
Ascorbate–Glutathione Oxidant Scavengers, Metabolome Analysis and Adaptation Mechanisms of Ion Exclusion in Sorghum under Salt Stress

 ,
,  ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Plant Material and Growth Conditions
2.3. Treatments
2.4. Raising of the Crop
2.5. Sampling
2.6. Physiological Indices
2.7. Na+: K+ Determination
2.8. Ascorbate–Glutathione Redox Pool
2.9. Antioxidant Molecules
2.10. Total Antioxidant Capacity
2.11. Polyphenolic Compounds
2.12. Compatible Osmolytes
2.13. Oxidative Stress Markers
2.14. Polyamines
2.15. Quantitative Real-Time PCR (qPCR) Gene Expression Analysis
2.16. Yield Components
2.17. Statistical Analysis
3. Results
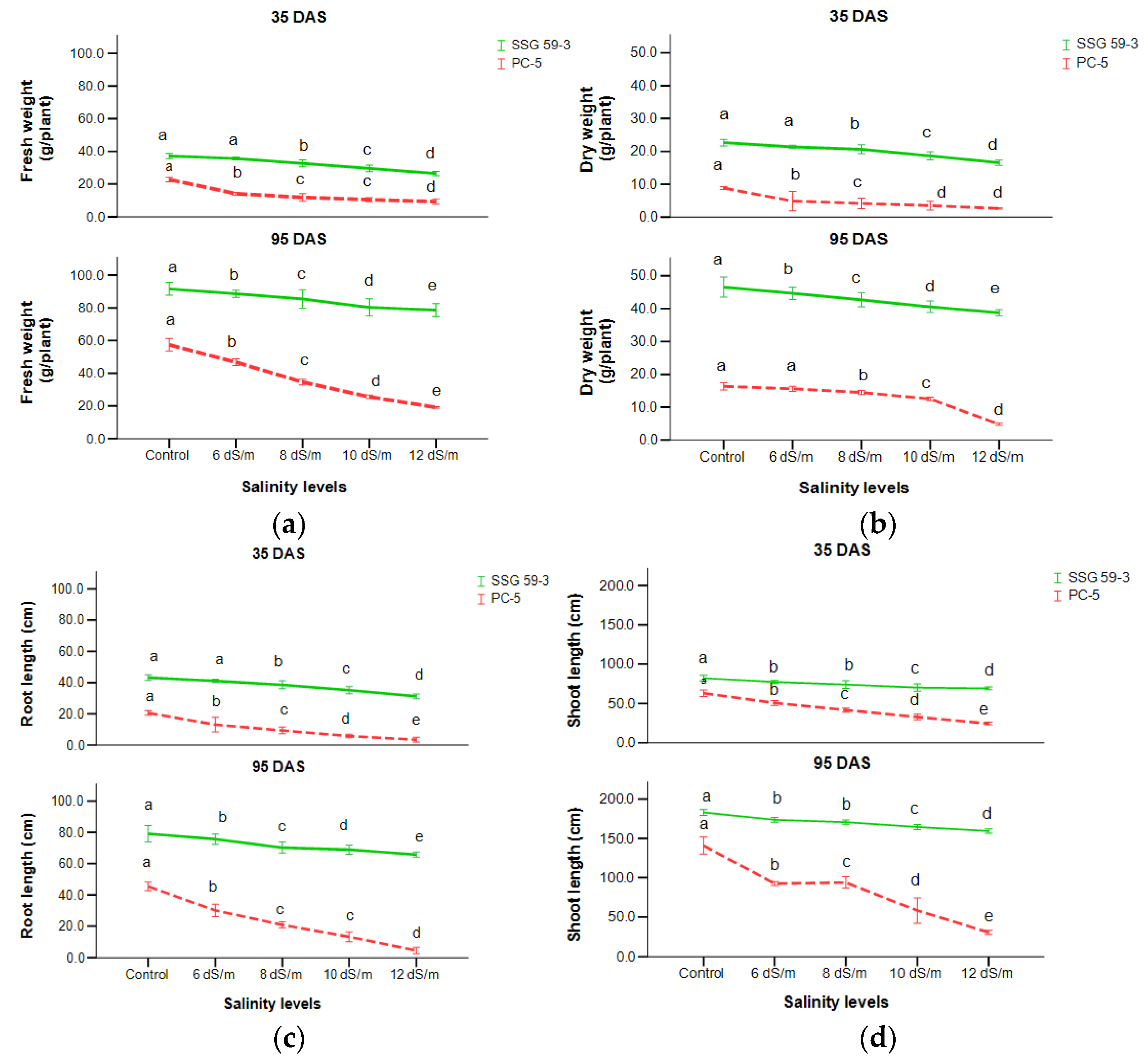
3.1. Biomass Accumulation
3.2. Morpho-Physiological Analysis
3.3. Na+: K+ Determination
3.4. ASC/GSH Cycle Components
3.5. Enzymatic Antioxidants
3.6. Antioxidant Capacity and Polyphenolic Compounds
3.7. Compatible Solutes
3.8. Oxidative Stress Markers
3.9. Polyamines
3.10. Gene Expression Analysis
3.11. Yield Traits
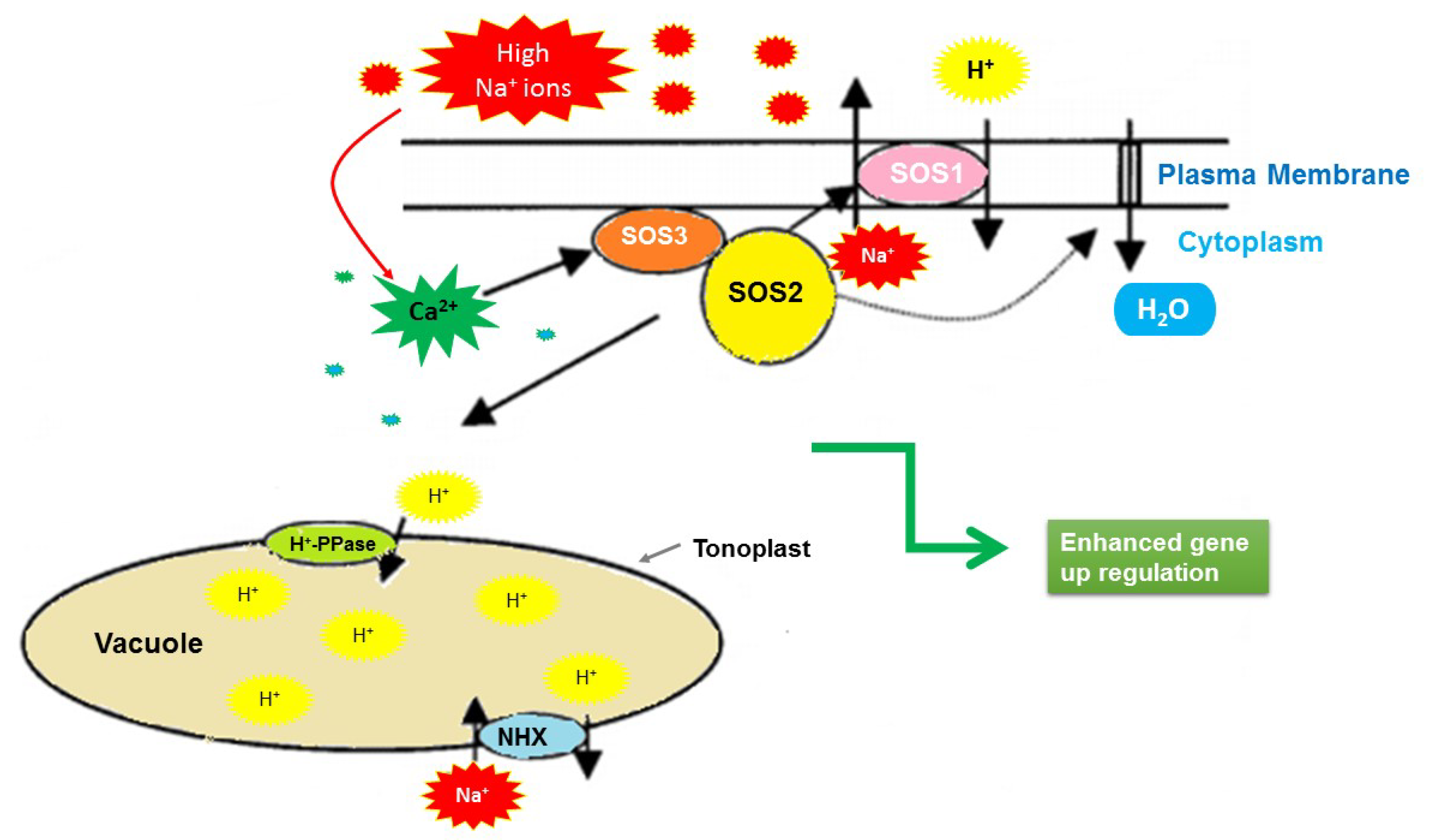
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, X.; Yao, Y.; Li, X.; Zhang, L.; Fan, S. Transcriptomic analysis identifies novel genes and pathways for salt stress responses in Suaeda salsa leaves. Sci. Rep. 2020, 10, 4236. [Google Scholar] [CrossRef]
- Punia, H.; Tokas, J.; Malik, A.; Singh, S.; Phogat, D.S.; Bhuker, A.; Mor, V.S.; Rani, A.; Sheokand, R.N. Discerning morpho-physiological and quality traits contributing to salinity tolerance acquisition in sorghum [Sorghum bicolor (L.) Moench]. S. Afr. J. Bot. 2021, 140, 409–418. [Google Scholar] [CrossRef]
- Corwin, D.L.; Scudiero, E. Review of soil salinity assessment for agriculture across multiple scales using proximal and/or remote sensors. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2019; Volume 158, pp. 1–130. ISBN 0065-2113. [Google Scholar]
- Munns, R.; Passioura, J.B.; Colmer, T.D.; Byrt, C.S. Osmotic adjustment and energy limitations to plant growth in saline soil. New Phytol. 2020, 225, 1091–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shokat, S.; Großkinsky, D.K. Tackling Salinity in Sustainable Agriculture—What Developing Countries May Learn from Approaches of the Developed World. Sustainability 2019, 11, 4558. [Google Scholar] [CrossRef] [Green Version]
- Abreu, I.A.; Farinha, A.P.; Negrão, S.; Gonçalves, N.; Fonseca, C.; Rodrigues, M.; Batista, R.; Saibo, N.J.M.; Oliveira, M.M. Coping with abiotic stress: Proteome changes for crop improvement. J. Proteom. 2013, 93, 145–168. [Google Scholar] [CrossRef]
- Chawla, S.; Jain, S.; Jain, V. Salinity induced oxidative stress and antioxidant system in salt-tolerant and salt-sensitive cultivars of rice (Oryza sativa L.). J. Plant Biochem. Biotechnol. 2013, 22, 27–34. [Google Scholar] [CrossRef]
- Punia, H.; Tokas, J.; Bhadu, S.; Mohanty, A.K.; Rawat, P.; Malik, A. Satpal Proteome dynamics and transcriptome profiling in sorghum [Sorghum bicolor (L.) Moench] under salt stress. 3 Biotech 2020, 10, 412. [Google Scholar] [CrossRef] [PubMed]
- AbdElgawad, H.; Zinta, G.; Hegab, M.M.; Pandey, R.; Asard, H.; Abuelsoud, W. High salinity induces different oxidative stress and antioxidant responses in maize seedlings organs. Front. Plant Sci. 2016, 7, 276. [Google Scholar] [CrossRef] [Green Version]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Punia, H.; Tokas, J.; Malik, A.; Rani, A.; Gupta, P.; Kumari, A.; Mor, V.S.; Bhuker, A.; Kumar, S. Solar Radiation and Nitrogen Use Efficiency for Sustainable Agriculture. In Resources Use Efficiency in Agriculture; Springer: Berlin/Heidelberg, Germany, 2020; pp. 177–212. [Google Scholar]
