
A Role for Human Renal Tubular Epithelial Cells in Direct Allo-Recognition by CD4+ T-Cells and the Effect of Ischemia-Reperfusion

 , , and
, , and 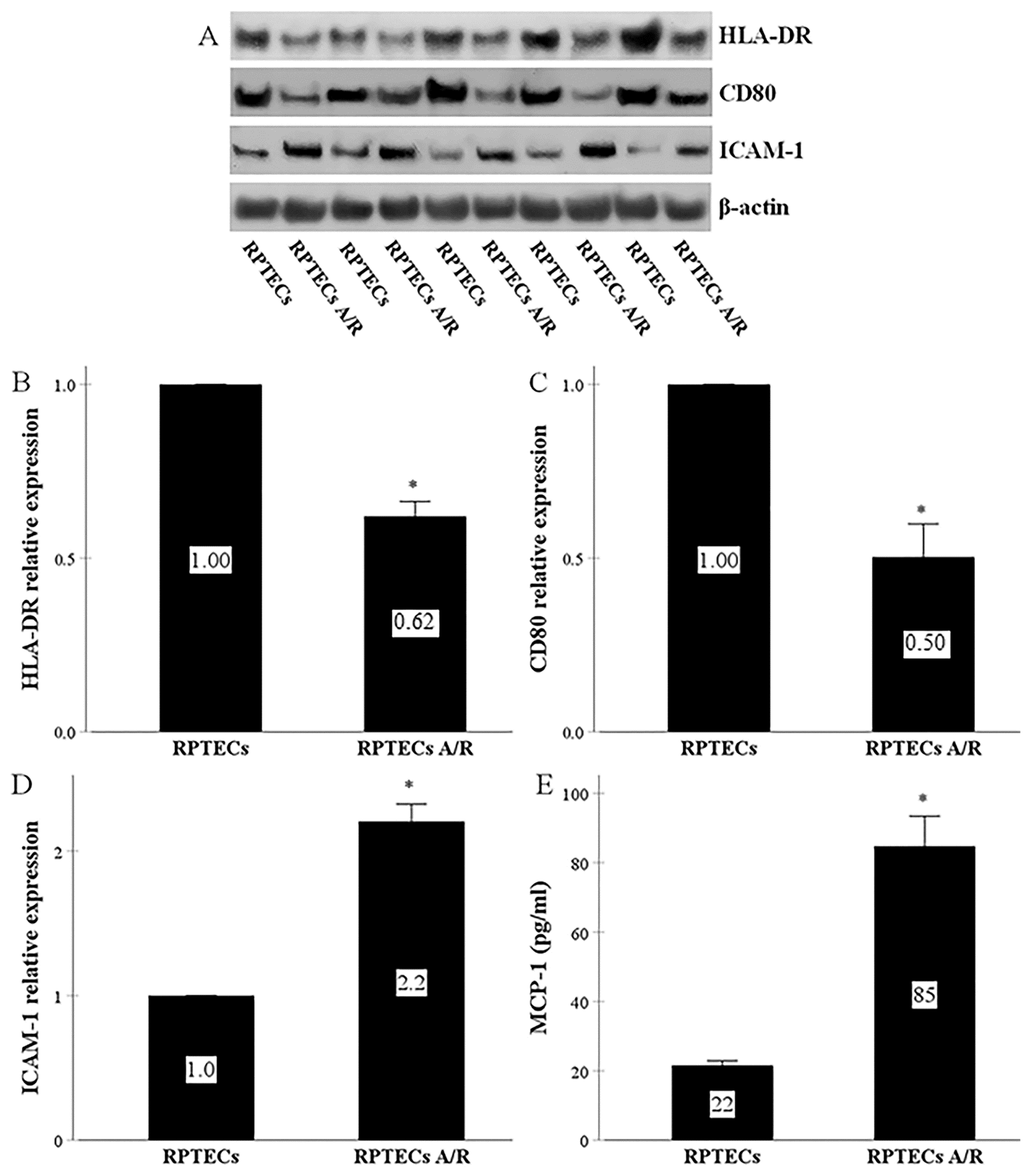{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. RPTECs Express All the Required Molecules for Direct