
Targeting Non-Replicating Mycobacterium tuberculosis and Latent Infection: Alternatives and Perspectives (Mini-Review)



Abstract
:
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Differences between the Definitions of Latency, Persistence, and Dormancy of Mycobacterium tuberculosis
3. Pathogenesis of Latent Tuberculosis Infection in Humans
4. In Vitro and In Vivo Models to Imitate Mycobacterium tuberculosis Latency
5. Alternatives for Latent Tuberculosis Infection Treatment
6. Early-Stage Compounds Active against Dormant and Non-Replicating Bacilli
6.1. Dormancy Survival Two-Component Regulatory System (DosRST)
6.2. Methionine Aminopeptidase (MetAP)
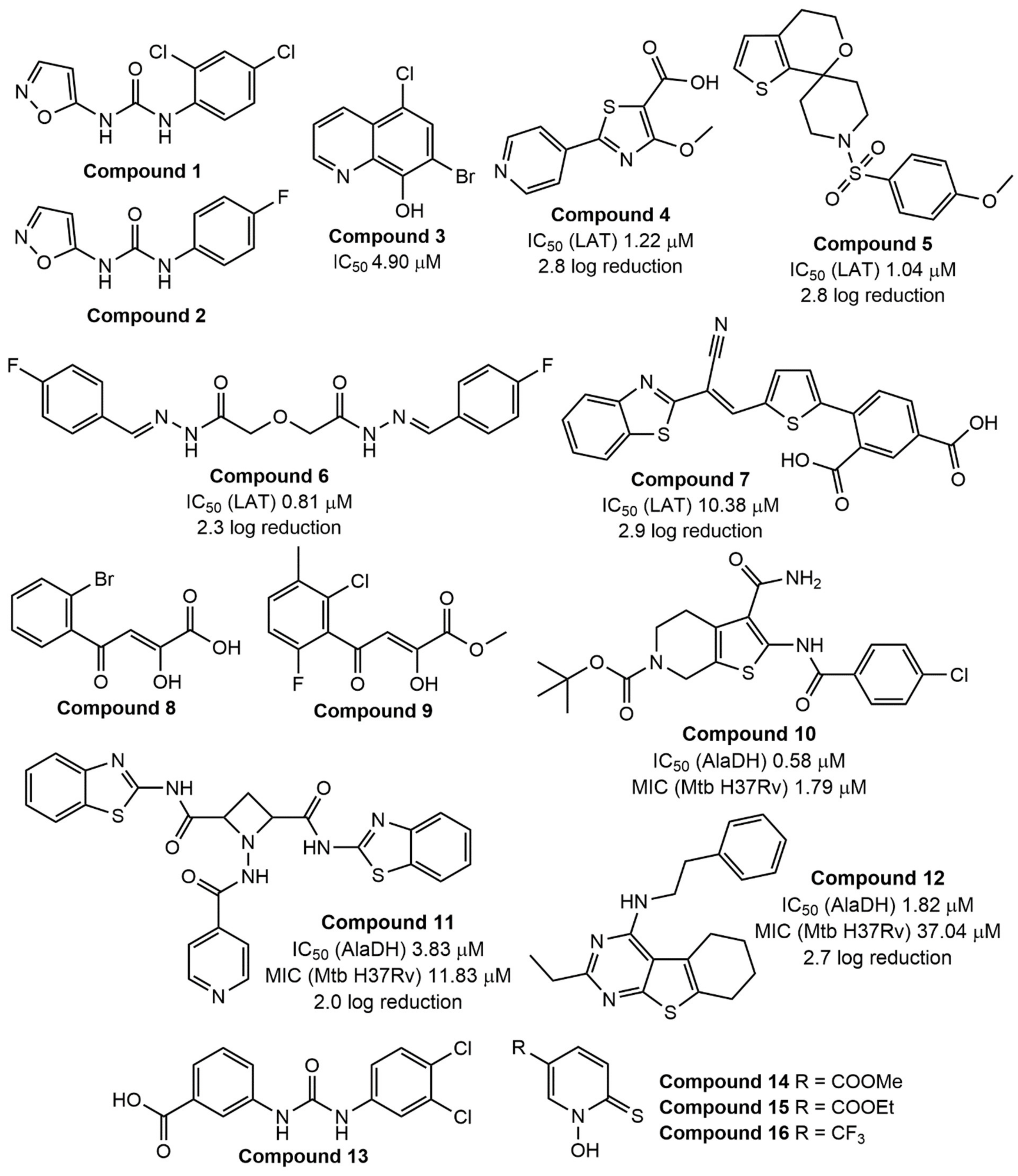
6.3. Lysine ε-Aminotransferase (LAT)
6.4. Isocitrate Lyase (ICL)
6.5. Malate Synthase (GlcB)
6.6. L-Alanine Dehydrogenase (L-AlaDH)
6.7. Cysteine Synthase (CysM)
6.8. Copper-Mediated Innate Immunity
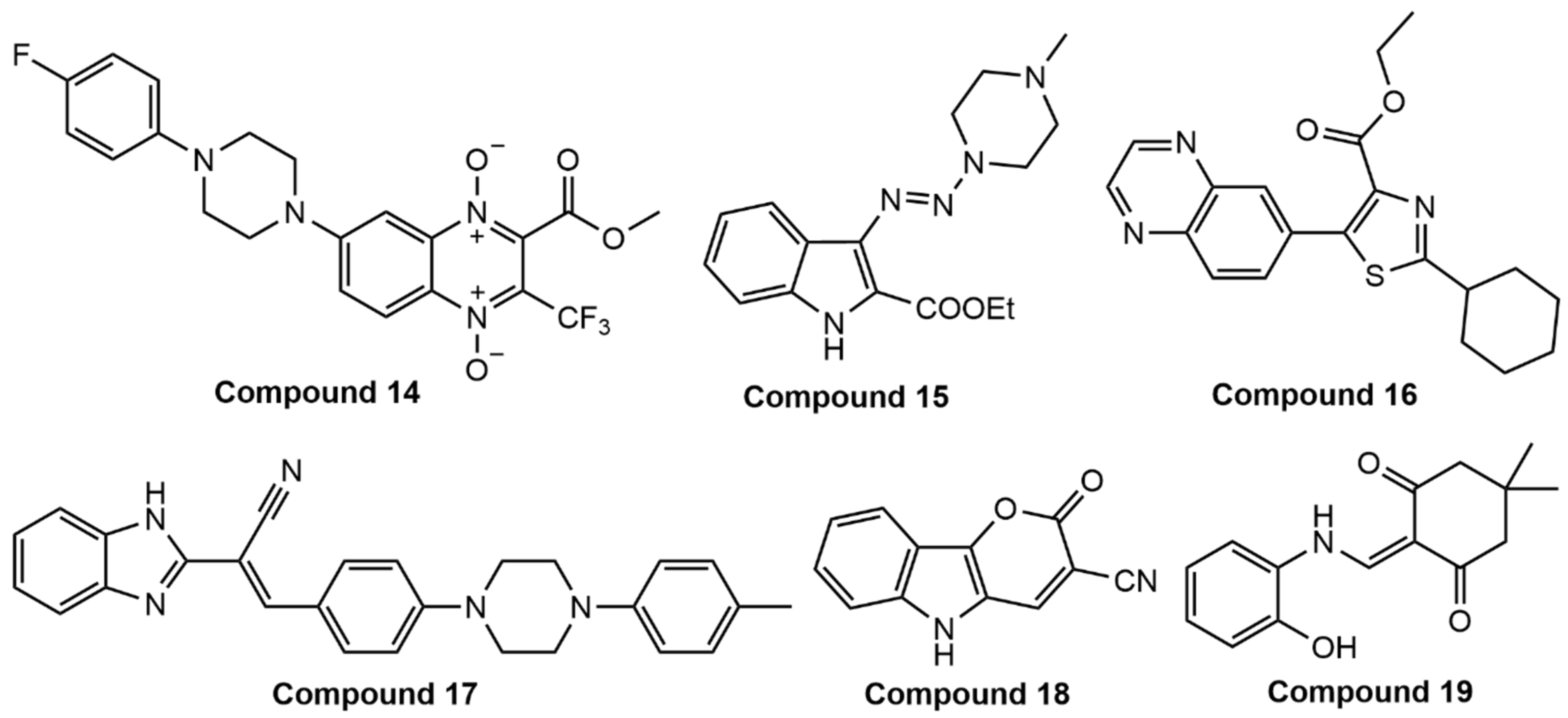
6.9. Unknown Targets
7. Concluding Remarks and Future Outlook
Funding
Conflicts of Interest
References
- World Health Organization. The top 10 causes of death. Available online: Who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 15 September 2021).
- WHO. Global Tuberculosis Report 2020; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- WHO. Global Tuberculosis Report 2019; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- WHO. Global Tuberculosis Report 2018; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Muñoz, L.; Stagg, H.R.; Abubakar, I. Diagnosis and Management of Latent Tuberculosis Infection. Cold Spring Harb. Perspect. Med. 2015, 5, a017830. [Google Scholar] [CrossRef] [Green Version]
- WHO. Latent Tuberculosis Infection—Executive Summary; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Cohen, A.; Mathiasen, V.D.; Schön, T.; Wejse, C. The global prevalence of latent tuberculosis: A systematic review and meta-analysis. Eur. Respir. J. 2019, 54, 1900655. [Google Scholar] [CrossRef]
- WHO. Latent Tuberculosis Infection: Updated and Consolidated Guidelines for Programmatic Management; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Koul, A.; Arnoult, E.; Lounis, N.; Guillemont, J.E.G.; Andries, K. The challenge of new drug discovery for tuberculosis. Nat. Cell Biol. 2011, 469, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Payne, D.J.; Gwynn, M.N.; Holmes, D.J.; Pompliano, D.L. Drugs for bad bugs: Confronting the challenges of antibacterial discovery. Nat. Rev. Drug Discov. 2007, 6, 29–40. [Google Scholar] [CrossRef] [PubMed]
- McCune, R.M.; Tompsett, R. Fate of Mycobacterium tuberculosis in mouse tissues as determined by the microbial enumeration technique. I. The persistence of drug-susceptible tubercle bacilli in the tissues despite prolonged antimicrobial therapy. J. Exp. Med. 1956, 104, 737–762. [Google Scholar] [CrossRef]
- McCune, R.M.; Feldmann, F.M.; Lambert, H.P.; McDermott, W. Microbial persistence. I. The capacity of tubercle bacilli to survive sterilization in mouse tissues. J. Exp. Med. 1966, 123, 445–468. [Google Scholar] [CrossRef]
- Kramnik, I.; Beamer, G. Mouse models of human TB pathology: Roles in the analysis of necrosis and the development of host-directed therapies. Semin. Immunopathol. 2016, 38, 221–237. [Google Scholar] [CrossRef] [Green Version]
- Kesavan, A.K.; Brooks, M.; Tufariello, J.; Chan, J.; Manabe, Y.C. Tuberculosis genes expressed during persistence and reactivation in the resistant rabbit model. Tuberculosis 2009, 89, 17–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenaerts, A.J.; Hoff, D.; Aly, S.; Ehlers, S.; Andries, K.; Cantarero, L.; Orme, I.M.; Basaraba, R.J. Location of persisting mycobacteria in a guinea pig model of tuberculosis revealed by r207910. Antimicrob. Agents Chemother. 2007, 51, 3338–3345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, P.L.; Rodgers, M.; Smith, L.; Bigbee, M.; Myers, A.; Bigbee, C.; Chiosea, I.; Capuano, S.V.; Fuhrman, C.; Klein, E.; et al. Quantitative Comparison of Active and Latent Tuberculosis in the Cynomolgus Macaque Model. Infect. Immun. 2009, 77, 4631–4642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veatch, A.V.; Kaushal, D. Opening Pandora’s Box: Mechanisms of Mycobacterium tuberculosis Resuscitation. Trends Microbiol. 