- Malik, A.; Mor, V.S.; Tokas, J.; Punia, H.; Malik, S.; Malik, K.; Sangwan, S.; Tomar, S.; Singh, P.; Singh, N. Biostimulant-Treated Seedlings under Sustainable Agriculture: A Global Perspective Facing Climate Change. Agronomy 2020, 11, 14. [Google Scholar] [CrossRef]
- Venkateswaran, K.; Elangovan, M.; Sivaraj, N. Origin, domestication and diffusion of Sorghum bicolor. In Breeding Sorghum for Diverse End Uses; Elsevier: Amsterdam, The Netherlands, 2019; pp. 15–31. [Google Scholar]
- USDA–Agricultural Research Service. Germplasm Resources Information Network (GRIN-Taxonomy); USDA: Washington, DC, USA, 2019.
- Wang, Y.-H.; Upadhyaya, H.D.; Kole, C. Genetics, Genomics and Breeding of Sorghum; CRC Press: Boca Raton, FL, USA, 2014; ISBN 1482210088. [Google Scholar]
- Punia, H.; Tokas, J.; Malik, A.; Satpal; Sangwan, S. Characterization of phenolic compounds and antioxidant activity in sorghum [Sorghum bicolor (L.) Moench] grains. Cereal Res. Commun. 2021. [Google Scholar] [CrossRef]
- Punia, H.; Tokas, J.; Malik, A.; Kumar, N. Enzymes as nanoadditives: A promising alternative for biofuel production. In Nanomaterials; Elsevier: Amsterdam, The Netherlands, 2021; pp. 631–662. [Google Scholar]
- Punia, H.; Tokas, J.; Malik, A.; Sangwan, S.; Rani, A.; Yashveer, S.; Alansi, S.; Hashim, M.J.; El-Sheikh, M.A. Genome-Wide Transcriptome Profiling, Characterization, and Functional Identification of NAC Transcription Factors in Sorghum under Salt Stress. Antioxidants 2021, 10, 1605. [Google Scholar] [CrossRef]
- Munns, R.; Gilliham, M. Salinity tolerance of crops—What is the cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotula, L.; Clode, P.L.; Striker, G.G.; Pedersen, O.; Läuchli, A.; Shabala, S.; Colmer, T.D. Oxygen deficiency and salinity affect cell-specific ion concentrations in adventitious roots of barley (Hordeum vulgare). New Phytol. 2015, 208, 1114–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goche, T.; Shargie, N.G.; Cummins, I.; Brown, A.P.; Chivasa, S.; Ngara, R. Comparative physiological and root proteome analyses of two sorghum varieties responding to water limitation. Sci. Rep. 2020, 10, 11835. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef]
- Talbi, S.; Romero-Puertas, M.C.; Hernández, A.; Terrón, L.; Ferchichi, A.; Sandalio, L.M. Drought tolerance in a Saharian plant Oudneya africana: Role of antioxidant defences. Environ. Exp. Bot. 2015, 111, 114–126. [Google Scholar] [CrossRef]
- Kravchik, M.; Bernstein, N. Effects of salinity on the transcriptome of growing maize leaf cells point at cell-age specificity in the involvement of the antioxidative response in cell growth restriction. BMC Genom. 2013, 14, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrs, H.D.; Weatherley, P.E. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Murphy, K.S. Modifications of the technique for determination of chlorophyll stability index in relation to studies of drought resistance in rice. Curr. Sci. 1962, 31, 470–471. [Google Scholar]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Aebi, H. [13] Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Kumar, K.B.; Khan, P.A. Peroxidase and polyphenol oxidase in excised ragi (Eleusine corocana cv PR 202) leaves during senescence. Indian J. Exp. Biol. 1982, 20, 412–416. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Halliwell, B.; Foyer, C.H. Properties and physiological function of a glutathione reductase purified from spinach leaves by affinity chromatography. Planta 1978, 139, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Nagalakshmi, N.; Prasad, M.N. V Responses of glutathione cycle enzymes and glutathione metabolism to copper stress in Scenedesmus bijugatus. Plant Sci. 2001, 160, 291–299. [Google Scholar] [CrossRef]
