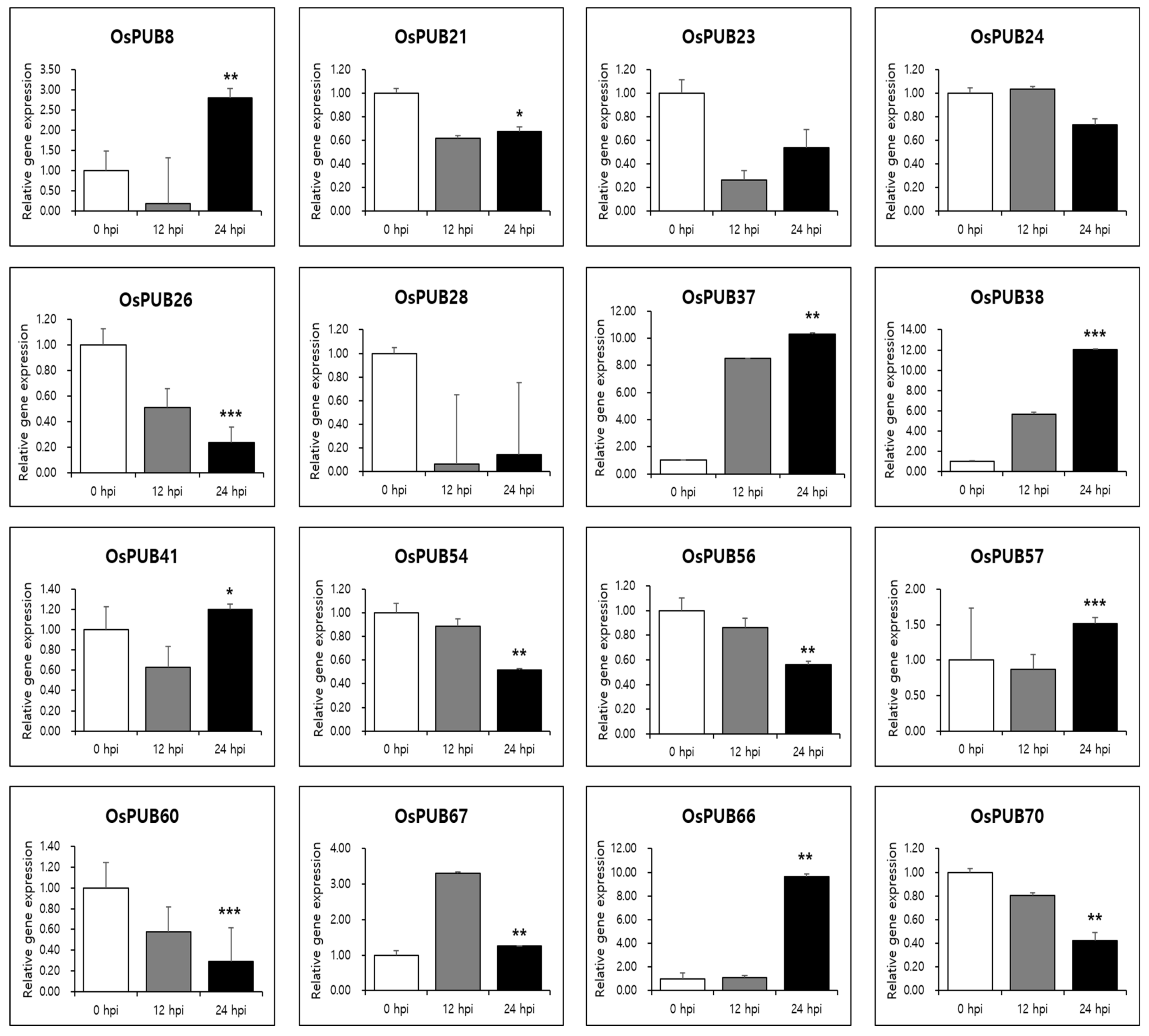
To further determine the responses of OsPUB genes to abiotic and biotic stresses, expression patterns of 77 OsPUB genes to drought stress and rice blast infection were examined using RNA-sequencing and in silico expression pattern analysis. Results revealed that OsPUB genes responded to both abiotic and biotic stresses, suggesting the authenticity of these genes and their potential role in stress response. In addition, quantitative real-time PCR was performed on 16 OsPUB genes, showing a common expression pattern to further confirm responses to abiotic and biotic stresses. Three biological replications were performed in all reactions. Results revealed that these genes were differentially expressed in leaves under normal conditions (0 HAT or no infection) (Figure 8) and under stress conditions (12 HAT, 24 HAT or 12 hpi, 24 hpi) (Figure 9). Interestingly, these 16 selected OsPUB genes showed a moderating expression pattern similar to RNA-sequencing or in silico expression analysis except for OsPUB24 and OsPUB57 under abiotic and biotic stresses. OsPUB8, OsPUB37, OsPUB38, OsPUB41, OsPUB57, OsPUB66, and OsPUB67 showed low expression levels in rice leaves under normal conditions [35,36,47]. However, they showed upregulated expression levels during drought stress. Among them, OsPUB38 and OsPUB57 showed significantly upregulated expression levels (>2-fold). In addition, nine genes (OsPUB21, OsPUB23, OsPUB24, OsPUB26, OsPUB28, OsPUB54, OsPUB56, OsPUB60, and OsPUB70) were significantly downregulated during stress treatment. Among these genes, OsPUB21 was confirmed to exhibit statistical significance between drought stress and rice blast infection expression analysis. The N. benthamiana CMPG1 gene homologous to the Arabidopsis PUB21 gene triggers cell death by the tomato receptor-like protein Cf-9 and the fungal elicitor Avr9 [41]. OsPUB57 revealed a higher expression in the resistant plants carrying the Pi9-resistant gene [48]. These results suggest that these selected genes of the Oryza sativa E3 ubiquitin ligase gene family are responsive to abiotic and biotic stresses, indicating potential roles of these genes in stress response.
