
Risk Mitigation of Immunogenicity: A Key to Personalized Retinal Gene Therapy

Abstract
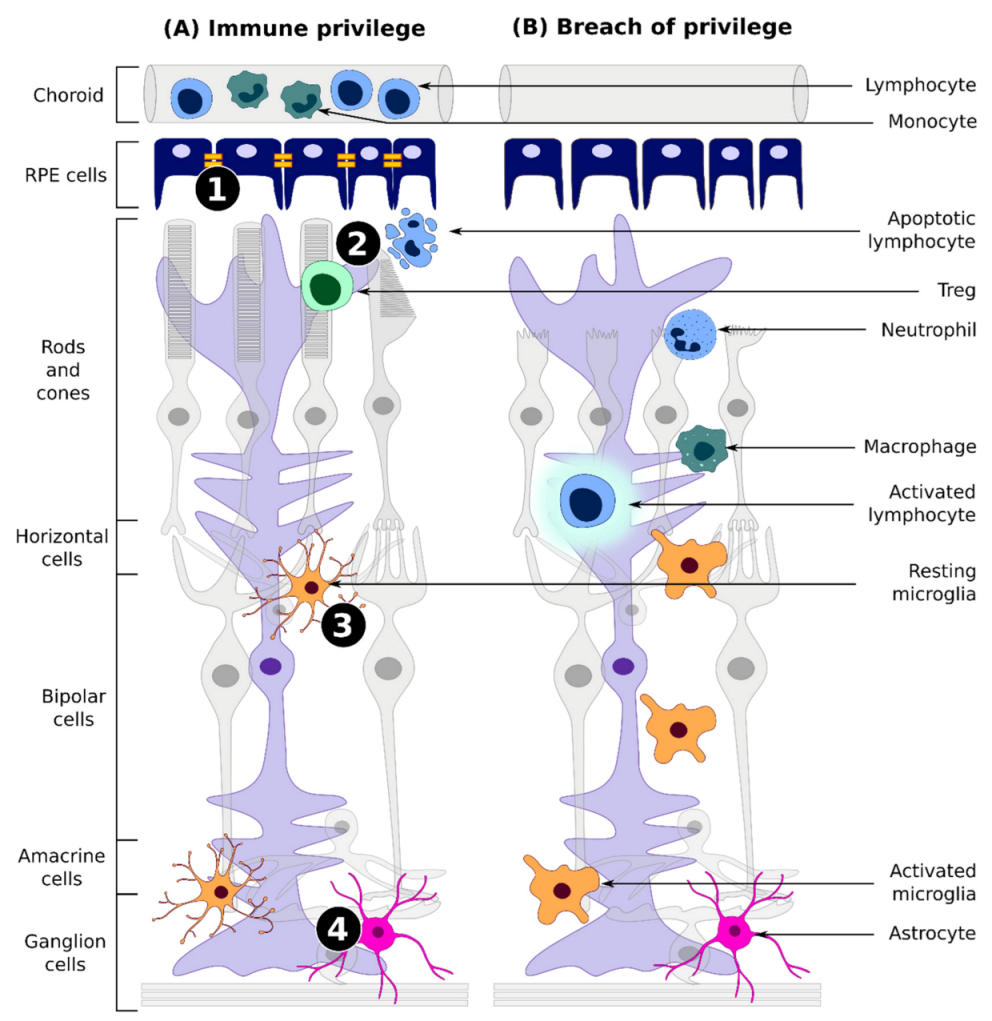
:1. Introduction

2. Immune Responses in Retinal Gene Therapy Trials

{kind=link}
{kind=link}
{kind=link}
| NCT Number | Sponsor | Study Type (1) | Vector | Immune Response | References | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Serotype | Promoter | Transgene | Dose (vg) | Injection (2) | Inflammation | Other | ||||
| Leber congenital amaurosis | ||||||||||
| NCT00516477 | Spark Therapeutics | PI, NR, OL | 2 | CBA | hRPE65v2 | 1.5 × 1010 to 1.5 × 1011 | SRI | no inflammation | transient increase in nAB in 2/3 patients | Maguire et al. 2009 [38] Simonelli et al. 2010 [39] |
| NCT01208389 | PI/II, NR, OL (follow-on study: injection of the contralateral eye) | 1.5 × 1011 | SRI | no inflammation | transient minor increase in anti-capsid Abs 1/10 no cell-mediated T cell responses detectable in peripheral blood | Bennett et al. 2012 & 2016 [40,41] | ||||
| NCT00999609 | PIII, R, OL | 1.5 × 1011 | SRI | 2/20 with transient mild bilateral inflammation in treated arm | N/A | Russell et al. 2017 [42] | ||||
| NCT00643747 | University College, London | PI/II, NR, OL | 2 | hRPE65p | hRPE65 | 1 × 1012 | SRI | in 5/8 with high dose; anterior uveitis in 1/8 | increased level of AAV2 NAbs and marginal increased circulating T cells with reactivity to AAV2 in 1 high-dose patient transiently increased circulating neutralizing antibodies to AAV2 in another high-dose patient | Bainbridge et al. 2015 [37] |
| NCT01496040 | Nantes University Hospital | PI/II, NR, OL | 2 | hRPE65p | hRPE65 | 1.22 × 1010 to 4.8 × 1010 | SRI | transient infra-clinical inflammation at D+4 in three patients more significant transient inflammation was observed in two patients | anti-AAV4 IgG antibodies in three patients (1 before + 2 after injection) weak and transient cellular response to RPE65 between | Le Meur et al. 2018 [43] |
| NCT02781480 | MeiraGTx UK II Ltd. | PI/II, NR, OL | 5 | hRPE65popt | hRPE65opt | 1 × 1011 to 1 × 1012 | SRI | mild uveitis in 3/9 low, 1/3 intermediate, and 1/3 high dose serious uveitis in 2/3 intermediate dose and 1/3 high dose | N/A | clinicaltrials.gov 24 November 2021 |
| NCT00481546 | University of Pennsylvania | PI, NR, OL | 2 | CBSB | hRPE65 | 5.96 × 1010 to 1.79 × 1011 | SRI | N/A | episodic humoral immune response in 4/15; modest increase in T-cell response by cultured ELISpot in 3/15 | Jacobson et al. 2012 [44] |
| NCT00749957 | Applied Genetic Technologies Corp | PI/II, NR, OL | 2 | CBSB | hRPE65 | 1.8 × 1011 to 6 × 1011 | SRI | in 3 patients, eye inflammation at highest dose | Titers of neutralizing antibodies to AAV increased in 5 of 12 patients | Weleber et al. 2016 [45] |
| NCT00821340 | Hadassah Medical Organization | PI | 2 | N/A | hRPE65 | N/A | SRI | N/A | N/A | |
| Choroideremia | ||||||||||
| NCT02341807 | Spark Therapeutics | PI/II, NR, OL | 2 | CAG | hCHM | 5 × 1010 to 1 × 1011 | SRI | N/A | N/A | Morgan et al. 2021 [46] |
| NCT01461213 | University of Oxford | PI/II, NR, OL | 2 | CAG | hCHM + WPRE | 0.6 × 1010 to 1 × 1011 | SRI | vector-related inflammation and vitritis in 1/14 at 2wpi; | N/A | MacLaren et al. 2014 [47] |
| NCT02077361 | Ian M. MacDonald | PI/II, NR, OL | 1 × 1011 | SRI | 1/6 severe intraretinal inflammation, leading to permanent structural and functional impairment of the retina | N/A | Dimopoulos et al. 2018 [48] | |||
| NCT02553135 | Byron Lam | PII, NR, OL | 1 × 1011 | SRI | 1/6 vitreous cells | significantly increased serum anti-AAV-2 neutralizing antibody after treatment 1/6 | Lam et al. 2019 [49] | |||
| NCT02671539 | STZ eye trial | PII, R, OL | 2 | CAG | hCHM + WPRE | 1 × 1011 | SRI | N/A | N/A | Fischer et al. 2019 [50] |
| NCT02407678 | University of Oxford | PII, R, OL | 1 × 1011 | SRI | N/A | N/A | ||||
| NCT03507686 | NightstaRx Ltd. | PII, NR, OL | N/A | SRI | N/A | N/A | ||||
| NCT03496012 | NightstaRx Ltd. | PIII, R, M | N/A | SRI | N/A | N/A | ||||
| X-linked retinitis pigmentosa | ||||||||||
| NCT03116113 | NightstaRx Ltd. | PI/II, R, M | 8 | hRK | coRPGR | 2 × 109 to 4 × 1011 | SRI | subretinal inflammation at high doses | N/A | Cehajic-Kapetanovic et al. 2020 [36] |
