
Isoprocurcumenol Supports Keratinocyte Growth and Survival through Epidermal Growth Factor Receptor Activation

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
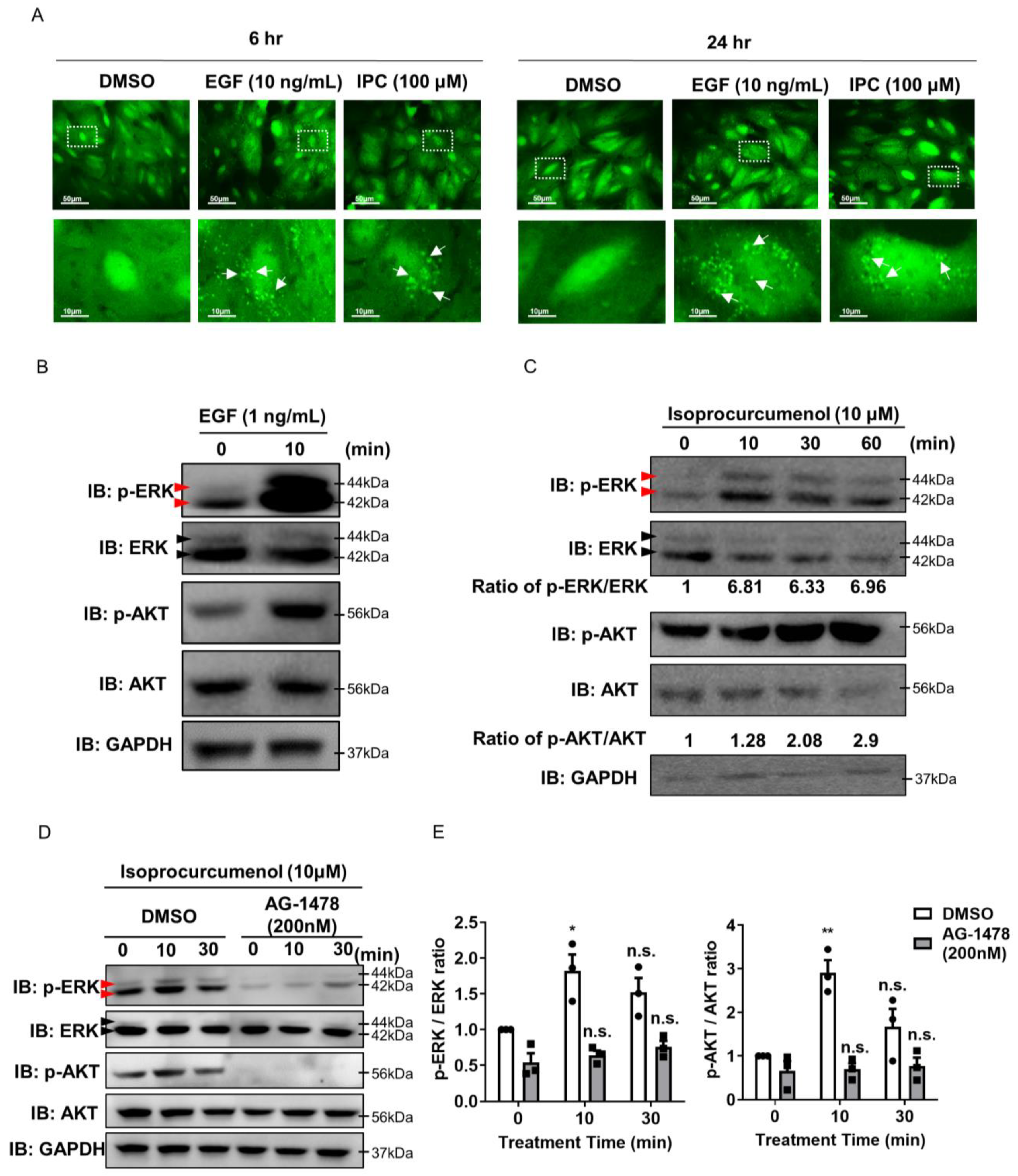
2.1. Recognizing Isoprocurcumenol as an EGF Analogue
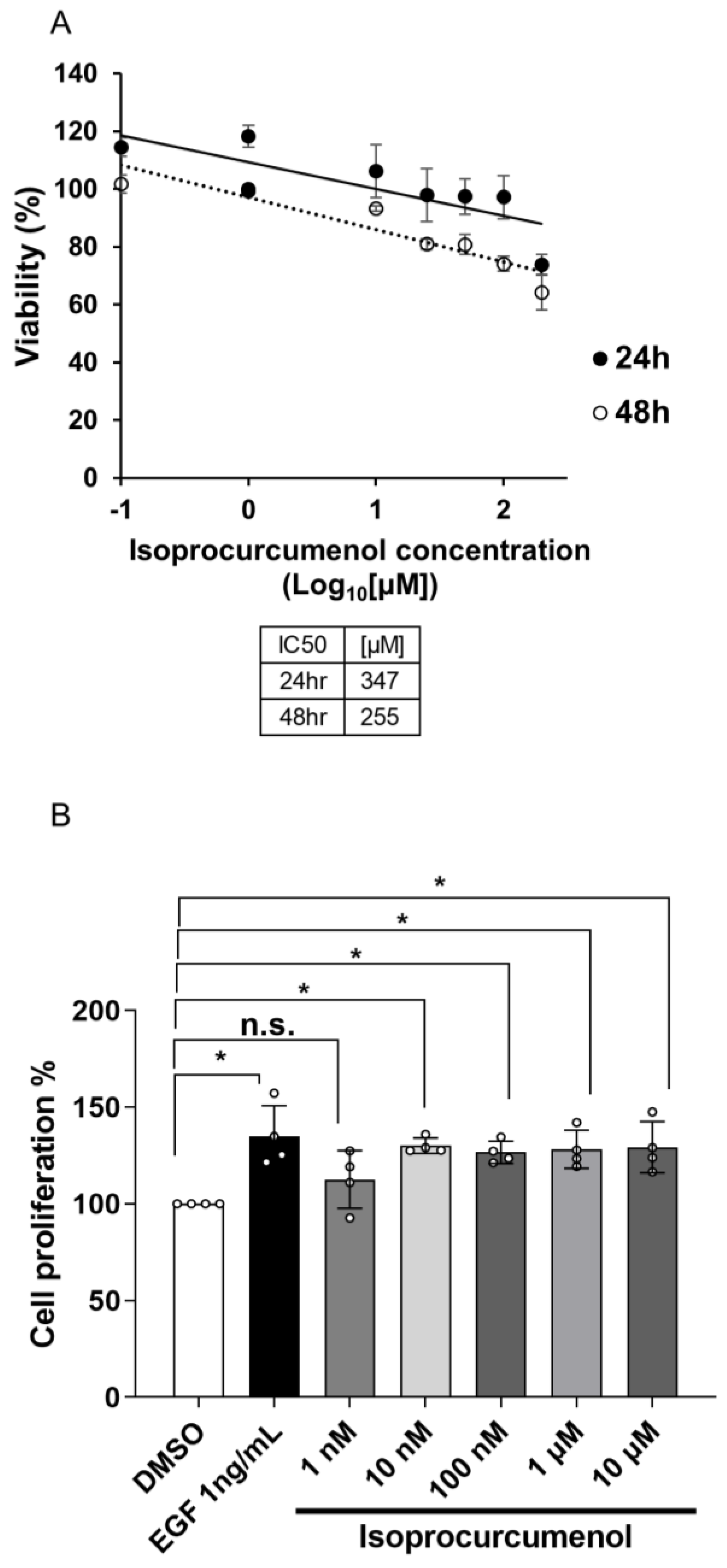
2.2. Measuring Appropriate Concentration of Isoprocurcumenol to Induce Cell Proliferation
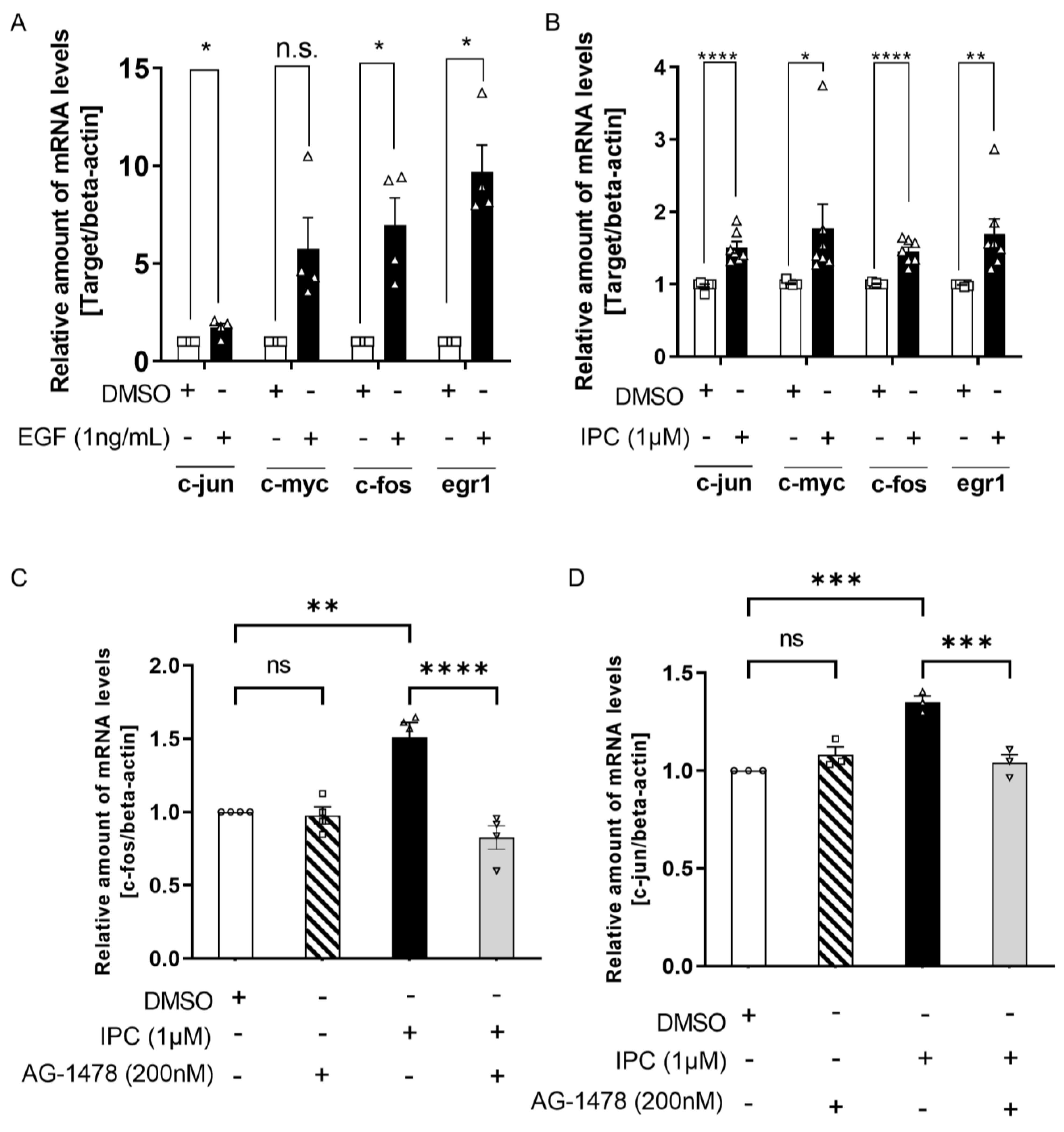
2.3. IPC Induces the Expression of Genes Related to Cell Growth and Proliferation
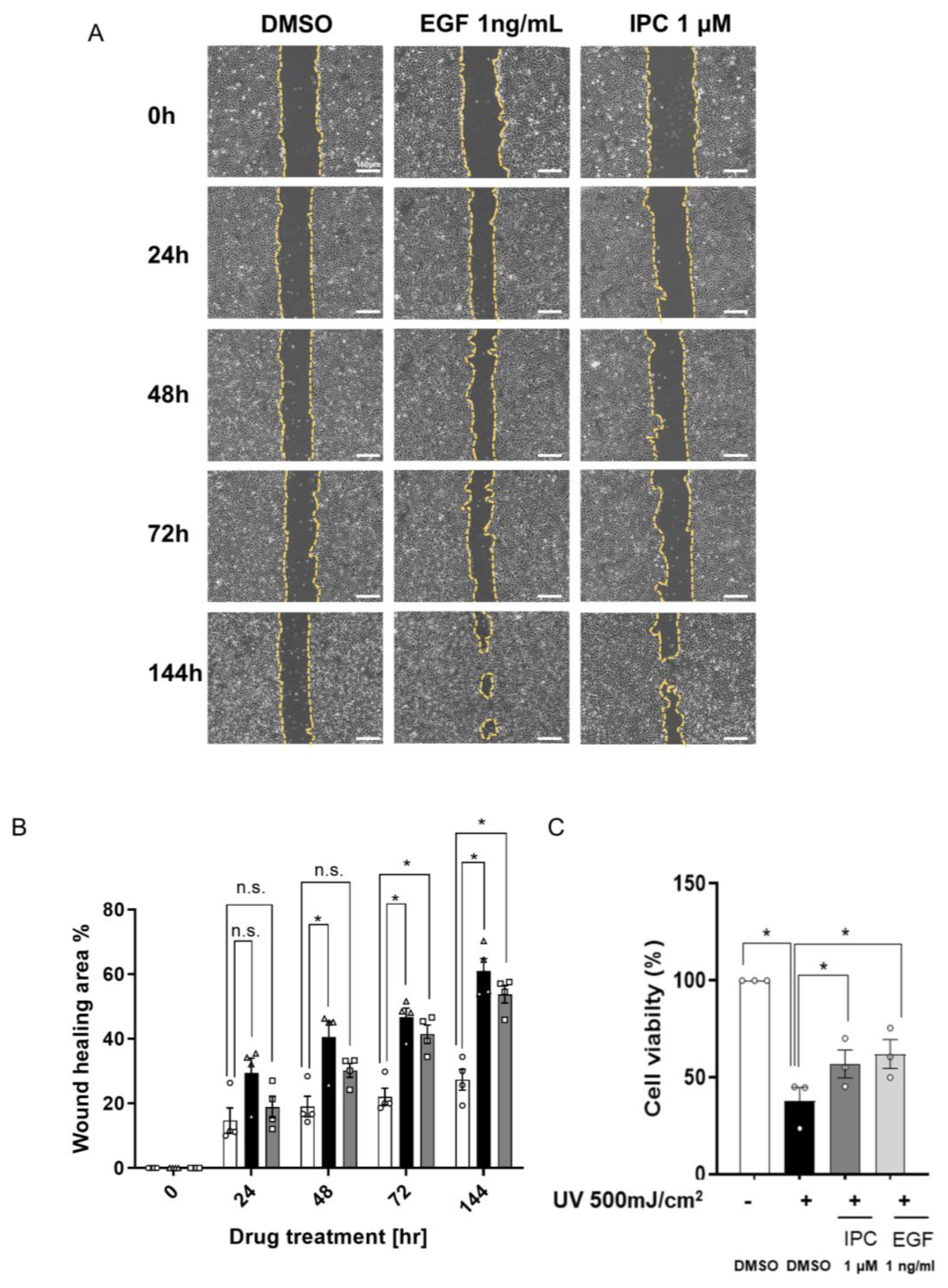
2.4. IPC Induces Cell Recovery and Wound Healing
3. Discussion
4. Material and Methods
4.1. Cell Culture and Compound Treatment
4.2. SDS-Polyacrylamide Gel Electrophoresis (PAGE) and Immunoblotting
4.3. Cell Growth Assay
4.4. Cell Viability (MTT) Assay
4.5. RNA Extraction and Quantitative PCR
4.6. Wound Healing Assay
4.7. UVB Irradiation
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Swann, G. The skin is the body’s largest organ. J. Vis. Commun. Med. 2010, 33, 148–149. [Google Scholar] [CrossRef] [PubMed]
- Roupe, G. Skin of the aging human being. Lakartidningen 2001, 98, 1091–1095. [Google Scholar] [PubMed]
- Soroka, Y.; Ma’or, Z.; Leshem, Y.; Verochovsky, L.; Neuman, R.; Brégégère, F.M.; Milner, Y. Aged keratinocyte phenotyping: Morphology, biochemical markers and effects of Dead Sea minerals. Exp. Gerontol. 2008, 43, 947–957. [Google Scholar] [CrossRef]
- Gunin, A.G.; Kornilova, N.K.; Petrov, V.V.; Vasil’eva, O.V. Age-related changes in the number and proliferation of fibroblasts in the human skin. Adv. Gerontol. 2011, 24, 43–47. [Google Scholar]
