
Investigation of PtSGT1 and PtSGT4 Function in Cellulose Biosynthesis in Populus tomentosa Using CRISPR/Cas9 Technology

Abstract
:1. Introduction
2. Results
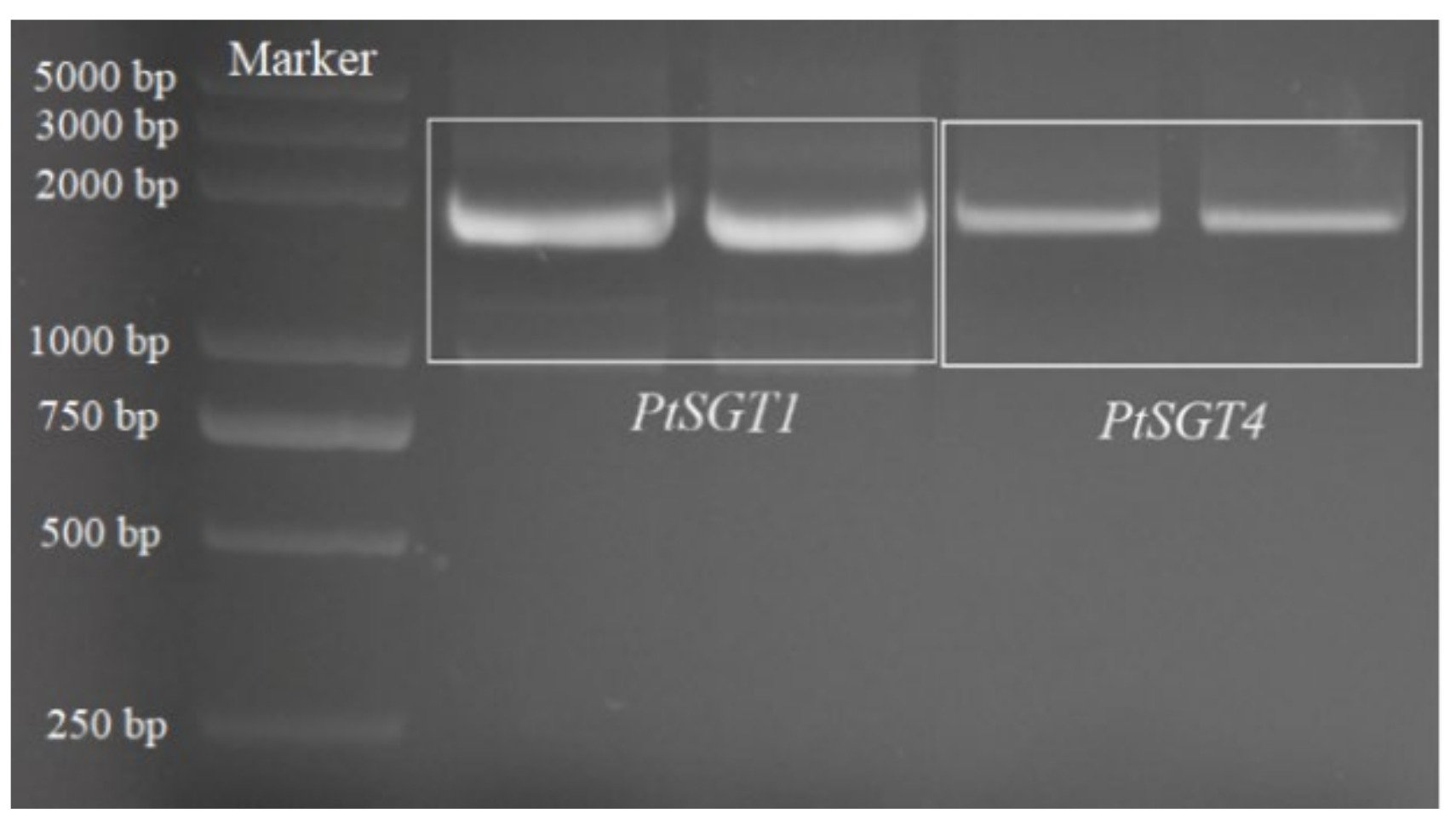
2.1. Cloning and Sequence Analysis of PtSGT1 and PtSGT4
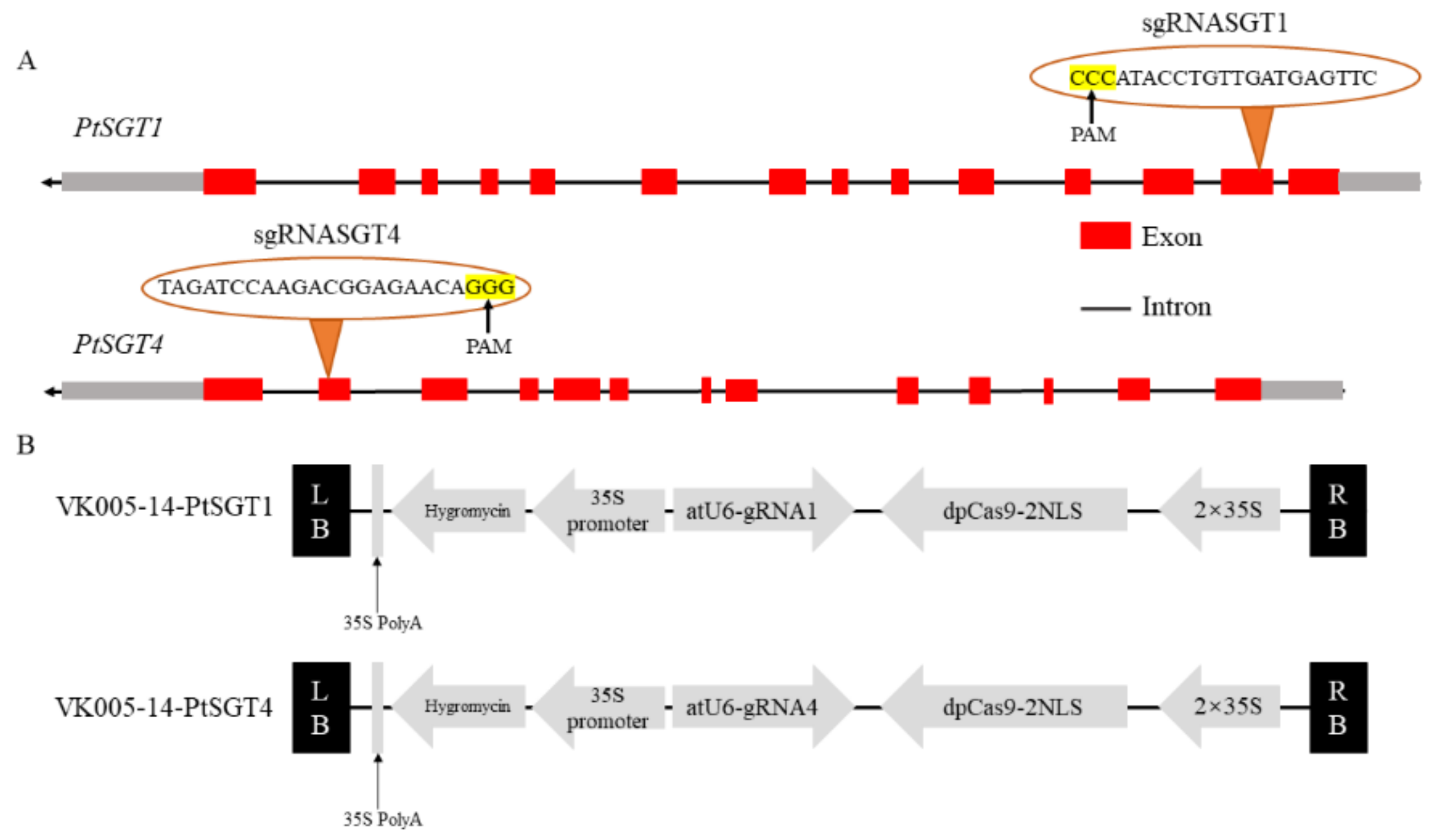
2.2. Plasmid Construction
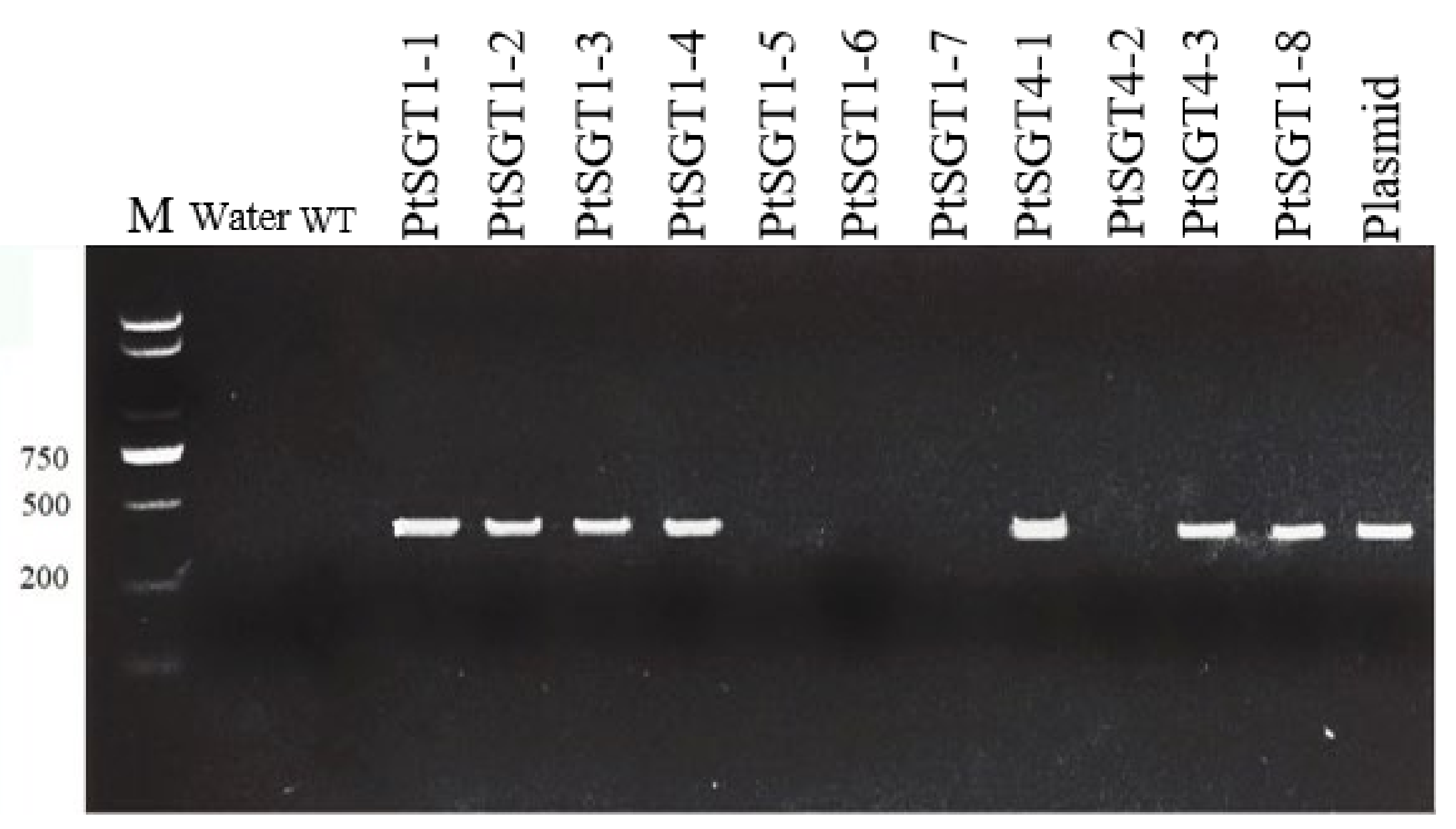
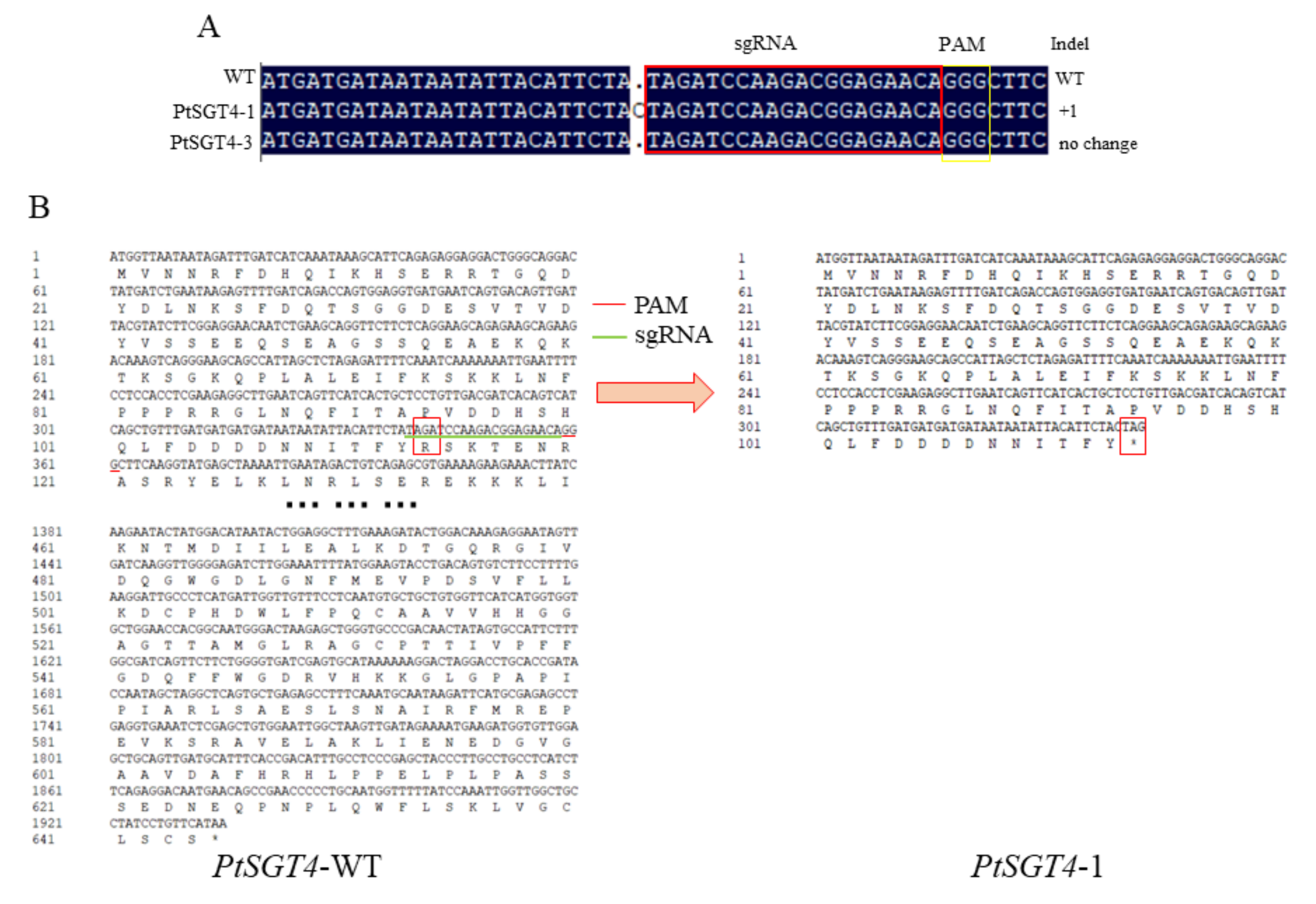
2.3. Genetic Transformation of P. tomentosa
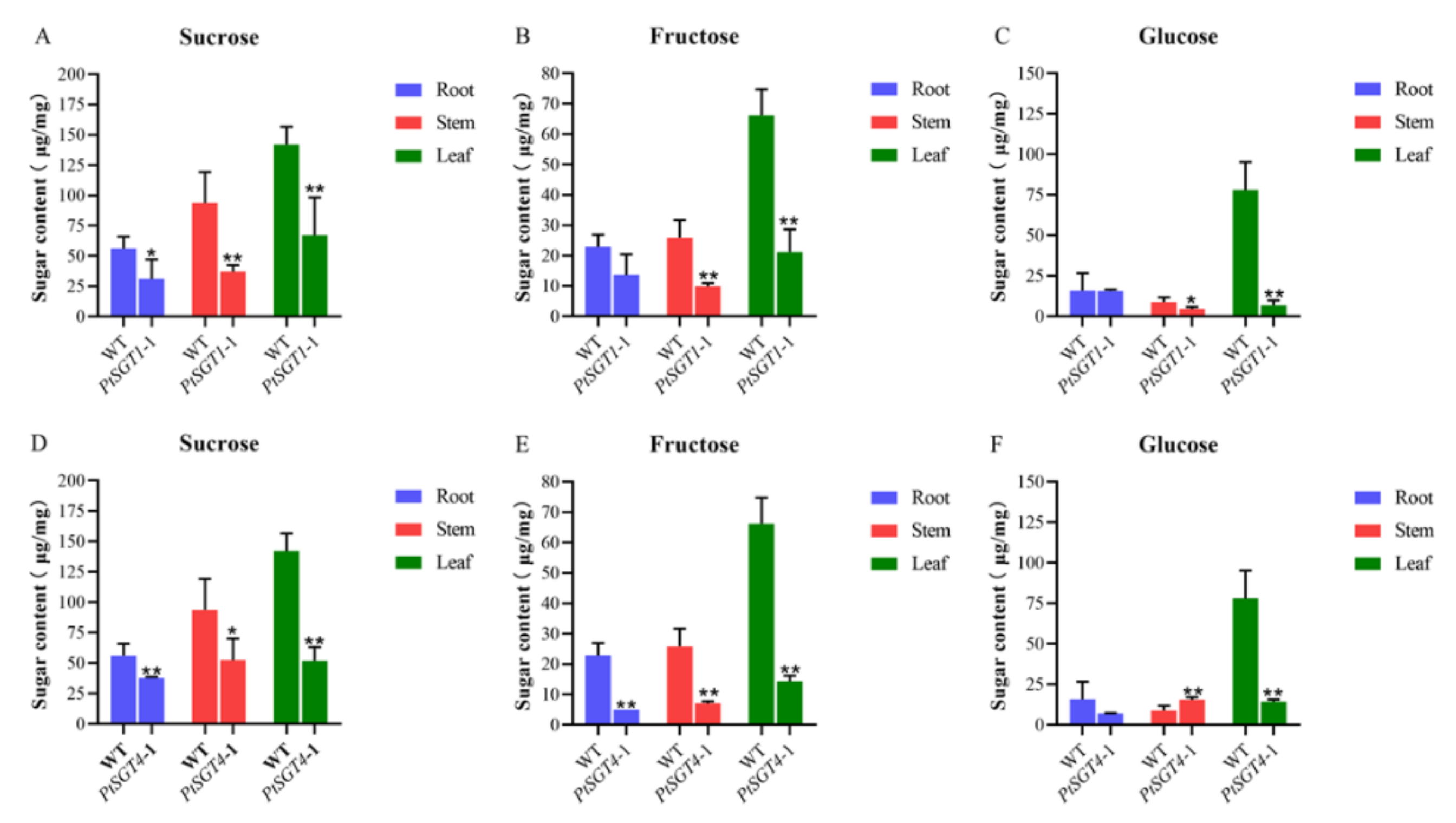
2.4. Sugar Extraction and Measurement
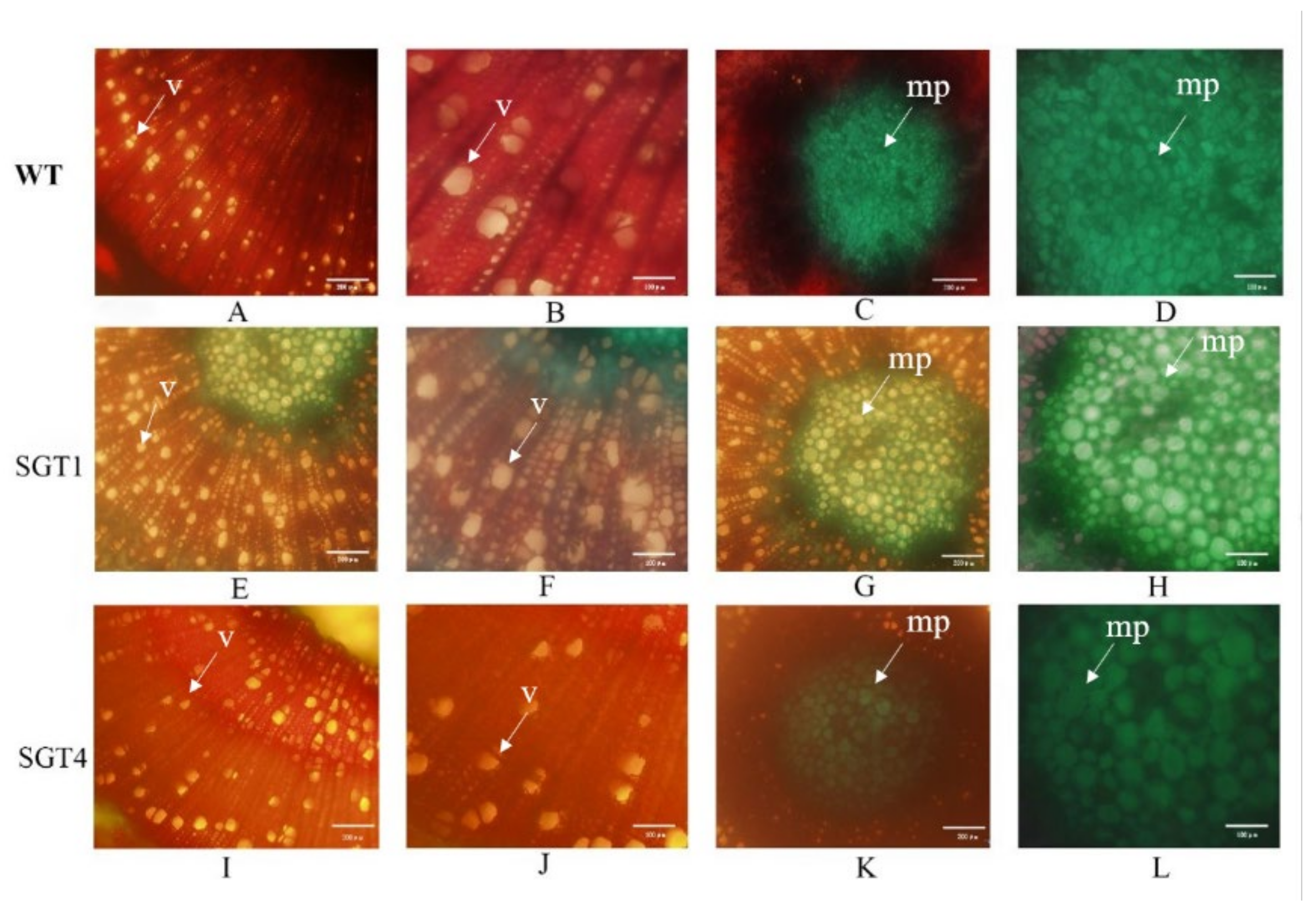
2.5. Sections of Xylem from Transgenic Plants
3. Discussion
4. Materials and Methods
4.1. Experimental Materials
4.2. Cloning and Sequence Analysis of PtSGT
4.3. Construction and Transformation of the PtSGT Vector and Molecular Identification of Transformed P. tomentosa
4.4. Sugar Concentration Measurements in Transgenic Line Tissues
4.5. Sections of Xylem from Transgenic Plants
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rohmer, M. The discovery of a mevalonate-independent pathway for isoprenoid biosynthesis in bacteria, algae and higher plants. Nat. Prod. Rep. 1999, 16, 565–574. [Google Scholar] [CrossRef]
