
Additive Behavioral Improvement after Combined Cell Therapy and Rehabilitation Despite Long-Term Microglia Presence in Stroke Rats

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. ADMSCs Suppressed the Proliferation of Peripheral Blood Mononuclear Cells (PBMCs) In Vitro
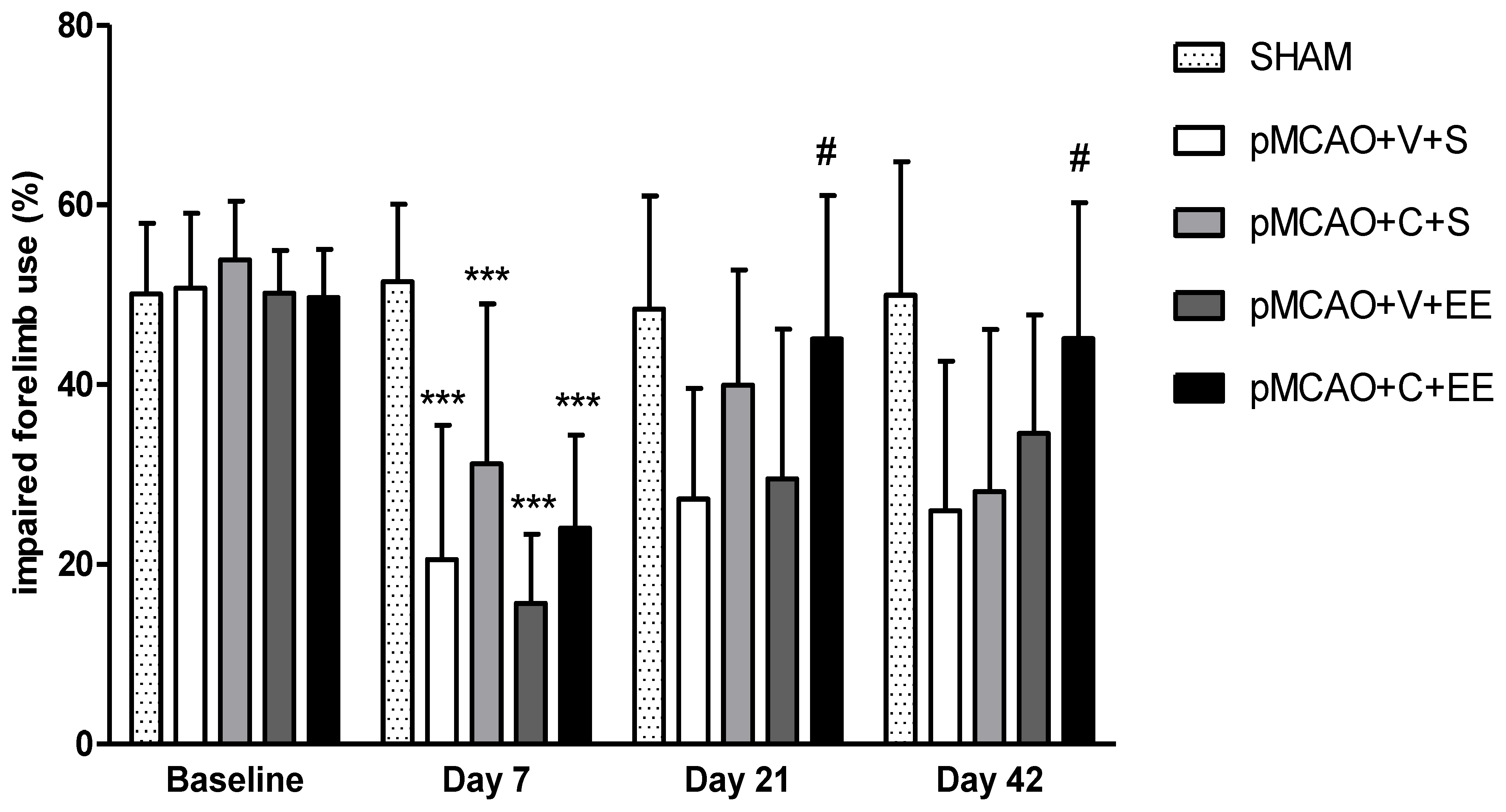
2.2. Combination Therapy Improved the Long-Term Behavioral Outcome
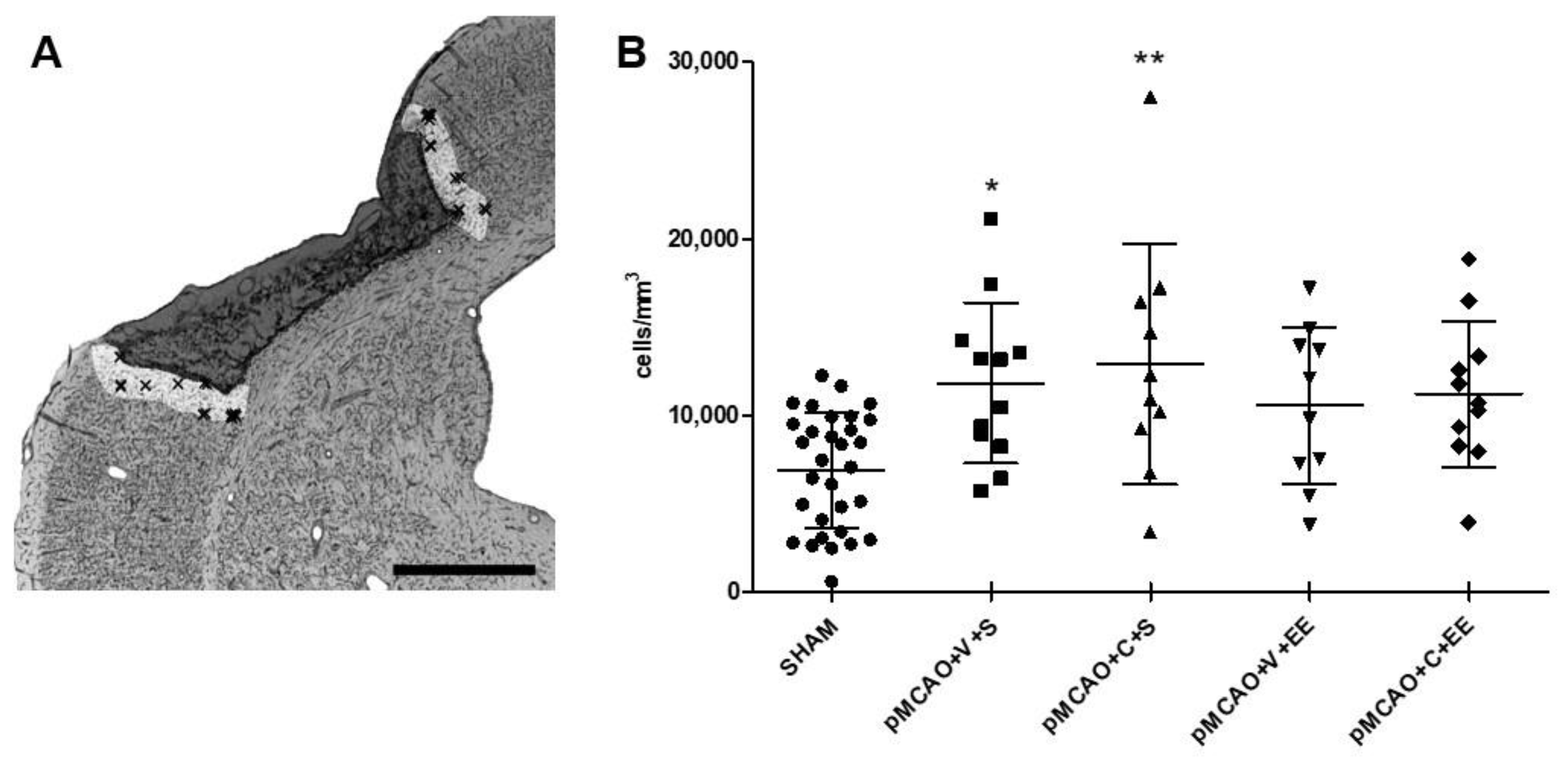
2.3. Stereological Quantification Revealed a Long-Term Increase in the Number of Microglia/Macrophages in the Perilesional Cortex
2.4. ADMSCs with or without Rehabilitation Did Not Change the Staining Intensity of Perilesional Microglia/Macrophages
2.5. No Changes Were Seen in Perilesional Microglia Ramifications after ADMSCs and/or Rehabilitation
2.6. Timing of Cell Delivery Did Not Affect the Long-Term Inflammatory Response
2.7. No Long-Term Correlations between Behavioral Outcome and Microglia Response
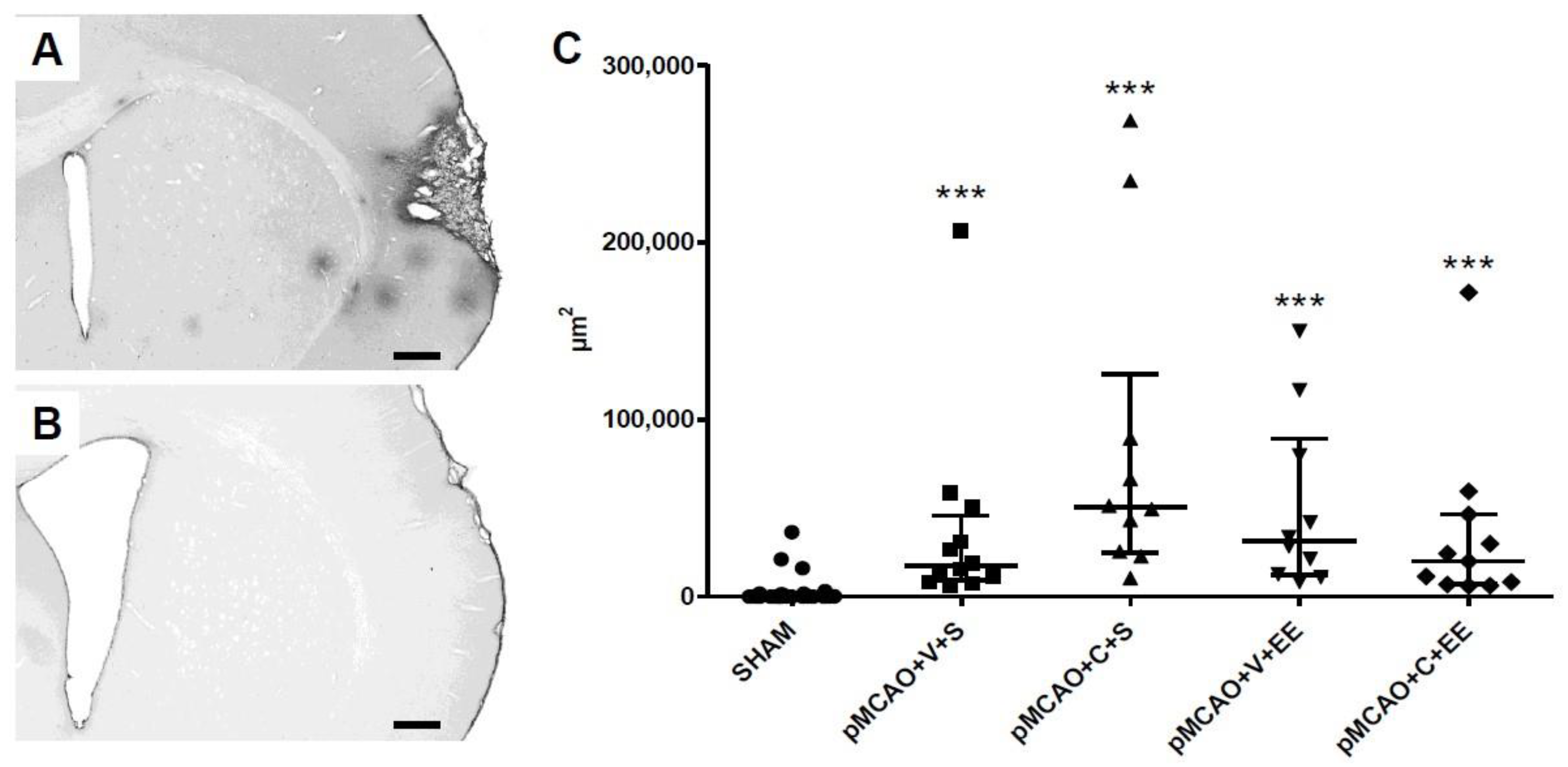
2.8. The Extravasation of Serum Immunoglobulin G (IgG) into Perilesional Cortex Was Increased in pMCAO Rats
3. Discussion
3.1. Time Course of Microglia Behavior
3.2. Effect of Intravenous Cell Infusion
3.3. Effect of Rehabilitation
3.4. Combined Cell Therapy and Enriched Environment
3.5. Long-Term Perilesional BBB Leakage in pMCAO Rats
3.6. Limitations of the Study
4. Materials and Methods
4.1. Animals
4.2. Isolation, Culturing, and Immunophenotyping of Human ADMSCs
4.3. Assessment of Immunomodulatory Potential of ADMSCs In Vitro
4.4. In Vivo Study Design
4.5. Behavioral Testing
4.6. Histology
4.6.1. Iba1 Immunohistochemistry
4.6.2. IgG Immunohistochemistry
4.7. Histological Analyses
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADMSCs | Adipose tissue-derived mesenchymal stem cells |
| ANOVA | Analysis of variance |
| BBB | Blood–brain barrier |
| BMMSCs | Bone-marrow-derived mesenchymal stem cells |
| BrdU | Bromodeoxyuridine |
| CCA | Common carotid artery |
| CD68 | Cluster of differentiation 68 |
| DAB | 3,3′-Diaminobenzidine |
| EE | Enriched environment |
| ELISA | Enzyme-linked immunosorbent assay |
| H2O2 | Hydrogen peroxide |
| Iba1 | Ionized calcium-binding adaptor molecule 1 |
| IgG | Immunoglobulin G |
| IQR | Interquartile range |
| MCA | Middle cerebral artery |
| MLR | Mixed lymphocyte reaction |
| MRI | Magnetic resonance imaging |
| NGS | Normal goat serum |
| PB | Phosphate buffer |
| PBMC | Peripheral blood mononuclear cell |
| PFA | Paraformaldehyde |
| pMCAO | Permanent middle cerebral artery occlusion |