2018, 26, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Wayne, L.G.; Hayes, L.G. An in vitro model for sequential study of shiftdown of Mycobacterium tuberculosis through two stages of nonreplicating persistence. Infect. Immun. 1996, 64, 2062–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Mangan, J.A.; Dhillon, J.; Sole, K.M.; Mitchison, D.A.; Butcher, P.D.; Coates, A.R.M. Detection of mRNA Transcripts and Active Transcription in Persistent Mycobacterium tuberculosis Induced by Exposure to Rifampin or Pyrazinamide. J. Bacteriol. 2000, 182, 6358–6365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betts, J.C.; Lukey, P.T.; Robb, L.C.; McAdam, R.A.; Duncan, K. Evaluation of a nutrient starvation model of Mycobacterium tuberculosis persistence by gene and protein expression profiling. Mol. Microbiol. 2002, 43, 717–731. [Google Scholar] [CrossRef]
- Shleeva, M.O.; Bagramyan, K.; Telkov, M.V.; Mukamolova, G.V.; Young, M.; Kell, D.; Kaprelyants, A.S. Formation and resuscitation of ‘non-culturable’ cells of Rhodococcus rhodochrous and Mycobacterium tuberculosis in prolonged stationary phase. Microbiology 2002, 148, 1581–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deb, C.; Lee, C.-M.; Dubey, V.S.; Daniel, J.; Abomoelak, B.; Sirakova, T.D.; Pawar, S.; Rogers, L.; Kolattukudy, P.E. A Novel In Vitro Multiple-Stress Dormancy Model for Mycobacterium tuberculosis Generates a Lipid-Loaded, Drug-Tolerant, Dormant Pathogen. PLoS ONE 2009, 4, e6077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sala, C.; Dhar, N.; Hartkoorn, R.; Zhang, M.; Ha, Y.H.; Schneider, P.; Cole, S.T. Simple Model for Testing Drugs against Nonreplicating Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2010, 54, 4150–4158. [Google Scholar] [CrossRef] [Green Version]
- Shleeva, M.O.; Kudykina, Y.K.; Vostroknutova, G.N.; Suzina, N.E.; Mulyukin, A.L.; Kaprelyants, A.S. Dormant ovoid cells of Mycobacterium tuberculosis are formed in response to gradual external acidification. Tuberculosis 2011, 91, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Salina, E.; Ryabova, O.; Kaprelyants, A.; Makarov, V. New 2-Thiopyridines as Potential Candidates for Killing both Actively Growing and Dormant Mycobacterium tuberculosis Cells. Antimicrob. Agents Chemother. 2013, 58, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Carranza, C.; Pedraza-Sanchez, S.; de Oyarzabal-Mendez, E.; Torres, M. Diagnosis for latent tuberculosis infection: New alternatives. Front. Immunol. 2020, 11, 2006. [Google Scholar] [CrossRef]
- Shulman, S.T. Clemens von Pirquet: A Remarkable Life and Career. J. Pediatr. Infect. Dis. Soc. 2016, 6, 376–379. [Google Scholar] [CrossRef] [Green Version]
- Bigger, J.W. Treatment of staphylococcal infections with penicillin by intermittent sterilization. Lancet 1944, 244, 497–500. [Google Scholar] [CrossRef]
- McDedmott, W. Microbial persistence. Yale J. Biol. Med. 1958, 30, 257–291. [Google Scholar]
- Gold, B.; Nathan, C. Targeting Phenotypically Tolerant Mycobacterium tuberculosis. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lillebaek, T.; Dirksen, A.; Vynnycky, E.; Baess, I.; Thomsen, V.Ø.; Andersen, Å.B. Stability of DNA Patterns and Evidence of Mycobacterium tuberculosis Reactivation Occurring Decades after the Initial Infection. J. Infect. Dis. 2003, 188, 1032–1039. [Google Scholar] [CrossRef] [Green Version]
- Colangeli, R.; Arcus, V.L.; Cursons, R.T.; Ruthe, A.; Karalus, N.; Coley, K.; Manning, S.D.; Kim, S.; Marchiano, E.; Alland, D. Whole genome sequencing of Mycobacterium tuberculosis reveals slow growth and low mutation rates during latent infections in humans. PLoS ONE 2014, 9, e91024. [Google Scholar]
- Dhar, N.; McKinney, J.; Manina, G. Phenotypic Heterogeneity in Mycobacterium tuberculosis. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S. Pathogenesis, Immunology, and Diagnosis of Latent Mycobacterium tuberculosis Infection. Clin. Dev. Immunol. 2011, 2011, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardona, P.J.; Ruiz-Manzano, J. On the nature of Mycobacterium tuberculosis-latent bacilli. Eur. Respir. J. 2004, 24, 1044–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, R.L. The Pathogenesis of Tuberculosis: The Early Infiltrate of Post-primary (Adult Pulmonary) Tuberculosis: A Distinct Disease Entity. Front. Immunol. 2018, 9, 2108. [Google Scholar] [CrossRef] [Green Version]
