
Aquaporin-1 Facilitates Transmesothelial Water Permeability: In Vitro and Ex Vivo Evidence and Possible Implications in Peritoneal Dialysis

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Characterization of HMC
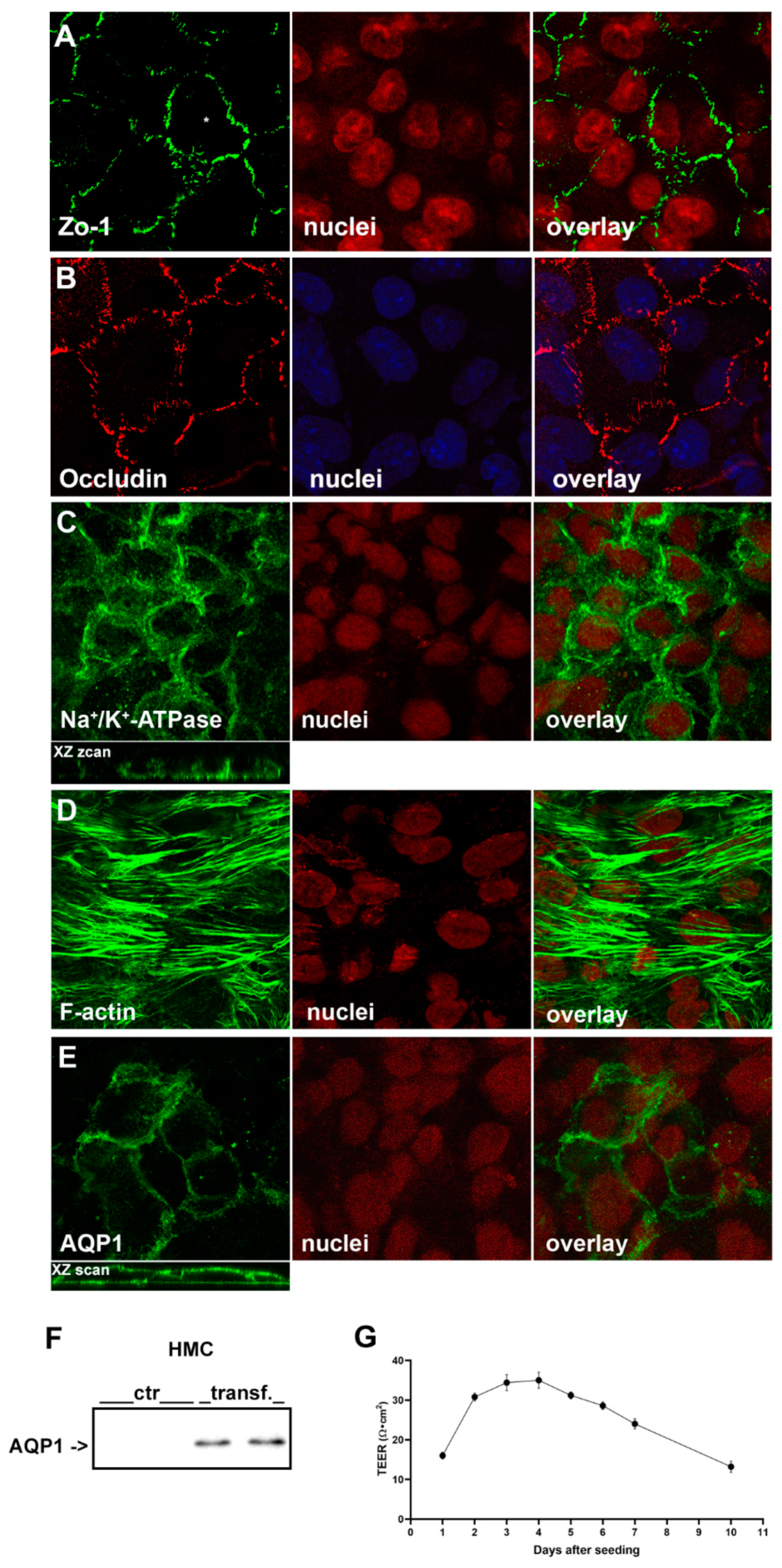
2.1.1. Immunofluorescence Confocal Analysis
2.1.2. Evaluation of HMC Monolayers Polarization
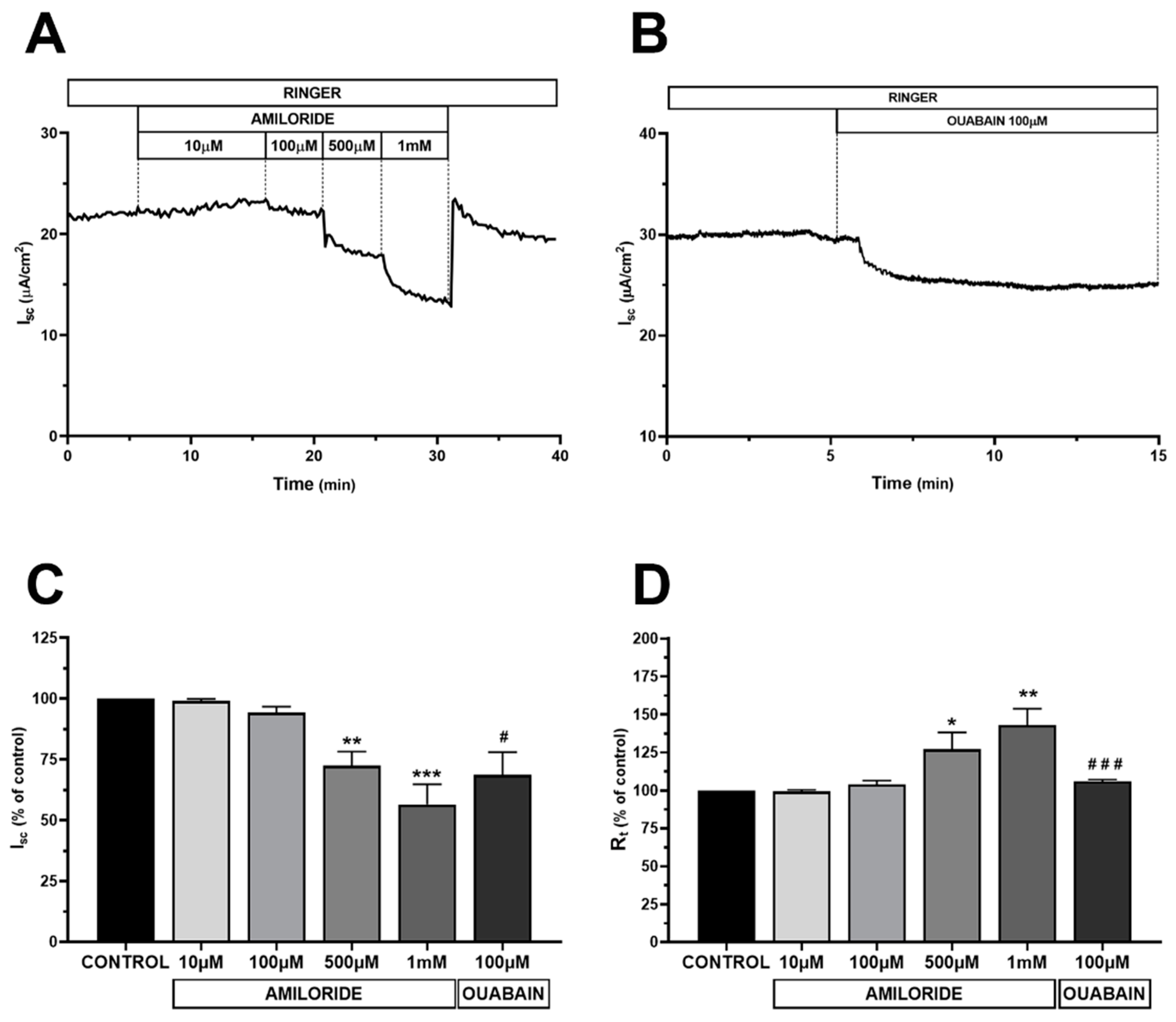
2.1.3. Electrophysiological Characterization of HMC Monolayers
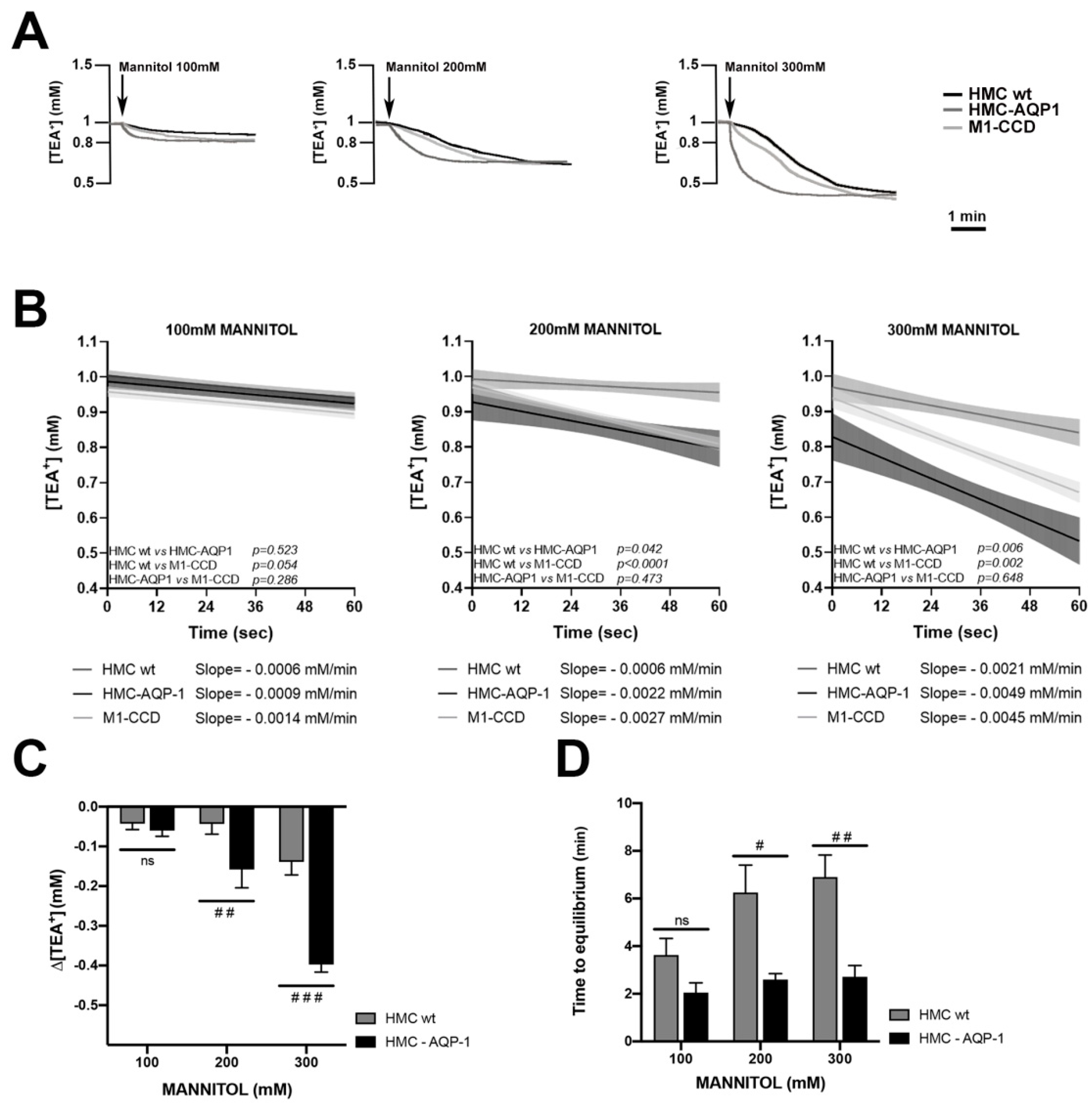
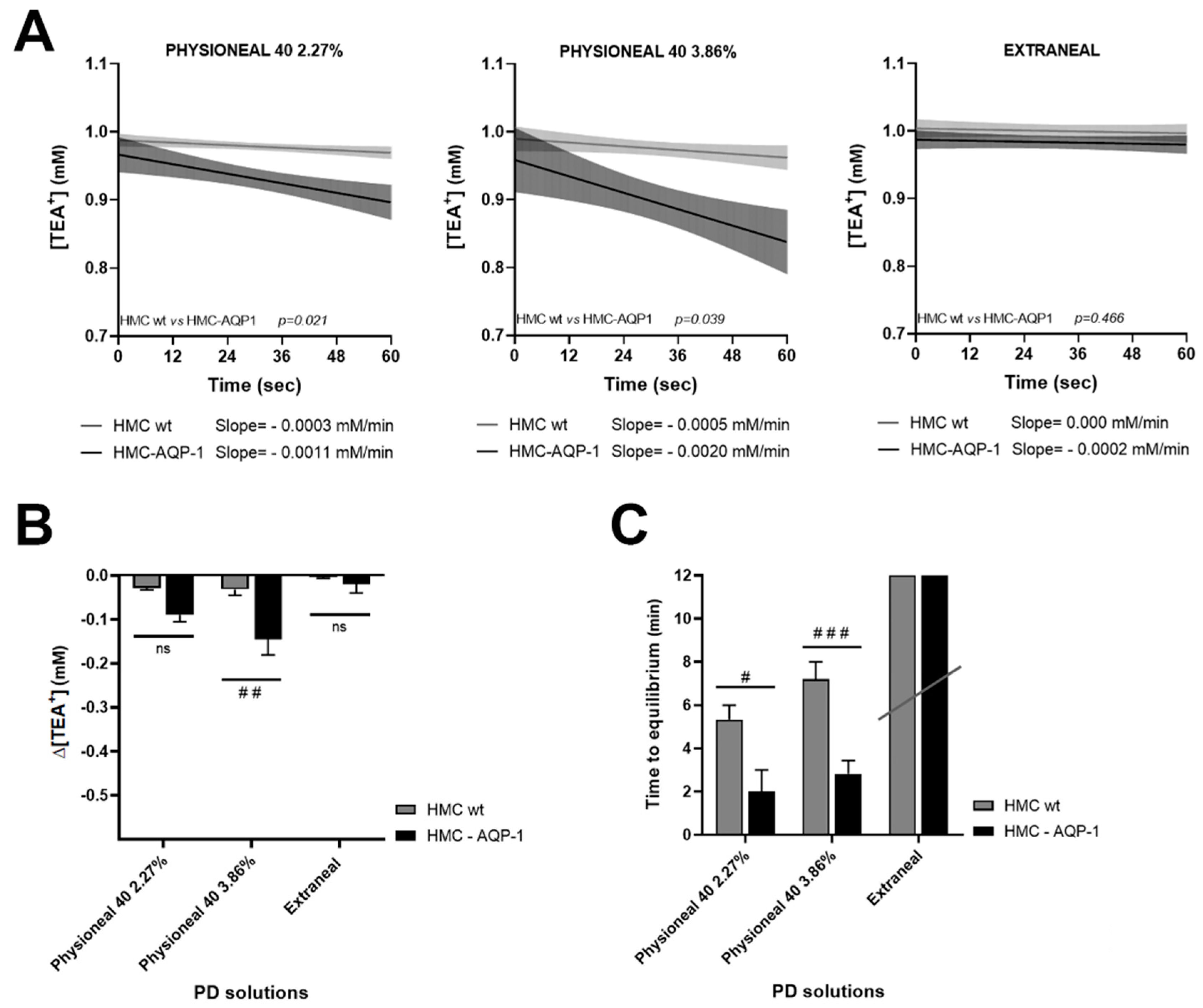
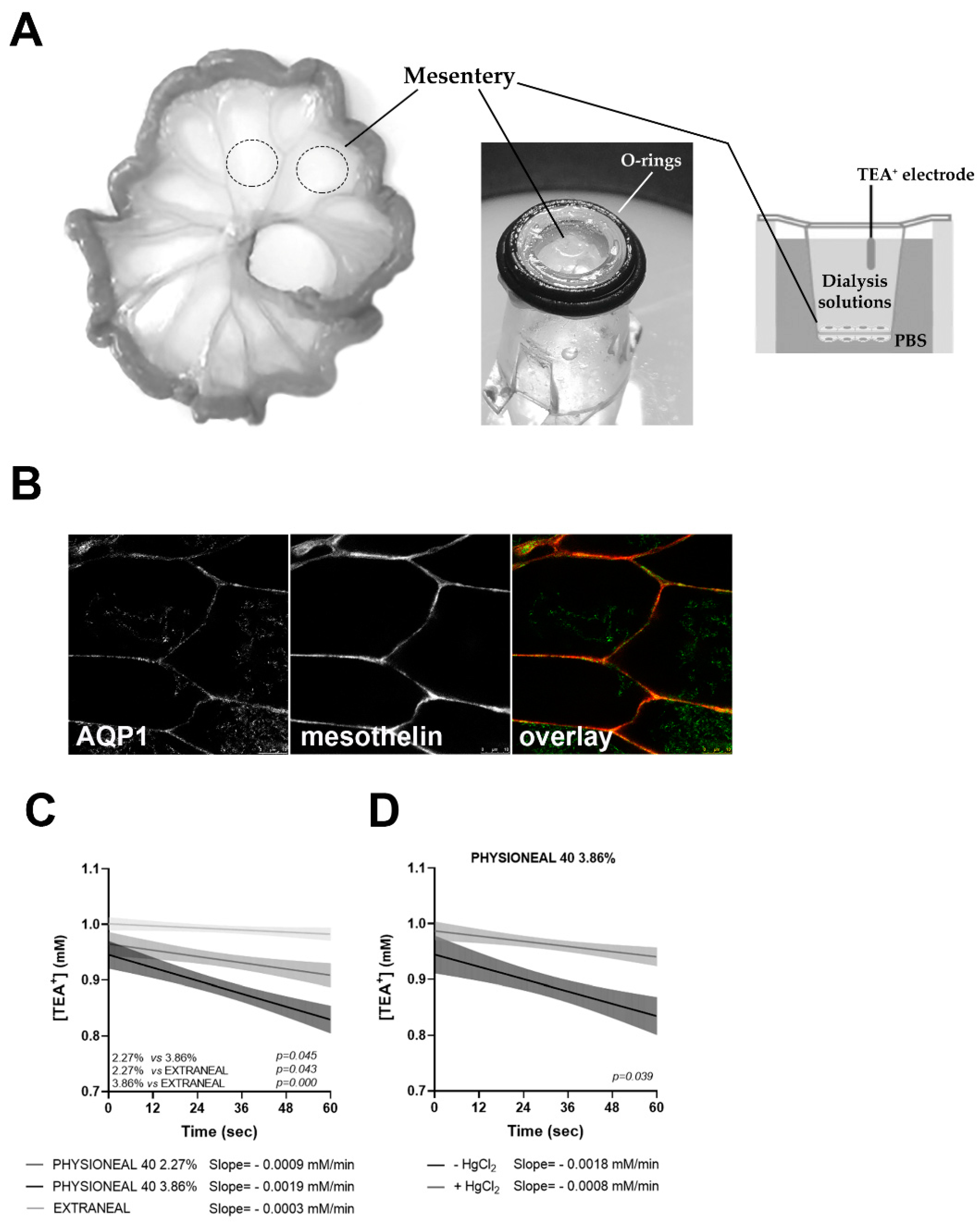
2.2. Real Time Measurements of Transmesothelial Water Flux across the HMC Monolayers and Mouse Mesentery: Role of AQP1
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture
4.3. Transepithelial Electrical Resistance Measurements
4.4. Immunofluorescence and Confocal Microscopy
4.5. Ussing Chamber Studies
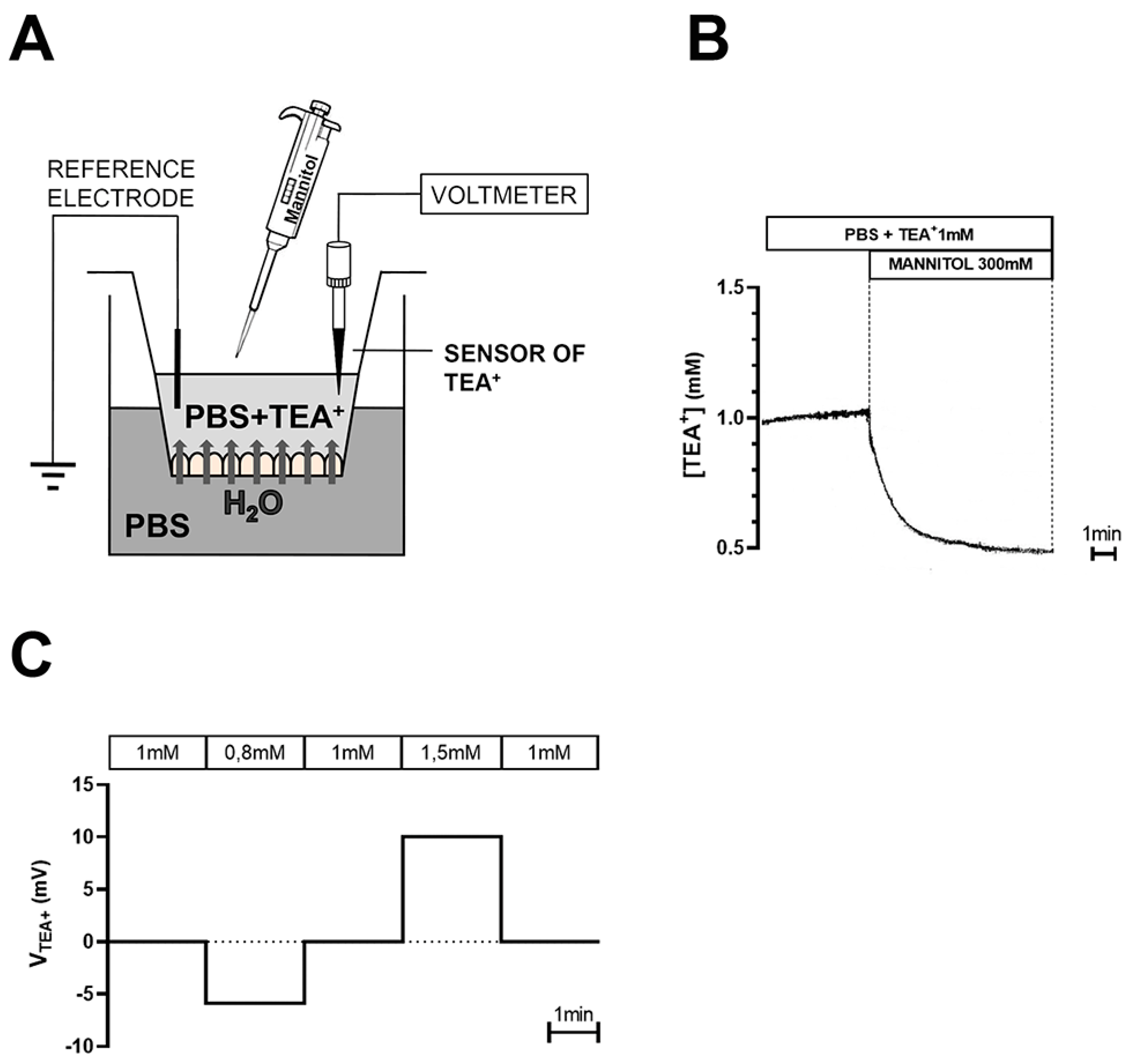
4.6. Transmesothelial Water Flux Measurements
4.7. Data Analysis and Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
Abbreviations
| PD | Peritoneal Dialysis |
| AQP1 | Aquaporin1 |
| UF | Ultrafiltration |
| TEER | TransEpithelial Electrical Resistance |
| ECM | Extracellular matrix |
| HMC | Human Mesothelial Cells |
References
- Devuyst, O.; Rippe, B. Water transport across the peritoneal membrane. Kidney Int. 2014, 85, 750–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutsaers, S.E. Mesothelial cells: Their structure, function and role in serosal repair. Respirology 2002, 7, 171–191. [Google Scholar] [CrossRef] [PubMed]
- Mehrotra, R.; Devuyst, O.; Davies, S.J.; Johnson, D.W. The current state of peritoneal dialysis. J. Am. Soc. Nephrol. 2016, 27, 3238–3252. [Google Scholar] [CrossRef] [Green Version]
- Rippe, B.; Levin, L. Computer simulations of ultrafiltration profiles for an icodextrin-based peritoneal fluid in CAPD. Kidney Int. 2000, 57, 2546–2556. [Google Scholar] [CrossRef] [Green Version]
- Rippe, B.; Stelin, G.; Haraldsson, B. Computer simulations of peritoneal fluid transport in CAPD. Kidney Int. 1991, 40, 315–325. [Google Scholar] [CrossRef] [Green Version]