| PX | Passages |
| RESSTORE | Regenerative Stem Cell Therapy for Stroke in Europe |
| RT | Room temperature |
| SD | Standard deviation |
| SHAM | Sham-operated |
| STEPS 4 | Stem Cells as an Emerging Paradigm in Stroke 4 |
| S-HRP | Streptavidin–horseradish peroxidase |
| TBS-T | Tris buffered saline + Triton |
| TLR | Toll-like receptor |
References
- Schroeter, M.; Jander, S.; Witte, O.W.; Stoll, G. Heterogeneity of the microglial response in photochemically induced focal ischemia of the rat cerebral cortex. Neuroscience 1999, 89, 1367–1377. [Google Scholar] [CrossRef]
- Taylor, R.A.; Sansing, L.H. Microglial responses after ischemic stroke and intracerebral hemorrhage. Clin. Dev. Immunol. 2013, 2013, 746068. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.R.; Ritzel, R.; McCullough, L.D.; Liu, F. Microglia and ischemic stroke, a double-edged sword. Int J. Physiol Pathophysiol Pharmacol. 2013, 5, 73–90. [Google Scholar] [PubMed]
- Pandya, R.S.; Mao, L.; Zhou, H.; Zhou, S.; Zeng, J.; Popp, A.J.; Wang, X. Central Nervous System Agents for Ischemic Stroke, Neuroprotection Mechanisms. Cent. Nerv. Syst. Agents Med. Chem. 2011, 11, 81–97. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Kim, N.; Yenari, M.A. Mechanisms and potential therapeutic applications of microglial activation after brain injury. CNS Neurosci. Ther. 2015, 21, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Ritzel, R.M.; Patel, A.R.; Grenier, J.M.; Crapser, J.; Verma, R.; Jellison, E.R.; McCullough, L.D. Functional differences between microglia and monocytes after ischemic stroke. J. Neuroinflamm. 2015, 12, 106. [Google Scholar] [CrossRef] [Green Version]
- Samary, C.S.; Ramos, A.B.; Maia, L.A.; Rocha, N.N.; Santos, C.L.; Magalhães, R.F.; Clevelario, A.L.; Pimentel-Coelho, P.M.; Mendez-Otero, R.; Cruz, F.F.; et al. Focal ischemic stroke leads to lung injury and reduces alveolar macrophage phagocytic capability in rats. Crit. Care 2018, 22, 249. [Google Scholar] [CrossRef] [Green Version]
- Kolosowska, N.; Keuters, M.H.; Wojciechowski, S.; Keksa-Goldsteine, V.; Laine, M.; Malm, T.; Goldsteins, G.; Koistinaho, J.; Dhungana, H. Peripheral administration of IL-13 induces anti-inflammatory microglial/macrophage responses and provides neuroprotection in Ischemic Stroke. Neurotherapeutics 2019, 16, 1304–1319. [Google Scholar] [CrossRef] [Green Version]