- Gibson, S.; Harrison, J.; Cox, J. Modelling a Silent Epidemic: A Review of the In Vitro Models of Latent Tuberculosis. Pathogens 2018, 7, 88. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.H.; Warit, S.; Wan, B.; Hwang, C.H.; Pauli, G.F.; Franzblau, S.G. Low-Oxygen-Recovery Assay for High-Throughput Screening of Compounds against Nonreplicating Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2007, 51, 1380–1385. [Google Scholar] [CrossRef] [Green Version]
- Loebel, R.O.; Shorr, E.; Richardson, H.B. The Influence of Foodstuffs upon the Respiratory Metabolism and Growth of Human Tubercle Bacilli. J. Bacteriol. 1933, 26, 139–166. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Sala, C.; Hartkoorn, R.C.; Dhar, N.; Mendoza-Losana, A.; Cole, S.T. Streptomycin-starved Mycobacterium tuberculosis 18b, a drug discovery tool for latent tuberculosis. Antimicrob. Agents Chemother. 2012, 56, 5782–5789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, T. Experimental studies on the mechanism of infection and immunity in tuberculosis from the analytical standpoint of streptomycin-dependent tubercle bacilli. 1. Isolation and biological characteristics of a streptomycin-dependent mutant, and effect of streptomycin administration on its pathogenicity in guinea pigs. Kekkaku 1955, 30, 4–8. [Google Scholar]
- Yang, H.J.; Wang, D.; Wen, X.; Weiner, D.M.; Via, L.E. One size fits all? Not in in vivo modeling of tuberculosis chemotherapeutics. Front. Cell. Infect. Microbiol. 2021, 11, 613149. [Google Scholar] [CrossRef] [PubMed]
- Manabe, Y.C.; Bishai, W.R. Latent Mycobacterium tuberculosis–persistence, patience and winning by waiting. Nat. Med. 2000, 6, 1327–1329. [Google Scholar] [CrossRef] [PubMed]
- Karakousis, P.C.; Yoshimatsu, T.; Lamichhane, G.; Woolwine, S.C.; Nuermberger, E.L.; Grosset, J.; Bishai, W.R. Dormancy Phenotype Displayed by Extracellular Mycobacterium tuberculosis within Artificial Granulomas in Mice. J. Exp. Med. 2004, 200, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Nuermberger, E.L. Preclinical Efficacy Testing of New Drug Candidates. Microbiol. Spectr. 2017, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Flynn, J.L.; Capuano, S.V.; Croix, D.; Pawar, S.; Myers, A.; Zinovik, A.; Klein, E. Non-human primates: A model for tuberculosis research. Tuberculosis 2003, 83, 116–118. [Google Scholar] [CrossRef]
- Kim, H.W.; Kim, J.S. Treatment of Latent Tuberculosis Infection and Its Clinical Efficacy. Tuberc. Respir. Dis. 2018, 81, 6–12. [Google Scholar] [CrossRef]
- Turetz, M.L.; Ma, K.C. Diagnosis and management of latent tuberculosis. Curr. Opin. Infect. Dis. 2016, 29, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Vilchèze, C.; Jacobs, W.R., Jr. The Isoniazid Paradigm of Killing, Resistance, and Persistence in Mycobacterium tuberculosis. J. Mol. Biol. 2019, 431, 3450–3461. [Google Scholar] [CrossRef] [PubMed]
- Sterling, T.R.; Villarino, M.E.; Borisov, A.S.; Shang, N.; Gordin, F.; Bliven-Sizemore, E.; Hackman, J.; Hamilton, C.D.; Menzies, D.; Kerrigan, A.; et al. Three months of rifapentine and isoniazid for latent tuberculosis infection. N. Engl. J. Med. 2011, 365, 2155–2166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andries, K.; Verhasselt, P.; Guillemont, J.; Göhlmann, H.W.; Neefs, J.M.; Winkler, H.; Van Gestel, J.; Timmerman, P.; Zhu, M.; Lee, E.; et al. A diarylquinoline drug active on the ATP synthase of Mycobacterium tuberculosis. Science 2005, 307, 223–227. [Google Scholar]
- Koul, A.; Vranckx, L.; Dendouga, N.; Balemans, W.; Wyngaert, I.V.D.; Vergauwen, K.; Göhlmann, H.W.; Willebrords, R.; Poncelet, A.; Guillemont, J.; et al. Diarylquinolines Are Bactericidal for Dormant Mycobacteria as a Result of Disturbed ATP Homeostasis. J. Biol. Chem. 2008, 283, 25273–25280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, S.P.S.; Alonso, S.; Rand, L.; Dick, T.; Pethe, K. The protonmotive force is required for maintaining ATP homeostasis and viability of hypoxic, nonreplicating Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2008, 105, 11945–11950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tasneen, R.; Li, S.Y.; Peloquin, C.A.; Taylor, D.; Williams, K.N.; Andries, K.; Mdluli, K.E.; Nuermberger, E.L. Sterilizing activity of novel TMC207- and PA-824-containing regimens in a murine model of tuberculosis. Antimicrob. Agents Chemother. 2011, 55, 5485–5492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Li, S.-Y.; Williams, K.N.; Andries, K.; Nuermberger, E.L. Short-Course Chemotherapy with TMC207 and Rifapentine in a Murine Model of Latent Tuberculosis Infection. Am. J. Respir. Crit. Care Med. 2011, 184, 732–737. [Google Scholar] [CrossRef] [Green Version]