- Hossain, M.A.; Nakano, Y.; Asada, K. Monodehydroascorbate reductase in spinach chloroplasts and its participation in regeneration of ascorbate for scavenging hydrogen peroxide. Plant Cell Physiol. 1984, 25, 385–395. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Smith, I.K. Stimulation of glutathione synthesis in photorespiring plants by catalase inhibitors. Plant Physiol. 1985, 79, 1044–1047. [Google Scholar] [CrossRef] [Green Version]
- Wellburn, A.R.; Lichtenthaler, H. Formulae and program to determine total carotenoids and chlorophylls a and b of leaf extracts in different solvents. In Advances in Photosynthesis Research; Springer: Berlin/Heidelberg, Germany, 1984; pp. 9–12. [Google Scholar]
- Benzie, I.F.F.; Strain, J.J. [2] Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1999; Volume 299, pp. 15–27. ISBN 0076-6879. [Google Scholar]
- Sompong, R.; Siebenhandl-Ehn, S.; Linsberger-Martin, G.; Berghofer, E. Physicochemical and antioxidative properties of red and black rice varieties from Thailand, China and Sri Lanka. Food Chem. 2011, 124, 132–140. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Swain, T.; Hillis, W.E. The phenolic constituents of Prunus domestica. I.—The quantitative analysis of phenolic constituents. J. Sci. Food Agric. 1959, 10, 63–68. [Google Scholar] [CrossRef]
- Delcour, J.A.; de Varebeke, D.J. A new colourimetric assay for flavanoids in pilsner beers. J. Inst. Brew. 1985, 91, 37–40. [Google Scholar] [CrossRef]
- Johnson, G.; Schaal, L.A. Relation of chlorogenic acid to scab resistance in potatoes. Science 1952, 115, 627–629. [Google Scholar] [CrossRef] [PubMed]
- Balbaa, S.I.; Zaki, A.Y.; El Shamy, A.M. Total Flavonoid and Rutin Content of the Different Organs of Sophora japonica L. J. Assoc. 1974, 57, 752–755. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R. Rapid assay for determination of water soluble quaternary ammonium compounds. Plant Soil 1983, 70, 303–307. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.t.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: II. Role of electron transfer. Arch. Biochem. Biophys. 1968, 125, 850–857. [Google Scholar] [CrossRef]
- Sinha, A.K. Colorimetric assay of catalase. Anal. Biochem. 1972, 47, 389–394. [Google Scholar] [CrossRef]
- Sullivan, C.Y. Selection for drought and heat tolerance in grain sorghum. In Stress Physiology in Crop Plants; Mussell, H., Staples, R.C., Eds.; John Wiley and Sons: New York, NY, USA, 1979; pp. 263–281. [Google Scholar]
- Malik, A.U.; Singh, Z. Endogenous free polyamines of mangos in relation to development and ripening. J. Am. Soc. Hortic. Sci. 2004, 129, 280–286. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 − ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Shabala, S.; Bose, J.; Hedrich, R. Salt bladders: Do they matter? Trends Plant Sci. 2014, 19, 687–691. [Google Scholar] [CrossRef]
- Singh, S.; Duhan, B.S.; Joshi, U.N.; Godara, A.S.; Arya, S. Response of yield, quality and economics of single cut forage sorghum genotypes to different nitrogen and phosphorus levels. Forage Res. 2015, 41, 170–175. [Google Scholar]