- Lambertsen, K.L.; Finsen, B.; Clausen, B.H. Post-stroke inflammation—target or tool for therapy? Acta Neuropathol. 2019, 137, 693–714. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.-T.; Wu, W.-F.; Deng, Y.-H.; Ge, J.-W. Modulators of microglia activation and polarization in ischemic stroke (Review). Mol. Med. Rep. 2020, 21, 2006–2018. [Google Scholar] [CrossRef] [Green Version]
- Rawlinson, C.; Jenkins, S.; Thei, L.; Dallas, M.L.; Chen, R. Post-Ischaemic Immunological Response in the Brain: Targeting Microglia in Ischaemic Stroke Therapy. Brain Sci. 2020, 10, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fumagalli, S.; Perego, C.; Pischiutta, F.; Zanier, E.R.; De Simoni, M.-G. The ischemic environment drives microglia and macrophage function. Front. Neurol. 2015, 6, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandvig, I.; Augestad, I.L.; Håberg, A.K.; Sandvig, A. Neuroplasticity in stroke recovery. The role of microglia in engaging and modifying synapses and networks. Eur. J. Neurosci. 2018, 47, 1414–1428. [Google Scholar] [CrossRef] [PubMed]
- Badimon, A.; Strasburger, H.J.; Ayata, P.; Chen, X.; Nair, A.; Ikegami, A.; Hwang, P.; Chan, A.T.; Graves, S.M.; Uweru, J.O.; et al. Negative feedback control of neuronal activity by microglia. Nature 2020, 586, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Stonesifer, C.; Corey, S.; Ghanekar, S.; Diamandis, Z.; Acosta, S.A.; Borlongan, C.V. Stem cell therapy for abrogating stroke-induced neuroinflammation and relevant secondary cell death mechanisms. Prog. Neurobiol. 2017, 158, 94–131. [Google Scholar] [CrossRef]
- Boshuizen, M.C.S.; Steinberg, G.K. Stem Cell-Based Immunomodulation After Stroke: Effects on brain repair processes. Stroke 2018, 49, 1563–1570. [Google Scholar] [CrossRef]
- Patrikoski, M.; Sivula, J.; Huhtala, H.; Helminen, M.; Salo, F.; Mannerström, B.; Miettinen, S. Different culture conditions modulate the immunological properties of adipose stem cells. Stem Cells Transl. Med. 2014, 3, 1220–1230. [Google Scholar] [CrossRef]
- Valencia, J.; Blanco, B.; Yáñez, R.; Vázquez, M.; Herrero Sánchez, C.; Fernández-García, M.; Rodríguez Serrano, C.; Pescador, D.; Blanco, J.F.; Hernando-Rodríguez, M.; et al. Comparative analysis of the immunomodulatory capacities of human bone marrow- and adipose tissue-derived mesenchymal stromal cells from the same donor. Cytotherapy 2016, 18, 1297–1311. [Google Scholar] [CrossRef]
- Schilling, M.; Besselmann, M.; Leonhard, C.; Mueller, M.; Ringelstein, E.B.; Kiefer, R. Microglial activation precedes and predominates over macrophage infiltration in transient focal cerebral ischemia: A study in green fluorescent protein transgenic bone marrow chimeric mice. Exp. Neurol. 2003, 183, 25–33. [Google Scholar] [CrossRef]