- Rippe, B.; Venturoli, D.; Simonsen, O.; de Arteaga, J. Fluid and electrolyte transport across the peritoneal membrane during CAPD according to the three-pore model. Perit. Dial. Int. 2004, 24, 10–27. [Google Scholar] [CrossRef]
- Stelin, G.; Rippe, B. A phenomenological interpretation of the variation in dialysate volume with dwell time in CAPD. Kidney Int. 1990, 38, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Agre, P.; Preston, G.M.; Smith, B.L.; Jung, J.S.; Raina, S.; Moon, C.; Guggino, W.B.; Nielsen, S. Aquaporin CHIP: The archetypal molecular water channel. Am. J. Physiol. Physiol. 1993, 265, F463–F476. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.S.; Preston, G.M.; Smith, B.L.; Guggino, W.B.; Agre, P. Molecular structure of the water channel through aquaporin CHIP. The hourglass model. J. Biol. Chem. 1994, 269, 14648–14654. [Google Scholar] [CrossRef]
- Nielsen, S.; Smith, B.L.; Christensen, E.I.; Agre, P. Distribution of the aquaporin CHIP in secretory and resorptive epithelia and capillary endothelia. Proc. Natl. Acad. Sci. USA 1993, 90, 7275–7279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, J.; Verbavatz, J.-M.; Rippe, A.; Boisdé, I.; Moulin, P.; Rippe, B.; Verkman, A.S.; Devuyst, O. Aquaporin-1 plays an essential role in water permeability and ultrafiltration during peritoneal dialysis. Kidney Int. 2006, 69, 1518–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Freichel, M.; Van Der Hoeven, F.; Nawroth, P.P.; Katus, H.; Kälble, F.; Zitron, E.; Schwenger, V. Novel endothelial cell-specific AQP1 knockout mice confirm the crucial role of endothelial AQP1 in ultrafiltration during peritoneal dialysis. PLoS ONE 2016, 11, e0145513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devuyst, O.; Goffin, E. Water and solute transport in peritoneal dialysis: Models and clinical applications. Nephrol. Dial. Transplant. 2008, 23, 2120–2123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rippe, B.; Krediet, R.T. Peritoneal physiology-transport of solutes. In The Textbook of Peritoneal Dialysis; Springer: Dordrecht, The Netherlands, 1994. [Google Scholar]
