
Increased Systemic Antioxidant Power Ameliorates the Aging-Related Reduction in Oocyte Competence in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
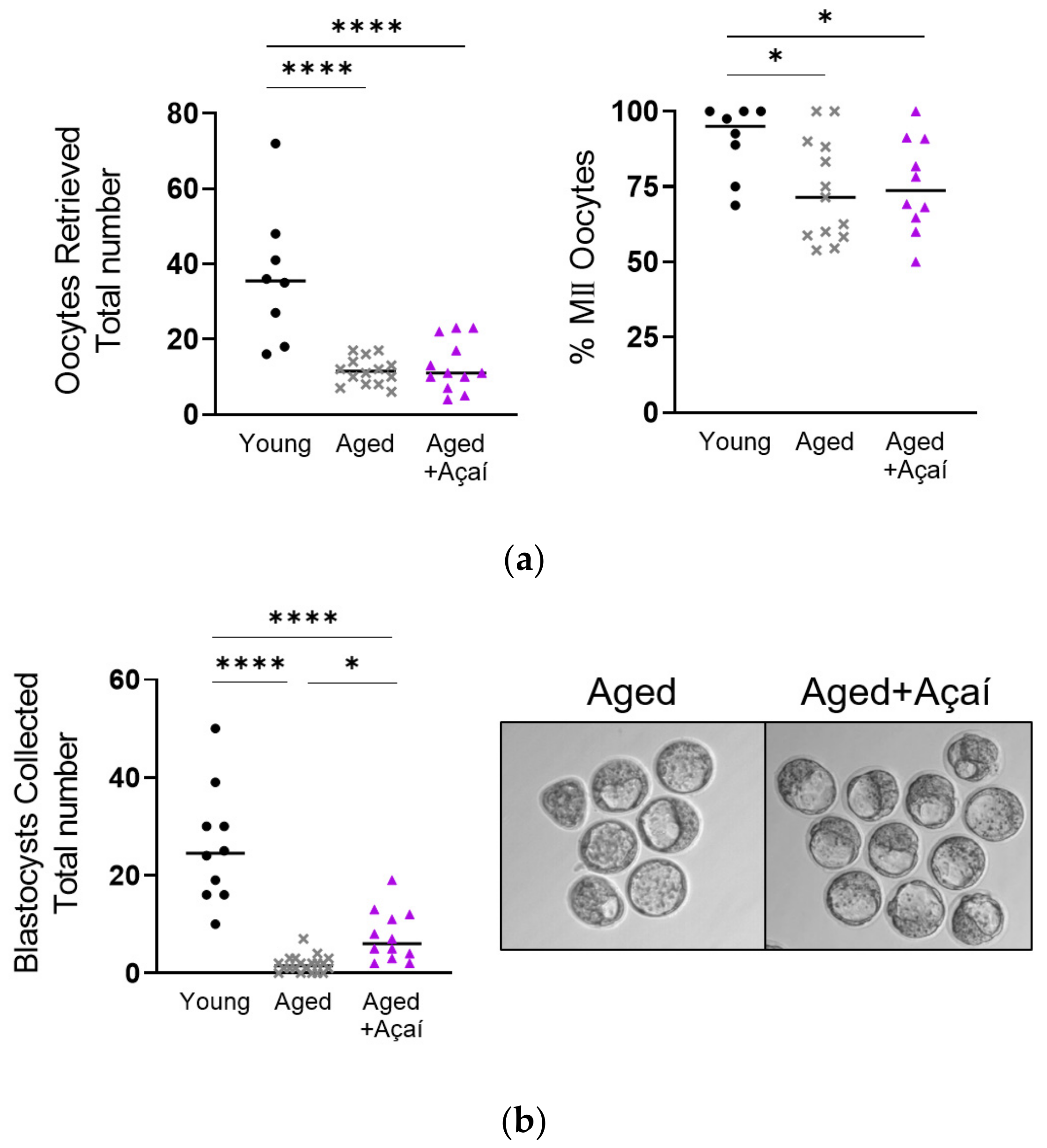
2.1. Reproductive Outcomes
2.2. Serum Changes following Aging and Antioxidant Supplementation
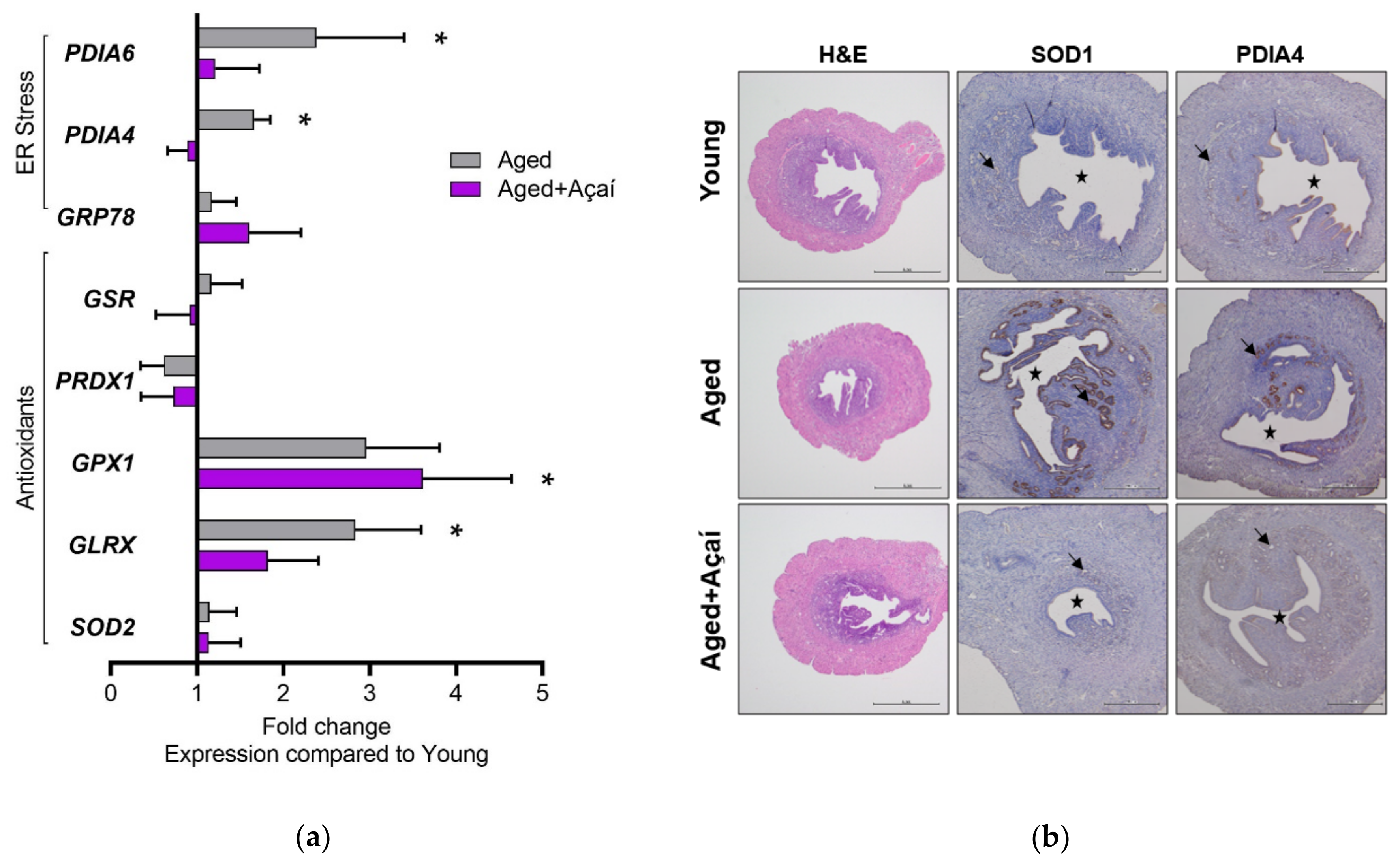
2.3. Effects of Aging and Açaí on Ovarian ER Stress
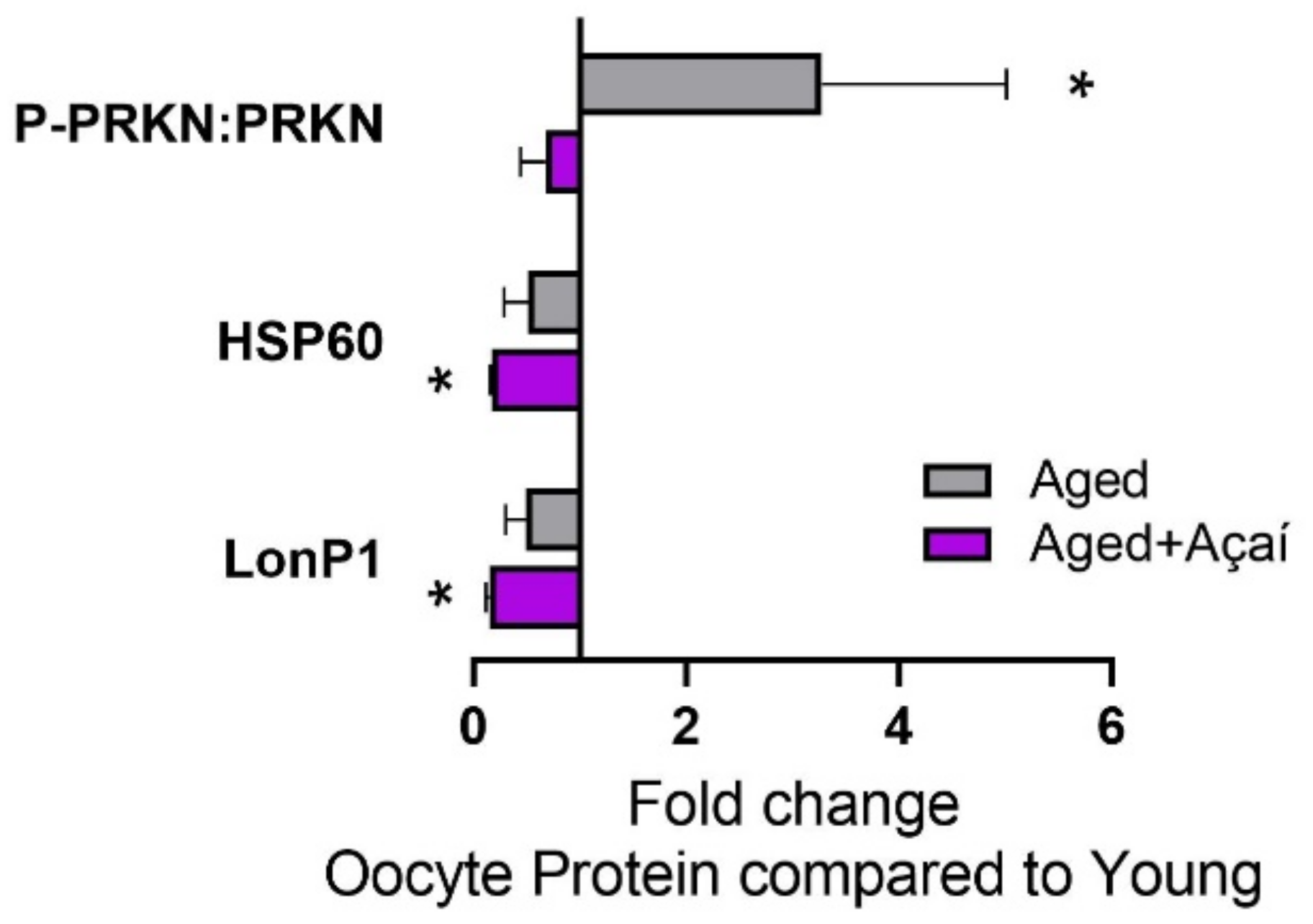
2.4. Alterations in Cellular Stress Pathways in the Oocyte
2.5. Molecular Effects of Aging and Açaí in the Uterus
3. Discussion
4. Materials and Methods
4.1. Animals and Antioxidant Supplementation
4.2. Isolation of the Oocytes and Preimplantation Embryos
4.3. Embryo Transfer
4.4. Serum DNA Damage and FRAP Assays
4.5. RNA Isolation and Quantitative Polymerase Chain Reaction
4.6. RNA In Situ Hybridization and Immunohistochemistry
4.7. Jess Simple Western Blotting
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Keefe, D.; Kumar, M.; Kalmbach, K. Oocyte competency is the key to embryo potential. Fertil. Steril. 2015, 103, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Llarena, N.; Hine, C. Reproductive Longevity and Aging: Geroscience Approaches to Maintain Long-Term Ovarian Fitness. J. Gerontol. Ser. A Boil. Sci. Med. Sci. 2020, 76, 1551–1560. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.A.; E Hamilton, B.; Osterman, M.J.K.; Driscoll, A.K. Births: Final Data for 2018. Natl. Vital Stat. Rep. 2019, 68, 1–47. [Google Scholar] [PubMed]
- La Marca, A.; Capuzzo, M.; Imbrogno, M.G.; Donno, V.; Spedicato, G.A.; Sacchi, S.; Minasi, M.G.; Spinella, F.; Greco, P.; Fiorentino, F.; et al. The complex relationship between female age and embryo euploidy. Minerva Obstet. Gynecol. 2021, 73, 103–110. [Google Scholar] [CrossRef]
- Greaney, J.; Wei, Z.; Homer, H. Regulation of chromosome segregation in oocytes and the cellular basis for female meiotic errors. Hum. Reprod. Updat. 2017, 24, 135–161. [Google Scholar] [CrossRef]
- Van Blerkom, J. Mitochondrial function in the human oocyte and embryo and their role in developmental competence. Mitochondrion 2011, 11, 797–813. [Google Scholar] [CrossRef]