- Rog, T.; Pasenkiewicz-Gierula, M.; Vattulainen, I.; Karttunen, M. Ordering effects of cholesterol and its analogues. Biochim. Biophys. Acta 2009, 1788, 97–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khelashvili, G.; Rappolt, M.; Chiu, S.; Pabst, G.; Harries, D. Impact of Sterol Tilt on Membrane Bending Rigidity in Cholesterol and 7DHC-Containing DMPC Membrane. Biophys. J. 2012, 1021, 413a. [Google Scholar] [CrossRef] [Green Version]
- Grille, S.; Zaslawski, A.; Thiele, S.; Plat, J.; Warnecke, D. The functions of steryl glycosides come to those who wait: Recent advances in plants, fungi, bacteria and animals. Prog. Lipid Res. 2010, 49, 262–288. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.K.; Madina, B.R.; Chaturvedi, P.; Sangwan, R.S.; Tuli, R. Molecular cloning and characterization of one member of 3beta-hydroxy sterol glucosyltransferase gene family in Withania somnifera. Arch. Biochem. Biophys. 2007, 460, 48–55. [Google Scholar] [CrossRef]
- Mackenzie, P.I.; Owens, I.S.; Burchell, B.; Bock, K.W.; Bairoch, A.; Belanger, A.; Fournel-Gigleux, S.; Green, M.; Hum, D.W.; Iyanagi, T.; et al. The UDP glycosyltransferase gene superfamily: Recommended nomenclature update based on evolutionary divergence. Pharmacogenetics 1997, 7, 255–269. [Google Scholar] [CrossRef] [PubMed]