- Stover, C.K.; Warrener, P.; VanDevanter, D.; Sherman, D.R.; Arain, T.M.; Langhorne, M.H.; Anderson, S.W.; Towell, J.A.; Yuan, Y.; McMurray, D.N.; et al. A small-molecule nitroimidazopyran drug candidate for the treatment of tuberculosis. Nat. Cell Biol. 2000, 405, 962–966. [Google Scholar] [CrossRef] [PubMed]
- Lenaerts, A.J.; Gruppo, V.; Marietta, K.S.; Johnson, C.M.; Driscoll, D.K.; Tompkins, N.M.; Rose, J.D.; Reynolds, R.C.; Orme, I.M. Preclinical Testing of the Nitroimidazopyran PA-824 for Activity against Mycobacterium tuberculosis in a Series of In Vitro and In Vivo Models. Antimicrob. Agents Chemother. 2005, 49, 2294–2301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Manjunatha, U.; Boshoff, H.I.M.; Ha, Y.H.; Niyomrattanakit, P.; Ledwidge, R.; Dowd, C.S.; Lee, I.Y.; Kim, P.; Zhang, L.; et al. PA-824 Kills Nonreplicating Mycobacterium tuberculosis by Intracellular NO Release. Science 2008, 322, 1392–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, N.K.; Illei, P.B.; Jain, S.K.; Karakousis, P.C. Characterization of a novel necrotic granuloma model of latent tuberculosis infection and reactivation in mice. Am. J. Pathol. 2014, 184, 2045–2055. [Google Scholar] [CrossRef] [Green Version]
- Dutta, N.; Karakousis, P.C. PA-824 is as effective as isoniazid against latent tuberculosis infection in C3HeB/FeJ mice. Int. J. Antimicrob. Agents 2014, 44, 564–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Hashizume, H.; Tomishige, T.; Nakamura, I.; Matsuba, M.; Fujiwara, M.; Kitamoto, R.; Hanaki, E.; Ohba, Y.; Matsumoto, M. Delamanid Kills Dormant Mycobacteria In Vitro and in a Guinea Pig Model of Tuberculosis. Antimicrob. Agents Chemother. 2017, 61, e02402-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidrônio, M.G.S.; Branco, A.P.O.C.; Abbadi, B.L.; Macchi, F.; Silveira, M.D.; Lock, G.D.A.; Costa, T.D.; de Araújo, D.M.; Cibulski, S.; Bizarro, C.V.; et al. Effects of tafenoquine against active, dormant and resistant Mycobacterium tuberculosis. Tuberculosis 2021, 128, 102089. [Google Scholar] [CrossRef] [PubMed]
- Konduri, S.; Pogaku, V.; Prashanth, J.; Krishna, V.S.; Sriram, D.; Basavoju, S.; Behera, J.N.; Rao, K.P. Sacubitril-Based Urea and Thiourea Derivatives as Novel Inhibitors for Anti-Tubercular against Dormant Tuberculosis. ChemistrySelect 2021, 6, 3869–3874. [Google Scholar] [CrossRef]
- Lemieux, M.R.; Siricilla, S.; Mitachi, K.; Eslamimehr, S.; Wang, Y.; Yang, D.; Pressly, J.D.; Kong, Y.; Park, F.; Franzblau, S.; et al. An antimycobacterial pleuromutilin analogue effective against dormant bacilli. Bioorganic Med. Chem. 2018, 26, 4787–4796. [Google Scholar] [CrossRef] [PubMed]
- Dutta, N.K.; Karakousis, C. Latent Tuberculosis Infection: Myths, Models, and Molecular Mechanisms. Microbiol. Mol. Biol. Rev. 2014, 78, 343–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahrt, T.C.; Deretic, V. Mycobacterium tuberculosis signal transduction system required for persistent infections. Proc. Natl. Acad. Sci. USA 2001, 98, 12706–12711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyagi, J.S.; Sharma, D. Signal transduction systems of mycobacteria with special reference to M. tuberculosis. Curr. Sci. 2004, 86, 93–102. [Google Scholar]
- Honaker, R.W.; Leistikow, R.L.; Bartek, I.L.; Voskuil, M.I. Unique Roles of DosT and DosS in DosR Regulon Induction and Mycobacterium tuberculosis Dormancy. Infect. Immun. 2009, 77, 3258–3263. [Google Scholar] [CrossRef] [Green Version]
- Sivaramakrishnan, S.; Ortiz de Montellano, P.R. The DosS-DosT/DosR mycobacterial sensor system. Biosensors 2013, 3, 259–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.-J.; Park, K.-J.; Ko, I.-J.; Kim, Y.M.; Oh, J.-I. Different Roles of DosS and DosT in the Hypoxic Adaptation of Mycobacteria. J. Bacteriol. 2010, 192, 4868–4875. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.K.; Thakur, T.; Desiraju, G.R.; Tyagi, J.S. Structure-Based Design of DevR Inhibitor Active against Nonreplicating Mycobacterium tuberculosis. J. Med. Chem. 2009, 52, 6324–6334. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Colvin, C.J.; Johnson, B.K.; Kirchhoff, P.D.; Wilson, M.; Jorgensen-Muga, K.; Larsen, S.D.; Abramovitch, R.B. Inhibitors of Mycobacterium tuberculosis DosRST signaling and persistence. Nat. Chem. Biol. 2017, 13, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, T.; Robert-Baudouy, J. Bacterial aminopeptidases: Properties and functions. FEMS Microbiol. Rev. 1996, 18, 319–344. [Google Scholar] [CrossRef] [PubMed]