- Marples, D. Aquaporins: Roles in renal function and peritoneal dialysis. Perit. Dial. Int. 2001, 21, 212–218. [Google Scholar] [CrossRef]
- Schoenicke, G.; Diamant, R.; Donner, A.; Roehrborn, A.; Grabensee, B.; Plum, J. Histochemical distribution and expression of aquaporin 1 in the peritoneum of patients undergoing peritoneal dialysis: Relation to peritoneal transport. Am. J. Kidney Dis. 2004, 44, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Corciulo, S.; Nicoletti, M.C.; Mastrofrancesco, L.; Milano, S.; Mastrodonato, M.; Carmosino, M.; Gerbino, A.; Corciulo, R.; Russo, R.; Svelto, M.; et al. AQP1-containing exosomes in peritoneal dialysis effluent as biomarker of dialysis efficiency. Cells 2019, 8, 330. [Google Scholar] [CrossRef] [Green Version]
- Piccapane, F.; Bonomini, M.; Castellano, G.; Gerbino, A.; Carmosino, M.; Svelto, M.; Arduini, A.; Procino, G. A novel formulation of glucose-sparing peritoneal dialysis solutions with L-carnitine improves biocompatibility on human mesothelial cells. Int. J. Mol. Sci. 2020, 22, 123. [Google Scholar] [CrossRef]
- Gerbino, A.; Fistetto, G.; Colella, M.; Hofer, A.M.; Debellis, L.; Caroppo, R.; Curci, S. Real time measurements of water flow in amphibian gastric glands: Modulation via the extracellular Ca2+-sensing receptor. J. Biol. Chem. 2007, 282, 13477–13486. [Google Scholar] [CrossRef] [Green Version]
- Brückner, B.R.; Nöding, H.; Skamrahl, M.; Janshoff, A. Mechanical and morphological response of confluent epithelial cell layers to reinforcement and dissolution of the F-actin cytoskeleton. Prog. Biophys. Mol. Biol. 2019, 144, 77–90. [Google Scholar] [CrossRef]
- Nie, H.G.; Tucker, T.; Su, X.F.; Na, T.; Peng, J.B.; Smith, P.R.; Idell, S.; Ji, H.L. Expression and regulation of epithelial Na+ channels by nucleotides in pleural mesothelial cells. Am. J. Respir. Cell Mol. Biol. 2009, 40, 543–554. [Google Scholar] [CrossRef]