- McConnell, J.S.; Francis, P.B.; Stark, C.R.; Glover, R.E. Plant responses of ultra narrow row cotton to nitrogen fertilization. J. Plant Nutr. 2008, 31, 1005–1017. [Google Scholar] [CrossRef]
- Gill, R.A.; Zang, L.; Ali, B.; Farooq, M.A.; Cui, P.; Yang, S.; Ali, S.; Zhou, W. Chromium-induced physio-chemical and ultrastructural changes in four cultivars of Brassica napus L. Chemosphere 2015, 120, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Menconi, M.; Sgherri, C.L.M.; Pinzino, C.; Navari-Lzzo, F. Activated oxygen production and detoxification in wheat plants subjected to a water deficit programme. J. Exp. Bot. 1995, 46, 1123–1130. [Google Scholar] [CrossRef]
- Patel, P.K.; Hemantaranjan, A.; Sarma, B.K.; Radha, S. Growth and antioxidant system under drought stress in chickpea (Cicer arietinum L.) as sustained by salicylic acid. J. Stress Physiol. Biochem. 2011, 7, 130–144. [Google Scholar]
- Siddique, M.R.B.; Hamid, A.; Islam, M.S. Drought stress effects on water relations of wheat. Bot. Bull. Acad. Sin. 2000, 41, 35–39. [Google Scholar]
- Sairam, R.K.; Srivastava, G.C.; Saxena, D.C. Increased antioxidant activity under elevated temperatures: A mechanism of heat stress tolerance in wheat genotypes. Biol. Plant. 2000, 43, 245–251. [Google Scholar] [CrossRef]
- Yang, Z.; Li, J.-L.; Liu, L.-N.; Xie, Q.; Sui, N. Photosynthetic regulation under salt stress and salt-tolerance mechanism of sweet sorghum. Front. Plant Sci. 2020, 10, 1722. [Google Scholar] [CrossRef]
- Netondo, G.W.; Onyango, J.C.; Beck, E. Sorghum and salinity: II. Gas exchange and chlorophyll fluorescence of sorghum under salt stress. Crop Sci. 2004, 44, 806–811. [Google Scholar] [CrossRef]
- Aftab, T.; Khan, M.M.A.; da Silva, J.A.T.; Idrees, M.; Naeem, M. Role of salicylic acid in promoting salt stress tolerance and enhanced artemisinin production in Artemisia annua L. J. Plant Growth Regul. 2011, 30, 425–435. [Google Scholar] [CrossRef]
- Dai, L.Y.; Zhang, L.J.; Jiang, S.J.; Yin, K. De Saline and alkaline stress genotypic tolerance in sweet sorghum is linked to sodium distribution. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2014, 64, 471–481. [Google Scholar]
- Ashraf, U.; Kanu, A.S.; Mo, Z.; Hussain, S.; Anjum, S.A.; Khan, I.; Abbas, R.N.; Tang, X. Lead toxicity in rice: Effects, mechanisms, and mitigation strategies—A mini review. Environ. Sci. Pollut. Res. 2015, 22, 18318–18332. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cao, X.; Zhu, Y.; Yang, X.; Zhang, K.; Xiao, Z.; Wang, H.; Zhao, J.; Zhang, L.; Li, G. Osa-miR398b boosts H2O2 production and rice blast disease-resistance via multiple superoxide dismutases. New Phytol. 2019, 222, 1507–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.-P.; Kim, S.-H.; Bang, J.-W.; Lee, H.-S.; Kwak, S.-S.; Kwon, S.-Y. Enhanced tolerance to oxidative stress in transgenic tobacco plants expressing three antioxidant enzymes in chloroplasts. Plant Cell Rep. 2007, 26, 591–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.; Gupta, N.K. High temperature induced antioxidative defense mechanism in seedlings of contrasting wheat genotypes. Indian J. Plant Physiol. 2005, 10, 73. [Google Scholar]
- Pal, M.; Singh, D.K.; Rao, L.S.; Singh, K.P. Photosynthetic characteristics and activity of antioxidant enzymes in salinity tolerant and sensitive rice cultivars. Indian J. Plant Physiol 2004, 9, 407–412. [Google Scholar]