- Lord, T.; Aitken, R.J. Oxidative stress and ageing of the post-ovulatory oocyte. Reproduction 2013, 146, R217–R227. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.; Luderer, U. Oxidative Damage Increases and Antioxidant Gene Expression Decreases with Aging in the Mouse Ovary. Biol. Reprod. 2011, 84, 775–782. [Google Scholar] [CrossRef] [Green Version]
- Ruder, E.H.; Hartman, T.J.; Blumberg, J.; Goldman, M.B. Oxidative stress and antioxidants: Exposure and impact on female fertility. Hum. Reprod. Updat. 2008, 14, 345–357. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Zheng, Y.; Li, J.; Yu, Y.; Zhang, W.; Song, M.; Liu, Z.; Min, Z.; Hu, H.; Jing, Y.; et al. Single-Cell Transcriptomic Atlas of Primate Ovarian Aging. Cell 2020, 180, 585–600.e19. [Google Scholar] [CrossRef]
- Fujii, J.; Iuchi, Y.; Okada, F. Fundamental roles of reactive oxygen species and protective mechanisms in the female reproductive system. Reprod. Biol. Endocrinol. 2005, 3, 43. [Google Scholar] [CrossRef] [Green Version]
- Freitas, C.; Neto, A.C.; Matos, L.; Silva, E.; Ribeiro, Â.; Silva-Carvalho, J.L.; Almeida, H. Follicular Fluid redox involvement for ovarian follicle growth. J. Ovarian Res. 2017, 10, 44. [Google Scholar] [CrossRef]
- Bianchi, S.; Macchiarelli, G.; Micara, G.; Linari, A.; Boninsegna, C.; Aragona, C.; Rossi, G.; Cecconi, S.; Nottola, S.A. Ultrastruc tural markers of quality are impaired in human metaphase II aged oocytes: A comparison between reproductive and in vitro aging. J. Assist. Reprod. Genet. 2015, 32, 1343–1358. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Wheeler, M.; Krisher, R.L. Disrupted Redox Homeostasis and Aberrant Redox Gene Expression in Porcine Oocytes Contribute to Decreased Developmental Competence1. Biol. Reprod. 2012, 87, 78. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial Reactive Oxygen Species (ROS) and ROS-Induced ROS Release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef] [Green Version]
- Ray, P.D.; Huang, B.-W.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell. Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef] [Green Version]
- Poprac, P.; Jomova, K.; Simunkova, M.; Kollar, V.; Rhodes, C.J.; Valko, M. Targeting Free Radicals in Oxidative Stress-Related Human Diseases. Trends Pharmacol. Sci. 2017, 38, 592–607. [Google Scholar] [CrossRef]