Allorecognition by CD4+ T-Cells and Produce MCP-1, whereas Anoxia-Reoxygenation Affects All the Above
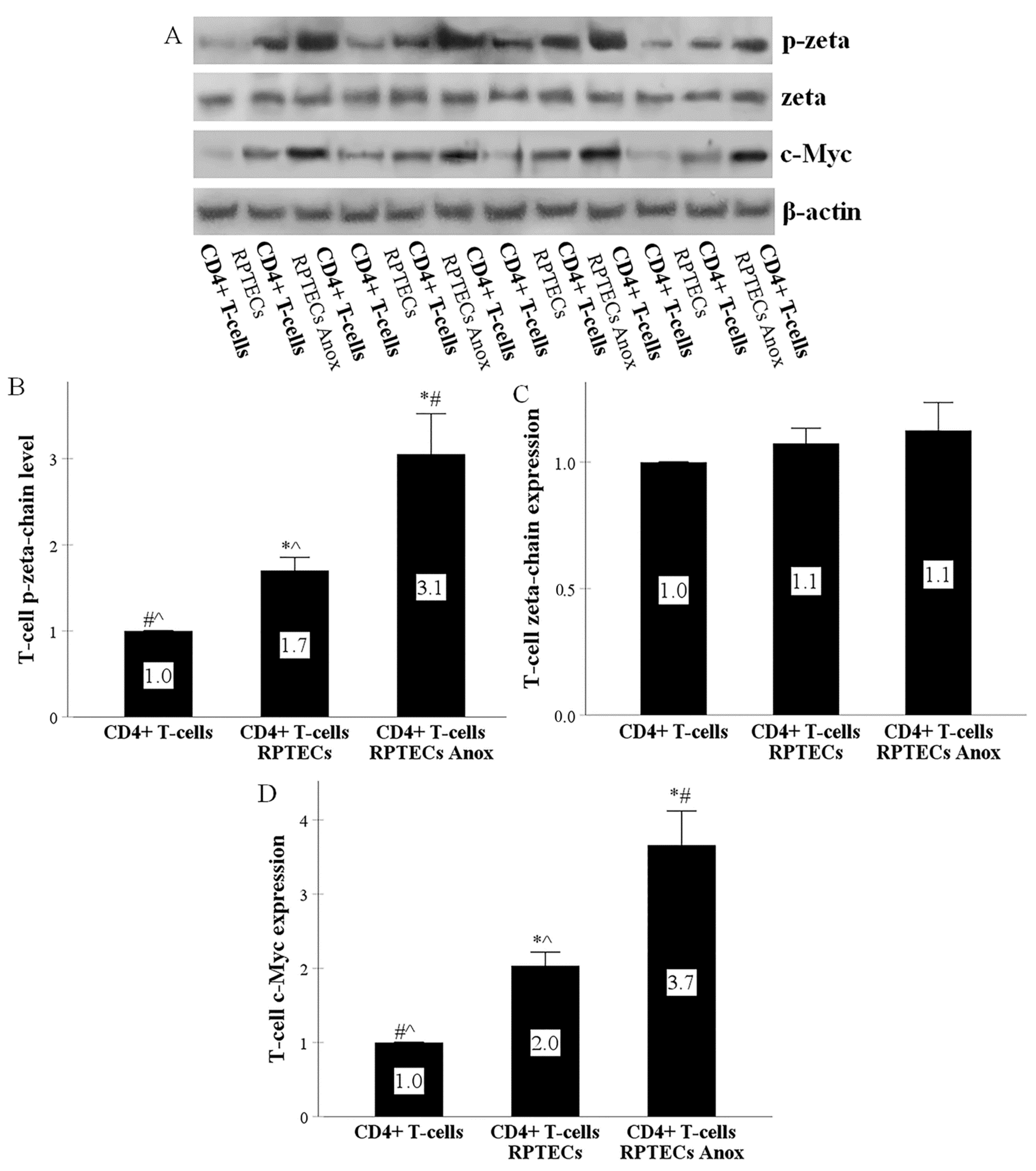
2.2. RPTECs Activate the T-Cell Receptor and the Co-Stimulation Signal Transduction Pathways in CD4+ T-Cells, and Anoxia-Reoxygenation Intensifies the above Signal Transduction Pathways Further
