
Combined In Silico, Ex Vivo, and In Vivo Assessment of L-17, a Thiadiazine Derivative with Putative Neuro- and Cardioprotective and Antidepressant Effects

 , , ,
, , , 

Abstract
:1. Introduction
2. Results
2.1. In Silico Predictions
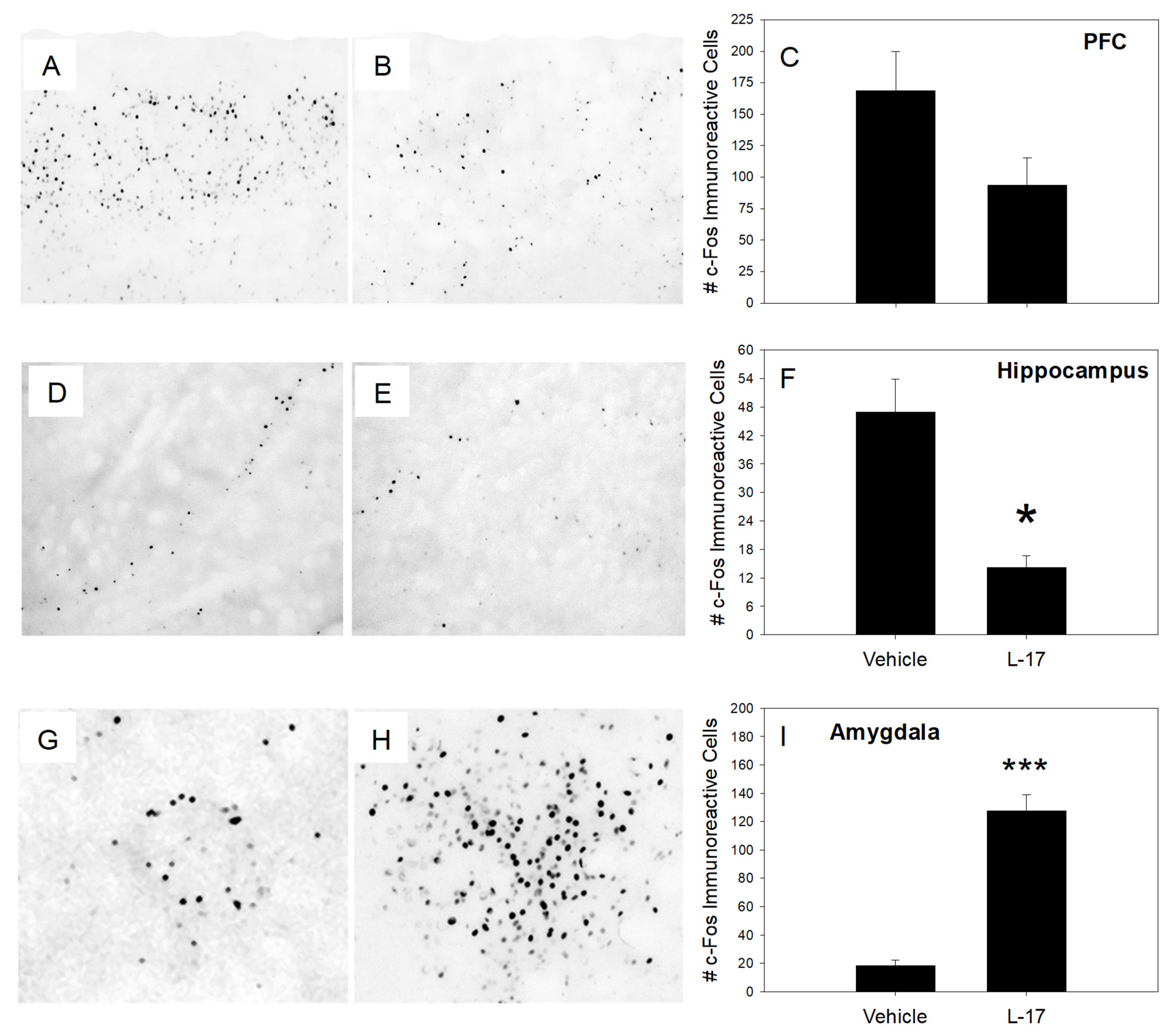
2.2. Proto-Oncogene Fos Expression
2.3. In Vivo Electrophysiology
3. Discussion
4. Materials and Methods
4.1. In Silico Predictions
4.2. Assessment of c-Fos Immunoreactivity
4.3. In Vivo Electrophysiology
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 3D | Three-dimensional |
| 5-HT | 5-hydroxytryptamine (serotonin) |
| 5-HT1AR | 5-HT1A Receptor |
| 5-HT3R | 5-HT3 Receptor |
| ANOVA | Analysis of variance |
| DRN | Dorsal raphe nucleus |
| L-17 | 2-morpholino-5-phenyl-6H-1,3,4-thiadiazine, hydrobromide |
| MI | Myocardial infarction |
| SERT | Serotonin transporter |
| SSRI | Selective serotonin reuptake inhibitor |
References
- Sarapultsev, A.P.; Chupakhin, O.N.; Sarapultsev, P.A.; Sidorova, L.P.; Tseitler, T.A. Pharmacologic Evaluation of Antidepressant Activity and Synthesis of 2-Morpholino-5-phenyl-6H-1,3,4-thiadiazine Hydrobromide. Pharmaceuticals 2016, 9, 27. [Google Scholar] [CrossRef] [Green Version]
- Sarapultsev, P.; Chupakhin, O.; Sarapultsev, A.; Rantsev, M.; Sidorova, L.; Medvedeva, S.; Danilova, I. New insights in to the treatment of myocardial infarction. Int. J. Exp. Pathol. 2012, 93, 18–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarapultsev, A.P.; Chupakhin, O.N.; Sarapultsev, P.A.; Rantsev, M.A.; Medvedeva, S.U.; Sidorova, L.P. Effect of a new class of compounds of the group of substituted 5R1, 6H2-1,3,4-thiadiazine-2-amines on the inflammatory and cytokine response in experimental myocardial infarction. Curr. Vasc. Pharmacol. 2015, 13, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Sarapultsev, A.P.; Chupakhin, O.N.; Sarapultsev, P.A.; Rantsev, M.A.; Medvedeva, S.U.; Sidorova, L.P.; Abidov, M.T.; Danilova, I.G. Modulation of inflammatory response improves myocardial infarct healing in rats. Curr. Pharm. Des. 2014, 20, 1980–1986. [Google Scholar] [CrossRef]
- Sarapultsev, A.; Chupakhin, O.; Rantsev, M.; Sarapultsev, P.; Danilova, I.; Medvedeva, S.; Sidorova, L.; Tseitler, T.; Brilliant, S.; Tseǐlikman, V. Effects of 1,3,4-thiadiazine compound with antidepressant properties in ligation model of acute pancreatitis. Gen. Physiol. Biophys. 2018, 37, 549–562. [Google Scholar] [CrossRef] [PubMed]
- Shevelev, O.B.; Illarionova, N.B.; Petrovski, D.V.; Sarapultsev, A.P.; Chupakhin, O.N.; Moshkin, M.P. Effects of a compound from the group of substituted thiadiazines with hypothermia inducing properties on brain metabolism in rats, a study in vivo and in vitro. PLoS ONE 2017, 12, e0180739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koprdova, R.; Csatlosova, K.; Durisova, B.; Bogi, E.; Majekova, M.; Dremencov, E.; Mach, M. Electrophysiology and Behavioral Assessment of the New Molecule SMe1EC2M3 as a Representative of the Future Class of Triple Reuptake Inhibitors. Molecules 2019, 24, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bullitt, E. Expression of c-fos-like protein as a marker for neuronal activity following noxious stimulation in the rat. J. Comp. Neurol. 