- Pierce, G.F.; Yanagihara, D.; Klopchin, K.; Danilenko, D.M.; Hsu, E.; Kenney, W.C.; Morris, C.F. Stimulation of all epithelial elements during skin regeneration by keratinocyte growth factor. J. Exp. Med. 1994, 179, 831–840. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Liu, F.; Wang, J.; Chen, X.; Xie, J.; Xiong, K. Epidermal stem cells in wound healing and their clinical applications. Stem Cell Res. Ther. 2019, 10, 229. [Google Scholar] [CrossRef] [Green Version]
- Nanba, D.; Toki, F.; Barrandon, Y.; Higashiyama, S. Recent advances in the epidermal growth factor receptor/ligand system biology on skin homeostasis and keratinocyte stem cell regulation. J. Dermatol. Sci. 2013, 72, 81–86. [Google Scholar] [CrossRef]
- Pastore, S.; Mascia, F.; Mariani, V.; Girolomoni, G. The Epidermal Growth Factor Receptor System in Skin Repair and Inflammation. J. Investig. Dermatol. 2008, 128, 1365–1374. [Google Scholar] [CrossRef] [Green Version]
- Oda, K.; Matsuoka, Y.; Funahashi, A.; Kitano, H. A comprehensive pathway map of epidermal growth factor receptor signaling. Mol. Syst. Biol. 2005, 1, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Gan, Y.; Shi, C.; Inge, L.; Hibner, M.; Balducci, J.; Huang, Y. Differential roles of ERK and Akt pathways in regulation of EGFR-mediated signaling and motility in prostate cancer cells. Oncogene 2010, 29, 4947–4958. [Google Scholar] [CrossRef] [Green Version]
