
Progressive Aggregation of 16 kDa Gamma-Zein during Seed Maturation in Transgenic Arabidopsis thaliana

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. 16γzf Accumulates in Arabidopsis Embryos without Interfering with the Synthesis and Processing of Endogenous Storage Proteins
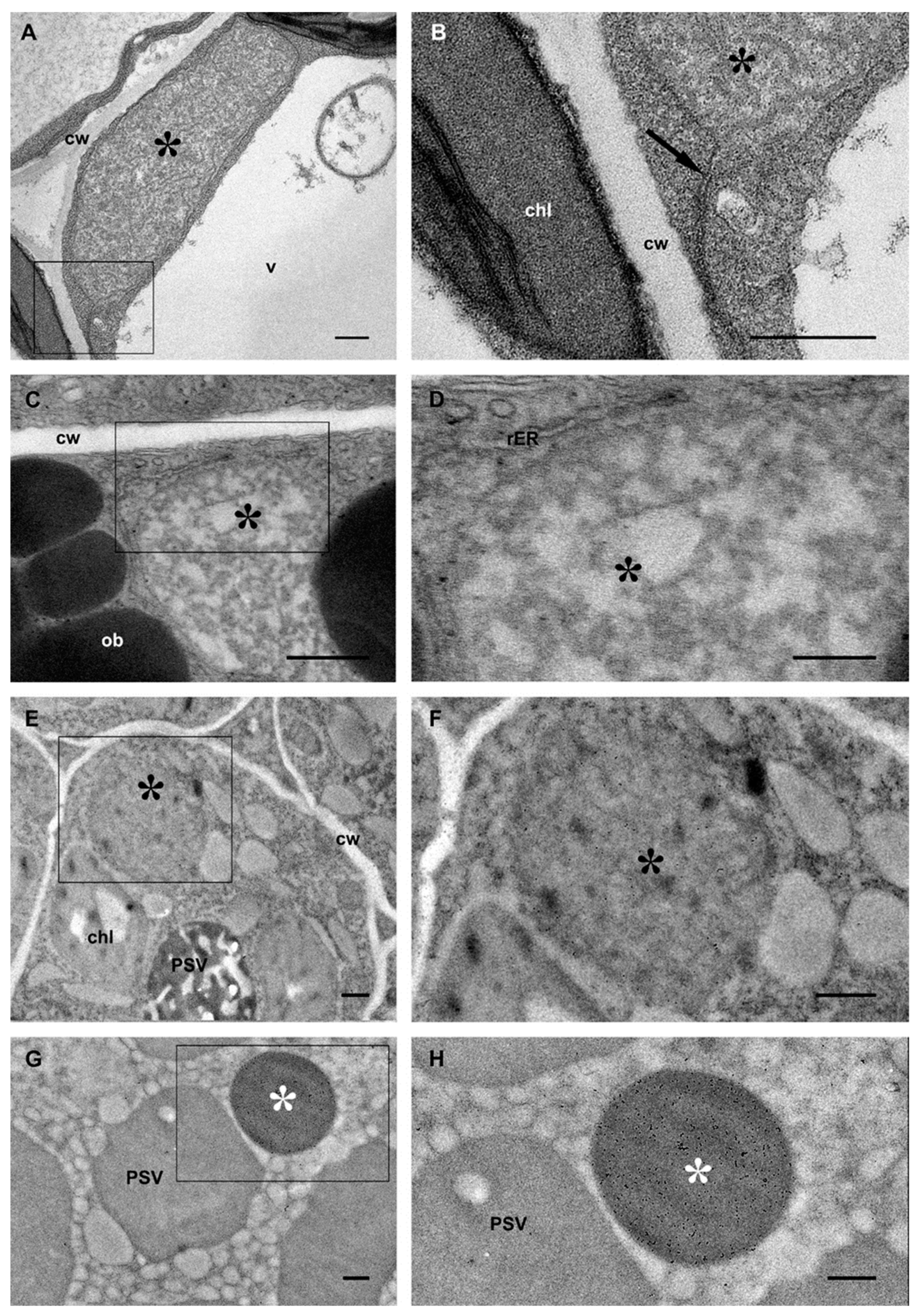
2.2. 16γzf Avoids Traffic to PSVs and Forms Electron-Dense Threads That Progressively Coalesce during Embryo Development
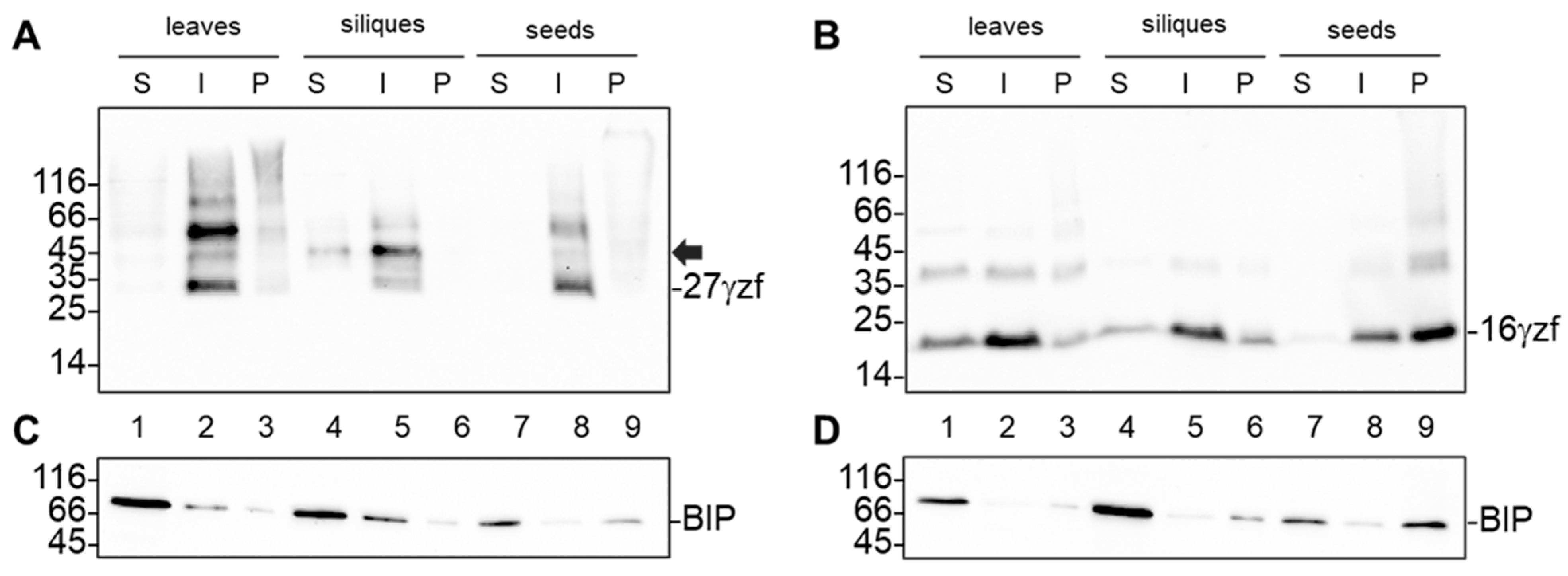
2.3. 16γzf Forms Insoluble Aggregates in Mature Embryos, Unlike 27γzf
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Antibodies
4.3. Total Protein Extraction from Leaves, Roots, and Dry Seeds
4.4. Protein Body Precipitation
4.5. Protein Sequential Extraction with Different Buffer and Centrifugation
4.6. Electron Microscopy
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hinz, G.; Hoh, B.; Hohl, I.; Robinson, D.G. Stratification of storage proteins in the protein storage vacuole of developing cotyledons of Pisum sativum L. J. Plant Physiol. 1995, 145, 437–442. [Google Scholar] [CrossRef]
- Lending, C.R.; Larkins, B.A. Changes in the zein composition of protein bodies during maize endosperm development. Plant Cell 1989, 1, 1011–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedrazzini, E.; Giovinazzo, G.; Bielli, A.; de Virgilio, M.; Frigerio, L.; Pesca, M.; Faoro, F.; Bollini, R.; Ceriotti, A.; Vitale, A. Protein quality control along the route to the plant vacuole. Plant Cell 1997, 9, 1869–1880. [Google Scholar] [PubMed] [Green Version]
- Chrispeels, M.J. The Golgi apparatus mediates the transport of phytohemagglutinin to the protein bodies in bean cotyledons. Planta 1983, 158, 140–151. [Google Scholar] [CrossRef]
- Castelli, S.; Vitale, A. The phaseolin vacuolar sorting signal promotes transient, strong membrane association and aggregation of the bean storage protein in transgenic tobacco. J. Exp. Bot. 2005, 56, 1379–1387. [Google Scholar] [CrossRef] [PubMed]
- Hillmer, S.; Movafeghi, A.; Robinson, D.G.; Hinz, G. Vacuolar storage proteins are sorted in the cis-cisternae of the pea cotyledon Golgi apparatus. J. Cell Biol. 2001, 152, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Hara-Nishimura, I.; Shimada, T.; Hatano, K.; Takeuchi, Y.; Nishimura, M. Transport of storage proteins to protein storage vacuoles is mediated by large precursor-accumulating vesicles. Plant Cell 1998, 10, 825–836. [Google Scholar] [CrossRef] [Green Version]