- Olaleye, O.; Raghunand, T.R.; Bhat, S.; He, J.; Tyagi, S.; Lamichhane, G.; Gu, P.; Zhou, J.; Zhang, Y.; Grosset, J.; et al. Methionine Aminopeptidases from Mycobacterium tuberculosis as Novel Antimycobacterial Targets. Chem. Biol. 2010, 17, 86–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- John, S.F.; Aniemeke, E.; Ha, N.P.; Chong, C.R.; Gu, P.; Zhou, J.; Zhang, Y.; Graviss, E.A.; Liu, J.O.; Olaleye, O.A. Characterization of 2-hydroxy-1-naphthaldehyde isonicotinoyl hydrazone as a novel inhibitor of methionine aminopeptidases from Mycobacterium tuberculosis. Tuberculosis 2016, 101, S73–S77. [Google Scholar] [CrossRef]
- Olaleye, O.; Raghunand, T.R.; Bhat, S.; Chong, C.; Gu, P.; Zhou, J.; Zhang, Y.; Bishai, W.R.; Liu, J.O. Characterization of clioquinol and analogues as novel inhibitors of methionine aminopeptidases from Mycobacterium tuberculosis. Tuberculosis 2011, 91, S61–S65. [Google Scholar] [CrossRef] [PubMed]
- Ekpenyong, O.; Gao, X.; Ma, J.; Cooper, C.; Nguyen, L.; Olaleye, O.A.; Liang, D.; Xie, H. Pre-clinical pharmacokinetics, tissue distribution and physicochemical studies of CLBQ14, a novel methionine aminopeptidase inhibitor for the treatment of infectious diseases. Drug Des. Dev. Ther. 2020, 14, 1263–1277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, X.; Li, Y.; Du, Q.; Huang, Q.; Guo, S.; Xu, M.; Lin, Y.; Liu, Z.; Xie, J. Mycobacterium lysine ε-aminotransferase is a novel alarmone metabolism related persister gene via dysregulating the intracellular amino acid level. Sci. Rep. 2016, 6, 19695. [Google Scholar] [CrossRef] [PubMed]
- Rittershaus, E.S.; Baek, S.-H.; Krieger, I.V.; Nelson, S.J.; Cheng, Y.-S.; Nambi, S.; Baker, R.E.; Leszyk, J.D.; Shaffer, S.A.; Sacchettini, J.C.; et al. A Lysine Acetyltransferase Contributes to the Metabolic Adaptation to Hypoxia in Mycobacterium tuberculosis. Cell Chem. Biol. 2018, 25, 1495–1505.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devi, P.B.; Sridevi, J.P.; Kakan, S.S.; Saxena, S.; Jeankumar, V.U.; Soni, V.; Anantaraju, H.S.; Yogeeswari, P.; Sriram, D. Discovery of novel lysine ε-aminotransferase inhibitors: An intriguing potential target for latent tuberculosis. Tuberculosis 2015, 95, 786–794. [Google Scholar] [CrossRef] [PubMed]
- Parthiban, B.D.; Saxena, S.; Chandran, M.; Jonnalagadda, P.S.; Yadav, R.; Srilakshmi, R.R.; Perumal, Y.; Dharmarajan, S. Design and development of Mycobacterium tuberculosis lysine ε-aminotransferase inhibitors for latent tuberculosis infection. Chem. Biol. Drug Des. 2016, 87, 265–274. [Google Scholar] [CrossRef]
- Reshma, R.S.; Jeankumar, V.U.; Kapoor, N.; Saxena, S.; Bobesh, K.A.; Vachaspathy, A.R.; Kolattukudy, P.E.; Sriram, D. Mycobacterium tuberculosis lysine-ε-aminotransferase a potential target in dormancy: Benzothiazole based inhibitors. Bioorganic Med. Chem. 2017, 25, 2761–2771. [Google Scholar] [CrossRef] [PubMed]
- Alluri, K.K.; Reshma, R.S.; Suraparaju, R.; Gottapu, S.; Sriram, D. Synthesis and evaluation of 4′,5′-dihydrospiro[piperidine-4,7′-thieno[2,3-c]pyran] analogues against both active and dormant Mycobacterium tuberculosis. Bioorganic Med. Chem. 2018, 26, 1462–1469. [Google Scholar] [CrossRef]
- Höner Zu Bentrup, K.; Miczak, A.; Swenson, D.L.; Russell, D.G. Characterization of activity and expression of isocitrate lyase in Mycobacterium avium and Mycobacterium tuberculosis. J. Bacteriol. 1999, 181, 7161–7167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, T.V.; Murkin, A.; Moynihan, M.M.; Harris, L.; Tyler, P.C.; Shetty, N.; Sacchettini, J.C.; Huang, H.-L.; Meek, T.D. Mechanism-based inactivator of isocitrate lyases 1 and 2 from Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2017, 114, 7617–7622. [Google Scholar] [CrossRef] [Green Version]