- García-Sánchez, A.; Miranda-Díaz, A.G.; Cardona-Muñoz, E.G. The Role of Oxidative Stress in Physiopathology and Pharmacological Treatment with Pro- and Antioxidant Properties in Chronic Diseases. Oxidative Med. Cell. Longev. 2020, 2020, e2082145. [Google Scholar] [CrossRef]
- Espino, J.; Macedo, M.; Lozano, G.; Ortiz, Á.; Rodríguez, C.; Rodríguez, A.B.; Bejarano, I. Impact of Melatonin Supplementation in Women with Unexplained Infertility Undergoing Fertility Treatment. Antioxidants 2019, 8, 338. [Google Scholar] [CrossRef] [Green Version]
- Giannubilo, S.R.; Orlando, P.; Silvestri, S.; Cirilli, I.; Marcheggiani, F.; Ciavattini, A.; Tiano, L. CoQ10 Supplementation in Patients Undergoing IVF-ET: The Relationship with Follicular Fluid Content and Oocyte Maturity. Antioxidants 2018, 7, 141. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Varela, C.; Labarta, E. Does Coenzyme Q10 Supplementation Improve Human Oocyte Quality? Int. J. Mol. Sci. 2021, 22, 9541. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Nisenblat, V.; Lu, C.; Li, R.; Qiao, J.; Zhen, X.; Wang, S. Pretreatment with coenzyme Q10 improves ovarian response and embryo quality in low-prognosis young women with decreased ovarian reserve: A randomized controlled trial. Reprod. Biol. Endocrinol. 2018, 16, 29. [Google Scholar] [CrossRef] [PubMed]
- Earling, M.; Beadle, T.; Niemeyer, E.D. Açai Berry (Euterpe oleracea) Dietary Supplements: Variations in Anthocyanin and Flavonoid Concentrations, Phenolic Contents, and Antioxidant Properties. Plant Foods Hum. Nutr. 2019, 74, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Dattilo, M.; D’Amato, G.; Caroppo, E.; Ménézo, Y. Erratum to: Improvement of gamete quality by stimulating and feeding the endogenous antioxidant system: Mechanisms, clinical results, insights on gene-environment interactions and the role of diet. J. Assist. Reprod. Genet. 2016, 34, 155. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, P.O.; Souza, M.O.; Silva, M.P.; Santos, G.T.; Silva, M.E.; Bermano, G.; Freitas, R.N. Açaí (Euterpe oleracea Martius) supplementation improves oxidative stress biomarkers in liver tissue of dams fed a high-fat diet and increases antioxidant enzymes’ gene expression in offspring. Biomed. Pharmacother. 2021, 139, 111627. [Google Scholar] [CrossRef]