- Ullmann, P.; Ury, A.; Rimmele, D.; Benveniste, P.; Bouvier-Nave, P. UDP-glucose sterol beta-D-glucosyltransferase, a plasma membrane-bound enzyme of plants: Enzymatic properties and lipid dependence. Biochimie 1993, 75, 713–723. [Google Scholar] [CrossRef]
- Warnecke, D.C.; Heinz, E. Purification of a Membrane-Bound UDP-Glucose: Sterol [beta]-D-Glucosyltransferase Based on Its Solubility in Diethyl Ether. Plant Physiol. 1994, 105, 1067–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warnecke, D.C.; Baltrusch, M.; Buck, F.; Wolter, F.P.; Heinz, E. UDP-glucose:sterol glucosyltransferase: Cloning and functional expression in Escherichia coli. Plant Mol. Biol. 1997, 35, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Kawagoe, Y.; Hogan, P.; Delmer, D. Sitosterol-beta-glucoside as primer for cellulose synthesis in plants. Science 2002, 295, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Goren, S.; Huber, S.C.; Granot, D. Comparison of a novel tomato sucrose synthase, SlSUS4, with previously described SlSUS isoforms reveals distinct sequence features and differential expression patterns in association with stem maturation. Planta 2011, 233, 1011–1023. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.L.; Llewellyn, D.J.; Furbank, R.T. Suppression of sucrose synthase gene expression represses cotton fiber cell initiation, elongation, and seed development. Plant Cell 2003, 15, 952–964. [Google Scholar] [CrossRef] [Green Version]