- Grant, S.; Qiao, L.; Dent, P. Roles of ERBB family receptor tyrosine kinases, and downstream signaling pathways, in the control of cell growth and survival. Front. Biosci. 2002, 7, 376–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldag, C.; Nogueira Teixeira, D.; Leventhal, P.S. Skin rejuvenation using cosmetic products containing growth factors, cytokines, and matrikines: A review of the literature. Clin. Cosmet. Investig. Dermatol. 2019, 9, 411–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, J.J.; Eum, W.S.; Kwon, H.S.; Koh, J.S.; Lee, S.Y.; Baek, J.H.; Cho, Y.-J.; Kim, D.W.; Han, K.H.; Choi, S.Y.; et al. Protective effects of skin permeable epidermal and fibroblast growth factor against ultraviolet-induced skin damage and human skin wrinkles. J. Cosmet. Dermatol. 2013, 12, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Alemdaroğlu, C.; Degim, Z.; Celebi, N.; Şengezer, M.; Alömeroglu, M.; Nacar, A. Investigation of epidermal growth factor containing liposome formulation effects on burn wound healing. J. Biomed. Mater. Res. A 2008, 85, 271–283. [Google Scholar] [CrossRef]
- Pouranvari, S.; Ebrahimi, F.; Javadi, G.; Maddah, B. Cloning, Expression, and Cost Effective Purification of Authentic Human Epidermal Growth Factor With High Activity. Iran. Red. Crescent. Med. J. 2016, 18, e24966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razis AF, A.; Ismail, E.N.; Hambali, Z.; Abdullah MN, H.; Ali, A.M.; Lila MA, M. Expression of recombinant human epidermal growth factor in Escherichia coli and characterization of its biological activity. Appl. Biochem. Biotechnol. 2008, 144, 249–261. [Google Scholar] [CrossRef]