- Ji, H.-L.; Nie, H.-G. Electrolyte and fluid transport in mesothelial cells. J. Epithel. Biol. Pharmacol. 2008, 1, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neher, E.; Lux, H.D. Rapid changes of potassium concentration at the outer surface of exposed single neurons during membrane current flow. J. Gen. Physiol. 1973, 61, 385–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson, C. Ion-selective microelectrodes and diffusion measurements as tools to explore the brain cell microenvironment. J. Neurosci. Methods 1993, 48, 199–213. [Google Scholar] [CrossRef]
- Stanfield, P.R. Tetraethylammonium ions and the potassium permeability of excitable cells. Rev. Physiol. Biochem. Pharmacol. 1983, 97, 1–67. [Google Scholar]
- Fejes-Toth, G.; Naray-Fejes-Toth, A. Differentiation of renal β-intercalated cells to α-intercalated and principal cells in culture. Proc. Natl. Acad. Sci. USA 1992, 89, 5487–5491. [Google Scholar] [CrossRef] [Green Version]
- Stoos, B.A.; Náray-Fejes-Tóth, A.; Carretero, O.A.; Ito, S.; Fejes-Tóth, G. Characterization of a mouse cortical collecting duct cell line. Kidney Int. 1991, 39, 1168–1175. [Google Scholar] [CrossRef] [Green Version]
- Mancinelli, R.; La Rovere, R.M.L.; Fulle, S.; Miscia, S.; Marchisio, M.; Pierdomenico, L.; Lanuti, P.; Procino, G.; Barbieri, C.; Svelto, M.; et al. Extracellular GTP is a potent water-transport regulator via aquaporin 5 plasma-membrane insertion in M1-CCD epithelial cortical collecting duct cells. Cell. Physiol. Biochem. 2014, 33, 731–746. [Google Scholar] [CrossRef]
- Preston, G.M.; Carroll, T.P.; Guggino, W.B.; Agre, P. Appearance of water channels in Xenopus oocytes expressing red cell CHIP28 protein. Science 1992, 256, 385–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devuyst, O.; Ni, J. Aquaporin-1 in the peritoneal membrane: Implications for water transport across capillaries and peritoneal dialysis. Biochim. Biophys. Acta Biomembr. 2006, 1758, 1078–1084. [Google Scholar] [CrossRef] [Green Version]