- Schrick, K.; Fujioka, S.; Takatsuto, S.; Stierhof, Y.D.; Stransky, H.; Yoshida, S.; Jurgens, G. A link between sterol biosynthesis, the cell wall, and cellulose in Arabidopsis. Plant J. 2004, 38, 227–243. [Google Scholar] [CrossRef]
- Li, X.; Xia, T.; Huang, J.; Guo, K.; Liu, X.; Chen, T.; Xu, W.; Wang, X.; Feng, S.; Peng, L. Distinct biochemical activities and heat shock responses of two UDP-glucose sterol glucosyltransferases in cotton. Plant Sci. 2014, 219, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.C.; Millet, Y.A.; Cheng, Z.; Bush, J.; Ausubel, F.M. Jasmonate signalling in Arabidopsis involves SGT1b-HSP70-HSP90 chaperone complexes. Nat. Plants 2015, 1, 15049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.I.; Noh, E.W.; Kim, H.J.; Shim, D. Overexpression of poplar GSTU51 confers selective tolerance to both mercury and methyl viologen but not to CDNB or cadmium in transgenic poplars. Plant Biotechnol. Rep. 2013, 7, 175–184. [Google Scholar] [CrossRef]
- Sannigrahi, P.; Ragauskas, A.J.; Tuskan, G.A.; Oak Ridge National Lab. ORNL, O.R.T.U. Poplar as a feedstock for biofuels: A review of compositional characteristics. Biofuels Bioprod. Biorefin. 2010, 4, 209–226. [Google Scholar] [CrossRef]