| NCT03252847 | MeiraGTx UK II Ltd. | PI/II, R, OL | 5 | hRK | RPGR | SRI | Inflammatory responses were observed in 2 out of the 3 patients in the high-dose cohort | N/A | Michaelides et al. 2020 [51] | |
| Retinitis pigmentosa | ||||||||||
| NCT04919473 | Nanoscope Therapeutics Inc. | PI/iIa, NR, OL | 2 | CMVp + mGLUR6 enhancer | MCO | 1.75 × 1011 to 3.5 × 1011 | IVT | N/A | N/A | clinicaltrials.gov 24 November 2021 |
| Age-related macular degeneration | ||||||||||
| NCT01494805 | Lions Eye Institute, Perth | PI/II (dose-escalation), R, M | 2 | CBA | sFLT1 | 1 × 1011 | SRI | 2 anterior chamber inflammation and 1 eye inflammation; two ocular aEs were considered possibly related to rAAV. sFLT-1 was eye inflammation, and anterior chamber inflammation, which was mild in nature and resolved without sequelae | IFNϒ T-cells against capsid in 1/21; 3 seroconverted; | Constable et al. 2016 [52] |
| NCT03066258 | Regenxbio Inc. | PI/iIa (dose-escalation), NR, OL | 8 | N/A | soluble anti-VEGF (monoclonal antibody fragment) | 3 × 109 to 2.5 × 1011 | SRI | post-operative inflammation in 36% of subjects resolved within days to weeks | N/A | Allen Ho et al. 2021 [53] (presentation) |
| NCT01024998 | Genzyme | PI (dose-escalation), NR, OL | 2 | CBA | sFLT1 | 2 × 108 to 2 × 1010 | IVT | 2/3 intraocular inflammation resolved with topical steroid | 62% of patients injected with 6 × 109 or 2 × 1010 had increase in anti-AAV2 antibodies | Heier et al. 2017 [54] |
| NCT03748784 | Adverum Biotechnologies, Inc. | PI, NR, OL | 2.7m8 | CMV | aflibercept | 2 × 1011 to 6 × 1011 | IVT | Ocular inflammation minimal and responsive to steroids in low-dose cohort | N/A | Adverum press release 01 October 2021 [55] |
| NCT03585556 | Janssen Research & Development, LLC | PI, NR, OL | 2 | CAG | sCD59 | 3.56 × 1011 to 1.071 × 1012 | IVT | N/A | N/A | |
| NCT03144999 | Janssen Research & Development, LLC | PI, NR, OL | 2 | CAG | sCD59 | IVT | N/A | N/A | ||
| Diabetic macular edema | ||||||||||
| NCT04418427 | Adverum Biotechnologies, Inc. | PII, R, OL | 2.7m8 | CMV | aflibercept | 2 × 1011 to 6 × 1011 | IVT | 5/12 increase ocular pressure and 1/12 loss of vision at high dose | N/A | Adverum press release 22 July 2021 [56] |
| X-linked retinoschisis | ||||||||||
| NCT02416622 | Applied Genetic Technologies Corp | PI/II, NR, OL | 2tYF | CBSB | hRS1 | 1 × 1011 to 6 × 1011 | IVT | 10/21 anterior chamber cells for intermediate and high doses | N/A | clinicaltrials.gov 24 November 2021 |
| Leber hereditary optic neuropathy | ||||||||||
| NCT02161380 | Byron Lam | PI | scAAV2 | CAG | ND4 subunit gene of complex I with targeting sequence of P1 isoform of subunit c of ATP synthase | 5 × 109 to 2.46 × 1010 | IVT | 1/14 mild uveitis medium dose and 1/14 uveitis low dose after 2 months, resolved spontaneously | 1/14 strong increase in NAbs | Guy et al. 2017 [57] |
3. The Immunological Environment of the Retina
3.1. Immune Privilege
3.2. Microglial-Based Surveillance and Its Impact on AAV Gene Therapy
3.3. DNA Sensors
3.4. The TLR9/MyD88 Pathway Is Activated upon rAAV Transduction
4. Factors Influencing Inflammatory Reactions in Response to rAAV
4.1. CpG Content
4.2. Combination of Viral and Bacterial Sequences
4.3. Route of Administration
4.4. Inflammatory Components of Retinal Diseases
5. Options to Reduce Immunological Risk
5.1. At the AAV Level
5.1.1. Modified Capsid
5.1.2. Evading DNA Sensing
5.1.3. Decreased Dose
5.2. At the Patient Level
5.2.1. Immunosuppressive Strategies
5.2.2. Clinical Trial Criteria
6. Conclusions and Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kuzmin, D.A.; Shutova, M.V.; Johnston, N.R.; Smith, O.P.; Fedorin, V.V.; Kukushkin, Y.S.; van der Loo, J.C.M.; Johnstone, E.C. The Clinical Landscape for AAV Gene Therapies. Nat. Rev. Drug Discov. 2021, 20, 173–174. [Google Scholar] [CrossRef] [PubMed]
- Astellas Provides Update on ASPIRO Clinical Trial of AT132 in Patients with X-Linked Myotubular Myopathy|Astellas Pharma Inc. GLOBAL WEBSITE. Available online: https://www.astellas.com/en/news/17121 (accessed on 25 November 2021).
- Solid Biosciences Announces Clinical Hold on SGT-001 Phase I/II Clinical Trial For Duchenne Muscular Dystrophy. Available online: https://www.solidbio.com/about/media/press-releases/solid-biosciences-announces-clinical-hold-on-sgt-001-phase-i-ii-clinical-trial-for-duchenne-muscular-dystrophy (accessed on 30 October 2021).