1990, 296, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Herdegen, T.; Leah, J.D. Inducible and constitutive transcription factors in the mammalian nervous system: Control of gene expression by Jun, Fos and Krox, and CREB/ATF proteins. Brain Res. Brain Res. Rev. 1998, 28, 370–490. [Google Scholar] [CrossRef]
- Slattery, D.A.; Morrow, J.A.; Hudson, A.L.; Hill, D.R.; Nutt, D.J.; Henry, B. Comparison of alterations in c-Fos and Egr-1 (zif268) expression throughout the rat brain following acute administration of different classes of antidepressant compounds. Neuropsychopharmacology 2005, 30, 1278–1287. [Google Scholar] [CrossRef] [PubMed]
- Koprdova, R.; Osacka, J.; Mach, M.; Kiss, A. Acute Impact of Selected Pyridoindole Derivatives on Fos Expression in Different Structures of the Rat Brain. Cell. Mol. Neurobiol. 2018, 38, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Cohen, B.M.; Cherkerzian, S.; Ma, J.; Ye, N.; Wager, C.; Lange, N. Cells in midline thalamus, central amygdala, and nucleus accumbens responding specifically to antipsychotic drugs. Psychopharmacology 2003, 167, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Morelli, M.; Pinna, A. Antidepressants and atypical neuroleptics induce Fos-like immunoreactivity in the central extended amygdala. Ann. N. Y. Acad. Sci. 1999, 877, 703–706. [Google Scholar] [CrossRef] [PubMed]
- Pinna, A.; Costa, G.; Contu, L.; Morelli, M. Fos expression induced by olanzapine and risperidone in the central extended amygdala. Eur. J. Pharmacol. 2019, 865, 172764. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, C.; Helboe, L. Regional pattern of binding and c-Fos induction by (R)- and (S)-citalopram in rat brain. Neuroreport 2003, 14, 2411–2414. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Miyata, M.; Nagata, T.; Izawa, Y.; Kawakami, Y. Intracerebroventricular fluvoxamine administration inhibited pain behavior but increased Fos expression in affective pain pathways. Pharmacol. Biochem. Behav. 2009, 91, 441–446. [Google Scholar] [CrossRef]
- Jongsma, M.E.; Sebens, J.B.; Bosker, F.J.; Korf, J. Effect of 5-HT1A receptor-mediated serotonin augmentation on Fos immunoreactivity in rat brain. Eur. J. Pharm. 2002, 455, 109–115. [Google Scholar] [CrossRef]
- Robertson, G.S.; Fibiger, H.C. Effects of olanzapine on regional C-Fos expression in rat forebrain. Neuropsychopharmacology 1996, 14, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Sebens, J.B.; Koch, T.; Ter Horst, G.J.; Korf, J. Olanzapine-induced Fos expression in the rat forebrain; cross-tolerance with haloperidol and clozapine. Eur. J. Pharm. 1998, 353, 13–21. [Google Scholar] [CrossRef]