- Bos, J.D.; Meinardi, M.M. The 500 Dalton rule for the skin penetration of chemical compounds and drugs. Exp. Dermatol. 2000, 9, 165–169. [Google Scholar] [CrossRef]
- Srimal, R. Turmeric: A brief review of medicinal properties. Fitoterapia 1997, 68, 483–493. [Google Scholar]
- Haddad, M.; Sauvain, M.; Deharo, E. Curcuma as a parasiticidal agent: A review. Planta Med. 2011, 77, 672–678. [Google Scholar] [CrossRef] [Green Version]
- Honda, S.; Aoki, F.; Tanaka, H.; Kishida, H.; Nishiyama, T.; Okada, S.; Matsumoto, I.; Abe, K.; Mae, T. Effects of ingested turmeric oleoresin on glucose and lipid metabolisms in obese diabetic mice: A DNA microarray study. J. Agric. Food Chem. 2006, 54, 9055–9062. [Google Scholar] [CrossRef]
- Cole, G.M.; Teter, B.; Frautschy, S.A. Neuroprotective effects of curcumin. Adv. Exp. Med. Biol. 2007, 595, 197–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, B. Study of secondary metabolite constituents and curcumin contents of six different species of genus Curcuma. J. Med. Plants Stud. 2015, 3, 116–119. [Google Scholar]
- Lee, J.; Jung, Y.; Shin, J.H.; Kim, H.K.; Moon, B.C.; Ryu, D.H.; Hwang, G.S. Secondary metabolite profiling of Curcuma species grown at different locations using GC/TOF and UPLC/Q-TOF MS. Molecules 2014, 19, 9535–9551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.; Lim, J.; Woo, K.-C.; Kim, K.-T. Piperonylic acid stimulates keratinocyte growth and survival by activating epidermal growth factor receptor (EGFR). Sci. Rep. 2018, 8, 162. [Google Scholar] [CrossRef] [Green Version]