- Konkle, B.A.; Walsh, C.; Escobar, M.A.; Josephson, N.C.; Young, G.; von Drygalski, A.; McPhee, S.W.J.; Samulski, R.J.; Bilic, I.; De La Rosa, M.; et al. BAX 335 Hemophilia B Gene Therapy Clinical Trial Results—Potential Impact of CpG Sequences on Gene Expression. Blood 2021, 137, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Reichel, F.F.; Dauletbekov, D.L.; Klein, R.; Peters, T.; Ochakovski, G.A.; Seitz, I.P.; Wilhelm, B.; Ueffing, M.; Biel, M.; Wissinger, B.; et al. AAV8 Can Induce Innate and Adaptive Immune Response in the Primate Eye. Mol. Ther. 2017, 25, 2648–2660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, W.; Earley, L.F.; Chai, Z.; Chen, X.; Sun, J.; He, T.; Deng, M.; Hirsch, M.L.; Ting, J.; Samulski, R.J.; et al. Double-Stranded RNA Innate Immune Response Activation from Long-Term Adeno-Associated Virus Vector Transduction. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Noma, H.; Mimura, T.; Yasuda, K.; Shimura, M. Role of Inflammation in Diabetic Macular Edema. OPH 2014, 232, 127–135. [Google Scholar] [CrossRef]
- Stein-Streilein, J. Immune Regulation and the Eye. Trends Immunol. 2008, 29, 548–554. [Google Scholar] [CrossRef]
- Zamiri, P.; Masli, S.; Kitaichi, N.; Taylor, A.W.; Streilein, J.W. Thrombospondin Plays a Vital Role in the Immune Privilege of the Eye. Investig. Ophthal. Vis. Sci. 2005, 46, 908–919. [Google Scholar] [CrossRef] [Green Version]
- Griffith, T.S.; Yu, X.; Herndon, J.M.; Green, D.R.; Ferguson, T.A. CD95-Induced Apoptosis of Lymphocytes in an Immune Privileged Site Induces Immunological Tolerance. Immunity 1996, 5, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Hori, J.; Wang, M.; Miyashita, M.; Tanemoto, K.; Takahashi, H.; Takemori, T.; Okumura, K.; Yagita, H.; Azuma, M. B7-H1-Induced Apoptosis as a Mechanism of Immune Privilege of Corneal Allografts. J. Immunol. 2006, 177, 5928. [Google Scholar] [CrossRef] [Green Version]
- Chandler, L.; Yusuf, I.; McClements, M.; Barnard, A.; MacLaren, R.; Xue, K. Immunomodulatory Effects of Hydroxychloroquine and Chloroquine in Viral Infections and Their Potential Application in Retinal Gene Therapy. Int. J. Mol. Sci. 2020, 21, 4972. [Google Scholar] [CrossRef]
- Chan, Y.K.; Wang, S.K.; Chu, C.J.; Copland, D.A.; Letizia, A.J.; Costa Verdera, H.; Chiang, J.J.; Sethi, M.; Wang, M.K.; Neidermyer, W.J.; et al. Engineering Adeno-Associated Viral Vectors to Evade Innate Immune and Inflammatory Responses. Sci. Transl. Med. 2021, 13, eabd3438. [Google Scholar] [CrossRef]
- Forrester, J.; Xu, H. Good News–Bad News: The Yin and Yang of Immune Privilege in the Eye. Front. Immunol. 2012, 3, 338. [Google Scholar] [CrossRef] [Green Version]
- Hori, J.; Vega, J.L.; Masli, S. Review of Ocular Immune Privilege in the Year 2010: Modifying the Immune Privilege of the Eye. Ocular Immunol. Inflamm. 2010, 18, 325–333. [Google Scholar] [CrossRef]
- Chen, M.; Luo, C.; Zhao, J.; Devarajan, G.; Xu, H. Immune Regulation in the Aging Retina. Prog. Retin. Eye Res. 2019, 69, 159–172. [Google Scholar] [CrossRef] [Green Version]
- Griffith, T.S.; Brunner, T.; Fletcher, S.M.; Green, D.R.; Ferguson, T.A. Fas Ligand-Induced Apoptosis as a Mechanism of Immune Privilege. Science 1995, 270, 1189–1192. [Google Scholar] [CrossRef]
- Taylor, A.W.; Ng, T.F. Negative Regulators That Mediate Ocular Immune Privilege. J. Leukoc. Biol. 2018, 103, 1179–1187. [Google Scholar] [CrossRef]
- Detrick, B.; Hooks, J.J. Immune Regulation in the Retina. Immunol Res. 2010, 47, 153–161. [Google Scholar] [CrossRef]
- Fu, S.; Dong, S.; Zhu, M.; Sherry, D.M.; Wang, C.; You, Z.; Haigh, J.J.; Le, Y.-Z. Müller Glia Are a Major Cellular Source of Survival Signals for Retinal Neurons in Diabetes. Diabetes 2015, 64, 3554–3563. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Ma, W.; Zhao, L.; Fariss, R.N.; Wong, W.T. Adaptive Müller Cell Responses to Microglial Activation Mediate Neuroprotection and Coordinate Inflammation in the Retina. J. Neuroinflamm. 2011, 8, 173. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.W.; Hsu, S.; Ng, T.F. The Role of Retinal Pigment Epithelial Cells in Regulation of Macrophages/Microglial Cells in Retinal Immunobiology. Front. Immunol. 2021, 12, 3256. [Google Scholar] [CrossRef]
- McMenamin, P.G.; Saban, D.R.; Dando, S.J. Immune Cells in the Retina and Choroid: Two Different Tissue Environments That Require Different Defenses and Surveillance. Prog. Retin. Eye Res. 2019, 70, 85–98. [Google Scholar] [CrossRef]
- Kettenmann, H.; Hanisch, U.-K.; Noda, M.; Verkhratsky, A. Physiology of Microglia. Physiol. Rev. 2011, 91, 461–553. [Google Scholar] [CrossRef]
- Detrick, B.; Hooks, J.J. The RPE Cell and the Immune System. In Retinal Pigment Epithelium in Health and Disease; Klettner, A.K., Dithmar, S., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 101–114. ISBN 978-3-030-28384-1. [Google Scholar]
- Kumar, A.; Pandey, R.K.; Miller, L.J.; Singh, P.K.; Kanwar, M. Müller Glia in Retinal Innate Immunity: A Perspective on Their Roles in Endophthalmitis. Crit. Rev. Immunol. 2013, 33, 119–135. [Google Scholar] [CrossRef]
- Medzhitov, R. Origin and Physiological Roles of Inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Xu, H.; Chen, M.; Forrester, J.V. Para-Inflammation in the Aging Retina. Prog. Retin. Eye Res. 2009, 28, 348–368. [Google Scholar] [CrossRef]
- Chen, M.; Xu, H. Parainflammation, Chronic Inflammation and Age-Related Macular Degeneration. J. Leukoc. Biol. 2015, 98, 713–725. [Google Scholar] [CrossRef] [Green Version]
- Murakami, Y.; Ishikawa, K.; Nakao, S.; Sonoda, K.-H. Innate Immune Response in Retinal Homeostasis and Inflammatory Disorders. Prog. Retin. Eye Res. 2020, 74, 100778. [Google Scholar] [CrossRef]
- Ronning, K.E.; Karlen, S.J.; Miller, E.B.; Burns, M.E. Molecular Profiling of Resident and Infiltrating Mononuclear Phagocytes during Rapid Adult Retinal Degeneration Using Single-Cell RNA Sequencing. Sci. Rep. 2019, 9, 4858. [Google Scholar] [CrossRef] [Green Version]
- Garafalo, A.V.; Cideciyan, A.V.; Héon, E.; Sheplock, R.; Pearson, A.; WeiYang Yu, C.; Sumaroka, A.; Aguirre, G.D.; Jacobson, S.G. Progress in Treating Inherited Retinal Diseases: Early Subretinal Gene Therapy Clinical Trials and Candidates for Future Initiatives. Prog. Retin. Eye Res. 2020, 77, 100827. [Google Scholar] [CrossRef]
- Trapani, I.; Auricchio, A. Seeing the Light after 25 Years of Retinal Gene Therapy. Trends Mol. Med. 2018, 24, 669–681. [Google Scholar] [CrossRef]
- Pennesi, M.E.; Schlecther, C.L. The Evolution of Retinal Gene Therapy: From Clinical Trials to Clinical Practice. Available online: https://pubmed.ncbi.nlm.nih.gov/31973830/ (accessed on 19 May 2020).