- Kornum, B.R.; Weikop, P.; Moller, A.; Ronn, L.C.; Knudsen, G.M.; Aznar, S. Serotonin depletion results in a decrease of the neuronal activation caused by rivastigmine in the rat hippocampus. Brain Res. 2006, 1073–1074, 262–268. [Google Scholar] [CrossRef]
- Filimonov, D.A.; Lagunin, A.A.; Gloriozova, T.A.; Rudik, A.V.; Druzhilovskii, D.S.; Pogodin, P.V.; Poroikov, V.V. Prediction of the Biological Activity Spectra of Organic Compounds Using the Pass Online Web Resource. Chem. Heterocycl. Compd. 2014, 50, 444–457. [Google Scholar] [CrossRef]
- Inte:Ligand: Your Partner for in Silico Drug Discovery. Available online: http://www.inteligand.com/ (accessed on 15 December 2021).
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amunts, K.; Kedo, O.; Kindler, M.; Pieperhoff, P.; Mohlberg, H.; Shah, N.J.; Habel, U.; Schneider, F.; Zilles, K. Cytoarchitectonic mapping of the human amygdala, hippocampal region and entorhinal cortex: Intersubject variability and probability maps. Anat. Embryol. 2005, 210, 343–352. [Google Scholar] [CrossRef] [PubMed]
- El Mansari, M.; Sánchez, C.; Chouvet, G.; Renaud, B.; Haddjeri, N. Effects of acute and long-term administration of escitalopram and citalopram on serotonin neurotransmission: An in vivo electrophysiological study in rat brain. Neuropsychopharmacology 2005, 30, 1269–1277. [Google Scholar] [CrossRef]
- Dremencov, E.; Csatlosova, K.; Durisova, B.; Moravcikova, L.; Lacinova, L.; Jezova, D. Effect of Physical Exercise and Acute Escitalopram on the Excitability of Brain Monoamine Neur ons: In Vivo Electrophysiological Study in Rats. Int. J. Neuropsychopharmacol. 2017, 20, 585–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Mansari, M.; Blier, P. In vivo electrophysiological assessment of the putative antidepressant Wf-516 in the rat raphe dorsalis, locus coeruleus and hippocampus. Naunyn-Schmiedebergs Arch. Pharmacol. 2008, 376, 351–361. [Google Scholar] [CrossRef]
- Piñeyro, G.; Blier, P.; Dennis, T.; de Montigny, C. Desensitization of the neuronal 5-HT carrier following its long-term blockade. J. Neurosci. Off. J. Soc. Neurosci. 1994, 14 Pt 2, 3036–3047. [Google Scholar] [CrossRef]
- Dremencov, E.; El Mansari, M.; Blier, P. Distinct electrophysiological effects of paliperidone and risperidone on the firing activity of rat serotonin and norepinephrine neurons. Psychopharmacology 2007, 194, 63–72. [Google Scholar] [CrossRef]
- PDBe logo Protein Data Bank in Europe. Available online: https://www.ebi.ac.uk/pdbe/ (accessed on 15 December 2021).
- Coleman, J.A.; Green, E.M.; Gouaux, E. X-ray structures and mechanism of the human serotonin transporter. Nature 2016, 532, 334–339. [Google Scholar] [CrossRef] [Green Version]
- ModBase: Database of Comparative Protein Structure Models. Available online: https://modbase.compbio.ucsf.edu/modbase-cgi/index.cgi (accessed on 15 December 2021).