- Tuskan, G.A.; DiFazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar]
- Park, S.; Oh, S.; Han, K.H. Large-scale computational analysis of poplar ESTs reveals the repertoire and unique features of expressed genes in the poplar genome. Mol. Breed. 2004, 14, 429–440. [Google Scholar] [CrossRef]
- Liu, T.; Fan, D.; Ran, L.; Jiang, Y.; Liu, R.; Luo, K. Highly efficient CRISPR/Cas9-mediated targeted mutagenesis of multiple genes in Populus. Hereditas 2015, 37, 1044–1052. [Google Scholar] [PubMed]
- Fan, D.; Liu, T.; Li, C.; Jiao, B.; Li, S.; Hou, Y.; Luo, K. Efficient CRISPR/Cas9-mediated Targeted Mutagenesis in Populus in the First Generation. Sci. Rep. 2015, 5, 12217. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Jacobs, T.B.; Xue, L.; Harding, S.A.; Tsai, C. Exploiting SNPs for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-coumarate: CoA ligase specificity and redundancy. New Phytol. 2015, 208, 298–301. [Google Scholar] [CrossRef]
- Yan, L.S.; Bin, G.; An, X. Cloning and Expression Analysis of Two Sitosterol Glycosyltransferase Genes in Populus tomentosa. Chin. J. Cell Biol. 2018, 40, 1130–1137. [Google Scholar]