- Schilling, M.; Besselmann, M.; Müller, M.; Strecker, J.K.; Ringelstein, E.B.; Kiefer, R. Predominant phagocytic activity of resident microglia over hematogenous macrophages following transient focal cerebral ischemia, an investigation using green fluorescent protein transgenic bone marrow chimeric mice. Exp. Neurol. 2005, 196, 290–297. [Google Scholar] [CrossRef]
- Schroeter, M.; Jander, S.; Huitinga, I.; Witte, O.W.; Stoll, G. Phagocytic response in photochemically induced infarction of rat cerebral cortex. The role of resident microglia. Stroke 1997, 28, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Kawabori, M.; Kacimi, R.; Kauppinen, T.; Calosing, C.; Kim, J.Y.; Hsieh, C.L.; Nakamura, M.C.; Yenari, M.A. Triggering receptor expressed on myeloid cells 2 (TREM2) deficiency attenuates phagocytic activities of microglia and exacerbates ischemic damage in experimental stroke. J. Neurosci. 2015, 35, 3384–3396. [Google Scholar] [CrossRef] [PubMed]
- Kurisu, K.; Zheng, Z.; Kim, J.Y.; Shi, J.; Kanoke, A.; Liu, J.; Hsieh, C.L.; Yenari, M.A. Triggering receptor expressed on myeloid cells-2 expression in the brain is required for maximal phagocytic activity and improved neurological outcomes following experimental stroke. J. Cereb. Blood Flow Metab. 2019, 39, 1906–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thored, P.; Heldmann, U.; Gomes-Leal, W.; Gisler, R.; Darsalia, V.; Taneera, J.; Nygren, J.M.; Jacobsen, S.-E.W.; Ekdahl, C.T.; Kokaia, Z.; et al. Long-term accumulation of microglia with proneurogenic phenotype concomitant with persistent neurogenesis in adult subventricular zone after stroke. Glia 2009, 57, 835–849. [Google Scholar] [CrossRef] [PubMed]
- Ladwig, A.; Walter, H.L.; Hucklenbroich, J.; Willuweit, A.; Langen, K.-J.; Fink, G.R.; Rueger, M.A.; Schroeter, M. Osteopontin Augments M2 Microglia Response and Separates M1- and M2-Polarized Microglial Activation in Permanent Focal Cerebral Ischemia. Mediat. Inflamm. 2017, 2017, 7189421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, H.W.; Filosa, J.A. A quantitative spatiotemporal analysis of microglia morphology during ischemic stroke and reperfusion. J. Neuroinflamm. 2013, 10, 782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekdahl, C.T.; Kokaia, Z.; Lindvall, O. Brain inflammation and adult neurogenesis: The dual role of microglia. Neuroscience 2009, 158, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Patrikoski, M.; Mannerström, B.; Miettinen, S. Perspectives for Clinical Translation of Adipose Stromal/Stem Cells. Stem Cells Int. 2019, 2019, 5858247. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Moisan, A.; Jolkkonen, J. Intravascular cell therapy in stroke: Predicting the future trends. Regen Med. 2019, 14, 63–68. [Google Scholar] [CrossRef]