- Vilhena, J.C.; Lopes de Melo Cunha, L.; Jorge, T.M.; de Lucena Machado, M.; de Andrade Soares, R.; Santos, I.B.; Freitas de Bem, G.; Fernandes-Santos, C.; Ognibene, D.T.; Soares de Moura, R.; et al. Açaí Reverses Adverse Cardiovascular Remodeling in Renovascular Hypertension: A Comparative Effect with Enalapril. J. Cardiovasc. Pharmacol. 2021, 77, 673–684. [Google Scholar] [CrossRef]
- Monteiro, C.; Filho, H.; Silva, F.G.O.; de Souza, M.F.F.; Sousa, J.A.O.; Franco Á, X.; Resende Â, C.; de Moura, R.S.; de Souza, M.H.L.; Soares, P.M.G.; et al. Euterpe oleracea Mart. (Açaí) attenuates experimental colitis in rats: Involvement of TLR4/COX-2/NF-ĸB. Inflammopharmacology 2021, 29, 193–204. [Google Scholar] [CrossRef]
- Katz-Jaffe, M.G.; Lane, S.L.; Parks, J.C.; McCallie, B.R.; Makloski, R.; Schoolcraft, W.B. Antioxidant Intervention Attenuates Aging-Related Changes in the Murine Ovary and Oocyte. Life 2020, 10, 250. [Google Scholar] [CrossRef]
- Budani, M.C.; Tiboni, G.M. Effects of Supplementation with Natural Antioxidants on Oocytes and Preimplantation Embryos. Antioxidants 2020, 9, 612. [Google Scholar] [CrossRef]
- Kawwass, J.F.; Monsour, M.; Crawford, S.; Kissin, D.M.; Session, D.R.; Kulkarni, A.D.; Jamieson, D.; National ART Surveillance System (NASS) Group. Trends and outcomes for donor oocyte cycles in the United States, 2000–2010. JAMA 2013, 310, 2426–2434. [Google Scholar] [CrossRef] [Green Version]
- Igarashi, H.; Takahashi, E.; Hiroi, M.; Doi, K. Aging-related changes in calcium oscillations in fertilized mouse oocytes. Mol. Reprod. Dev. 1997, 48, 383–390. [Google Scholar] [CrossRef]
- Brenjian, S.; Moini, A.; Yamini, N.; Kashani, L.; Faridmojtahedi, M.; Bahramrezaie, M.; Khodarahmian, M.; Amidi, F. Resveratrol treatment in patients with polycystic ovary syndrome decreased pro-inflammatory and endoplasmic reticulum stress markers. Am. J. Reprod. Immunol. 2019, 83, e13186. [Google Scholar] [CrossRef]