3. Discussion
PUB genes in rice are known to play an important role in plant responses to abiotic and biotic stresses. Functional analysis revealed a detailed molecular mechanism involving PUB proteins in response to abiotic and biotic stresses [4,49,50]. To date, characteristics and functions of the PUB gene family have been identified in several plant species such as Arabidopsis [51], Soybean [52], Chinese cabbage [53], and Barley [54]. In this study, we performed a genome-wide analysis of PUB genes in rice by investigating their chromosome locations, phylogenetic relationships, gene structures, cis-acting elements, and gene expression profiles under various stress treatments. The classification of OsPUB proteins differs from those of other gene families based on additional domains besides the U-box, not based on genetic homology. Thus, in addition to the U-box domain, 77 OsPUB proteins can be grouped into three major groups and one minor group based on various other domains such as ARM/HEAT, GKL, TPR, and kinase. Mutations occurring at U-box motif positions often interfere with the structure and function of these enzymes [55,56]. Major PUB proteins whose biological functions have been identified belong to the PUB/ARM group [4]. The ARM repeat is a tandem repeat motif of about 40 amino acids in length that can bind to S-locus receptor kinase (SRK). It is an essential component of the signaling pathway downstream of SRK [38]. For example, PHOR1, a Solanum tuberculosum U-box protein with ARM repeats, appears to play a role in the signaling of gibberellin [57]. NtCMPG1, a Nicotiana benthamiana homologous with the Arabidopsis U-box protein AtPUB20/21, has been found to be essential for plant defense and disease resistance [41]. According to subsequent genetic structure and motif analysis, genes within the same subfamily were relatively preserved. However, properties of the OsPUB gene and its protein varied widely depending on different classifications. In addition to the U-box domain, 77 OsPUB proteins were found to be able to bind to different domains including ARM/HEAT, TPR domain, GLKs, and kinase. Eight members of PUB genes in rice only housed the domain. The TPR domain was found in OsPUB67 and OsPUB70. WD40 repeats were found in OsPUB 71 and OsPUB72. Therefore, PUB might be functionally diverse in plants. Analysis of the cis-acting regulatory elements in the promoter indicated that the PUB gene family was involved in stress-related mechanisms, hormone regulation, and cellular development. In our study, ABRE (abscisic acid response) was detected in seven OsPUB genes. However, CGTCA-motif (MeJA-responsiveness) and TGACG-motif (MeJA-responsiveness) were detected in putative promoter regions of all 16 rice OsPUB genes. This result indicates that the PUB gene might play a significant role during abiotic and biotic stresses, inducing ABA signal transduction and methyl jasmonate treatment in rice (Table 2). Based on RNA-Seq data under abiotic and biotic stress treatment conditions, expression levels of 16 OsPUB genes showing a common expression pattern were investigated using qRT-PCR. Interestingly, these 16 selected OsPUB genes showed a moderating expression pattern similar to RNA-sequencing or in silico expression analysis except for OsPUB24 and OsPUB57 under abiotic and biotic stresses. Among these genes, OsPUB21 was confirmed to exhibit statistical significance in expression analysis under both drought stress and rice blast infection conditions. In addition, OsPUB57 revealed a higher expression in the resistant plants carrying the Pi9-resistant gene [48]. These results suggest that these selected genes of the Oryza sativa E3 ubiquitin ligase gene family are responsive to abiotic and biotic stress treatments, indicating potential roles of these genes in stress response [28,29,58].
In summary, characteristics of 77 OsPUB genes in rice were identified. Systematic and comprehensive analyses of the OsPUB gene family were performed, including analysis of conserved domains, phylogenetic relationships, gene structures, chromosome locations, cis-acting elements, and expression patterns. Through transcriptome analysis, we confirmed that 16 OsPUB genes show similar expression patterns in drought stress and rice blast infection response pathways. Numerous cis-acting elements were found in promoter sequences of 16 OsPUB genes, indicating that the OsPUB genes might be involved in complex regulatory networks to control hormones, responses to stress, and cellular development. We performed qRT-PCR on 16 OsPUB genes under drought stress and rice blast infection to further identify the reliability of transcriptome and cis-element analysis data. It was confirmed that expression patterns were similar to RNA-sequencing analysis results except for 2 out of 16 genes. Transcriptome and qRT-PCR analyses indicated that sixteen OsPUB genes might play an important role in plant stress resistance mechanisms by showing expression patterns common to multiple stress response pathways. Taken together, these results indicate that genome-wide analysis of the OsPUB genes can provide a solid basis for functional analysis of rice E3 ubiquitin ligase genes. In addition, our study provides a theoretical basis for the future cultivation and breeding of crops suffering from environmental stress by characterizing and classifying U-box E3 ubiquitin ligase in rice, gene expression patterns, and cis-acting regulatory elements. The molecular information of the U-box E3 ubiquitin ligase gene family in rice, including gene expression patterns and cis-acting regulatory elements, could be useful for a future crop breeding program for producing stress-tolerant crops.
4. Materials and Methods
4.1. Identification of U-box E3 Ubiquitin Ligase Gene Family in Rice
All genomic and protein sequences of PUB genes of Oryza sativa were retrieved from Gramene (http://www.gramene.org accessed on 3 August 2020), Rice Genome Annotation Project (rice.plantbiolog.msu.edu accessed on 3 August 2020), and NCBI (http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/gene accessed on 3 August 2020). Molecular weight, pI (isoelectric point), instability index, aliphatic index, GRAVY (grand average of hydropathicity), number of amino acids, and number of nucleotides were analyzed using ProtParam and the compute pI/Mw tool from ExPasy (http://www.expasy.org/tools/ accessed on 3 August 2020). Chromosomal locations of the OsPUB genes family were mapped using the MapGene2Chromosome V2 program (http://mg2c.iask.in/mg2c_v2.0/ accessed on 7 May 2021).
4.2. Phylogenetic and Gene Strucure Analyses of OsPUB Gene Family
To identify and visualize the structural organization of the rice U-box E3 gene family, exon and intron structures of individual OsPUB genes were determined using GSDS (Gene structure display server; http://gsds.cbi.pku.edu.cn/ accessed on 20 November 2020) via the alignment of cDNAs with their corresponding genomic DNA sequences. The multiple sequence alignment of 77 OsPUB genes was performed using ClustalW. Results of the multiple sequence alignment were then used to construct a phylogenetic tree with MEGA version 7.0 (https://www.megasoftware.net/ accessed on 15 September 2021). Unrooted trees were constructed with the Neighbor-Joining method using the following parameters: Poisson correction, pairwise deletion, and 1000 bootstrap replicates. Conserved motifs in the rice U-box E3 gene family were identified using MEME suite (http://meme-suite.org/ accessed on 15 September 2021). A total of 10 motifs and a width limit of 200 amino acids with other default parameters were used for the analysis.
4.3. Prediction of Cis-Acting Elements in OsPUB Genes
PUB family gene promoters (3000 bp upstream of the translation start site) were obtained from the Rice Annotation Project Database (https://rapdb.dna.affrc.go.jp/index.html accessed on 23 August 2021). Stress response and hormone-related cis-acting regulatory elements in promoter sequences were surveyed using the PlantCARE database (http://bioinformatics.psb.ugent.be/webtools/plantcare/html accessed on 28 August 2021).