- McKinney, J.D.; Höner zu Bentrup, K.; Muñoz-Elías, E.J.; Miczak, A.; Chen, B.; Chan, W.T.; Swenson, D.; Sacchettini, J.C.; Jacobs, W.R., Jr.; Russell, D.G. Persistence of Mycobacterium tuberculosis in macrophages and mice requires the glyoxylate shunt enzyme isocitrate lyase. Nature 2000, 406, 735–738. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Elías, E.J.; McKinney, J.D. Mycobacterium tuberculosis isocitrate lyases 1 and 2 are jointly required for in vivo growth and virulence. Nat. Med. 2005, 11, 638–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gengenbacher, M.; Rao, S.P.S.; Pethe, K.; Dick, T. Nutrient-starved, non-replicating Mycobacterium tuberculosis requires respiration, ATP synthase and isocitrate lyase for maintenance of ATP homeostasis and viability. Microbiology 2009, 156, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.; Sharma, S.; Hoener zu Bentrup, K.; McKinney, J.D.; Russell, D.G.; Jacobs, W.R., Jr.; Sacchettini, J.C. Structure of isocitrate lyase, a persistence factor of Mycobacterium tuberculosis. Nat. Struct. Biol. 2000, 7, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Kwai, B.X.C.; Collins, A.J.; Middleditch, M.J.; Sperry, J.; Bashiri, G.; Leung, I.K.H. Itaconate is a covalent inhibitor of the Mycobacterium tuberculosis isocitrate lyase. RSC Med. Chem. 2020, 12, 57–61. [Google Scholar] [CrossRef]
- Bhusal, R.P.; Bashiri, G.; Kwai, B.X.C.; Sperry, J.; Leung, I.K.H. Targeting isocitrate lyase for the treatment of latent tuberculosis. Drug Discov. Today 2017, 22, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Krátký, M.; Vinšová, J.; Novotná, E.; Mandíková, J.; Wsól, V.; Trejtnar, F.; Ulmann, V.; Stolaříková, J.; Fernandes, S.; Bhat, S.; et al. Salicylanilide derivatives block Mycobacterium tuberculosis through inhibition of isocitrate lyase and methionine aminopeptidase. Tuberculosis 2012, 92, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhou, S.; Deng, Q.; Li, X.; Meng, J.; Guan, Y.; Li, C.; Xiao, C. Identification of a novel inhibitor of isocitrate lyase as a potent antitubercular agent against both active and non-replicating Mycobacterium tuberculosis. Tuberculosis 2016, 97, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Krátký, M.; Janďourek, O.; Baranyai, Z.; Novotná, E.; Stolaříková, J.; Bősze, S.; Vinšová, J. Phenolic N-monosubstituted carbamates: Antitubercular and toxicity evaluation of multi-targeting compounds. Eur. J. Med. Chem. 2019, 181, 111578. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.V.; Mellott, D.M.; Moghadamchargari, Z.; Chen, K.; Krieger, I.; Laganowsky, A.; Sacchettini, J.C.; Meek, T.D. Covalent inactivation of Mycobacterium tuberculosis isocitrate lyase by cis-2,3-epoxy-succinic acid. ACS Chem. Biol. 2021, 16, 463–470. [Google Scholar] [CrossRef]
- Krieger, I.V.; Freundlich, J.S.; Gawandi, V.B.; Roberts, J.P.; Gawandi, V.B.; Sun, Q.; Owen, J.L.; Fraile, M.T.; Huss, S.I.; Lavandera, J.-L.; et al. Structure-Guided Discovery of Phenyl-diketo Acids as Potent Inhibitors of M. tuberculosis Malate Synthase. Chem. Biol. 2012, 19, 1556–1567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puckett, S.; Trujillo, C.; Wang, Z.; Eoh, H.; Ioerger, T.R.; Krieger, I.; Sacchettini, J.; Schnappinger, D.; Rhee, K.Y.; Ehrt, S. Glyoxylate detoxification is an essential function of malate synthase required for carbon assimilation in Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2017, 114, E2225–E2232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, J.-A.; Oh, J.-I. Alanine dehydrogenases in mycobacteria. J. Microbiol. 2019, 57, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Giffin, M.M.; Shi, L.; Gennaro, M.L.; Sohaskey, C.D. Role of alanine dehydrogenase of Mycobacterium tuberculosis during recovery from hypoxic nonreplicating persistence. PLoS ONE 2016, 11, e0155522. [Google Scholar] [CrossRef] [PubMed]
- Saxena, S.; Samala, G.; Sridevi, J.P.; Devi, P.B.; Yogeeswari, P.; Sriram, D. Design and development of novel Mycobacterium tuberculosis l-alanine dehydrogenase inhibitors. Eur. J. Med. Chem. 2015, 92, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Reshma, R.S.; Saxena, S.; Bobesh, K.A.; Jeankumar, V.U.; Gunda, S.; Yogeeswari, P.; Sriram, D. Design and development of new class of Mycobacterium tuberculosis l-alanine dehydrogenase inhibitors. Bioorganic Med. Chem. 2016, 24, 4499–4508. [Google Scholar] [CrossRef]