- Price, K.L.; Lillestol, R.K.; Ulens, C.; Lummis, S.C.R. Palonosetron–5-HT3 Receptor Interactions As Shown by a Binding Protein Cocrystal Structure. ACS Chem. Neurosci. 2016, 7, 1641–1646. [Google Scholar] [CrossRef]
- Jaśkowska, J.; Zaręba, P.; Śliwa, P.; Pindelska, E.; Satała, G.; Majka, Z. Microwave-Assisted Synthesis of Trazodone and Its Derivatives as New 5-HT(1A) Ligands: Binding and Docking Studies. Molecules 2019, 24, 1609. [Google Scholar] [CrossRef] [Green Version]
- MarvinSketch—Create and Design. Available online: https://chemaxon.com/products/marvin (accessed on 15 December 2021).
- Stewart Computational Chemistry. Available online: http://openmopac.net/ (accessed on 15 December 2021).
- UniProtKB. Available online: https://www.uniprot.org/help/uniprotkb (accessed on 15 December 2021).
- DrugBank Online. Available online: https://go.drugbank.com/ (accessed on 15 December 2021).
- IUPHAR/BPS Guide to PHARMACOLOGY. Available online: https://www.guidetopharmacology.org/ (accessed on 15 December 2021).
- Majercikova, Z.; Weering, H.; Scsukova, S.; Mikkelsen, J.D.; Kiss, A. A new approach of light microscopic immunohistochemical triple-staining: Combination of Fos labeling with diaminobenzidine-nickel and neuropeptides labeled with Alexa488 and Alexa555 fluorescent dyes. Endocr. Regul. 2012, 46, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. Paxino’s and Watson’s The Rat Brain in Stereotaxic Coordinates, 7th ed.; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Csatlosova, K.; Bogi, E.; Durisova, B.; Grinchii, D.; Paliokha, R.; Moravcikova, L.; Lacinova, L.; Jezova, D.; Dremencov, E. Maternal immune activation in rats attenuates the excitability of monoamine-secreting neurons in adult offspring in a sex-specific way. Eur. Neuropsychopharmacol. 2020, 43, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Grinchii, D.; Paliokha, R.; Tseilikman, V.; Dremencov, E. Inhibition of cytochrome P450 by proadifen diminishes the excitability of brain serotonin neurons in rats. Gen. Physiol. Biophys. 2018, 37, 711–713. [Google Scholar] [CrossRef] [PubMed]
- Vandermaelen, C.P.; Aghajanian, G.K. Electrophysiological and pharmacological characterization of serotonergic dorsal raphe neurons recorded extracellularly and intracellularly in rat brain slices. Brain Res. 1983, 289, 109–119. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | SERT | 5-HT3 | 5-HT1A | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Molecule | ΔE, kcal/mol | pK | RA | ΔE, kcal/mol | pK | RA | ΔE, kcal/mol | pK | RA |
| L-17 | −8.1 | 5.90 | 0.87 | −6.6 | 4.81 | 0.94 | −7.9 | 5.78 | 0.85 |
| Fluoxetine | −9.3 | 6.78 | - | - | - | - | - | - | - |
| Granisetron | - | - | - | −7.0 | 5.10 | - | - | - | - |
| WAY100135 | - | - | - | - | - | - | −9.3 | 6.78 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarapultsev, A.; Vassiliev, P.; Grinchii, D.; Kiss, A.; Mach, M.; Osacka, J.; Balloova, A.; Paliokha, R.; Kochetkov, A.; Sidorova, L.; et al. Combined In Silico, Ex Vivo, and In Vivo Assessment of L-17, a Thiadiazine Derivative with Putative Neuro- and Cardioprotective and Antidepressant Effects. Int. J. Mol. Sci. 2021, 22, 13626. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413626
Sarapultsev A, Vassiliev P, Grinchii D, Kiss A, Mach M, Osacka J, Balloova A, Paliokha R, Kochetkov A, Sidorova L, et al. Combined In Silico, Ex Vivo, and In Vivo Assessment of L-17, a Thiadiazine Derivative with Putative Neuro- and Cardioprotective and Antidepressant Effects. International Journal of Molecular Sciences. 2021; 22(24):13626. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413626
Chicago/Turabian StyleSarapultsev, Alexey, Pavel Vassiliev, Daniil Grinchii, Alexander Kiss, Mojmir Mach, Jana Osacka, Alexandra Balloova, Ruslan Paliokha, Andrey Kochetkov, Larisa Sidorova, and et al. 2021. "Combined In Silico, Ex Vivo, and In Vivo Assessment of L-17, a Thiadiazine Derivative with Putative Neuro- and Cardioprotective and Antidepressant Effects" International Journal of Molecular Sciences 22, no. 24: 13626. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413626