4.4. Plant Materials and Stress Treatment
For screening drought stress tolerance, four-week-old plants (O. sativa spp. japonica via Dongjin) in the greenhouse were transplanted into soils with a 5% moisture content. Screening was then performed without watering until the drought phenotype had appeared. The soil moisture content was measured using a Pro check device (Decagon devices, Pullman, WA, USA). Leaf samples were collected at 0, 1, and 3 days after treatment. Rice samples infected with Magnaporthe oryzae strains were collected from an experimental field of Chungbuk National University to screen gene expression against biological stress. Inoculated leaves with three biological replicates were harvested at 0 hpi, 12 hpi, and 24 hpi. The collected leaves were immediately frozen in liquid nitrogen and stored at −80 °C until RNA extraction. Total RNA was extracted from leaf tissues using the RNeasy Plant Mini Kit (QIAGEN, Germantown, MD, USA) according to the manufacturer’s instructions. The relative purity and concentration of RNA were estimated using the NanoDrop One (Thermo Fisher Scientific, Thermo Fisher Scientific, MA, USA) and checked on a 1.2% agarose gel. RNA samples were then stored in a −80 °C freezer.
4.5. cDNA Library Construction and RNA-Sequencing
Prior to library preparation, total RNAs of all samples were pooled for each experimental group. First-strand cDNAs were synthesized using the Oligo (dT)20 primer and ReverTra AceTM qPCR RT Master Mix (TOYOBO, Osaka, Japan). RNA-seq libraries for sequencing were prepared in triplicate and constructed according to the manufacturer’s protocol provided by Illumina Inc. (San Diego, CA, USA). Transcription profiles of rice seedlings were determined by RNA-seq on a DNACARE platform (http://www.dnacare.co.kr/web/ accessed on 16 December 2020) using Illumina HiSeq 2500 (Biomarker Technology Co., Rohnert Park, CA, USA), which generated raw data of 150-bp paired-end reads. RNA-seq reads were sorted after filtering adaptor sequences and low-quality bases. They were then mapped to the Os-Nipponbare-Reference IRGSP-1.0 reference genome (http://rapdb.dna.affrc.go.jp/download/archive/irgsp1/IRGSP-1.0_genome.fasta.gz accessed on 16 December 2020) using Trimmomatic v0.38. Complete library read alignment files were integrated from RAPDB (http://rapdb.dna.affrc.go.jp/download/archive/irgsp1 accessed on 16 December 2020) and MSU 7.0 databases (ftp://ftp.plantbiology.msu.edu/pub/data/Eukaryotic_Projects/o_sativa/annotation_dbs/pseudomolecules/version_7.0/all.dir/all.gff3 accessed on 16 December 2020) using HISAT2.
4.6. Differential Expression Analysis
Expression levels of genes were calculated and normalized to fragments per kilobase of transcript per killion fragments mapped (FPKM) to identify differentially expressed genes (DEGs). StringTie v1.3.4d is a software for estimating gene and transcript expression levels from RNA-Seq data. The measurement result of the transcript expression level through assembly was used for DEG analysis. DESeq is a tool for analyzing the difference in expression levels of RNA-Seq expression profiles, and it provides a normalization between comparative samples that minimizes the loss of biological significance by implementing various statistical methods. DEGs were first selected using Student’s t-test (p < 0.05) and log2FC (fold change) > 1 between before stress treatment and after stress treatment.
4.7. Expression Analysis of OsPUB Genes
For qRT-PCR, first-strand cDNAs were synthesized using the Oligo (dT)20 primer and ReverTra AceTM qPCR RT Master Mix (TOYOBO, Osaka, Japan). Primer sets were designed through primer3plus software (Supplementary Table S2). qRT-PCR was performed using iQ SYBR Green Supermix (Bio-Rad, Hercules, CA, USA) on a BioRad CFX96 Detection System (Bio-Rad Laboratories, Hercules, CA, USA) according to the manufacturer’s instructions. Primers for actin as an internal control were used to normalize the results of real-time qRT-PCR.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/ijms222112088/s1.
Author Contributions
Experimental analysis and writing article, M.-S.K.; review and editing, K.-K.K.; supervision, idea development, and editing, Y.-G.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a grant (PJ01483602) from the Rural Development Administration of Korea.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This research was supported by a grant (PJ01483602) from the Rural Development Administration of Korea.
Conflicts of Interest
The authors have no conflicts of interest relevant to this study to disclose.
References
- Mukhopadhyay, D.; Riezman, H. Proteasome-independent functions of ubiquitin in endocytosis and signaling. Science 2007, 315, 201–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Qu, B.; Dou, S.; Li, L.; Yin, D.; Pang, Z.; Zhou, Z.; Tian, M.; Liu, G.; Xie, Q.; et al. The E3 ligase OsPUB15 interacts with the receptor-like kinase PID2 and regulates plant cell death and innate immunity. BMC Plant Biol. 2015, 15, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatakeyama, S.; Yada, M.; Matsumoto, M.; Ishida, N.; Nakayama, K. U box proteins as a new family of ubiquitin-protein ligases. J. Biol. Chem. 2001, 276, 33111–33120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, S.K.; Ryu, M.Y.; Kim, J.H.; Hong, J.S.; Oh, T.R.; Kim, W.T.; Yang, S.W. RING E3 ligases: Key regulatory elements are involved in abiotic stress responses in plants. BMB Rep. 2017, 50, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Small, J.; Vierstra, R. The ubiquitin 26S proteasome proteolytic pathway. Annu. Rev. Plant Biol. 2004, 55, 555–590. [Google Scholar] [CrossRef]
- Donna, Y.; Daphne, R. The diversity of plant Y-box E3 ubiquitin ligases: From upstream activators to downstream target substrates. J. Exp. Bot. 2009, 40, 1109–1121. [Google Scholar]
- Dielen, A.S.; Badaoui, S.; Candresse, T.; German-Retana, S. The ubiquitin/26S proteasome system in plant-pathogen interactions: A never-ending hide-and-seek game. Mol. Plant Pathol. 2010, 11, 293–308. [Google Scholar] [CrossRef]