- Mu, J.; Bakreen, A.; Juntunen, M.; Korhonen, P.; Oinonen, E.; Cui, L.; Myllyniemi, M.; Zhao, S.; Miettinen, S.; Jolkkonen, J. Combined Adipose Tissue-Derived Mesenchymal Stem Cell Therapy and Rehabilitation in Experimental Stroke. Front. Neurol. 2019, 10, 235. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Wei, X.; Li, J.; Heine, L.A.; Rosenwasser, R.; Iacovitti, L. Changes in host blood factors and brain glia accompanying the functional recovery after systemic administration of bone marrow stem cells in ischemic stroke rats. Cell Transpl. 2010, 19, 1073–1084. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huang, M.; Zhao, R.; Zhao, C.; Liu, Y.; Zou, H.; Chen, L.; Guan, Y.; Zhang, Y.A. Intravenously Delivered Allogeneic Mesenchymal Stem Cells Bidirectionally Regulate Inflammation and Induce Neurotrophic Effects in Distal Middle Cerebral Artery Occlusion Rats Within the First 7 Days After Stroke. CPB 2018, 46, 1951–1970. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, J.; Zhang, C.L.; Wang, L.; Lu, D.; Katakowski, M.; Gao, Q.; Shen, L.H.; Zhang, J.; Lu, M.; et al. Gliosis and brain remodeling after treatment of stroke in rats with marrow stromal cells. Glia 2005, 49, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.-H.; Choi, C.; Chang, D.-J.; Shin, D.-A.; Lee, N.; Jeon, I.; Sung, J.-H.; Lee, H.; Hong, K.-S.; Ko, J.J.; et al. Early neuroprotective effect with lack of long-term cell replacement effect on experimental stroke after intra-arterial transplantation of adipose-derived mesenchymal stromal cells. Cytotherapy 2015, 17, 1090–1103. [Google Scholar] [CrossRef] [PubMed]
- Gómez-de Frutos, M.C.; Laso-García, F.; Diekhorst, L.; Otero-Ortega, L.; Fuentes, B.; Jolkkonen, J.; Detante, O.; Moisan, A.; Martínez-Arroyo, A.; Díez-Tejedor, E.; et al. Intravenous delivery of adipose tissue-derived mesenchymal stem cells improves brain repair in hyperglycemic stroke rats. Stem Cell Res. Ther. 2019, 10, 212. [Google Scholar] [CrossRef] [Green Version]
- Mering, S.; Jolkkonen, J. Proper housing conditions in experimental stroke studies-special emphasis on environmental enrichment. Front. Neurosci. 2015, 9, 106. [Google Scholar] [CrossRef] [Green Version]