- Lin, X.; Dai, Y.; Tong, X.; Xu, W.; Huang, Q.; Jin, X.; Li, C.; Zhou, F.; Zhou, H.; Lin, X.; et al. Excessive oxidative stress in cumulus granulosa cells induced cell senescence contributes to endometriosis-associated infertility. Redox Biol. 2020, 30, 101431. [Google Scholar] [CrossRef]
- Lin, T.; Lee, J.E.; Kang, J.W.; Shin, H.Y.; Bin Lee, J.; Jin, D.I. Endoplasmic Reticulum (ER) Stress and Unfolded Protein Response (UPR) in Mammalian Oocyte Maturation and Preimplantation Embryo Development. Int. J. Mol. Sci. 2019, 20, 409. [Google Scholar] [CrossRef] [Green Version]
- May-Panloup, P.; Boucret, L.; De La Barca, J.-M.C.; Desquiret-Dumas, V.; Ferré-L’Hotellier, V.; Morinière, C.; Descamps, P.; Procaccio, V.; Reynier, P. Ovarian ageing: The role of mitochondria in oocytes and follicles. Hum. Reprod. Updat. 2016, 22, 725–743. [Google Scholar] [CrossRef] [Green Version]
- Chocron, E.S.; Munkácsy, E.; Pickering, A.M. Cause or casualty: The role of mitochondrial DNA in aging and age-associated disease. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1865, 285–297. [Google Scholar] [CrossRef]
- Simsek-Duran, F.; Li, F.; Ford, W.; Swanson, R.J.; Jones, H.W., Jr.; Castora, F.J. Age-Associated Metabolic and Morphologic Changes in Mitochondria of Individual Mouse and Hamster Oocytes. PLoS ONE 2013, 8, e64955. [Google Scholar] [CrossRef] [Green Version]
- Sykora, P.; Kanno, S.; Akbari, M.; Kulikowicz, T.; Baptiste, B.A.; Leandro, G.S.; Lu, H.; Tian, J.; May, A.; Becker, K.A.; et al. DNA Polymerase Beta Participates in Mitochondrial DNA Repair. Mol. Cell. Biol. 2017, 37, e00237–e00317. [Google Scholar] [CrossRef] [Green Version]
- Kelley, M.R.; Parsons, S.H. Redox regulation of the DNA repair function of the human AP endonuclease Ape1/ref-1. Antioxid. Redox Signal 2001, 3, 671–683. [Google Scholar] [CrossRef]
- Bravard, A.; Vacher, M.; Moritz, E.; Vaslin, L.; Hall, J.; Epe, B.; Radicella, J.P. Oxidation Status of Human OGG1-S326C Polymorphic Variant Determines Cellular DNA Repair Capacity. Cancer Res. 2009, 69, 3642–3649. [Google Scholar] [CrossRef] [Green Version]
- Cuneo, M.J.; London, R.E. Oxidation state of the XRCC1 N-terminal domain regulates DNA polymerase beta binding affinity. Proc. Natl. Acad. Sci. USA 2010, 107, 6805–6810. [Google Scholar] [CrossRef] [Green Version]