- Downes, B.P.; Stupar, R.M.; Gingerich, D.J.; Vierstra, R.D. The HECT ubiquitin-protein ligase (UPL) family in Arabidopsis: UPL3 has a specific role in trichome development. Plant J. 2003, 35, 729–742. [Google Scholar] [CrossRef]
- Stone, S.L.; Hauksdottir, H.; Troy, A.; Herschleb, J.; Kraft, E.; Callis, J. Functional analysis of the RING-type ubiquitin ligase family of Arabidopsis. Plant Physiol. 2005, 137, 13–30. [Google Scholar] [CrossRef] [Green Version]
- Yee, D.; Goring, D.R. The diversity of plant U-box E3 ubiquitin ligases: From upstream activators to downstream target substrates. J. Exp. Bot. 2009, 60, 1109–1121. [Google Scholar] [CrossRef] [Green Version]
- Aravind, L.; Koonin, E.V. The U box is a modified RING finger—A common domain in ubiquitination. Curr. Biol. 2000, 10, R132–R134. [Google Scholar] [CrossRef] [Green Version]
- Borden, K.L.; Freemont, P.S. The RING finger domain: A recent example of a sequence-structure family. Curr. Opin. Struct. Biol. 1996, 6, 395–401. [Google Scholar] [CrossRef]
- Saurin, A.J.; Borden, K.L.; Boddy, M.N.; Freemont, P.S. Does this have a familiar RING? Trends Biochem. Sci. 1996, 21, 208–214. [Google Scholar] [CrossRef]
- Mudgil, Y.; Shiu, S.H.; Stone, S.L.; Salt, J.N.; Goring, D.R. A large complement of the predicted Arabidopsis ARM repeat proteins are members of the U-box E3 ubiquitin ligase family. Plant Physiol. 2004, 134, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Samuel, M.A.; Salt, J.N.; Shiu, S.H.; Goring, D.R. Multifunctional arm repeat domains in plants. Int. Rev. Cytol. 2006, 253, 1–26. [Google Scholar]
- Wiborg, J.; O’Shea, C.; Skriver, K. Biochemical function of typical and variant Arabidopsis thaliana U-box E3 ubiquitin-protein ligases. Biochem. J. 2008, 413, 447–457. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.H.; Arnaud, D.; Belcram, H.; Falentin, C.; Rouault, P.; Piel, N.; Lucas, M.O.; Just, J.; Renard, M.; Delourme, R.; et al. A dominant point mutation in a RINGv E3 ubiquitin ligase homoeologous gene leads to cleistogamy in Brassica napus. Plant Cell 2012, 24, 4875–4891. [Google Scholar] [CrossRef] [Green Version]
- Vierstra, R.D. The ubiquitin-26S proteasome system at the nexus of plant biology. Nat. Rev. Mol. Cell Biol. 2009, 10, 385–397. [Google Scholar] [CrossRef]
- Finley, D.; Ulrich, H.D.; Sommer, T.; Kaiser, P. The ubiquitin–proteasome system of Saccharomyces cerevisiae. Genetics 2012, 192, 319–360. [Google Scholar] [CrossRef] [Green Version]
- Ju, H.W.; Min, J.H.; Chung, M.S.; Kim, C.S. The atrzf1 mutation of the novel RING-type E3 ubiquitin ligase increases proline contents and enhances drought tolerance in Arabidopsis. Plant Sci. 2013, 203–204, 1–7. [Google Scholar] [CrossRef]
- Adler, G.; Konrad, Z.; Zamir, L.; Mishra, A.K.; Raveh, D.; Bar-Zvi, D. The Arabidopsis paralogs, PUB46 and PUB48, encoding U-box E3 ubiquitin ligases, are essential for plant response to drought stress. BMC Plant Biol. 2017, 17, 8. [Google Scholar] [CrossRef] [Green Version]
- Smalle, J.; Kurepa, J.; Yang, P.; Emborg, T.J.; Babiychuk, E.; Kushnir, S.; Vierstra, R.D. The pleiotropic role of the 26S proteasome subunit RPN10 in Arabidopsis growth and development supports a substrate-specific function in abscisic acid signaling. Plant Cell 2003, 15, 965–980. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Kurepa, J.; Smalle, J.A. The Arabidopsis 26S proteasome subunit RPN1a is required for optimal plant growth and stress responses. Plant Cell Physiol. 2009, 50, 1721–1725. [Google Scholar] [CrossRef] [Green Version]
- Kurepa, J.; Toh-E, A.; Smalle, J.A. 26S proteasome regulatory particle mutants have increased oxidative stress tolerance. Plant J. 2008, 53, 102–114. [Google Scholar] [CrossRef]
- Sakamoto, T.; Kamiya, T.; Sako, K.; Yamaguchi, J.; Yamagami, M.; Fujiwara, T. Arabidopsis thaliana 26S proteasome subunits RPT2a and RPT5a are crucial for zinc deficiency-tolerance. Biosci. Biotechnol. Biochem. 2011, 75, 561–567. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.W.; Callis, J. Independent modulation of Arabidopsis thaliana poly-ubiquitin mRNAs in different organs and in response to environmental changes. Plant J. 1997, 11, 1017–1027. [Google Scholar] [CrossRef]
- Christensen, A.H.; Sharrock, R.A.; Quail, P.H. Maize poly-ubiquitin genes: Structure, thermal perturbation of expression and transcript splicing, and promoter activity following transfer to protoplasts by electroporation. Plant Mol. Biol. 1992, 18, 675–689. [Google Scholar] [CrossRef]
- Kang, H.; Zhang, M.; Zhou, S.; Guo, Q.; Chen, F.; Wu, J.; Wang, W. Overexpression of wheat ubiquitin gene, Ta-Ub2, improves abiotic stress tolerance of Brachypodium distachyon. Plant Sci. 2016, 248, 102–115. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Zhang, J.; Gao, Q.; Xing, S.; Li, F.; Wang, W. Drought tolerance through overexpression of mono-ubiquitin in transgenic tobacco. J. Plant Physiol. 2008, 165, 1745–1755. [Google Scholar] [CrossRef] [PubMed]
- Zhiguo, E.; Zhang, Y.; Li, T.; Wang, L.; Zhao, H. Characterization of the Ubiquitin-Conjugating Enzyme Gene Family in Rice and Evaluation of Expression Profiles under Abiotic Stresses and Hormone Treatments. PLoS ONE 2015, 10, e0122621. [Google Scholar]
- Chung, E.; Cho, C.W.; So, H.A.; Kang, J.S.; Chung, Y.S.; Lee, J.H. Overexpression of VrUBC1, a Mung Bean E2 Ubiquitin-Conjugating Enzyme, Enhances Osmotic Stress Tolerance in Arabidopsis. PLoS ONE 2013, 8, e66056. [Google Scholar] [CrossRef] [Green Version]
- Wan, X.; Mo, A.; Liu, S.; Yang, L.; Li, L. Constitutive expression of a peanut ubiquitin-conjugating enzyme gene in Arabidopsis confers improved water-stress tolerance through regulation of stress-responsive gene expression. J. Biosci. Bioeng. 2011, 111, 478–484. [Google Scholar] [CrossRef]
- Zhou, G.A.; Chang, R.Z.; Qiu, L.J. Overexpression of soybean ubiquitin-conjugating enzyme gene GmUBC2 confers enhanced drought and salt tolerance through modulating abiotic stress-responsive gene expression in Arabidopsis. Plant Mol. Biol. 2010, 72, 357–367. [Google Scholar] [CrossRef] [Green Version]