- Samala, G.; Devi, P.B.; Saxena, S.; Gunda, S.; Yogeeswari, P.; Sriram, D. Anti-tubercular activities of 5,6,7,8-tetrahydrobenzo[4,5]thieno[2,3-d ]pyrimidin-4-amine analogues endowed with high activity toward non-replicative Mycobacterium tuberculosis. Bioorganic Med. Chem. 2016, 24, 5556–5564. [Google Scholar] [CrossRef] [PubMed]
- Schnell, R.; Sriram, D.; Schneider, G. Pyridoxal-phosphate dependent mycobacterial cysteine synthases: Structure, mechanism and potential as drug targets. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2015, 1854, 1175–1183. [Google Scholar] [CrossRef] [Green Version]
- Sareen, D.; Newton, G.L.; Fahey, R.C.; Buchmeier, N.A. Mycothiol is essential for growth of Mycobacterium tuberculosis Erdman. J. Bacteriol. 2003, 185, 6736–6740. [Google Scholar] [CrossRef] [PubMed]
- Brunner, K.; Maric, S.; Reshma, R.S.; Almqvist, H.; Seashore-Ludlow, B.; Gustavsson, A.-L.; Poyraz, O.; Yogeeswari, P.; Lundbäck, T.; Vallin, M.; et al. Inhibitors of the Cysteine Synthase CysM with Antibacterial Potency against Dormant Mycobacterium tuberculosis. J. Med. Chem. 2016, 59, 6848–6859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunner, K.; Steiner, E.M.; Reshma, R.S.; Sriram, D.; Schnell, R.; Schneider, G. Profiling of in vitro activities of urea-based inhibitors against cysteine synthases from Mycobacterium tuberculosis. Bioorganic Med. Chem. Lett. 2017, 27, 4582–4587. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Skaar, E.P. Metal limitation and toxicity at the interface between host and pathogen. FEMS Microbiol. Rev. 2014, 38, 1235–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djoko, K.Y.; Goytia, M.M.; Donnelly, P.S.; Schembri, M.A.; Shafer, W.M.; McEwan, A.G. Copper(II)-Bis(Thiosemicarbazonato) Complexes as Antibacterial Agents: Insights into Their Mode of Action and Potential as Therapeutics. Antimicrob. Agents Chemother. 2015, 59, 6444–6453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladomersky, E.; Petris, M.J. Copper tolerance and virulence in bacteria. Metallomics 2015, 7, 957–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neyrolles, O.; Wolschendorf, F.; Mitra, A.; Niederweis, M. Mycobacteria, metals, and the macrophage. Immunol. Rev. 2015, 264, 249–263. [Google Scholar] [CrossRef] [PubMed]
- Rowland, J.; Niederweis, M. Resistance mechanisms of Mycobacterium tuberculosis against phagosomal copper overload. Tuberculosis 2012, 92, 202–210. [Google Scholar] [CrossRef] [Green Version]
- Salina, E.G.; Huszár, S.; Zemanová, J.; Keruchenko, J.; Riabova, O.; Kazakova, E.; Grigorov, A.; Azhikina, T.; Kaprelyants, A.; Mikušová, K.; et al. Copper-related toxicity in replicating and dormant Mycobacterium tuberculosis caused by 1-hydroxy-5-R-pyridine-2(1H)-thiones. Metallomics 2018, 10, 992–1002. [Google Scholar] [CrossRef] [PubMed]
- Macomber, L.; Imlay, J.A. The iron-sulfur clusters of dehydratases are primary intracellular targets of copper toxicity. Proc. Natl. Acad. Sci. USA 2009, 106, 8344–8349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chillappagari, S.; Seubert, A.; Trip, H.; Kuipers, O.P.; Marahiel, M.A.; Miethke, M. Copper Stress Affects Iron Homeostasis by Destabilizing Iron-Sulfur Cluster Formation in Bacillus subtilis. J. Bacteriol. 2010, 192, 2512–2524. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.D.; Kehl-Fie, T.E.; Rosch, J.W. Copper intoxication inhibits aerobic nucleotide synthesis in Streptococcus pneumonia. Metallomics 2015, 7, 786–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santivañez-Veliz, M.; Perez-Silanes, S.; Torres, E.; Moreno-Viguri, E. Design and synthesis of novel quinoxaline derivatives as potential candidates for treatment of multidrug-resistant and latent tuberculosis. Bioorganic Med. Chem. Lett. 2016, 26, 2188–2193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikonenko, B.V.; Kornienko, A.; Majorov, K.; Ivanov, P.; Kondratieva, T.; Korotetskaya, M.; Apt, A.S.; Salina, E.; Velezheva, V. In Vitro Activity of 3-Triazeneindoles against Mycobacterium tuberculosis and Mycobacterium avium. Antimicrob. Agents Chemother. 2016, 60, 6422–6424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karale, U.B.; Krishna, V.S.; Krishna, E.V.; Choudhari, A.S.; Shukla, M.; Gaikwad, V.R.; Mahizhaveni, B.; Chopra, S.; Misra, S.; Sarkar, D.; et al. Synthesis and biological evaluation of 2,4,5-trisubstituted thiazoles as antituberculosis agents effective against drug-resistant tuberculosis. Eur. J. Med. Chem. 2019, 178, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Bonnett, S.A.; Dennison, D.; Files, M.; Bajpai, A.; Parish, T. A class of hydrazones are active against non-replicating Mycobacterium tuberculosis. PLoS ONE 2018, 13, e0198059. [Google Scholar] [CrossRef] [Green Version]