- Bhattacharjee, S.; Mukherjee, A.K. Heat and salinity induced oxidative stress and changes in protein profile in Amaranthus lividus L. Indian J. Plant Physiol. 2006, 11, 41–47. [Google Scholar]
- de Azevedo Neto, A.D.; Prisco, J.T.; Enéas-Filho, J.; de Abreu, C.E.B.; Gomes-Filho, E. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. Exp. Bot. 2006, 56, 87–94. [Google Scholar] [CrossRef]
- Murthy, S.M.; Devaraj, V.R.; Anitha, P.; Tejavathi, D.H. Studies on the activities of antioxidant enzymes under induced drought stress in in vivo and in vitro plants of Macrotyloma uniflorum (Lam.) Verdc. Recent Res. Sci. Technol. 2012, 4, 34–37. [Google Scholar]
- Randhawa, N.; Kaur, J. Antioxidant responses of Chickpea genotypes exposed to moisture stress. Int. J. Adv. Res. 2015, 3, 950–955. [Google Scholar]
- Sairam, R.K.; Rao, K.V.; Srivastava, G.C. Differential response of wheat genotypes to long term salinity stress in relation to oxidative stress, antioxidant activity and osmolyte concentration. Plant Sci. 2002, 163, 1037–1046. [Google Scholar] [CrossRef]
- Zeeshan, M.; Lu, M.; Sehar, S.; Holford, P.; Wu, F. Comparison of biochemical, anatomical, morphological, and physiological responses to salinity stress in wheat and barley genotypes deferring in salinity tolerance. Agronomy 2020, 10, 127. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Anjum, N.A.; Hasanuzzaman, M.; Gill, R.; Trivedi, D.K.; Ahmad, I.; Pereira, E.; Tuteja, N. Glutathione and glutathione reductase: A boon in disguise for plant abiotic stress defense operations. Plant Physiol. Biochem. 2013, 70, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Wang, Z.; Zhao, Q.; Mao, J.; Speiser, A.; Wirtz, M.; Hell, R.; Zhu, J.; Xiang, C. Sulfate availability affects ABA levels and germination response to ABA and salt stress in Arabidopsis thaliana. Plant J. 2014, 77, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Salama, A.Z.; Lazova, G.N.; Stoinova, Z.G.; Popova, L.P.; El-Fouly, M.M. Effect of zinc deficiency on photosynthesis in chick-pea and maize plants. Comptes Rendus L’academie Bulg. Sci. 2002, 55, 3–65. [Google Scholar]
- Ngo, T.; Zhao, Y. Retaining green pigments on thermally processed peels-on green pears. J. Food Sci. 2005, 70, C568–C574. [Google Scholar] [CrossRef]
- Lau, T.S.L.; Eno, E.; Goldstein, G.; Smith, C.; Christopher, D.A. Ambient levels of UV-B in Hawaii combined with nutrient deficiency decrease photosynthesis in near-isogenic maize lines varying in leaf flavonoids: Flavonoids decrease photoinhibition in plants exposed to UV-B. Photosynthetica 2006, 44, 394–403. [Google Scholar] [CrossRef]
- Dykes, L.; Rooney, L.W.; Waniska, R.D.; Rooney, W.L. Phenolic compounds and antioxidant activity of sorghum grains of varying genotypes. J. Agric. Food Chem. 2005, 53, 6813–6818. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Santhakumar, A.B.; Chinkwo, K.A.; Wu, G.; Johnson, S.K.; Blanchard, C.L. Characterization of phenolic compounds and antioxidant activity in sorghum grains. J. Cereal Sci. 2018, 84, 103–111. [Google Scholar] [CrossRef]
- Shen, S.; Huang, R.; Li, C.; Wu, W.; Chen, H.; Shi, J.; Chen, S.; Ye, X. Phenolic compositions and antioxidant activities differ significantly among sorghum grains with different applications. Molecules 2018, 23, 1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Johnson, S.K.; Bornman, J.F.; Bennett, S.J.; Clarke, M.W.; Singh, V.; Fang, Z. Growth temperature and genotype both play important roles in sorghum grain phenolic composition. Sci. Rep. 2016, 6, 21835. [Google Scholar] [CrossRef] [PubMed]