- Bahmani, R.; Kim, D.; Lee, B.D.; Hwang, S. Over-expression of tobacco UBC1 encoding a ubiquitin-conjugating enzyme increases cadmium tolerance by activating the 20S/26S proteasome and by decreasing Cd accumulation and oxidative stress in tobacco (Nicotiana tabacum). Plant Mol. Biol. 2017, 94, 433–451. [Google Scholar] [CrossRef]
- Zeng, L.R.; Park, C.H.; Venu, R.C.; Gough, J.; Wang, G.L. Classification, expression pattern, and E3 ligase activity assay of rice U-box-containing proteins. Mol. Plant 2008, 1, 800–815. [Google Scholar] [CrossRef] [Green Version]
- Yoo, Y.H.; Jiang, X.; Jung, K.H. An Abiotic Stress Responsive U-Box E3 Ubiquitin Ligase Is Involved in OsGI-Mediating Diurnal Rhythm Regulating Mechanism. Plants 2020, 9, 1071. [Google Scholar] [CrossRef]
- Riggleman, B.; Wieschaus, E.; Schedl, P. Molecular analysis of the armadillo locus: Uniformly distributed transcripts and a protein with novel internal repeats are associated with a Drosophila segment polarity gene. Genes Dev. 1989, 3, 96–113. [Google Scholar] [CrossRef] [Green Version]
- Huber, A.H.; Weis, W.I. The structure of the beta-catenin/E-cadherin complex and the molecular basis of diverse ligand recognition by beta-catenin. Cell 2001, 105, 391–402. [Google Scholar] [CrossRef] [Green Version]
- Andrade, M.A.; Bork, P. HEAT repeats in the Huntington’s disease protein. Nat. Genet. 1995, 11, 115–116. [Google Scholar] [CrossRef]
- Azevedo, C.; Santos-Rosa, M.J.; Shirasu, K. The U-box protein family in plants. Trends Plant Sci. 2001, 6, 354–358. [Google Scholar] [CrossRef]
- Gonzalez-Lamothe, R.; Tsitsigiannis, D.I.; Ludwig, A.A.; Panicot, M.; Shirasu, K.; Jones, J.D. The U-box protein CMPG1 is required for efficient activation of defense mechanisms triggered by multiple resistance genes in tobacco and tomato. Plant Cell 2006, 18, 1067–1083. [Google Scholar] [CrossRef] [Green Version]
- Das, A.K.; Cohen, P.W.; Barford, D. The structure of the tetratricopeptide repeats of protein phosphatase 5: Implications for TPR-mediated protein-protein interactions. EMBO J. 1998, 17, 1192–1199. [Google Scholar] [CrossRef] [Green Version]
- D’Andrea, L.D.; Regan, L. TPR proteins: The versatile helix. Trends Biochem. Sci. 2003, 28, 655–662. [Google Scholar] [CrossRef]
- Hanks, S.K.; Quinn, A.M.; Hunter, T. The protein kinase family: Conserved features and deduced phylogeny of the catalytic domains. Science 1988, 241, 42–52. [Google Scholar] [CrossRef]
- Manning, G.; Plowman, G.D.; Hunter, T.; Sudarsanam, S. Evolution of protein kinase signaling from yeast to man. Trends Biochem. Sci. 2002, 27, 514–520. [Google Scholar] [CrossRef]
- Siberil, Y.; Doireau, P.; Gantet, P. Plant bZIP G-box binding factors. Modular structure and activation mechanisms. Eur. J. Biochem. 2001, 268, 5655–5666. [Google Scholar] [CrossRef]
- Qin, Q.; Wang, Y.X.; Huang, L.Y.; Du, F.P.; Zhao, X.Q.; Li, Z.K.; Wang, W.S.; Fu, Y.B. A U-box E3 ubiquitin ligase OsPUB67 is positively involved in drought tolerance in rice. Plant Mol. Biol. 2020, 102, 89–107. [Google Scholar] [CrossRef]
- Qu, S.H.; Liu, G.F.; Zhou, B.; Bellizzi, M.; Zeng, L.R.; Dai, L.Y.; Han, H.; Wang, G.L. The broad-spectrum blast resistance gene Pi9 encodes an NBS-LRR protein and is a member of a multigene family in rice. Genetics 2006, 172, 1901–1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.J.; Yi, J.; Yoon, J.; Cho, L.H.; Ping, J.; Jeong, H.J.; Cho, S.K.; Kim, W.T.; An, G. OsPUB15, an E3 ubiquitin ligase, functions to reduce cellular oxidative stress during seedling establishment. Plant J. 2011, 65, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Seo, D.H.; Ryu, M.Y.; Jammes, F.; Hwang, J.H.; Turek, M.; Kang, B.G.; Kwak, J.M.; Kim, W.T. Roles of four Arabidopsis U-box E3 ubiquitin ligases in negative regulation of abscisic acid-mediated drought stress responses. Plant Physiol. 2012, 160, 556–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.C.; Wu, Y.R.; Huang, X.H.; Sun, J.; Xie, Q. AtPUB19, a U-box E3 ubiquitin ligase, negatively regulates abscisic acid and drought responses in Arabidopsis thaliana. Mol. Plant 2011, 4, 938–946. [Google Scholar] [CrossRef]
- Wang, N.; Liu, Y.; Cong, Y.; Wang, T.; Zhong, X.; Yang, S.; Li, Y.; Gai, J. Genome-Wide Identification of Soybean U-Box E3 Ubiquitin Ligases and Roles of GmPUB8 in Negative Regulation of Drought Stress Response in Arabidopsis. Plant Cell Physiol. 2016, 57, 1189–1209. [Google Scholar] [CrossRef] [Green Version]
- Luo, Q.; Li, Y.; Wang, W.; Fei, X.; Deng, X. Genome-wide survey and expression analysis of Chlamydomonas reinhardtii U-box E3 ubiquitin ligases (CrPUBs) reveal a functional lipid metabolism module. PLoS ONE 2015, 10, e0122600. [Google Scholar] [CrossRef]
- Ryu, M.Y.; Cho, S.K.; Hong, Y.; Kim, J.; Kim, J.H.; Kim, G.M.; Chen, Y.J.; Knoch, E.; Møller, B.L.; Kim, W.T.; et al. Classification of barley U-box E3 ligases and their expression patterns in response to drought and pathogen stresses. BMC Genom. 2019, 20, 326. [Google Scholar] [CrossRef]
- Zeng, L.R.; Qu, S.; Bordeos, A.; Yang, C.; Baraoidan, M.; Yan, H.; Xie, Q.; Nahm, B.H.; Leung, H.; Wang, G.L. Spotted leaf11, a negative regulator of plant cell death and defense, encodes a U-box/armadillo repeat protein endowed with E3 ubiquitin ligase activity. Plant Cell 2004, 16, 2795–2808. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.K.; Chung, H.S.; Ryu, M.Y.; Park, M.J.; Lee, M.M.; Bahk, Y.Y.; Kim, J.; Pai, H.S.; Kim, W.T. Heterologous expression and molecular and cellular characterization of CaPUB1 encoding a hot pepper U-Box E3 ubiquitin ligase homolog. Plant Physiol. 2006, 142, 1664–1682. [Google Scholar] [CrossRef] [Green Version]
- Amador, V.; Monte, E.; Garcia-Martinez, J.L.; Prat, S. Gibberellins signal nuclear import of PHOR1, a photoperiod-responsive protein with homology to Drosophila armadillo. Cell 2001, 106, 343–354. [Google Scholar] [CrossRef]
- Park, J.W.; Gileung, L.; Jin-Baek, K.; Hong-Il, C. RNA Sequencing-Based Transcriptome Analysis in Response to Different Types and Doses of Ionizing Radiation in Rice. Plant Breed. Biotech. 2021, 9, 213–226. [Google Scholar] [CrossRef]
Figure 1.