- Rippe, B.; Venturoli, D. Simulations of osmotic ultrafiltration failure in CAPD using a serial three-pore membrane/fiber matrix model. Am. J. Physiol.-Ren. Physiol. 2007, 292, F1035–F1043. [Google Scholar] [CrossRef] [Green Version]
- Breborowicz, A.; Knapowski, J. Transmesothelial ultrafiltration in vitro. Perit. Dial. Bull. 1986, 6, 124–127. [Google Scholar] [CrossRef]
- Stylianou, E.; Jenner, L.A.; Davies, M.; Coles, G.A.; Williams, J.D. Isolation, culture and characterization of human peritoneal mesothelial cells. Kidney Int. 1990, 37, 1563–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuse, M.; Hirase, T.; Itoh, M.; Nagafuchi, A.; Yonemura, S.; Tsukita, S.; Tsukita, S. Occludin: A novel integral membrane protein localizing at tight junctions. J. Cell Biol. 1993, 127, 1617–1626. [Google Scholar] [CrossRef] [Green Version]
- Hirase, T.; Staddon, J.M.; Saitou, M.; Ando-Akatsuka, Y.; Itoh, M.; Furuse, M.; Fujimoto, K.; Tsukita, S.; Rubin, L.L. Occludin as a possible determinant of tight junction permeability in endothelial cells. J. Cell Sci. 1997, 110, 1603–1613. [Google Scholar] [CrossRef]
- Ito, T.; Yorioka, N.; Kyuden, Y.; Asakimori, Y.; Kiribayashi, K.; Ogawa, T.; Kohno, N. Effect of glucose polymer on the intercellular junctions of cultured human peritoneal mesothelial cells. Nephron. Clin. Pract. 2003, 11, 1969–1979. [Google Scholar] [CrossRef]
- Horiuchi, T.; Matsunaga, K.; Banno, M.; Nakano, Y.; Nishimura, K.; Hanzawa, C.; Miyamoto, K.I.; Nomura, S.; Ohta, Y. HPMCs induce greater intercellular delocalization of tight junction-associated proteins due to a higher susceptibility to H2O2 compared with HUVECs. Perit. Dial. Int. 2009, 29, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Retana, C.; Sanchez, E.; Perez-Lopez, A.; Cruz, A.; Lagunas, J.; Cruz, C.; Vital, S.; Reyes, J.L. Alterations of intercellular junctions in peritoneal mesothelial cells from patients undergoing dialysis: Effect of retinoic acid. Perit. Dial. Int. 2015, 35, 275–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonetti, D.A.; Wolpert, E.B.; DeMaio, L.; Harhaj, N.S.; Scaduto, R.C. Hydrocortisone decreases retinal endothelial cell water and solute flux coincident with increased content and decreased phosphorylation of occludin. J. Neurochem. 2002, 80, 667–677. [Google Scholar] [CrossRef]