- Shewry, P.R.; Napier, J.A.; Tatham, A.S. Seed storage proteins: Structures and biosynthesis. Plant Cell 1995, 7, 945–956. [Google Scholar] [CrossRef] [Green Version]
- Pedrazzini, E.; Mainieri, D.; Marrano, C.; Vitale, A. Where do protein bodies of cereal seeds come from? Front. Plant Sci. 2016, 7, 1139. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.U.; Sheteiwy, M.; Lihua, N.; Khan, M.M.U.; Han, Z. An update on the maize zein-gene family in the post-genomics era. Food Prod. Process. Nutr. 2019, 1, 13. [Google Scholar] [CrossRef] [Green Version]
- Larkins, B.A. Proteins of the Kernel. In Corn: Chemistry and Technology, 3rd ed.; Elsevier Science B.V.: Amsterdam, The Netherlands, 2019; pp. 319–336. [Google Scholar] [CrossRef]
- Yao, D.; Qi, W.; Li, X.; Yang, Q.; Yan, S.; Ling, H.; Wang, G.; Wang, G.; Song, R. Maize opaque10 encodes a cereal-specific protein that is essential for the proper distribution of zeins in endosperm protein bodies. PloS Genet. 2016, 12, e1006270. [Google Scholar] [CrossRef] [PubMed]
- Geli, M.I.; Torrent, M.; Ludevid, D. Two structural domains mediate two sequential events in gamma-zein targeting: Protein endoplasmic reticulum retention and protein body formation. Plant Cell 1994, 6, 1911–1922. [Google Scholar] [CrossRef] [PubMed]
- Mainieri, D.; Marrano, C.; Prinsi, B.; Maffi, D.; Tschofen, M.; Espen, L.; Stoger, E.; Faoro, F.; Pedrazzini, E.; Vitale, A. Maize 16-kD gamma-zein forms very unusual disulfide-bonded polymers in the endoplasmic reticulum: Implications for prolamin evolution. J. Exp. Bot. 2018, 69, 5013–5027. [Google Scholar] [CrossRef]
- Mainieri, D.; Morandini, F.; Maitrejean, M.; Saccani, A.; Pedrazzini, E.; Vitale, A. Protein body formation in the endoplasmic reticulum as an evolution of storage protein sorting to vacuoles: Insights from maize gamma-zein. Front. Plant Sci. 2014, 5, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.-H.; Messing, J. Organization of the prolamin gene family provides insight into the evolution of the maize genome and gene duplications in grass species. Proc. Natl. Acad. Sci. USA 2008, 105, 14330–14335. [Google Scholar] [CrossRef] [Green Version]