- Jiang, W.; Zhou, H.; Bi, H.; Fromm, M.; Yang, B.; Weeks, D.P. Demonstration of CRISPR/Cas9/sgRNA-mediated targeted gene modification in Arabidopsis, tobacco, sorghum and rice. Nucleic Acids Res. 2013, 41, e188. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, Y.; Zhang, D.; Dai, X.; Estelle, M.; Zhao, Y. Auxin binding protein 1 (ABP1) is not required for either auxin signaling or Arabidopsis development. Proc. Natl. Acad. Sci. USA 2015, 112, 2275–2280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowder, L.G.; Zhang, D.; Baltes, N.J.; Paul, J.R.; Tang, X.; Zheng, X.; Voytas, D.F.; Hsieh, T.F.; Zhang, Y.; Qi, Y. A CRISPR/Cas9 Toolbox for Multiplexed Plant Genome Editing and Transcriptional Regulation. Plant Physiol. 2015, 169, 971–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuster, M.; Schweizer, G.; Reissmann, S.; Kahmann, R. Genome editing in Ustilago maydis using the CRISPR-Cas system. Fungal Genet. Biol. 2016, 89, 3–9. [Google Scholar] [CrossRef]
- Andersson, M.; Turesson, H.; Nicolia, A.; Falt, A.; Samuelsson, M.; Hofvander, P. Efficient targeted multiallelic mutagenesis in tetraploid potato (Solanum tuberosum) by transient CRISPR-Cas9 expression in protoplasts. Plant Cell Rep. 2017, 36, 117–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chilcoat, D.; Liu, Z.B.; Sander, J. Use of CRISPR/Cas9 for Crop Improvement in Maize and Soybean. Prog. Mol. Biol. Transl. Sci. 2017, 149, 27–46. [Google Scholar]