- Kaneda, K.I.; Miyamoto, K.; Nomura, S.; Horiuchi, T. Intercellular localization of occludins and ZO-1 as a solute transport barrier of the mesothelial monolayer. J. Artif. Organs 2006, 9, 241–250. [Google Scholar] [CrossRef]
- Witowski, J.; Breborowicz, A.; Topley, N.; Martis, L.; Knapowski, J.; Oreopoulos, D.G. Insulin stimulates the activity of Na+/N+-ATPase in human peritoneal mesothelial cells. Perit. Dial. Int. 1997, 17, 186–193. [Google Scholar] [CrossRef]
- Hatzoglou, C.H.; Gourgoulianis, K.I.; Molyvdas, P.A. Effects of SNP, ouabain, and amiloride on electrical potential profile of isolated sheep pleura. J. Appl. Physiol. 2001, 90, 1565–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agostoni, E.; Zocchi, L. Solute-coupled liquid absorption from the pleural space. Respir. Physiol. 1990, 81, 19–27. [Google Scholar] [CrossRef]
- Zocchi, L.; Agostoni, E.; Cremaschi, D. Electrolyte transport across the pleura of rabbits. Respir. Physiol. 1991, 86, 125–138. [Google Scholar] [CrossRef]
- Herrlich, A.; Leitch, V.; King, L.S. Role of proneuregulin 1 cleavage and human epidermal growth factor receptor activation in hypertonic aquaporin induction. Proc. Natl. Acad. Sci. USA 2004, 101, 15799–15804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenq, W.; Cooper, D.R.; Bittle, P.; Ramirez, G. Aquaporin-1 Expression in Proximal Tubule Epithelial Cells of Human Kidney Is Regulated by Hyperosmolarity and Contrast Agents. Biochem. Biophys. Res. Commun. 1999, 256, 240–248. [Google Scholar] [CrossRef]
- Umenishi, F.; Schrier, R.W. Identification and characterization of a novel hypertonicity-responsive element in the human aquaporin-1 gene. Biochem. Biophys. Res. Commun. 2002, 292, 771–775. [Google Scholar] [CrossRef]
- Umenishi, F.; Schrier, R.W. Hypertonicity-induced aquaporin-1 (AQP1) expression is mediated by the activation of MAPK pathways and hypertonicity-responsive element in the AQP1 gene. J. Biol. Chem. 2003, 278, 15765–15770. [Google Scholar] [CrossRef] [Green Version]