- Seitz, I.P.; Michalakis, S.; Wilhelm, B.; Reichel, F.F.; Ochakovski, G.A.; Zrenner, E.; Ueffing, M.; Biel, M.; Wissinger, B.; Bartz-Schmidt, K.U.; et al. Superior Retinal Gene Transfer and Biodistribution Profile of Subretinal Versus Intravitreal Delivery of AAV8 in Nonhuman Primates. Investig. Ophthalmol. Vis. Sci. 2017, 58, 5792–5801. [Google Scholar] [CrossRef] [PubMed]
- Cehajic-Kapetanovic, J.; Xue, K.; de la Camara, C.M.-F.; Nanda, A.; Davies, A.; Wood, L.J.; Salvetti, A.P.; Fischer, M.D.; Aylward, J.W.; Barnard, A.R.; et al. Retinal Gene Therapy in X-Linked Retinitis Pigmentosa Caused by Mutations in RPGR: Results at 6 Months in a First in Human Clinical Trial. Nat. Med. 2020, 26, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Bainbridge, J.W.B.; Mehat, M.S.; Sundaram, V.; Robbie, S.J.; Barker, S.E.; Ripamonti, C.; Georgiadis, A.; Mowat, F.M.; Beattie, S.G.; Gardner, P.J.; et al. Long-Term Effect of Gene Therapy on Leber’s Congenital Amaurosis. N. Engl. J. Med. 2015, 372, 1887–1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maguire, A.M.; High, K.A.; Auricchio, A.; Wright, J.F.; Pierce, E.A.; Testa, F.; Mingozzi, F.; Bennicelli, J.L.; Ying, G.; Rossi, S.; et al. Age-Dependent Effects of RPE65 Gene Therapy for Leber’s Congenital Amaurosis: A Phase 1 Dose-Escalation Trial. Lancet 2009, 374, 1597–1605. [Google Scholar] [CrossRef] [Green Version]
- Simonelli, F.; Maguire, A.M.; Testa, F.; Pierce, E.A.; Mingozzi, F.; Bennicelli, J.L.; Rossi, S.; Marshall, K.; Banfi, S.; Surace, E.M.; et al. Gene Therapy for Leber’s Congenital Amaurosis Is Safe and Effective through 1.5 Years after Vector Administration. Mol. Ther. 2010, 18, 643–650. [Google Scholar] [CrossRef]
- Bennett, J.; Ashtari, M.; Wellman, J.; Marshall, K.A.; Cyckowski, L.L.; Chung, D.C.; McCague, S.; Pierce, E.A.; Chen, Y.; Bennicelli, J.L.; et al. AAV2 Gene Therapy Readministration in Three Adults with Congenital Blindness. Sci. Transl. Med. 2012, 4, 120ra15. [Google Scholar] [CrossRef] [Green Version]
- Bennett, J.; Wellman, J.; Marshall, K.A.; McCague, S.; Ashtari, M.; DiStefano-Pappas, J.; Elci, O.U.; Chung, D.C.; Sun, J.; Wright, J.F.; et al. Safety and Durability of Effect of Contralateral-Eye Administration of AAV2 Gene Therapy in Patients with Childhood-Onset Blindness Caused by RPE65 Mutations: A Follow-on Phase 1 Trial. Lancet 2016, 388, 661–672. [Google Scholar] [CrossRef] [Green Version]
- Russell, S.; Bennett, J.; Wellman, J.A.; Chung, D.C.; Yu, Z.-F.; Tillman, A.; Wittes, J.; Pappas, J.; Elci, O.; McCague, S.; et al. Efficacy and Safety of Voretigene Neparvovec (AAV2-HRPE65v2) in Patients with RPE65-Mediated Inherited Retinal Dystrophy: A Randomised, Controlled, Open-Label, Phase 3 Trial. Lancet 2017, 390, 849–860. [Google Scholar] [CrossRef]
- Le Meur, G.; Lebranchu, P.; Billaud, F.; Adjali, O.; Schmitt, S.; Bézieau, S.; Péréon, Y.; Valabregue, R.; Ivan, C.; Darmon, C.; et al. Safety and Long-Term Efficacy of AAV4 Gene Therapy in Patients with RPE65 Leber Congenital Amaurosis. Mol. Ther. 2018, 26, 256–268. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, S.G.; Cideciyan, A.V.; Ratnakaram, R.; Heon, E.; Schwartz, S.B.; Roman, A.J.; Peden, M.C.; Aleman, T.S.; Boye, S.L.; Sumaroka, A.; et al. Gene Therapy for Leber Congenital Amaurosis Caused by RPE65 Mutations: Safety and Efficacy in 15 Children and Adults Followed up to 3 Years. Arch. Ophthalmol. 2012, 130, 9–24. [Google Scholar] [CrossRef] [Green Version]
- Weleber, R.G.; Pennesi, M.E.; Wilson, D.J.; Kaushal, S.; Erker, L.R.; Jensen, L.; McBride, M.T.; Flotte, T.R.; Humphries, M.; Calcedo, R.; et al. Results at 2 Years after Gene Therapy for RPE65-Deficient Leber Congenital Amaurosis and Severe Early-Childhood–Onset Retinal Dystrophy. Ophthalmology 2016, 123, 1606–1620. [Google Scholar] [CrossRef] [Green Version]
- Morgan, J.I.W.; Jiang, Y.Y.; Vergilio, G.K.; Serrano, L.W.; Pearson, D.J.; Bennett, J.; Maguire, A.M.; Aleman, T.S. Short-Term Assessment of Subfoveal Injection of AAV2-HCHM Gene Augmentation in Choroideremia Using Adaptive Optics Ophthalmoscopy. bioRxiv 2021. [Google Scholar] [CrossRef]
- MacLaren, R.E.; Groppe, M.; Barnard, A.R.; Cottriall, C.L.; Tolmachova, T.; Seymour, L.; Clark, K.R.; During, M.J.; Cremers, F.P.M.; Black, G.C.M.; et al. Retinal Gene Therapy in Patients with Choroideremia: Initial Findings from a Phase 1/2 Clinical Trial. Lancet 2014, 383, 1129–1137. [Google Scholar] [CrossRef] [Green Version]
- Dimopoulos, I.S.; Hoang, S.C.; Radziwon, A.; Binczyk, N.M.; Seabra, M.C.; MacLaren, R.E.; Somani, R.; Tennant, M.T.S.; MacDonald, I.M. Two-Year Results After AAV2-Mediated Gene Therapy for Choroideremia: The Alberta Experience. Am. J. Ophthalmol. 2018, 193, 130–142. [Google Scholar] [CrossRef]
- Lam, B.L.; Davis, J.L.; Gregori, N.Z.; MacLaren, R.E.; Girach, A.; Verriotto, J.D.; Rodriguez, B.; Rosa, P.R.; Zhang, X.; Feuer, W.J. Choroideremia Gene Therapy Phase 2 Clinical Trial: 24-Month Results. Am. J. Ophthalmol. 2019, 197, 65–73. [Google Scholar] [CrossRef]
- Fischer, M.D.; Ochakovski, G.A.; Beier, B.; Seitz, I.P.; Vaheb, Y.; Kortuem, C.; Reichel, F.F.L.; Kuehlewein, L.; Kahle, N.A.; Peters, T.; et al. Efficacy and Safety of Retinal Gene Therapy Using Adeno-Associated Virus Vector for Patients With Choroideremia: A Randomized Clinical Trial. JAMA Ophthalmol. 2019, 137, 1247–1254. [Google Scholar] [CrossRef]
- Michaelides, M.; Besirli, C.; Khan, K.; Yang, Y.; Wong, C.; Sahel, J.; Shah, M.; Tee, J.; Kumaran, N.; Georgiadis, A.; et al. AAV5-RPGR Gene Therapy for RPGR-Associated X-Linked Retinitis Pigmentosa: 12-Month Results From a Phase 1/2 Clinical Trial. In Proceedings of the AAO Virtual Meeting, Virtual, 13–15 November 2020. [Google Scholar]
- Constable, I.J.; Pierce, C.M.; Lai, C.-M.; Magno, A.L.; Degli-Esposti, M.A.; French, M.A.; McAllister, I.L.; Butler, S.; Barone, S.B.; Schwartz, S.D.; et al. Phase 2a Randomized Clinical Trial: Safety and Post Hoc Analysis of Subretinal RAAV.SFLT-1 for Wet Age-Related Macular Degeneration. EBioMedicine 2016, 14, 168–175. [Google Scholar] [CrossRef] [Green Version]
- REGENXBIO Announces Additional Positive Long-Term and Interim Phase I/IIa Trial Update for RGX-314 for the Treatment of Wet AMD|REGENXBIO Inc. Available online: http://ir.regenxbio.com/news-releases/news-release-details/regenxbio-announces-additional-positive-long-term-and-interim/ (accessed on 30 October 2021).
- Heier, J.S.; Kherani, S.; Desai, S.; Dugel, P.; Kaushal, S.; Cheng, S.H.; Delacono, C.; Purvis, A.; Richards, S.; Le-Halpere, A.; et al. Intravitreous Injection of AAV2-SFLT01 in Patients with Advanced Neovascular Age-Related Macular Degeneration: A Phase 1, Open-Label Trial. Lancet 2017, 390, 50–61. [Google Scholar] [CrossRef]
- Adverum Presents 2-Year OPTIC Data Demonstrating Sustained Durability and Promising Safety Profile from Single Intravitreal Injection of ADVM-022 in Wet AMD Patients Who Previously Required Frequent Anti-VEGF Injections. Available online: https://investors.adverum.com/news/news-details/2021/Adverum-Presents-2-Year-OPTIC-Data-Demonstrating-Sustained-Durability-and-Promising-Safety-Profile-from-Single-Intravitreal-Injection-of-ADVM-022-in-Wet-AMD-Patients-Who-Previously-Required-Frequent-Anti-VEGF-Injections/default.aspx (accessed on 30 October 2021).