- Khoddami, A.; Truong, H.H.; Liu, S.Y.; Roberts, T.H.; Selle, P.H. Concentrations of specific phenolic compounds in six red sorghums influence nutrient utilisation in broiler chickens. Anim. Feed Sci. Technol. 2015, 210, 190–199. [Google Scholar] [CrossRef]
- Qureshi, M.I.; Abdin, M.Z.; Ahmad, J.; Iqbal, M. Effect of long-term salinity on cellular antioxidants, compatible solute and fatty acid profile of Sweet Annie (Artemisia annua L.). Phytochemistry 2013, 95, 215–223. [Google Scholar] [CrossRef]
- Anjum, S.A.; Xie, X.; Wang, L.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Pottosin, I.; Bonales-Alatorre, E.; Shabala, S. Choline but not its derivative betaine blocks slow vacuolar channels in the halophyte Chenopodium quinoa: Implications for salinity stress responses. FEBS Lett. 2014, 588, 3918–3923. [Google Scholar] [CrossRef]
- Matysik, J.; Alia; Bhalu, B.; Mohanty, P. Molecular mechanisms of quenching of reactive oxygen species by proline under stress in plants. Curr. Sci. 2002, 525–532. [Google Scholar]
- Arafa, A.A.; Khafagy, M.A.; El-Banna, M.F. The effect of glycinebetaine or ascorbic acid on the salt-stress induced damages in sorghum plant cells. Int. J. Bot. 2007, 3, 251–259. [Google Scholar] [CrossRef]
- Murata, N.; Mohanty, P.S.; Hayashi, H.; Papageorgiou, G.C. Glycinebetaine stabilizes the association of extrinsic proteins with the photosynthetic oxygen-evolving complex. FEBS Lett. 1992, 296, 187–189. [Google Scholar] [CrossRef] [Green Version]
- Saneoka, H.; Ishiguro, S.; Moghaieb, R.E.A. Effect of salinity and abscisic acid on accumulation of glycinebetaine and betaine aldehyde dehydrogenase mRNA in Sorghum leaves (Sorghum bicolor). J. Plant Physiol. 2001, 158, 853–859. [Google Scholar] [CrossRef]
- Gill, P.K.; Sharma, A.D.; Singh, P.; Bhullar, S.S. Changes in germination, growth and soluble sugar contents of Sorghum bicolor (L.) Moench seeds under various abiotic stresses. Plant Growth Regul. 2003, 40, 157–162. [Google Scholar] [CrossRef]
- Bailly, C.; El-Maarouf-Bouteau, H.; Corbineau, F. From intracellular signaling networks to cell death: The dual role of reactive oxygen species in seed physiology. C. R. Biol. 2008, 331, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Motos, J.R.; Penella, C.; Hernández, J.A.; Díaz-Vivancos, P.; Sánchez-Blanco, M.J.; Navarro, J.M.; Gómez-Bellot, M.J.; Barba-Espín, G. Towards a sustainable agriculture: Strategies involving phytoprotectants against salt stress. Agronomy 2020, 10, 194. [Google Scholar] [CrossRef] [Green Version]
- Forghani, A.H.; Almodares, A.; Ehsanpour, A.A. The role of gibberellic acid and paclobutrazol on oxidative stress responses induced by in vitro salt stress in sweet sorghum. Russ. J. Plant Physiol. 2020, 67, 555–563. [Google Scholar] [CrossRef]
- Selote, D.S.; Khanna-Chopra, R. Drought-induced spikelet sterility is associated with an inefficient antioxidant defence in rice panicles. Physiol. Plant. 2004, 121, 462–471. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R.S. Drought induces oxidative stress and enhances the activities of antioxidant enzymes in growing rice seedlings. Plant Growth Regul. 2005, 46, 209–221. [Google Scholar] [CrossRef]