- Keiner, S.; Wurm, F.; Kunze, A.; Witte, O.W.; Redecker, C. Rehabilitative therapies differentially alter proliferation and survival of glial cell populations in the perilesional zone of cortical infarcts. Glia 2008, 56, 516–527. [Google Scholar] [CrossRef]
- Ma, Y.; He, M.; Qiang, L. Exercise Therapy Downregulates the Overexpression of TLR4, TLR2, MyD88 and NF-κB after Cerebral Ischemia in Rats. Int. J. Mol. Sci. 2013, 14, 3718–3733. [Google Scholar] [CrossRef]
- Quattromani, M.J.; Cordeau, P.; Ruscher, K.; Kriz, J.; Wieloch, T. Enriched housing down-regulates the Toll-like receptor 2 response in the mouse brain after experimental stroke. Neurobiol. Dis. 2014, 66, 66–73. [Google Scholar] [CrossRef]
- Piao, C.-S.; Stoica, B.A.; Wu, J.; Sabirzhanov, B.; Zhao, Z.; Cabatbat, R.; Loane, D.J.; Faden, A.I. Late exercise reduces neuroinflammation and cognitive dysfunction after traumatic brain injury. Neurobiol. Dis. 2013, 54, 252–263. [Google Scholar] [CrossRef] [Green Version]
- Ito, A.; Kubo, N.; Liang, N.; Aoyama, T.; Kuroki, H. Regenerative Rehabilitation for Stroke Recovery by Inducing Synergistic Effects of Cell Therapy and Neurorehabilitation on Motor Function: A Narrative Review of Pre-Clinical Studies. Int. J. Mol. Sci. 2020, 21, 3135. [Google Scholar] [CrossRef] [PubMed]
- Wahl, A.-S.; Schwab, M.E. Finding an optimal rehabilitation paradigm after stroke: Enhancing fiber growth and training of the brain at the right moment. Front. Hum. Neurosci. 2014, 8, 381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, Y.; Li, M.-Z.; Yang, L.; Feng, X.-F.; Lei, J.-F.; Zhang, N.; Zhao, Y.-Y.; Zhao, H. The three-phase enriched environment paradigm promotes neurovascular restorative and prevents learning impairment after ischemic stroke in rats. Neurobiol. Dis. 2020, 146, 105091. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Sasaki, M.; Kataoka-Sasaki, Y.; Nakazaki, M.; Nagahama, H.; Suzuki, J.; Tateyama, D.; Oka, S.; Namioka, T.; Namioka, A.; et al. Synergic Effects of Rehabilitation and Intravenous Infusion of Mesenchymal Stem Cells After Stroke in Rats. Phys. Ther. 2016, 96, 1791–1798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Boer, A.; Storm, A.; Gomez-Soler, M.; Smolders, S.; Rué, L.; Poppe, L.; B Pasquale, E.; Robberecht, W.; Lemmens, R. Environmental enrichment during the chronic phase after experimental stroke promotes functional recovery without synergistic effects of EphA4 targeted therapy. Hum. Mol. Genet. 2020, 29, 605–617. [Google Scholar] [CrossRef] [Green Version]