- Sirim, M.M.; Krishna, V.S.; Sriram, D.; Unsal Tan, O. Novel benzimidazole-acrylonitrile hybrids and their derivatives: Design, synthesis and antimycobacterial activity. Eur. J. Med. Chem. 2020, 188, 112010. [Google Scholar] [CrossRef] [PubMed]
- Monakhova, N.; Korduláková, J.; Vocat, A.; Egorova, A.; Lepioshkin, A.; Salina, E.G.; Nosek, J.; Repková, E.; Zemanová, J.; Jurdáková, H.; et al. Design and synthesis of pyrano[3,2-b]indolones showing antimycobacterial activity. ACS Infect. Dis. 2021, 7, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Rather, M.A.; Bhat, Z.S.; Lone, A.M.; Maqbool, M.; Bhat, B.A.; Ahmad, Z. In vitro potency of 2-(((2-hydroxyphenyl)amino)methylene)-5,5-dimethylcyclohexane-1,3-dione against drug-resistant and non-replicating persisters of Mycobacterium tuberculosis. J. Glob. Antimicrob. Resist. 2021, 25, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Krishna, V.S.; Zheng, S.; Rekha, E.M.; Nallangi, R.; Sai Prasad, D.V.; George, S.E.; Guddat, L.W.; Sriram, D. Design and development of ((4-methoxyphenyl)carbamoyl) (5-(5-nitrothiophen-2-yl)-1,3,4-thiadiazol-2-yl)amide analogues as Mycobacterium tuberculosis ketol-acid reductoisomerase inhibitors. Eur. J. Med. Chem. 2020, 193, 112178. [Google Scholar] [CrossRef] [PubMed]
- Konduri, S.; Bhargavi, D.; Prashanth, J.; Krishna, V.S.; Sriram, D.; Rao, K.P. Design and Synthesis of “Chloropicolinate Amides and Urea Derivatives” as Novel Inhibitors for Mycobacterium tuberculosis. ACS Omega 2021, 6, 1657–1667. [Google Scholar] [CrossRef] [PubMed]
- Konduri, S.; Prashanth, J.; Krishna, V.S.; Sriram, D.; Behera, J.N.; Siegel, D.; Rao, K.P. Design and synthesis of purine connected piperazine derivatives as novel inhibitors of Mycobacterium tuberculosis. Bioorganic Med. Chem. Lett. 2020, 30, 127512. [Google Scholar] [CrossRef] [PubMed]
- Verma, J.; Subbarao, N. Designing novel inhibitors against cyclopropane mycolic acid synthase 3 (PcaA): Targeting dormant state of Mycobacterium tuberculosis. J. Biomol. Struct. Dyn. 2021, 39, 6339–6354. [Google Scholar] [CrossRef] [PubMed]
- Hatherill, M.; White, R.G.; Hawn, T.R. Clinical Development of New TB Vaccines: Recent Advances and Next Steps. Front. Microbiol. 2020, 10, 3154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangtani, P.; Abubakar, I.; Ariti, C.; Beynon, R.; Pimpin, L.; Fine, P.E.M.; Rodrigues, L.C.; Smith, P.; Lipman, M.; Whiting, P.; et al. Protection by BCG Vaccine against Tuberculosis: A Systematic Review of Randomized Controlled Trials. Clin. Infect. Dis. 2014, 58, 470–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tait, D.R.; Hatherill, M.; Van Der Meeren, O.; Ginsberg, A.M.; Van Brakel, E.; Salaun, B.; Scriba, T.J.; Akite, E.J.; Ayles, H.M.; Bollaerts, A.; et al. Final analysis of a trial of M72/AS01E vaccine to prevent tuberculosis. N. Engl. J. Med. 2019, 381, 2429–2439. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Bustamante, C.A.; Dedrick, R.M.; Garlena, R.A.; Russell, D.A.; Hatfull, G.F. Toward a phage cocktail for tuberculosis: Susceptibility and tuberculocidal action of mycobacteriophages against diverse Mycobacterium tuberculosis strains. mBio 2021, 12, e00973-21. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Egorova, A.; Salina, E.G.; Makarov, V. Targeting Non-Replicating Mycobacterium tuberculosis and Latent Infection: Alternatives and Perspectives (Mini-Review). Int. J. Mol. Sci. 2021, 22, 13317. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413317
Egorova A, Salina EG, Makarov V. Targeting Non-Replicating Mycobacterium tuberculosis and Latent Infection: Alternatives and Perspectives (Mini-Review). International Journal of Molecular Sciences. 2021; 22(24):13317. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413317
Chicago/Turabian StyleEgorova, Anna, Elena G. Salina, and Vadim Makarov. 2021. "Targeting Non-Replicating Mycobacterium tuberculosis and Latent Infection: Alternatives and Perspectives (Mini-Review)" International Journal of Molecular Sciences 22, no. 24: 13317. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413317