- Reddy, P.S.; Jogeswar, G.; Rasineni, G.K.; Maheswari, M.; Reddy, A.R.; Varshney, R.K.; Kishor, P.B.K. Proline over-accumulation alleviates salt stress and protects photosynthetic and antioxidant enzyme activities in transgenic sorghum [Sorghum bicolor (L.) Moench]. Plant Physiol. Biochem. 2015, 94, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Flores, H.E.; Galston, A.W. Analysis of polyamines in higher plants by high performance liquid chromatography. Plant Physiol. 1982, 69, 701–706. [Google Scholar] [CrossRef] [Green Version]
- Fraga, M.F.; Rodríguez, R.; Cañal, M.J. Reinvigoration of Pinus radiata is associated with partial recovery of juvenile-like polyamine concentrations. Tree Physiol. 2003, 23, 205–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slocum, R.D.; Flores, H.E.; Galston, A.W.; Weinstein, L.H. Improved method for HPLC analysis of polyamines, agmatine and aromatic monoamines in plant tissue. Plant Physiol. 1989, 89, 512–517. [Google Scholar] [CrossRef]
- Punia, H.; Madan, S.; Malik, A.; Sethi, S.K. Stability analysis for quality attributes in durum wheat (Triticum durum L.) genotypes. Bangladesh J. Bot. 2019, 48, 967–972. [Google Scholar] [CrossRef]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef] [Green Version]
- Knight, H.; Trewavas, A.J.; Knight, M.R. Cold calcium signaling in Arabidopsis involves two cellular pools and a change in calcium signature after acclimation. Plant Cell 1996, 8, 489–503. [Google Scholar]
- Shi, H.; Ishitani, M.; Kim, C.; Zhu, J.-K. The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc. Natl. Acad. Sci. USA 2000, 97, 6896–6901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Punia, H.; Tokas, J.; Mor, V.S.; Bhuker, A.; Malik, A.; Singh, N.; Alsahli, A.A.; Hefft, D.I. Deciphering reserve mobilization, antioxidant potential, and expression analysis of starch synthesis in sorghum seedlings under salt stress. Plants 2021, 10, 2463. [Google Scholar] [CrossRef]
- Tokas, J.; Punia, H.; Malik, A.; Sangwan, S.; Devi, S.; Malik, S. Growth performance, nutritional status, forage yield and photosynthetic use efficiency of sorghum [Sorghum bicolor (L.) Moench] under salt stress. Range Manag. Agrofor 2021, 42, 59–70. [Google Scholar]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession No. | Primers | Sequence (5′-3′) |
|---|---|---|---|
| SOS1 | XM_015763865.2 | Forward | GGCTCAAGGCCAACTGGTAT |
| Reverse | TTTCGGGCCTCCCTATCTCA | ||
| SOS2 | KP330207.1 | Forward | TGCATCAGTACTGTGGCTGG |
| Reverse | ATTTAGAAGCCGCACACGGA | ||
| NHX-2 | EU482408.2 | Forward | CGATGGGTGAACGAGTCCAT |
| Reverse | GTTGCAAAAGTATGTCTGGCA | ||
| V-PPase-11 | GQ469975.1 | Forward | GCTACGGCGACTACCTCATC |
| Reverse | CCTTCGGAGATAGCGTTCTG | ||
| CIPK24 | XM_002438609.2 | Forward | TCTCCAGGAGCCAAGTCATT |
| Reverse | CAAACCATGGGTCTGCTCTT | ||
| PP2A | XM_002448914.2 | Forward | AAAAGGCTGCAGAAACGAAG |
| Reverse | GCTTCAATTGGGGCAGATAA |
| Genotypes | DPPH (mg TE/100 g) | FRAP mg (TE/100 g) | ABTS (mg/100 g) | TPC (mg GAE /100 g) | TFC (mg RE/100 g) | Flavanols (mg/100 g DW) | o-HP (mg/100 g DW) |
|---|---|---|---|---|---|---|---|
| SSG 59-3 | 22.4 ± 0.11 a | 16.4 ± 0.1 a | 43.7 ± 0.23 a | 940.7 ± 6.7 a | 45.3 ± 0.35 a | 0.292 ± 0.052 a | 0.924 ± 0.21 a |
| PC-5 | 9.7 ± 0.07 b | 7.1 ± 0.25 b | 20.2 ± 0.18 b | 291.2 ± 3.8 b | 17.2 ± 0.11 b | 0.117 ± 0.23 b | 0.243 ± 0.05 b |
| 35 DAS | 95 DAS | ||||||
|---|---|---|---|---|---|---|---|
| Genotypes | Treatments | Put * | Spd * | Spm * | Put | Spd | Spm |
| SSG 59-3 | Control | 270.9 ± 1.2 a | 246.8 ± 1.1 e | 15.5 ± 0.98 a | 214.6 ± 1.3 a | 195.3 ± 0.96 e | 11.1 ± 0.12 a |