- Liebigt, S.; Schlegel, N.; Oberland, J.; Witte, O.W.; Redecker, C.; Keiner, S. Effects of rehabilitative training and anti-inflammatory treatment on functional recovery and cellular reorganization following stroke. Exp. Neurol. 2012, 233, 776–782. [Google Scholar] [CrossRef]
- Boltze, J.; Modo, M.M.; Mays, R.W.; Taguchi, A.; Jolkkonen, J.; Savitz, S.I.; STEPS 4 Consortium. Stem cells as an emerging paradigm in Stroke 4, advancing and accelerating preclinical research. Stroke 2019, 50, 3299–3306. [Google Scholar] [CrossRef]
- Rust, R.; Weber, R.Z.; Grönnert, L.; Mulders, G.; Maurer, M.A.; Hofer, A.-S.; Sartori, A.M.; Schwab, M.E. Anti-Nogo-A antibodies prevent vascular leakage and act as pro-angiogenic factors following stroke. Sci. Rep. 2019, 9, 20040. [Google Scholar] [CrossRef]
- Obermeier, B.; Daneman, R.; Ransohoff, R.M. Development, maintenance and disruption of the blood-brain barrier. Nat. Med. 2013, 19, 1584–1596. [Google Scholar] [CrossRef]
- Strbian, D.; Durukan, A.; Pitkonen, M.; Marinkovic, I.; Tatlisumak, E.; Pedrono, E.; Abo-Ramadan, U.; Tatlisumak, T. The blood–brain barrier is continuously open for several weeks following transient focal cerebral ischemia. Neuroscience 2008, 153, 175–181. [Google Scholar] [CrossRef]
- Sladojevic, N.; Stamatovic, S.M.; Johnson, A.M.; Choi, J.; Hu, A.; Dithmer, S.; Blasig, I.E.; Keep, R.F.; Andjelkovic, A.V. Claudin-1-Dependent Destabilization of the Blood–Brain Barrier in Chronic Stroke. J. Neurosci. 2019, 39, 743–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Rosenberg, G.A. Blood-Brain Barrier Breakdown in Acute and Chronic Cerebrovascular Disease. Stroke 2011, 42, 3323–3328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, L.; Song, Z.; Mi, J.; Hou, P.; Xie, C.; Shi, J.; Li, Y.; Manaenko, A. The Effects and Underlying Mechanisms of Cell Therapy on Blood-Brain Barrier Integrity After Ischemic Stroke. Curr. Neuropharmacol. 2020, 18, 1213–1226. [Google Scholar] [CrossRef] [PubMed]
- Heindl, S.; Gesierich, B.; Benakis, C.; Llovera, G.; Duering, M.; Liesz, A. Automated Morphological Analysis of Microglia after Stroke. Front Cell Neurosci. 2018, 12, 106. [Google Scholar] [CrossRef]
- Young, K.; Morrison, H. Quantifying Microglia Morphology from Photomicrographs of Immunohistochemistry Prepared Tissue Using Image. JoVE J. Vis. Exp. 2018, 136, e57648. [Google Scholar]