- Adverum Provides Update on ADVM-022 and the INFINITY Trial in Patients with Diabetic Macular Edema. Available online: https://investors.adverum.com/news/news-details/2021/Adverum-Provides-Update-on-ADVM-022-and-the-INFINITY-Trial-in-Patients-with-Diabetic-Macular-Edema/default.aspx (accessed on 30 October 2021).
- Guy, J.; Feuer, W.J.; Davis, J.L.; Porciatti, V.; Gonzalez, P.J.; Koilkonda, R.D.; Yuan, H.; Hauswirth, W.W.; Lam, B.L. Gene Therapy for Leber Hereditary Optic Neuropathy: Low- and Medium-Dose Visual Results. Ophthalmology 2017, 124, 1621–1634. [Google Scholar] [CrossRef]
- Forrester, J.V.; McMenamin, P.G.; Dando, S.J. CNS Infection and Immune Privilege. Nat. Rev. Neurosci. 2018, 19, 655–671. [Google Scholar] [CrossRef]
- Qian, Y.-W.; Li, C.; Jiang, A.-P.; Ge, S.; Gu, P.; Fan, X.; Li, T.-S.; Jin, X.; Wang, J.-H.; Wang, Z.-L. HIV-1 Gp120 Glycoprotein Interacting with Dendritic Cell-Specific Intercellular Adhesion Molecule 3-Grabbing Non-Integrin (DC-SIGN) Down-Regulates Tight Junction Proteins to Disrupt the Blood Retinal Barrier and Increase Its Permeability. J. Biol. Chem. 2016, 291, 22977–22987. [Google Scholar] [CrossRef] [Green Version]
- McKimmie, C.S.; Roy, D.; Forster, T.; Fazakerley, J.K. Innate Immune Response Gene Expression Profiles of N9 Microglia Are Pathogen-Type Specific. J. Neuroimmunol. 2006, 175, 128–141. [Google Scholar] [CrossRef]
- Lyons, A.; Lynch, A.M.; Downer, E.J.; Hanley, R.; O’Sullivan, J.B.; Smith, A.; Lynch, M.A. Fractalkine-Induced Activation of the Phosphatidylinositol-3 Kinase Pathway Attentuates Microglial Activation in Vivo and in Vitro. J. Neurochem. 2009, 110, 1547–1556. [Google Scholar] [CrossRef]
- Harrison, J.K.; Jiang, Y.; Chen, S.; Xia, Y.; Maciejewski, D.; McNamara, R.K.; Streit, W.J.; Salafranca, M.N.; Adhikari, S.; Thompson, D.A.; et al. Role for Neuronally Derived Fractalkine in Mediating Interactions between Neurons and CX3CR1-Expressing Microglia. Proc. Natl. Acad. Sci. USA 1998, 95, 10896–10901. [Google Scholar] [CrossRef] [Green Version]
- Beguier, F.; Housset, M.; Roubeix, C.; Augustin, S.; Zagar, Y.; Nous, C.; Mathis, T.; Eandi, C.; Benchaboune, M.; Drame-Maigné, A.; et al. The 10q26 Risk Haplotype of Age-Related Macular Degeneration Aggravates Subretinal Inflammation by Impairing Monocyte Elimination. Immunity 2020, 53, 429–441.e8. [Google Scholar] [CrossRef]
- Liu, B.; Wei, L.; Meyerle, C.; Tuo, J.; Sen, H.N.; Li, Z.; Chakrabarty, S.; Agron, E.; Chan, C.-C.; Klein, M.L.; et al. Complement Component C5a Promotes Expression of IL-22 and IL-17 from Human T Cells and Its Implication in Age-Related Macular Degeneration. J. Transl. Med. 2011, 9, 111. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.; Liu, B.; Tuo, J.; Shen, D.; Chen, P.; Li, Z.; Liu, X.; Ni, J.; Dagur, P.; Sen, H.N.; et al. Hypomethylation of the IL17RC Promoter Associates with Age-Related Macular Degeneration. Cell Rep. 2012, 2, 1151–1158. [Google Scholar] [CrossRef] [Green Version]
- DiSabato, D.J.; Quan, N.; Godbout, J.P. Neuroinflammation: The Devil Is in the Details. J. Neurochem. 2016, 139, 136–153. [Google Scholar] [CrossRef] [Green Version]
- Verkhratsky, A.; Noda, M.; Parpura, V. Microglia: Structure and Function. In Brain Mapping; Elsevier: Amsterdam, The Netherlands, 2015; pp. 109–113. ISBN 978-0-12-397316-0. [Google Scholar]
- Cherry, J.D.; Olschowka, J.A.; O’Banion, M. Neuroinflammation and M2 Microglia: The Good, the Bad, and the Inflamed. J. Neuroinflamm. 2014, 11, 98. [Google Scholar] [CrossRef] [Green Version]
- Jha, M.K.; Jo, M.; Kim, J.-H.; Suk, K. Microglia-Astrocyte Crosstalk: An Intimate Molecular Conversation. Neuroscientist 2019, 25, 227–240. [Google Scholar] [CrossRef]
- Ransohoff, R.M. A Polarizing Question: Do M1 and M2 Microglia Exist? Nat. Neurosci. 2016, 19, 987–991. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Roubeix, C.; Sennlaub, F.; Saban, D.R. Perspective: Microglia and Monocyte Distinct Roles in Degenerative Diseases of the Retina. Trends Neurosci. 2020, 43, 433–449. [Google Scholar] [CrossRef] [PubMed]
- Mathys, H.; Adaikkan, C.; Gao, F.; Young, J.Z.; Manet, E.; Hemberg, M.; Jager, P.L.D.; Ransohoff, R.M.; Regev, A.; Tsai, L.-H. Temporal Tracking of Microglia Activation in Neurodegeneration at Single-Cell Resolution. Cell Rep. 2017, 21, 366–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammond, T.R.; Dufort, C.; Dissing-Olesen, L.; Giera, S.; Young, A.; Wysoker, A.; Walker, A.J.; Gergits, F.; Segel, M.; Nemesh, J.; et al. Single-Cell RNA Sequencing of Microglia throughout the Mouse Lifespan and in the Injured Brain Reveals Complex Cell-State Changes. Immunity 2019, 50, 253–271.e6. [Google Scholar] [CrossRef] [Green Version]
- Young, A.M.H.; Kumasaka, N.; Calvert, F.; Hammond, T.R.; Knights, A.; Panousis, N.; Park, J.S.; Schwartzentruber, J.; Liu, J.; Kundu, K.; et al. A Map of Transcriptional Heterogeneity and Regulatory Variation in Human Microglia. Nat. Genet. 2021, 53, 861–868. [Google Scholar] [CrossRef]
- Smith, J.A.; Das, A.; Ray, S.K.; Banik, N.L. Role of Pro-Inflammatory Cytokines Released from Microglia in Neurodegenerative Diseases. Brain Res. Bull. 2012, 87, 10–20. [Google Scholar] [CrossRef]