- Gaziev, A.I.; Abdullaev, S.; Podlutsky, A. Mitochondrial function and mitochondrial DNA maintenance with advancing age. Biogerontology 2014, 15, 417–438. [Google Scholar] [CrossRef]
- Ding, W.X.; Yin, X.M. Mitophagy: Mechanisms, pathophysiological roles, and analysis. Biol. Chem. 2012, 393, 547–564. [Google Scholar] [CrossRef] [Green Version]
- Pollecker, K.; Sylvester, M.; Voos, W. Proteomic analysis demonstrates the role of the quality control protease LONP1 in mitochondrial protein aggregation. J. Biol. Chem. 2021, 297, e101134. [Google Scholar] [CrossRef]
- Van Blerkom, J.; Davis, P.W.; Lee, J. Fertilization and early embryolgoy: ATP content of human oocytes and developmental potential and outcome after in-vitro fertilization and embryo transfer. Hum. Reprod. 1995, 10, 415–424. [Google Scholar] [CrossRef]
- Igarashi, H.; Takahashi, T.; Takahashi, E.; Tezuka, N.; Nakahara, K.; Takahashi, K.; Kurachi, H. Aged Mouse Oocytes Fail to Readjust Intracellular Adenosine Triphosphates at Fertilization1. Biol. Reprod. 2005, 72, 1256–1261. [Google Scholar] [CrossRef]
- Wang, L.; Tang, J.; Wang, L.; Tan, F.; Song, H.; Zhou, J.; Li, F. Oxidative stress in oocyte aging and female reproduction. J. Cell. Physiol. 2021. [Google Scholar] [CrossRef]
- Shaw, P.; Chattopadhyay, A. Nrf2–ARE signaling in cellular protection: Mechanism of action and the regulatory mechanisms. J. Cell. Physiol. 2019, 235, 3119–3130. [Google Scholar] [CrossRef]
- Ryoo, I.G.; Kwak, M.K. Regulatory crosstalk between the oxidative stress-related transcription factor Nfe2l2/Nrf2 and mitochondria. Toxicol. Appl. Pharmacol. 2018, 359, 24–33. [Google Scholar] [CrossRef]
- Chen, S.; Lu, Y.; Chen, Y.; Xu, J.; Chen, L.; Zhao, W.; Wang, T.; Wang, H.; Wang, P. The effect of Bu Shen Huo Xue Tang on autoimmune premature ovarian insufficiency via Modulation of the Nrf2/Keap1 signaling pathway in mice. J. Ethnopharmacol. 2021, 273, 113996. [Google Scholar] [CrossRef]
- Liu, X.; Lin, X.; Zhang, S.; Guo, C.; Li, J.; Mi, Y.; Zhang, C. Lycopene ameliorates oxidative stress in the aging chicken ovary via activation of Nrf2/HO-1 pathway. Aging 2018, 10, 2016–2036. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, N.; Zeng, Z.; Tang, L.; Zhao, S.; Zhou, F.; Zhou, L.; Xia, W.; Zhu, C.; Rao, M. Humanin regulates oxidative stress in the ovaries of polycystic ovary syndrome patients via the Keap1/Nrf2 pathway. Mol. Hum. Reprod. 2021, 27, 2. [Google Scholar] [CrossRef] [PubMed]