- Ichihashi, M.; Ueda, M.; Budiyanto, A.; Bito, T.; Oka, M.; Fukunaga, M.; Tsuru, K.; Horikawa, T. UV-induced skin damage. Toxicology 2003, 189, 21–39. [Google Scholar] [CrossRef]
- Kim, H.; Kong, W.H.; Seong, K.Y.; Sung, D.K.; Jeong, H.; Kim, J.K.; Yang, S.Y.; Hahn, S.K. Hyaluronate-Epidermal Growth Factor Conjugate for Skin Wound Healing and Regeneration. Biomacromolecules 2016, 17, 3694–3705. [Google Scholar] [CrossRef]
- He, X.-G.; Lin, L.-Z.; Lian, L.-Z.; Lindenmaier, M. Liquid chromatography–electrospray mass spectrometric analysis of curcuminoids and sesquiterpenoids in turmeric (Curcuma longa). J. Chromatogr. A 1998, 818, 127–132. [Google Scholar] [CrossRef]
- Ohshiro, M.; Kuroyanagi, M.; Ueno, A. Structures of sesquiterpenes from Curcuma longa. Phytochemistry 1990, 29, 2201–2205. [Google Scholar] [CrossRef]
- Normanno, N.; De Luca, A.; Bianco, C.; Strizzi, L.; Mancino, M.; Maiello, M.R.; Carotenuto, A.; Feo, G.D.; Caponigro, F.; Salomon, D.S. Epidermal growth factor receptor (EGFR) signaling in cancer. Gene 2006, 366, 2–16. [Google Scholar] [CrossRef]
- Tomas, A.; Futter, C.E.; Eden, E.R. EGF receptor trafficking: Consequences for signaling and cancer. Trends Cell Biol. 2014, 24, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, B.; Liebermann, D. Apoptotic signaling by c-MYC. Oncogene 2008, 27, 6462–6472. [Google Scholar] [CrossRef] [Green Version]
- Palmer, H.J.; Tuzon, C.T.; Paulson, K.E. Age-dependent Decline in Mitogenic Stimulation of Hepatocytes. J. Biol. Chem. 1999, 274, 11424–11430. [Google Scholar] [CrossRef] [Green Version]