- Meng, X.; Hu, X.; Liu, Q.; Song, X.; Gao, C.; Li, J.; Wang, K. Robust genome editing of CRISPR-Cas9 at NAG PAMs in rice. Sci. China Life Sci. 2018, 61, 122–125. [Google Scholar] [CrossRef] [PubMed]
- Svitashev, S.; Young, J.K.; Schwartz, C.; Gao, H.; Falco, S.C.; Cigan, A.M. Targeted Mutagenesis, Precise Gene Editing, and Site-Specific Gene Insertion in Maize Using Cas9 and Guide RNA. Plant Physiol. 2015, 169, 931–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Gao, K.; Lei, B.; Zhou, J.; Guo, T.; An, X. Altered sucrose metabolism and plant growth in transgenic Populus tomentosa with altered sucrose synthase PtSS3. Transgenic Res. 2020, 29, 125–134. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Name of Plasmid | sgRNA Target Sequence (5′→ 3′) |
|---|---|---|
| PtSGT1 | VK005-14-PtSGT1 | CCCATACCTGTTGATGAGTTC |
| PtSGT4 | VK005-14-PtSGT4 | TAGATCCAAGACGGAGAACAGGG |
| Primer | Sequence (5′→3′) |
|---|---|
| Universal detection primer F | TCTTCAAAGTCCCACATCGC (Source: VK005-14) |
| Universal detection primer R | ACGCTAAGGGAATGCTTGTAT (Source: VK005-14) |
| Primer | Sequence (5′→3′) | Purpose |
|---|---|---|
| PtSGT1F | ATGGCGGAGTCGCAGC | Used for PtSGT1 and PtSGT4 amplification |