Chromosome mapping of U-box E3 ubiquitin ligase gene family in Oryza sativa. The rice U-box gene family is divided into different classes shown by different colors: red color, Class 2; green color, Class 3; blue color, Class 4; black color, Class 1, Class 5, Class 6, Class 7, and Class 8. Chromosome number is shown at the top of the bar. The scale bar on the left indicates chromosome length (Mb).
Figure 1.
Chromosome mapping of U-box E3 ubiquitin ligase gene family in Oryza sativa. The rice U-box gene family is divided into different classes shown by different colors: red color, Class 2; green color, Class 3; blue color, Class 4; black color, Class 1, Class 5, Class 6, Class 7, and Class 8. Chromosome number is shown at the top of the bar. The scale bar on the left indicates chromosome length (Mb).

Figure 2.
Phylogenetic tree of U-box E3 ubiquitin ligase gene family members in Oryza sativa constructed with the Neighbor-Joining method. The rice U-box gene family is grouped into different classes shown by different colors. Blue color represents Class 2. Pink color indicates Class 3. Yellow color represents Class 4. White color indicates Class 1, Class 5, Class 6, Class 7, and Class 8.
Figure 2.
Phylogenetic tree of U-box E3 ubiquitin ligase gene family members in Oryza sativa constructed with the Neighbor-Joining method. The rice U-box gene family is grouped into different classes shown by different colors. Blue color represents Class 2. Pink color indicates Class 3. Yellow color represents Class 4. White color indicates Class 1, Class 5, Class 6, Class 7, and Class 8.

Figure 3.
Exon/intron distribution of 77 rice U-box gene family members was analyzed with the GSDS tool. Coding sequences were compared with their corresponding genomic sequences. The orange box represents the CDS. The continuous black line represents the intron region. Blue boxes represent upstream/downstream regions.
Figure 3.
Exon/intron distribution of 77 rice U-box gene family members was analyzed with the GSDS tool. Coding sequences were compared with their corresponding genomic sequences. The orange box represents the CDS. The continuous black line represents the intron region. Blue boxes represent upstream/downstream regions.

Figure 4.
Gene structure and conserved protein sequence patterns of OsPUB gene family. (A) Conserved motifs in OsPUB genes. These motifs were identified with the MEME program. Ten conserved motifs are displayed with different colored boxes. (B) Sequence logos of 10 conserved motifs. The height of the letter showing amino acid residue at each position represents the degree of conservation. Numbers on the x-axis represent residue positions in motifs. The y-axis represents content measured in bits.
Figure 4.
Gene structure and conserved protein sequence patterns of OsPUB gene family. (A) Conserved motifs in OsPUB genes. These motifs were identified with the MEME program. Ten conserved motifs are displayed with different colored boxes. (B) Sequence logos of 10 conserved motifs. The height of the letter showing amino acid residue at each position represents the degree of conservation. Numbers on the x-axis represent residue positions in motifs. The y-axis represents content measured in bits.

Figure 5.
Expression patterns of 77 rice U-box gene family members in leaves under drought stress. (A) Heatmap showing expression of total differential expressed genes with increased or decreased trend under drought treatment conditions. (B) Total number of upregulated and downregulated genes. (C) Venn diagram of upregulated genes in the two samples vs. control. (D) Venn diagram of downregulated genes in the two samples vs. control. Two samples are 1 DAT and 3 DAT.
Figure 5.
Expression patterns of 77 rice U-box gene family members in leaves under drought stress. (A) Heatmap showing expression of total differential expressed genes with increased or decreased trend under drought treatment conditions. (B) Total number of upregulated and downregulated genes. (C) Venn diagram of upregulated genes in the two samples vs. control. (D) Venn diagram of downregulated genes in the two samples vs. control. Two samples are 1 DAT and 3 DAT.

Figure 6.
Expression patterns of 77 rice U-box gene family members in indica rice plants infected with Magnaporthe oryzae. (A) Heatmap showing expression of total differential expressed genes with increased or decreased trend. (B) Total number of upregulated and downregulated genes. (C) Venn diagram of upregulated genes in the two samples vs. control. (D) Venn diagram of downregulated genes in the two samples vs. control. Two samples are 12 hpi and 24 hpi.
Figure 6.
Expression patterns of 77 rice U-box gene family members in indica rice plants infected with Magnaporthe oryzae. (A) Heatmap showing expression of total differential expressed genes with increased or decreased trend. (B) Total number of upregulated and downregulated genes. (C) Venn diagram of upregulated genes in the two samples vs. control. (D) Venn diagram of downregulated genes in the two samples vs. control. Two samples are 12 hpi and 24 hpi.

Figure 7.