- Prat, S.; Cortadas, J.; Puigdomenech, P.; Palau, J. Nucleic acid (cDNA) and amino acid sequences of the maize endosperm protein glutelin-2. Nucleic Acids Res. 1985, 13, 1493–1504. [Google Scholar] [CrossRef]
- Vitale, A.; Smaniotto, E.; Longhi, R.; Galante, E. Reduced soluble proteins associated with maize endosperm protein bodies. J. Exp. Bot. 1982, 33, 439–448. [Google Scholar] [CrossRef]
- Shimada, T.; Fuji, K.; Tamura, K.; Kondo, M.; Nishimura, M.; Hara-Nishimura, I. Vacuolar sorting receptor for seed storage proteins in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2003, 100, 16095–16100. [Google Scholar] [CrossRef] [Green Version]
- Pobre, K.F.R.; Poet, G.J.; Hendershot, L.M. The endoplasmic reticulum (ER) chaperone BiP is a master regulator of ER functions: Getting by with a little help from ERdj friends. J. Biol. Chem. 2019, 294, 2098–2108. [Google Scholar] [CrossRef] [Green Version]
- Vitale, A.; Boston, R. Endoplasmic reticulum quality control and the unfolded protein response: Insights from plants. Traffic 2008, 9, 1581–1588. [Google Scholar] [CrossRef]
- Brocca, L.; Zuccaro, M.; Frugis, G.; Mainieri, D.; Marrano, C.; Ragni, L.; Klein, E.; Vitale, A.; Pedrazzini, E. Two γ-zeins induce the unfolded protein response. Plant Physiol. 2021, 187, 1428–1444. [Google Scholar] [CrossRef] [PubMed]
- Fukasawa, T.; Haranishimura, I.; Nishimura, M. Biosynthesis, intracellular transport and in vitro processing of 11S globulin precursor proteins of developing castor bean endosperm. Plant Cell Physiol. 1988, 29, 339–345. [Google Scholar]
- Kinney, A.J.; Jung, R.; Herman, E.M. Cosuppression of the alpha subunits of beta-conglycinin in transgenic soybean seeds induces the formation of endoplasmic reticulum-derived protein bodies. Plant Cell 2001, 13, 1165–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molvig, L.; Tabe, L.M.; Eggum, B.O.; Moore, A.E.; Craig, S.; Spencer, D.; Higgins, T.J.V. Enhanced methionine levels and increased nutritive value of seeds of transgenic lupins (Lupinus angustifolius L.) expressing a sunflower seed albumin gene. Proc. Natl. Acad. Sci. USA 1997, 94, 8393–8398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wandelt, C.I.; Khan, M.R.I.; Craig, S.; Schroeder, H.E.; Spencer, D.; Higgins, T.J.V. Vicilin with carboxy-terminal KDEL is retained in the endoplasmic reticulum and accumulates to high levels in the leaves of transgenic plants. Plant J. 1992, 2, 181–192. [Google Scholar] [PubMed] [Green Version]