- Mangin, G.; Cogo, A.; Moisan, A.; Bonnin, P.; Maïer, B.; Kubis, N.; RESSTORE Consortium. Intravenous administration of human adipose derived-mesenchymal stem cells is not efficient in diabetic or hypertensive mice subjected to focal cerebral Ischemia. Front. Neurosci. 2019, 13, 718. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, K.R. Evaluation of cellular and humoral immune responses to allogeneic adipose-derived stem/stromal cells. Methods Mol. Biol. 2011, 702, 133–150. [Google Scholar]
- Corbett, D.; Carmichael, S.T.; Murphy, T.H.; Jones, T.A.; Schwab, M.E.; Jolkkonen, J.; Clarkson, A.N.; Dancause, N.; Weiloch, T.; Johansen-Berg, H.; et al. Enhancing the alignment of the preclinical and clinical stroke recovery research pipeline: Consensus-based core recommendations from the Stroke Recovery and Rehabilitation Roundtable translational working group. Int. J. Stroke 2017, 12, 462–471. [Google Scholar] [CrossRef]
- Geuna, S.; Herrera-Rincon, C. Update on stereology for light microscopy. Cell Tissue Res. 2015, 360, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Santos-Galdiano, M.; Perez-Rodriguez, D.; Anuncibay-Soto, B.; Font-Belmonte, E.; Ugidos, I.F.; Perez-Garcia, C.C.; Fernandez-Lopez, A. Celecoxib treatment improves neurological deficit and reduces selective neuronal loss and glial response in rats following transient middle cerebral artery occlusion. J. Pharmacol. Exp. Ther. 2018, 367, 528–542. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SHAM | 2.94 (2.63; 3.26) |
| pMCAO + V + S | 3.33 (2.88; 4.35) |
| pMCAO + C + S | 3.69 (3.42; 4.25) ** |
| pMCAO + V + EE | 3.31 (3.07; 3.92) |
| pMCAO + C + EE | 3.32 (3.07; 3.57) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakreen, A.; Juntunen, M.; Dunlop, Y.; Ugidos, I.F.; Malm, T.; Miettinen, S.; Jolkkonen, J. Additive Behavioral Improvement after Combined Cell Therapy and Rehabilitation Despite Long-Term Microglia Presence in Stroke Rats. Int. J. Mol. Sci. 2021, 22, 1512. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041512
Bakreen A, Juntunen M, Dunlop Y, Ugidos IF, Malm T, Miettinen S, Jolkkonen J. Additive Behavioral Improvement after Combined Cell Therapy and Rehabilitation Despite Long-Term Microglia Presence in Stroke Rats. International Journal of Molecular Sciences. 2021; 22(4):1512. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041512
Chicago/Turabian StyleBakreen, Abdulhameed, Miia Juntunen, Yannick Dunlop, Irene F. Ugidos, Tarja Malm, Susanna Miettinen, and Jukka Jolkkonen. 2021. "Additive Behavioral Improvement after Combined Cell Therapy and Rehabilitation Despite Long-Term Microglia Presence in Stroke Rats" International Journal of Molecular Sciences 22, no. 4: 1512. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041512