- Leypoldt, J.K.; Mistry, C.D. Ultrafiltration in peritoneal dialysis. In The Textbook of Peritoneal Dialysis; Springer: Dordrecht, The Netherlands, 1994. [Google Scholar]
- Morelle, J.; Sow, A.; Fustin, C.-A.; Fillée, C.; Garcia-Lopez, E.; Lindholm, B.; Goffin, E.; Vandemaele, F.; Rippe, B.; Öberg, C.M.; et al. Mechanisms of crystalloid versus colloid osmosis across the peritoneal membrane. J. Am. Soc. Nephrol. 2018, 29, 1875–1886. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Lou, Y.; Cui, W.; Zhang, Y.; Dong, W.; Sun, J.; Miao, L. Effect of aquaporin 1 on mouse peritoneal mesothelial cells after a long-term peritoneal dialysis. Ther. Apher. Dial. 2021, 25, 88–96. [Google Scholar] [CrossRef]
- Liu, S.M.; Li, J.; Wang, Y.; Ye, R.G.; Lindholm, B.; Wang, T. Methods to improve the preservation of peritoneal tissues. Adv. Perit. Dial. 2001, 17, 61–65. [Google Scholar]
- Kobayashi, H.; Yokoo, H.; Yanagita, T.; Satoh, S.; Kis, B.; Deli, M.; Niwa, M.; Wada, A. Induction of aquaporin 1 by dexamethasone in lipid rafts in immortalized brain microvascular endothelial cells. Brain Res. 2006, 1123, 12–19. [Google Scholar] [CrossRef]
- Guan, Y.; Chen, J.; Zhan, Y.; Lu, H. Effects of dexamethasone on C6 cell proliferation, migration and invasion through the upregulation of AQP1. Oncol. Lett. 2018, 15, 7595–7602. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Huang, B.; Wang, Y.; Tong, C.; Xie, P.; Fan, R.; Gao, Z. Emodin ameliorates acute lung injury induced by severe acute pancreatitis through the up-regulated expressions of AQP1 and AQP5 in lung. Clin. Exp. Pharmacol. Physiol. 2016, 43, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Wang, G.; Li, B.; Xiao, K.; Ma, Z.; Huang, H.; Wang, X.; Bai, C. Anti-asthmatic agents alleviate pulmonary edema by upregulating AQP1 and AQP5 expression in the lungs of mice with OVA-induced asthma. Respir. Physiol. Neurobiol. 2012, 181, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Amore, A.; Loiacono, E.; Balegno, S.; Manniello, D.; Peruzzi, L.; Camilla, R.; Minieri, V.; Daprà, V.; Qian, J.; et al. Effect of glucose degradation products, glucose-containing dialysate and icodextrin on AQP1 and eNOS expression in cultured endothelial cells. J. Nephrol. 2009, 22, 117–122. [Google Scholar]