- Arcalis, E.; Stadlmann, J.; Marcel, S.; Drakakaki, G.; Winter, V.; Rodriguez, J.; Fischer, R.; Altmann, F.; Stoger, E. The changing fate of a secretory glycoprotein in developing maize endosperm. Plant Physiol. 2010, 153, 693–702. [Google Scholar] [CrossRef] [Green Version]
- Hundertmark, M.; Hincha, D.K. LEA (Late Embryogenesis Abundant) proteins and their encoding genes in Arabidopsis thaliana. BMC Genomics 2008, 9, 118. [Google Scholar] [CrossRef] [Green Version]
- Waters, E.R.; Vierling, E. Plant small heat shock proteins-evolutionary and functional diversity. New Phytol. 2020, 227, 24–37. [Google Scholar] [CrossRef] [Green Version]
- Leprince, O.; Pellizzaro, A.; Berriri, S.; Buitink, J. Late seed maturation: Drying without dying. J. Exp. Bot. 2017, 68, 827–841. [Google Scholar] [CrossRef] [Green Version]
- Furuki, T.; Shimizu, T.; Chakrabortee, S.; Yamakawa, K.; Hatanaka, R.; Takahashi, T.; Kikawada, T.; Okuda, T.; Mihara, H.; Tunnacliffe, A.; et al. Effects of Group 3 LEA protein model peptides on desiccation-induced protein aggregation. BBA-Proteins Proteom. 2012, 1824, 891–897. [Google Scholar] [CrossRef]
- Feeney, M.; Kittelmann, M.; Menassa, R.; Hawes, C.; Frigerio, L. Protein Storage Vacuoles Originate from Remodeled Preexisting Vacuoles in Arabidopsis thaliana. Plant Physiol. 2018, 177, 241–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arcalis, E.; Mainieri, D.; Vitale, A.; Stöger, E.; Pedrazzini, E. Progressive Aggregation of 16 kDa Gamma-Zein during Seed Maturation in Transgenic Arabidopsis thaliana. Int. J. Mol. Sci. 2021, 22, 12671. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222312671
Arcalis E, Mainieri D, Vitale A, Stöger E, Pedrazzini E. Progressive Aggregation of 16 kDa Gamma-Zein during Seed Maturation in Transgenic Arabidopsis thaliana. International Journal of Molecular Sciences. 2021; 22(23):12671. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222312671
Chicago/Turabian StyleArcalis, Elsa, Davide Mainieri, Alessandro Vitale, Eva Stöger, and Emanuela Pedrazzini. 2021. "Progressive Aggregation of 16 kDa Gamma-Zein during Seed Maturation in Transgenic Arabidopsis thaliana" International Journal of Molecular Sciences 22, no. 23: 12671. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222312671