- Sierra, A.; Abiega, O.; Shahraz, A.; Neumann, H. Janus-Faced Microglia: Beneficial and Detrimental Consequences of Microglial Phagocytosis. Front. Cell. Neurosci. 2013, 7. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.C.; Neher, J.J. Microglial Phagocytosis of Live Neurons. Nat. Rev. Neurosci. 2014, 15, 209–216. [Google Scholar] [CrossRef]
- Zahid, A.; Ismail, H.; Li, B.; Jin, T. Molecular and Structural Basis of DNA Sensors in Antiviral Innate Immunity. Front. Immunol. 2020, 11, 3094. [Google Scholar] [CrossRef]
- Motwani, M.; Pesiridis, S.; Fitzgerald, K.A. DNA Sensing by the CGAS–STING Pathway in Health and Disease. Nat. Rev. Genet. 2019, 20, 657–674. [Google Scholar] [CrossRef]
- Lugrin, J.; Martinon, F. The AIM2 Inflammasome: Sensor of Pathogens and Cellular Perturbations. Immunol. Rev. 2018, 281, 99–114. [Google Scholar] [CrossRef]
- Jakobsen, M.R.; Paludan, S.R. IFI16: At the Interphase between Innate DNA Sensing and Genome Regulation. Cytokine Growth Factor Rev. 2014, 25, 649–655. [Google Scholar] [CrossRef]
- Lin, S.-C.; Lo, Y.-C.; Wu, H. Helical Assembly in the MyD88-IRAK4-IRAK2 Complex in TLR/IL-1R Signalling. Nature 2010, 465, 885–890. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like Receptors and the Control of Immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef]
- Latz, E.; Visintin, A.; Espevik, T.; Golenbock, D.T. Mechanisms of TLR9 Activation. J. Endotoxin. Res. 2004, 10, 406–412. [Google Scholar] [CrossRef] [Green Version]
- Nie, Y.; Wang, Y.-Y. Innate Immune Responses to DNA Viruses. Protein Cell 2013, 4, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, O.; Akira, S. Innate Immunity to Virus Infection. Immunol. Rev. 2009, 227, 75–86. [Google Scholar] [CrossRef]
- Huang, X.; Yang, Y. Targeting the TLR9–MyD88 Pathway in the Regulation of Adaptive Immune Responses. Expert Opin. Ther. Targets 2010, 14, 787–796. [Google Scholar] [CrossRef] [Green Version]
- Chinnery, H.R.; McLenachan, S.; Binz, N.; Sun, Y.; Forrester, J.V.; Degli-Esposti, M.A.; Pearlman, E.; McMenamin, P.G. TLR9 Ligand CpG-ODN Applied to the Injured Mouse Cornea Elicits Retinal Inflammation. Am. J. Pathol. 2012, 180, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Deguine, J.; Barton, G.M. MyD88: A Central Player in Innate Immune Signaling. F1000Prime Rep. 2014, 6. [Google Scholar] [CrossRef]
- Rogers, G.L.; Suzuki, M.; Zolotukhin, I.; Markusic, D.M.; Morel, L.M.; Lee, B.; Ertl, H.C.J.; Herzog, R.W. Unique Roles of TLR9- and MyD88-Dependent and -Independent Pathways in Adaptive Immune Responses to AAV-Mediated Gene Transfer. J. Innate Immun. 2015, 7, 302–314. [Google Scholar] [CrossRef]
- Martino, A.T.; Suzuki, M.; Markusic, D.M.; Zolotukhin, I.; Ryals, R.C.; Moghimi, B.; Ertl, H.C.J.; Muruve, D.A.; Lee, B.; Herzog, R.W. The Genome of Self-Complementary Adeno-Associated Viral Vectors Increases Toll-like Receptor 9-Dependent Innate Immune Responses in the Liver. Blood 2011, 117, 6459–6468. [Google Scholar] [CrossRef] [Green Version]
- Rossi, A.; Dupaty, L.; Aillot, L.; Zhang, L.; Gallien, C.; Hallek, M.; Odenthal, M.; Adriouch, S.; Salvetti, A.; Büning, H. Vector Uncoating Limits Adeno-Associated Viral Vector-Mediated Transduction of Human Dendritic Cells and Vector Immunogenicity. Sci. Rep. 2019, 9, 3631. [Google Scholar] [CrossRef]
- Wang, S.K.; Lapan, S.W.; Hong, C.M.; Krause, T.B.; Cepko, C.L. In Situ Detection of Adeno-Associated Viral Vector Genomes with SABER-FISH. Mol. Ther.-Methods Clin. Dev. 2020, 19, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Wu, D.M.; Xue, Y.; Wang, S.K.; Chung, M.J.; Ji, X.; Rana, P.; Zhao, S.R.; Mai, S.; Cepko, C.L. AAV Cis-Regulatory Sequences Are Correlated with Ocular Toxicity. Proc. Natl. Acad. Sci. USA 2019, 116, 5785–5794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, J.F. Quantification of CpG Motifs in RAAV Genomes: Avoiding the Toll. Mol. Ther. 2020, 28, 1756–1758. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, G.; Krieg, A.M. Mechanism and Function of a Newly Identified CpG DNA Motif in Human Primary B Cells. J. Immunol. 2000, 164, 944–953. [Google Scholar] [CrossRef]
- Pohar, J.; Kužnik Krajnik, A.; Jerala, R.; Benčina, M. Minimal Sequence Requirements for Oligodeoxyribonucleotides Activating Human TLR9. J. Immunol. 2015, 194, 3901–3908. [Google Scholar] [CrossRef] [Green Version]
- Vabret, N.; Bhardwaj, N.; Greenbaum, B.D. Sequence-Specific Sensing of Nucleic Acids. Trends Immunol. 2017, 38, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Yaung, S.J.; Esvelt, K.M.; Church, G.M. CRISPR/Cas9-Mediated Phage Resistance Is Not Impeded by the DNA Modifications of Phage T4. PLoS ONE 2014, 9, e98811. [Google Scholar] [CrossRef] [Green Version]
- Ruan, G.-X.; Barry, E.; Yu, D.; Lukason, M.; Cheng, S.H.; Scaria, A. CRISPR/Cas9-Mediated Genome Editing as a Therapeutic Approach for Leber Congenital Amaurosis 10. Mol. Ther. 2017, 25, 331–341. [Google Scholar] [CrossRef] [Green Version]
- Editas Medicine Announces Positive Initial Clinical Data from Ongoing Phase 1/2 BRILLIANCE Clinical Trial of EDIT-101 for LCA10|Editas Medicine. Available online: https://ir.editasmedicine.com/news-releases/news-release-details/editas-medicine-announces-positive-initial-clinical-data-ongoing (accessed on 25 November 2021).