- Kong, H.J.; Mooney, D.J. Microenvironmental regulation of biomacromolecular therapies. Nat. Rev. Drug. Discov. 2007, 6, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Hamdi OA, A.; Ye, L.J.; Kamarudin MN, A.; Hazni, H.; Paydar, M.; Looi, C.Y.; Shilpi, J.A.; Kadir, H.A.; Awang, K. Neuroprotective and Antioxidant Constituents from Curcuma zedoaria Rhizomes. Rec. Nat. Prod. 2015, 9, 349–355. [Google Scholar]
- Choi, J.H.; Kim, S.H.; Jeong, Y.H.; Kim, S.W.; Min, K.T.; Kim, K.T. hnRNP Q Regulates Internal Ribosome Entry Site-Mediated fmr1 Translation in Neurons. Mol. Cell. Biol. 2019, 39, e00371-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, P.K.; Lee, K.H.; Kim, J.H.; Tae, S.; Ham, S.; Jeong, Y.H.; Kim, S.W.; Kang, B.; Kim, H.M.; Kim, K.T.; et al. hnRNP K Supports High-Amplitude D Site-Binding Protein mRNA (Dbp mRNA) Oscillation To Sustain Circadian Rhythms. Mol. Cell. Biol. 2020, 40, e00537-19. [Google Scholar] [CrossRef]
- Kwon, P.K.; Kim, H.M.; Kang, B.; Kim, S.W.; Hwang, S.M.; Im, S.H.; Roh, T.-Y.; Kim, K.T. hnRNP K supports the maintenance of RORγ circadian rhythm through ERK signaling. FASEB J. 2021, 35, e21507. [Google Scholar] [CrossRef]
- Kwon, P.K.; Kim, H.M.; Kim, S.W.; Kang, B.; Yi, H.; Ku, H.O.; Roh, T.-Y.; Kim, K.T. The Poly(C) Motif in the Proximal Promoter Region of the D Site-Binding Protein Gene (Dbp) Drives Its High-Amplitude Oscillation. Mol. Cell. Biol. 2019, 39, e00101-19. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.H.; Kim, S.H.; Kim, H.J.; Kim, W.; Lee, H.R.; Jung, Y.; Choi, J.-H.; Hong, K.Y.; Jang, S.K.; Kim, K.T.; et al. AUF1 contributes to Cryptochrome1 mRNA degradation and rhythmic translation. Nucleic Acids Res. 2014, 42, 3590–3606. [Google Scholar] [CrossRef]