| PtSGT1R | TCAGGGACAACCAAAACATTTACT | |
| PtSGT4F | ATGGTTAATAATAGATTTGATCATC | |
| PtSGT4R | TTATGAACAGGATAGGCAG | |
| PtSGT1Ft | TATGCTTGTAATACCGATGC | Used for mutation sequencing analysis |
| PtSGT1Rt | TGAGTAGCCAGTCTAACACGAT | |
| PtSGT4Ft | CAGAAGACAAAGTCAGGGAA | |
| PtSGT4Rt | TCTAACAACTCCGAAGCAAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, Y.; Li, S.; Miao, D.; Huang, S.; Guo, B.; Li, S.; An, X.-M. Investigation of PtSGT1 and PtSGT4 Function in Cellulose Biosynthesis in Populus tomentosa Using CRISPR/Cas9 Technology. Int. J. Mol. Sci. 2021, 22, 13200. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413200
Xue Y, Li S, Miao D, Huang S, Guo B, Li S, An X-M. Investigation of PtSGT1 and PtSGT4 Function in Cellulose Biosynthesis in Populus tomentosa Using CRISPR/Cas9 Technology. International Journal of Molecular Sciences. 2021; 22(24):13200. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413200
Chicago/Turabian StyleXue, Yinxuan, Siyan Li, Deyu Miao, Sai Huang, Bin Guo, Shanwen Li, and Xin-Min An. 2021. "Investigation of PtSGT1 and PtSGT4 Function in Cellulose Biosynthesis in Populus tomentosa Using CRISPR/Cas9 Technology" International Journal of Molecular Sciences 22, no. 24: 13200. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413200