- Reichel, F.F.; Peters, T.; Wilhelm, B.; Biel, M.; Ueffing, M.; Wissinger, B.; Bartz-Schmidt, K.U.; Klein, R.; Michalakis, S.; Fischer, M.D.; et al. Humoral Immune Response After Intravitreal But Not After Subretinal AAV8 in Primates and Patients. Investig. Ophthalmol. Vis. Sci. 2018, 59, 1910–1915. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Miller, R.; Han, P.-Y.; Pang, J.; Dinculescu, A.; Chiodo, V.; Hauswirth, W.W. Intraocular Route of AAV2 Vector Administration Defines Humoral Immune Response and Therapeutic Potential. Mol. Vis. 2008, 14, 1760–1769. [Google Scholar]
- Li, W.; Kong, F.; Li, X.; Dai, X.; Liu, X.; Zheng, Q.; Wu, R.; Zhou, X.; Lü, F.; Chang, B.; et al. Gene Therapy Following Subretinal AAV5 Vector Delivery Is Not Affected by a Previous Intravitreal AAV5 Vector Administration in the Partner Eye. Mol. Vis. 2009, 15, 267–275. [Google Scholar]
- Vendomèle, J.; Dehmani, S.; Khebizi, Q.; Galy, A.; Fisson, S. Subretinal Injection of HY Peptides Induces Systemic Antigen-Specific Inhibition of Effector CD4+ and CD8+ T-Cell Responses. Front. Immunol. 2018, 9, 504. [Google Scholar] [CrossRef]
- Hageman, G.S.; Luthert, P.J.; Victor Chong, N.H.; Johnson, L.V.; Anderson, D.H.; Mullins, R.F. An Integrated Hypothesis That Considers Drusen as Biomarkers of Immune-Mediated Processes at the RPE-Bruch’s Membrane Interface in Aging and Age-Related Macular Degeneration. Prog. Retin Eye Res. 2001, 20, 705–732. [Google Scholar] [CrossRef]
- Hageman, G.S.; Anderson, D.H.; Johnson, L.V.; Hancox, L.S.; Taiber, A.J.; Hardisty, L.I.; Hageman, J.L.; Stockman, H.A.; Borchardt, J.D.; Gehrs, K.M.; et al. A Common Haplotype in the Complement Regulatory Gene Factor H (HF1/CFH) Predisposes Individuals to Age-Related Macular Degeneration. Proc. Natl. Acad. Sci. USA 2005, 102, 7227–7232. [Google Scholar] [CrossRef] [Green Version]
- Edwards, A.O.; Ritter, R.; Abel, K.J.; Manning, A.; Panhuysen, C.; Farrer, L.A. Complement Factor H Polymorphism and Age-Related Macular Degeneration. Science 2005, 308, 421–424. [Google Scholar] [CrossRef] [Green Version]
- Haines, J.L.; Hauser, M.A.; Schmidt, S.; Scott, W.K.; Olson, L.M.; Gallins, P.; Spencer, K.L.; Kwan, S.Y.; Noureddine, M.; Gilbert, J.R.; et al. Complement Factor H Variant Increases the Risk of Age-Related Macular Degeneration. Science 2005, 308, 419–421. [Google Scholar] [CrossRef] [Green Version]
- Klein, R.J.; Zeiss, C.; Chew, E.Y.; Tsai, J.-Y.; Sackler, R.S.; Haynes, C.; Henning, A.K.; SanGiovanni, J.P.; Mane, S.M.; Mayne, S.T.; et al. Complement Factor H Polymorphism in Age-Related Macular Degeneration. Science 2005, 308, 385–389. [Google Scholar] [CrossRef]
- Yoshida, N.; Ikeda, Y.; Notomi, S.; Ishikawa, K.; Murakami, Y.; Hisatomi, T.; Enaida, H.; Ishibashi, T. Clinical Evidence of Sustained Chronic Inflammatory Reaction in Retinitis Pigmentosa. Ophthalmology 2013, 120, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Bartel, M.; Schaffer, D.; Büning, H. Enhancing the Clinical Potential of AAV Vectors by Capsid Engineering to Evade Pre-Existing Immunity. Front. Microbiol. 2011, 2, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, J.-H.; Lim, K.; Schaffer, D.V. Library Selection and Directed Evolution Approaches to Engineering Targeted Viral Vectors. Biotechnol. Bioeng. 2007, 98, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Pavlou, M.; Schön, C.; Occelli, L.M.; Rossi, A.; Meumann, N.; Boyd, R.F.; Bartoe, J.T.; Siedlecki, J.; Gerhardt, M.J.; Babutzka, S.; et al. Novel AAV Capsids for Intravitreal Gene Therapy of Photoreceptor Disorders. EMBO Mol. Med. 2021, 13, e13392. [Google Scholar] [CrossRef]
- Santiago-Ortiz, J.; Ojala, D.S.; Westesson, O.; Weinstein, J.R.; Wong, S.Y.; Steinsapir, A.; Kumar, S.; Holmes, I.; Schaffer, D.V. AAV Ancestral Reconstruction Library Enables Selection of Broadly Infectious Viral Variants. Gene Ther. 2015, 22, 934–946. [Google Scholar] [CrossRef] [Green Version]
- Zinn, E.; Pacouret, S.; Khaychuk, V.; Turunen, H.T.; Carvalho, L.S.; Andres-Mateos, E.; Shah, S.; Shelke, R.; Maurer, A.C.; Plovie, E.; et al. In Silico Reconstruction of the Viral Evolutionary Lineage Yields a Potent Gene Therapy Vector. Cell Rep. 2015, 12, 1056–1068. [Google Scholar] [CrossRef] [Green Version]
- Mével, M.; Bouzelha, M.; Leray, A.; Pacouret, S.; Guilbaud, M.; Penaud-Budloo, M.; Alvarez-Dorta, D.; Dubreil, L.G.; Gouin, S.; Philippe Combal, J.; et al. Chemical Modification of the Adeno-Associated Virus Capsid to Improve Gene Delivery. Chem. Sci. 2020, 11, 1122–1131. [Google Scholar] [CrossRef] [Green Version]
- Faust, S.M.; Bell, P.; Cutler, B.J.; Ashley, S.N.; Zhu, Y.; Rabinowitz, J.E.; Wilson, J.M. CpG-Depleted Adeno-Associated Virus Vectors Evade Immune Detection. J. Clin. Investig. 2013, 123, 2994–3001. [Google Scholar] [CrossRef] [Green Version]
- Bertolini, T.B.; Shirley, J.L.; Zolotukhin, I.; Li, X.; Kaisho, T.; Xiao, W.; Kumar, S.R.P.; Herzog, R.W. Effect of CpG Depletion of Vector Genome on CD8+ T Cell Responses in AAV Gene Therapy. Front. Immunol. 2021, 12, 672449. [Google Scholar] [CrossRef]
- Cukras, C.; Wiley, H.E.; Jeffrey, B.G.; Sen, H.N.; Turriff, A.; Zeng, Y.; Vijayasarathy, C.; Marangoni, D.; Ziccardi, L.; Kjellstrom, S.; et al. Retinal AAV8-RS1 Gene Therapy for X-Linked Retinoschisis: Initial Findings from a Phase I/IIa Trial by Intravitreal Delivery. Mol. Ther. 2018, 26, 2282–2294. [Google Scholar] [CrossRef] [Green Version]
- Beltran, W.A.; Boye, S.L.; Boye, S.E.; Chiodo, V.A.; Lewin, A.S.; Hauswirth, W.W.; Aguirre, G.D. RAAV2/5 Gene-Targeting to Rods: Dose-Dependent Efficiency and Complications Associated with Different Promoters. Gene Ther. 2010, 17, 1162–1174. [Google Scholar] [CrossRef]
- Ramachandran, P.S.; Lee, V.; Wei, Z.; Song, J.Y.; Casal, G.; Cronin, T.; Willett, K.; Huckfeldt, R.; Morgan, J.I.W.; Aleman, T.S.; et al. Evaluation of Dose and Safety of AAV7m8 and AAV8BP2 in the Non-Human Primate Retina. Hum. Gene Ther. 2017, 28, 154–167. [Google Scholar] [CrossRef] [Green Version]
- Tobias, P.; Philipp, S.I.; Stylianos, M.; Martin, B.; Barbara, W.; Felix, R.; Alexander, O.G.; Eberhart, Z.; Marius, U.; Birgit, K.; et al. Safety and Toxicology of Ocular Gene Therapy with Recombinant AAV Vector RAAV.HCNGA3 in Nonhuman Primates. Hum. Gene Ther. Clin. Dev. 2019, 30, 50–56. [Google Scholar] [CrossRef]
- Dufour, V.L.; Cideciyan, A.V.; Ye, G.-J.; Song, C.; Timmers, A.; Habecker, P.L.; Pan, W.; Weinstein, N.M.; Swider, M.; Durham, A.C.; et al. Toxicity and Efficacy Evaluation of an Adeno-Associated Virus Vector Expressing Codon-Optimized RPGR Delivered by Subretinal Injection in a Canine Model of X-Linked Retinitis Pigmentosa. Hum. Gene Ther. 2020, 31, 253–267. [Google Scholar] [CrossRef]
- Ye, G.; Budzynski, E.; Sonnentag, P.; Nork, T.M.; Miller, P.E.; Sharma, A.K.; Ver Hoeve, J.N.; Smith, L.M.; Arndt, T.; Calcedo, R.; et al. Safety and Biodistribution Evaluation in Cynomolgus Macaques of RAAV2tYF-PR1.7-HCNGB3, a Recombinant AAV Vector for Treatment of Achromatopsia. Hum. Gene Ther. Clin. Dev. 2016, 27, 37–48. [Google Scholar] [CrossRef] [Green Version]
- Song, C.; Dufour, V.L.; Cideciyan, A.V.; Ye, G.-J.; Swider, M.; Newmark, J.A.; Timmers, A.M.; Robinson, P.M.; Knop, D.R.; Chulay, J.D.; et al. Dose Range Finding Studies with Two RPGR Transgenes in a Canine Model of X-Linked Retinitis Pigmentosa Treated with Subretinal Gene Therapy. Hum. Gene Ther. 2020, 31, 743–755. [Google Scholar] [CrossRef]
- Ye, G.-J.; Komáromy, A.M.; Zeiss, C.; Calcedo, R.; Harman, C.D.; Koehl, K.L.; Stewart, G.A.; Iwabe, S.; Chiodo, V.A.; Hauswirth, W.W.; et al. Safety and Efficacy of AAV5 Vectors Expressing Human or Canine CNGB3 in CNGB3-Mutant Dogs. Hum. Gene Clin. Dev. 2017, 28, 197–207. [Google Scholar] [CrossRef]
- McCarty, D.M. Self-Complementary AAV Vectors; Advances and Applications. Mol. Ther. 2008, 16, 1648–1656. [Google Scholar] [CrossRef]
- Wu, T.; Töpfer, K.; Lin, S.-W.; Li, H.; Bian, A.; Zhou, X.Y.; High, K.A.; Ertl, H.C. Self-Complementary AAVs Induce More Potent Transgene Product-Specific Immune Responses Compared to a Single-Stranded Genome. Mol. Ther 2012, 20, 572–579. [Google Scholar] [CrossRef] [Green Version]
- Chan, Y.K.; Dick, A.D.; Hall, S.M.; Langmann, T.; Scribner, C.L.; Mansfield, B.C. Inflammation in Viral Vector-Mediated Ocular Gene Therapy: A Review and Report From a Workshop Hosted by the Foundation Fighting Blindness, 9/2020. Transl. Vis. Sci. Technol. 2021, 10, 3. [Google Scholar] [CrossRef]
- Chu, W.S.; Ng, J. Immunomodulation in Administration of RAAV: Preclinical and Clinical Adjuvant Pharmacotherapies. Front. Immunol. 2021, 12, 858. [Google Scholar] [CrossRef]
- FRANCISCO, E.M. Zolgensma. Available online: https://www.ema.europa.eu/en/medicines/human/EPAR/zolgensma (accessed on 30 October 2021).