| 60 mM | 336.5 ± 1.4 b | 271.2 ± 1.2 d | 19.5 ± 0.65 b | 265.9 ± 1.2 b | 228.2 ± 1.3 d | 14.5 ± 0.16 b | |
| 80 mM | 382.5 ± 1.8 c | 287.5 ± 1.4 c | 25.6 ± 0.62 c | 319.6 ± 1.1 c | 245.8 ± 1.4 c | 17.6 ± 0.19 c | |
| 100 mM | 457.6 ± 2.3 d | 314.2 ± 1.9 b | 39.6 ± 0.34 d | 384.7 ± 1.9 d | 294.7 ± 1.5 b | 20.6 ± 0.21 d | |
| 120 mM | 496.4 ± 2.8 e | 357.9 ± 2.0 a | 45.2 ± 0.64 e | 402.6 ± 2.5 e | 320.1 ± 2.3 a | 23.9 ± 0.25 e | |
| PC-5 | Control | 155.7 ± 1.3 a | 123.6 ± 0.68 e | 6.4 ± 0.23 a | 98.6 ± 0.26 a | 88.6 ± 0.67 e | 4.6 ± 0.03 a |
| 60 mM | 191.8 ± 1.2 b | 134.2 ± 1.1 d | 10.8 ± 0.37 b | 110.6 ± 1.3 b | 99.3 ± 0.95 d | 5.7 ± 0.12 b | |
| 80 mM | 233.5 ± 1.4 c | 141.5 ± 1.2 c | 14.6 ± 0.56 c | 138.6 ± 0.98 c | 117.3 ± 1.2 c | 8.5 ± 0.06 c | |
| 100 mM | 297.7 ± 1.5 d | 164.3 ± 1.3 b | 18.6 ± 0.61 d | 180.2 ± 1.2 d | 126.8 ± 1.1 b | 10.5 ± 0.16 d | |
| 120 mM | 324.1 ± 1.7 e | 189.4 ± 1.4 a | 26.6 ± 0.62 e | 213.6 ± 1.4 e | 140.3 ± 1.6 a | 12.7 ± 0.20 e | |
| Stages | 35 DAS | 95 DAS | ||||||
|---|---|---|---|---|---|---|---|---|
| Genotypes | SSG 59-3 | PC-5 | SSG 59-3 | PC-5 | SSG 59-3 | PC-5 | SSG 59-3 | PC-5 |
| Character | Plant Height | Number of Leaves/Plant | Plant Height | Number of Leaves/Plant | ||||
| Control | 77.3 ± 1.2 a | 67.7 ± 2.1 a | 10.1 ± 0.23 a | 6 ± 0.06 a | 190.1 ± 22.6 a | 154.2 ± 11.3 a | 13 ± 0.06 a | 8 ± 0.11 a |
| 60 mM | 72.3 ± 2.3 b | 53.1 ± 1.1 b | 9.1 ± 0.45 b | 5 ± 0.11 b | 176.5 ± 15.6 b | 123.4 ± 13.2 b | 10 ± 0.11 b | 7 ± 0.10 b |
| 80 mM | 67.8 ± 2.4 c | 41.6 ± 1.5 c | 8.2 ± 0.12 c | 4 ± 0.14 c | 158.6 ± 14.2 c | 81.4 ± 13.5 c | 9 ± 0.06 c | 6 ± 0.13 c |
| 100 mM | 57.7 ± 1.6 d | 34.5 ± 1.9 d | 7.3 ± 0.16 d | 4 ± 0.16 c | 145.1 ± 13.3 d | 64.4 ± 14.6 d | 8 ± 0.07 d | 6 ± 0.06 c |
| 120 mM | 49.2 ± 1.9 e | 24.6 ± 1.4 e | 6.1 ± 0.21 e | 3 ± 0.09 d | 128.1 ± 12.6 e | 53.6 ± 12.8 e | 7 ± 0.09 e | 4 ± 0.09 d |
| Physiological Maturity (95 DAS) | ||||
|---|---|---|---|---|
| Genotypes | SSG 59-3 | PC-5 | SSG 59-3 | PC-5 |
| Character | Number of Tiller/Plant | Seed Yield/Plant (g) | ||
| Control | 5.03 ± 0.10 a | 3 ± 0.13 a | 15.25 ± 0.13 a | 9.56 ± 0.11 a |
| 60 mM | 4 ± 0.09 b | 2 ± 0.06 b | 13.26 ± 0.18 b | 7.35 ± 0.09 b |
| 80 mM | 3 ± 0.04 c | 1.73 ± 0.1 c | 12.43 ± 0.16 c | 6.12 ± 0.04 c |
| 100 mM | 2.6 ± 0.05 d | 1.33 ± 0.09 d | 11.32 ± 0.14 d | 5.48 ± 0.16 d |
| 120 mM | 2 ± 0.03 d | 0.95 ± 1.2 e | 9.57 ± 0.16 e | 3.26 ± 0.05 e |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Punia, H.; Tokas, J.; Malik, A.; Bajguz, A.; El-Sheikh, M.A.; Ahmad, P. Ascorbate–Glutathione Oxidant Scavengers, Metabolome Analysis and Adaptation Mechanisms of Ion Exclusion in Sorghum under Salt Stress. Int. J. Mol. Sci. 2021, 22, 13249. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413249
Punia H, Tokas J, Malik A, Bajguz A, El-Sheikh MA, Ahmad P. Ascorbate–Glutathione Oxidant Scavengers, Metabolome Analysis and Adaptation Mechanisms of Ion Exclusion in Sorghum under Salt Stress. International Journal of Molecular Sciences. 2021; 22(24):13249. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413249
Chicago/Turabian StylePunia, Himani, Jayanti Tokas, Anurag Malik, Andrzej Bajguz, Mohamed A. El-Sheikh, and Parvaiz Ahmad. 2021. "Ascorbate–Glutathione Oxidant Scavengers, Metabolome Analysis and Adaptation Mechanisms of Ion Exclusion in Sorghum under Salt Stress" International Journal of Molecular Sciences 22, no. 24: 13249. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413249