Distribution of stress-related cis-acting elements in promoter sequences of 16 OsPUB genes. Cis-elements distributed on the sense strand and the reverse strand are indicated above and below black lines, respectively. The 3 kb sequences upstream of the Transcription Start Site (TSS) of OsPUB genes can be estimated using the scale per 500 bp at the above. 3-AF1 binding site: light responsive element; ABRE: cis-acting element involved in the abscisic acid responsiveness; ARE: cis-acting regulatory element essential for the anaerobic induction; AT1-motif: part of a light responsive module; CGTCA-motif: cis-acting regulatory element involved in the MeJA-responsiveness; chu-Unit 1 m1: part of a light responsive element; GATA-motif: part of a light responsive element; G-box: cis-acting regulatory element involved in light responsiveness; GC-motif: enhancer-like element involved in anoxic specific inducibility; GT1-motif: light responsive element; I-box: part of a light responsive element; LTR: cis-acting element involved in low-temperature responsiveness; O2-site: cis-acting regulatory element involved in zein metabolism regulation; P-box: gibberellin-responsive element; Sp1: light responsive element; TATA-box: core promoter element around −30 of transcription start; TATC-box: core promoter element around −30 of transcription start; TGACG-motif: cis-acting regulatory element involved in MeJA-responsiveness. The cis-acting regulatory elements of Oryza sativa are indicated by a bold border and written inside a box. Cis-elements for other crops (Zea mays, Hordeum vulgare, Triticum aestivum, and Solanum tuberosum) are displayed only with thin borders. * The regions having several boxes (such as 1800 bp of OsPUB21) are with several cis-elements in a 100 bp interval. The closest box on the line means near to the starting bp of each multiple-box region.
Figure 7.
Distribution of stress-related cis-acting elements in promoter sequences of 16 OsPUB genes. Cis-elements distributed on the sense strand and the reverse strand are indicated above and below black lines, respectively. The 3 kb sequences upstream of the Transcription Start Site (TSS) of OsPUB genes can be estimated using the scale per 500 bp at the above. 3-AF1 binding site: light responsive element; ABRE: cis-acting element involved in the abscisic acid responsiveness; ARE: cis-acting regulatory element essential for the anaerobic induction; AT1-motif: part of a light responsive module; CGTCA-motif: cis-acting regulatory element involved in the MeJA-responsiveness; chu-Unit 1 m1: part of a light responsive element; GATA-motif: part of a light responsive element; G-box: cis-acting regulatory element involved in light responsiveness; GC-motif: enhancer-like element involved in anoxic specific inducibility; GT1-motif: light responsive element; I-box: part of a light responsive element; LTR: cis-acting element involved in low-temperature responsiveness; O2-site: cis-acting regulatory element involved in zein metabolism regulation; P-box: gibberellin-responsive element; Sp1: light responsive element; TATA-box: core promoter element around −30 of transcription start; TATC-box: core promoter element around −30 of transcription start; TGACG-motif: cis-acting regulatory element involved in MeJA-responsiveness. The cis-acting regulatory elements of Oryza sativa are indicated by a bold border and written inside a box. Cis-elements for other crops (Zea mays, Hordeum vulgare, Triticum aestivum, and Solanum tuberosum) are displayed only with thin borders. * The regions having several boxes (such as 1800 bp of OsPUB21) are with several cis-elements in a 100 bp interval. The closest box on the line means near to the starting bp of each multiple-box region.

Figure 8.
Expression patterns of OsPUB genes in response to drought stress treatment analyzed by qRT-PCR. Relative gene expression level was normalized against rice Actin gene. Values are presented as means ± standard error. Three independent biological repeats were performed. Bars represent standard deviations (SD) of three technical replicates. A significant difference is indicated by an asterisk according to t-test (* p < 0.05, ** p < 0.01, and *** p < 0.001).
Figure 8.
Expression patterns of OsPUB genes in response to drought stress treatment analyzed by qRT-PCR. Relative gene expression level was normalized against rice Actin gene. Values are presented as means ± standard error. Three independent biological repeats were performed. Bars represent standard deviations (SD) of three technical replicates. A significant difference is indicated by an asterisk according to t-test (* p < 0.05, ** p < 0.01, and *** p < 0.001).

Figure 9.
Expression patterns of OsPUB genes in response to infection of Magnaporthe oryzae analyzed by qRT-PCR. Relative gene expression was normalized against rice Actin gene. Values are presented as means ± standard error. Three independent biological repeats were performed. Bars represent standard deviations (SD) of three technical replicates. A significant difference is indicated by an asterisk according to t-test (* p < 0.05, ** p < 0.01, and *** p < 0.001).
Figure 9.
Expression patterns of OsPUB genes in response to infection of Magnaporthe oryzae analyzed by qRT-PCR. Relative gene expression was normalized against rice Actin gene. Values are presented as means ± standard error. Three independent biological repeats were performed. Bars represent standard deviations (SD) of three technical replicates. A significant difference is indicated by an asterisk according to t-test (* p < 0.05, ** p < 0.01, and *** p < 0.001).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
A list of OsPUB genes showing the same expression pattern under abiotic stress (drought condition) and biotic stress (blast infection).
Table 1.
A list of OsPUB genes showing the same expression pattern under abiotic stress (drought condition) and biotic stress (blast infection).
| No. | Gene Name | Locus No. | Class | Common Regulation | Drought Stress | Blast Infection | ||
|---|---|---|---|---|---|---|---|---|
| Log2FC (0 DAT vs. 1 DAT) | Log2FC (0 DAT vs. 3 DAT) | Log2FC (0 hpi vs. 12 hpi) | Log2FC (0 hpi vs. 24 hpi) | |||||
| 1 | OsPUB08 | LOC_Os02g28720 | 2 | UP | 1.223 ** | 2.485 | 0.208 | 0.209 |
| 2 | OsPUB21 | LOC_Os02g49520 | 2 | DOWN | −3.078 ** | −5.808 ** | −1.501 * | −1.686 ** |
| 3 | OsPUB23 | LOC_Os12g17900 | 2 | DOWN | −0.442 | −0.694 | −0.039 | −0.136 |
| 4 | OsPUB24 | LOC_Os03g45420 | 2 | UP | 1.850 * | 3.365 ** | 1.416 | 1.456 ** |
| 5 | OsPUB26 | LOC_Os03g60140 | 5 | DOWN | −0.338 | −1.248 | −0.402 | −0.679 |
| 6 | OsPUB28 | LOC_Os01g67500 | 2 | DOWN | −0.071 ** | −1.764 *** | −0.739 ** | −0.752 ** |
| 7 | OsPUB37 | LOC_Os12g06410 | 3 | UP | 0.069 * | 0.248 * | 1.884 ** | 2.165 *** |
| 8 | OsPUB38 | LOC_Os04g35680 | 3 | UP | 0.493 | 1.910 ** | 3.422 | 3.852 |
| 9 | OsPUB41 | LOC_Os03g13740 | 3 | UP | 2.724 * | 3.576 ** | 3.578 ** | 3.723 * |
| 10 | OsPUB54 | LOC_Os10g40100 | 4 | DOWN | −8.452 | −0.141 | −0.686 | −0.878 |
| 11 | OsPUB56 | LOC_Os09g39640 | 4 | DOWN | −0.468 | −1.457 | −0.149 | −0.228 |
| 12 | OsPUB57 | LOC_Os03g31070 | 4 | DOWN | −1.264 * | −2.056 ** | −0.406 | −0.718 * |
| 13 | OsPUB60 | LOC_Os02g44599 | 4 | DOWN | −0.768 | −2.068 | −0.527 | −0.567 |
| 14 | OsPUB66 | LOC_Os05g01460 | 7 | UP | 0.749 | 1.935 | 0.158 | 0.247 |
| 15 | OsPUB67 | LOC_Os10g40490 | 7 | UP | 3.412 ** | 4.337 ** | 1.478 ** | 1.531 ** |
| 16 | OsPUB70 | LOC_Os06g06760 | 8 | DOWN | −3.998 | −1.096 | −0.104 | −0.482 |
* Three independent biological repeats were performed. A significant difference is indicated by asterisks according to t-test (* p < 0.05, ** p < 0.01, *** p < 0.001).