- Mingozzi, F.; Anguela, X.M.; Pavani, G.; Chen, Y.; Davidson, R.J.; Hui, D.J.; Yazicioglu, M.; Elkouby, L.; Hinderer, C.J.; Faella, A.; et al. Overcoming Preexisting Humoral Immunity to AAV Using Capsid Decoys. Sci Transl Med. 2013, 5, 194ra92. [Google Scholar] [CrossRef] [Green Version]
- Ng, C.C.; Sy, A.; Cunningham, E.T. Rituximab for Non-Infectious Uveitis and Scleritis. J. Ophthalmic Inflamm. Infect. 2021, 11, 23. [Google Scholar] [CrossRef]
- Finn, J.D.; Hui, D.; Downey, H.D.; Dunn, D.; Pien, G.C.; Mingozzi, F.; Zhou, S.; High, K.A. Proteasome Inhibitors Decrease AAV2 Capsid Derived Peptide Epitope Presentation on MHC Class I Following Transduction. Mol. Ther. 2010, 18, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Hamer, M.; Stamatakis, E. Inflammation as an Intermediate Pathway in the Association between Psychosocial Stress and Obesity. Physiol. Behav. 2008, 94, 536–539. [Google Scholar] [CrossRef]
- Pathanapitoon, K.; Dodds, E.M.; Cunningham, E.T.; Rothova, A. Clinical Spectrum of HLA-B27-Associated Ocular Inflammation. Ocul. Immunol. Inflamm. 2017, 25, 569–576. [Google Scholar] [CrossRef] [Green Version]
- Bouquet, C.; Vignal Clermont, C.; Galy, A.; Fitoussi, S.; Blouin, L.; Munk, M.R.; Valero, S.; Meunier, S.; Katz, B.; Sahel, J.A.; et al. Immune Response and Intraocular Inflammation in Patients With Leber Hereditary Optic Neuropathy Treated With Intravitreal Injection of Recombinant Adeno-Associated Virus 2 Carrying the ND4 Gene: A Secondary Analysis of a Phase 1/2 Clinical Trial. JAMA Ophthalmol. 2019, 137, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Jabs, D.A.; Nussenblatt, R.B.; Rosenbaum, J.T. Standardization of Uveitis Nomenclature (SUN) Working Group Standardization of Uveitis Nomenclature for Reporting Clinical Data. Results of the First International Workshop. Am. J. Ophthalmol. 2005, 140, 509–516. [Google Scholar] [CrossRef]


| Type of Element | Sequence | Size of Sequence | Number of CG Sequences | RF1 |
|---|---|---|---|---|
| AAV sequences | ITR2 left (1) | 145 | 16 | 11.03% |
| ITR2 right (1) | 145 | 16 | 11.03% | |
| ITR5 left (1) | 166 | 16 | 9.64% | |
| ITR5 right (1) | 166 | 16 | 9.64% | |
| Promoter | CMV (1) | 581 | 32 | 5.51% |
| CBA (1) | 1557 | 181 | 11.62% | |
| CAG (1) | 1733 | 186 | 10.73% | |
| hRPE65 (1) | 1382 | 5 | 0.36% | |
| hRK (1) | 241 | 10 | 4.15% | |
| hRho (1) | 848 | 9 | 1.06% | |
| U6 (1) | 249 | 6 | 2.41% | |
| GFAP (1) | 696 | 19 | 2.73% | |
| EF1a (1) | 1179 | 94 | 7.97% | |
| hSyn (1) | 485 | 56 | 11.55% | |
| SV40 (1) | 269 | 9 | 3.35% | |
| hPGK (1) | 507 | 62 | 12.23% | |
| HSP (=P1) (2) | 600 | 83 | 13.83% | |
| VMD2 (2) | 600 | 8 | 1.33% | |
| Rs1 (2) | 600 | 19 | 3.17% | |
| Transgene | hRPE65 (1) | 1602 | 21 | 1.31% |
| hRDH12 (1) | 960 | 33 | 3.44% | |
| SaCas9 (1) | 3156 | 168 | 5.32% | |
| hRDH8 (1) | 936 | 43 | 4.59% | |
| ChR2 (3) | 930 | 65 | 6.99% | |
| BDNF (3) | 3985 | 87 | 2.18% | |
| GNAT2 (3) | 1515 | 29 | 1.91% | |
| CNTF (3) | 1902 | 27 | 1.42% | |
| eGFP (1) | 717 | 60 | 8.37% | |
| SpCas9 (1) | 4275 | 254 | 5.94% | |
| Rho (3) | 1047 | 52 | 4.97% | |
| VEGFA (3) | 1239 | 95 | 7.67% | |
| PDE6b (3) | 2565 | 138 | 5.38% | |
| RDCVF (3) | 639 | 50 | 7.82% | |
| Cas12a (CpfI) (3) | 3903 | 33 | 0.85% | |
| RPE1 (3) | 717 | 29 | 4.04% | |
| Mertk (3) | 3000 | 77 | 2.57% | |
| RPGR (3) | 2448 | 29 | 1.18% | |
| sFLT1 (3) | 147 | 1 | 0.68% | |
| Rs1 (3) | 675 | 27 | 4.00% | |
| ND4 (4) | 1378 | 31 | 2.25% | |
| Suppl. elements | IRES (1) | 587 | 29 | 4.94% |
| WPRE (1) | 592 | 37 | 6.25% | |
| SV40 PolyA (1) | 131 | 0 | 0.00% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varin, J.; Morival, C.; Maillard, N.; Adjali, O.; Cronin, T. Risk Mitigation of Immunogenicity: A Key to Personalized Retinal Gene Therapy. Int. J. Mol. Sci. 2021, 22, 12818. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222312818
Varin J, Morival C, Maillard N, Adjali O, Cronin T. Risk Mitigation of Immunogenicity: A Key to Personalized Retinal Gene Therapy. International Journal of Molecular Sciences. 2021; 22(23):12818. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222312818
Chicago/Turabian StyleVarin, Juliette, Clément Morival, Noémien Maillard, Oumeya Adjali, and Therese Cronin. 2021. "Risk Mitigation of Immunogenicity: A Key to Personalized Retinal Gene Therapy" International Journal of Molecular Sciences 22, no. 23: 12818. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222312818