- Odendaal, A.Y.; Schauss, A.G. Chapter 18—Potent Antioxidant and Anti-Inflammatory Flavonoids in the Nutrient-Rich Amazonian Palm Fruit, Açaí (Euterpe spp.). In Polyphenols in Human Health and Disease; Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 219–239. [Google Scholar]
- Sugino, N.; Shimamura, K.; Takiguchi, S.; Tamura, H.; Ono, M.; Nakata, M.; Nakamura, Y.; Ogino, K.; Uda, T.; Kato, H. Changes in activity of superoxide dismutase in the human endometrium throughout the menstrual cycle and in early pregnancy. Hum. Reprod. 1996, 11, 1073–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, C.; Pires, M.D.A.; Santos, D.; Carreira, R.P. Distribution of superoxide dismutase 1 and glutathione peroxidase 1 in the cyclic canine endometrium. Theriogenology 2016, 86, 738–748. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, A.; DeMayo, F.J.; E Spencer, T. Uterine Glands: Developmental Biology and Functional Roles in Pregnancy. Endocr. Rev. 2019, 40, 1424–1445. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lane, S.L.; Parks, J.C.; Russ, J.E.; Khan, S.A.; Schoolcraft, W.B.; Yuan, Y.; Katz-Jaffe, M.G. Increased Systemic Antioxidant Power Ameliorates the Aging-Related Reduction in Oocyte Competence in Mice. Int. J. Mol. Sci. 2021, 22, 13019. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313019
Lane SL, Parks JC, Russ JE, Khan SA, Schoolcraft WB, Yuan Y, Katz-Jaffe MG. Increased Systemic Antioxidant Power Ameliorates the Aging-Related Reduction in Oocyte Competence in Mice. International Journal of Molecular Sciences. 2021; 22(23):13019. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313019
Chicago/Turabian StyleLane, Sydney L., Jason C. Parks, Jennifer E. Russ, Shaihla A. Khan, William B. Schoolcraft, Ye Yuan, and Mandy G. Katz-Jaffe. 2021. "Increased Systemic Antioxidant Power Ameliorates the Aging-Related Reduction in Oocyte Competence in Mice" International Journal of Molecular Sciences 22, no. 23: 13019. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313019