Table 2.
Category of cis-elements extracted from 3000 bp upstream region of OsPUB genes using PlantCARE.
Table 2.
Category of cis-elements extracted from 3000 bp upstream region of OsPUB genes using PlantCARE.
| Function | Cis-Element | Sequence | Presence of Cis-Element in PUB Genes | |
|---|---|---|---|---|
| In PUB Genes | In Diffenrent Species | |||
| Cis-acting element involved in the abscisic acid responsiveness | ABRE | GCCGCGTGGC | OsPUB8, OsPUB21, OsPUB60, OsPUB67 | Oryza sativa |
| GACACGTGGC | OsPUB37, OsPUB70 | Triticum aestuvum | ||
| CGCACGTGTC | OsPUB21, OsPUB38, OsPUB60 | Hordeum vulgare | ||
| Cis-acting regulatory element essential for the anaerobic induction | ARE | AACCA | OsPUB8, OsPUB21, OsPUB23, OsPUB24, OsPUB26, OsPUB28, OsPUB37, OsPUB38, OsPUB41, OsPUB54, OsPUB56, OsPUB57, OsPUB60, OsPUB66, OsPUB67, OsPUB70 | Zea mays |
| Cis-acting regulatory element involved in the MeJA-responsiveness | TGACG | TGACG | OsPUB8, OsPUB21, OsPUB23, OsPUB24, OsPUB26, OsPUB28, OsPUB37, OsPUB38, OsPUB41, OsPUB54, OsPUB56, OsPUB57, OsPUB60, OsPUB66, OsPUB67, OsPUB70 | Hordeum vulgare |
| CGTCA | CGTCA | |||
| Cis-acting regulatory element involved in zein metabolism regulation | O2-site | GATGACATGG | OsPUB37, OsPUB70 | Zea mays |
| GATGATGTGG | OsPUB21, OsPUB26, OsPUB56, OsPUB57 | |||
| GTTGACGTGA | OsPUB21, OsPUB23, OsPUB24, OsPUB57, OsPUB60 | |||
| Enhancer-like element involved in anoxic specific inducibility | GC-motif | CCCCCG | OsPUB8, OsPUB21, OsPUB28, OsPUB37, OsPUB54, OsPUB57, OsPUB60, OsPUB70 | Zea mays |
| Cis-acting element involved in gibberellin-responsiveness | TATC-box | TATCCCA | OsPUB23, OsPUB54 | Oryza sativa |
| Gibberellin-responsive element | P-box | CCTTTTG | OSPUB8, OsPUB21, OsPUB23, OsPUB24, OsPUB38, OsPUB60, OsPUB66, OsPUB67 | Oryza sativa |
| Cis-acting element involved in low-temperature responsiveness | LTR | CCGAAA | OsPUB8, OsPUB21, OsPUB23, OsPUB37, OsPUB41, OsPUB60, OsPUB66 | Hordeum vulgare |
| Cis-acting regulatory element involved in light responsiveness | G-box | TCCACATGGCA | OsPUB41, OsPUB56 | Triticum aestivum |
| CACGTC | OsPUB8, OsPUB21, OsPUB23, OsPUB24, OsPUB26, OsPUB37, OsPUB38, OsPUB41, OsPUB54, OsPUB56, OsPUB57, OsPUB60, OsPUB66, OsPUB67 | Zea mays | ||
| Part of a light responsive element | chs-Unit 1 m1 | ACCTAACCCGC | OsPUB8 | Hordeum vulgare |
| GATA-motif | AAGGATAAGG | OsPUB8, OsPUB24, OsPUB41, OsPUB56, OsPUB66 | Solanum tuberosum | |
| GTGGC-motif | CAGCGTGTGGC | OsPUB54 | Hordeum vulgare | |
| I-box | AGATAAGG | OsPUB37, OsPUB57 | Triticum aestivum | |
| gGATAAGGTG | OsPUB8, OsPUB37, OsPUB38, OsPUB41, OsPUB60, OsPUB67 | Zea mays | ||
| AT1-motif | AATTATTTTTTATT | OsPUB8, OsPUB41, OsPUB54, OsPUB66, OsPUB67 | Solanum tuberosum | |
| Light responsive element | 3-AF1 binding site | TAAGAGAGGAA | OsPUB56 | Solanum tuberosum |
| GT1-motif | GTGTGTGAA | OsPUB28 | ||
| Sp1 | GGGCGG | OsPUB8, OsPUB21, OsPUB23, OsPUB24, OsPUB26, OsPUB28, OsPUB54, OsPUB57, OsPUB60, OsPUB70 | Oryza sativa | |
| Cis-regulatory element involved in endosperm expression | GCN4_motif | TGAGTCA | OsPUB8, OsPUB21, OsPUB23, OsPUB24, OsPUB37, OsPUB60, OsPUB66 | Oryza sativa |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, M.-S.; Kang, K.-K.; Cho, Y.-G. Molecular and Functional Analysis of U-box E3 Ubiquitin Ligase Gene Family in Rice (Oryza sativa). Int. J. Mol. Sci. 2021, 22, 12088. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222112088
AMA Style
Kim M-S, Kang K-K, Cho Y-G. Molecular and Functional Analysis of U-box E3 Ubiquitin Ligase Gene Family in Rice (Oryza sativa). International Journal of Molecular Sciences. 2021; 22(21):12088. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222112088
Chicago/Turabian StyleKim, Me-Sun, Kwon-Kyoo Kang, and Yong-Gu Cho. 2021. "Molecular and Functional Analysis of U-box E3 Ubiquitin Ligase Gene Family in Rice (Oryza sativa)" International Journal of Molecular Sciences 22, no. 21: 12088. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222112088
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.