- Lee, K.H.; Woo, K.C.; Kim, D.Y.; Kim, T.D.; Shin, J.; Park, S.M.; Jang, S.K.; Kim, K.T. Rhythmic interaction between Period1 mRNA and hnRNP Q leads to circadian time-dependent translation. Mol. Cell. Biol. 2012, 32, 717–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunes, J.P.S.; Dias, A.A.M. ImageJ macros for the user-friendly analysis of soft-agar and wound-healing assays. Biotechniques 2018, 62, 175–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, P.K.; Kim, S.W.; De, R.; Jeong, S.W.; Kim, K.-T. Isoprocurcumenol Supports Keratinocyte Growth and Survival through Epidermal Growth Factor Receptor Activation. Int. J. Mol. Sci. 2021, 22, 12579. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212579
Kwon PK, Kim SW, De R, Jeong SW, Kim K-T. Isoprocurcumenol Supports Keratinocyte Growth and Survival through Epidermal Growth Factor Receptor Activation. International Journal of Molecular Sciences. 2021; 22(22):12579. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212579
Chicago/Turabian StyleKwon, Paul Kwangho, Sung Wook Kim, Ranjit De, Sung Woo Jeong, and Kyong-Tai Kim. 2021. "Isoprocurcumenol Supports Keratinocyte Growth and Survival through Epidermal Growth Factor Receptor Activation" International Journal of Molecular Sciences 22, no. 22: 12579. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212579