- Bonomini, M.; Zammit, V.; Divino-Filho, J.C.; Davies, S.J.; Di Liberato, L.; Arduini, A.; Lambie, M. The osmo-metabolic approach: A novel and tantalizing glucose-sparing strategy in peritoneal dialysis. J. Nephrol. 2021, 34, 503–519. [Google Scholar] [CrossRef]
- Iolascon, A.; Aglio, V.; Tamma, G.; D’Apolito, M.; Addabbo, F.; Procino, G.; Simonetti, M.C.; Montini, G.; Gesualdo, L.; Debler, E.W.; et al. Characterization of two novel missense mutations in the AQP2 gene causing nephrogenic diabetes insipidus. Nephron Physiol. 2006, 105, p33–p41. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piccapane, F.; Gerbino, A.; Carmosino, M.; Milano, S.; Arduini, A.; Debellis, L.; Svelto, M.; Caroppo, R.; Procino, G. Aquaporin-1 Facilitates Transmesothelial Water Permeability: In Vitro and Ex Vivo Evidence and Possible Implications in Peritoneal Dialysis. Int. J. Mol. Sci. 2021, 22, 12535. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212535
Piccapane F, Gerbino A, Carmosino M, Milano S, Arduini A, Debellis L, Svelto M, Caroppo R, Procino G. Aquaporin-1 Facilitates Transmesothelial Water Permeability: In Vitro and Ex Vivo Evidence and Possible Implications in Peritoneal Dialysis. International Journal of Molecular Sciences. 2021; 22(22):12535. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212535
Chicago/Turabian StylePiccapane, Francesca, Andrea Gerbino, Monica Carmosino, Serena Milano, Arduino Arduini, Lucantonio Debellis, Maria Svelto, Rosa Caroppo, and Giuseppe Procino. 2021. "Aquaporin-1 Facilitates Transmesothelial Water Permeability: In Vitro and Ex Vivo Evidence and Possible Implications in Peritoneal Dialysis" International Journal of Molecular Sciences 22, no. 22: 12535. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212535