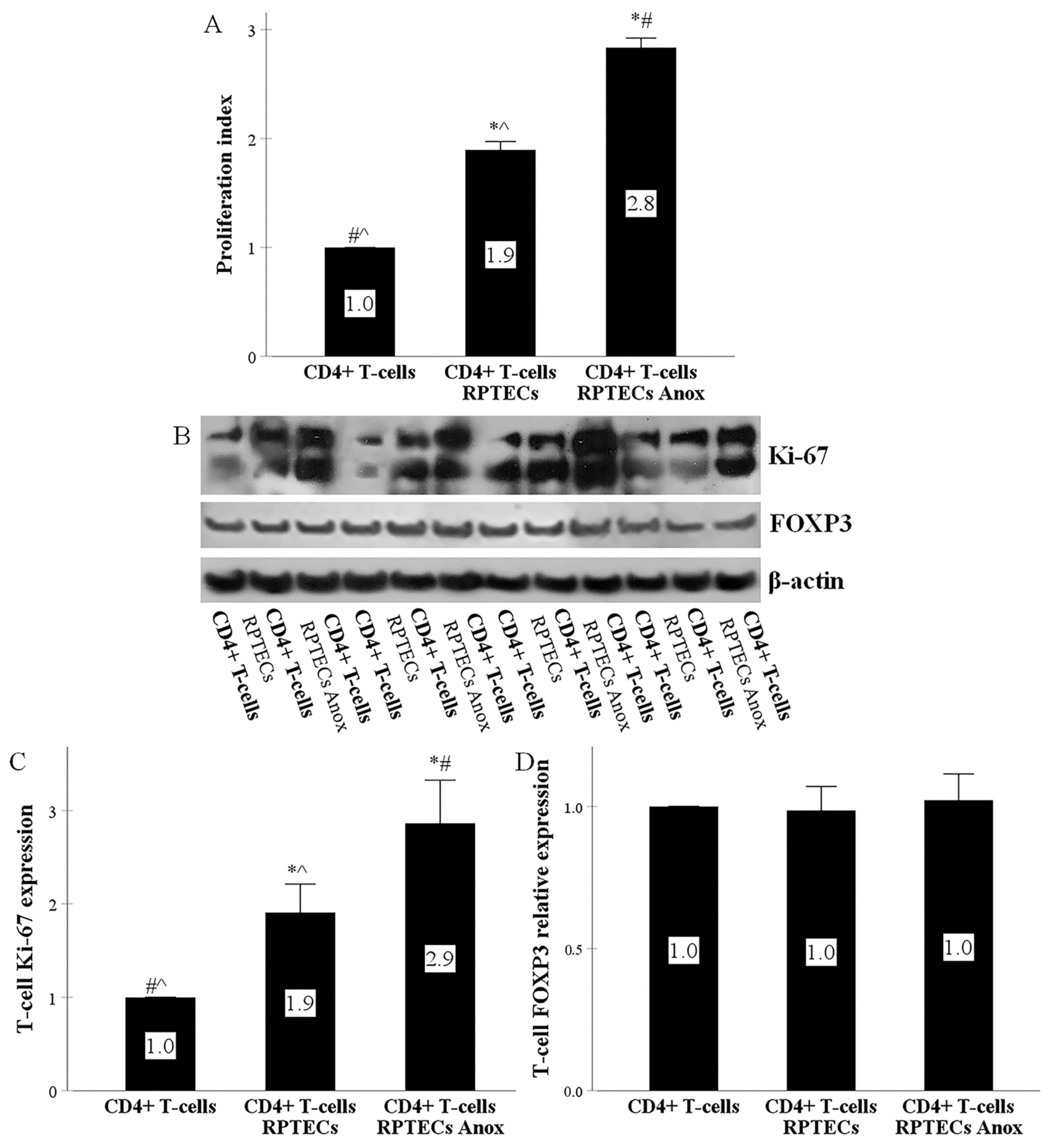
2.3. RPTECs Trigger CD4+ T-Cells to Proliferate, Anoxia-Reoxygenation Further Increases Proliferation, whereas CD4+ T-Cell Differentiation to Regulatory T-Cells Does Not Occur
3. Discussion
4. Materials and Methods
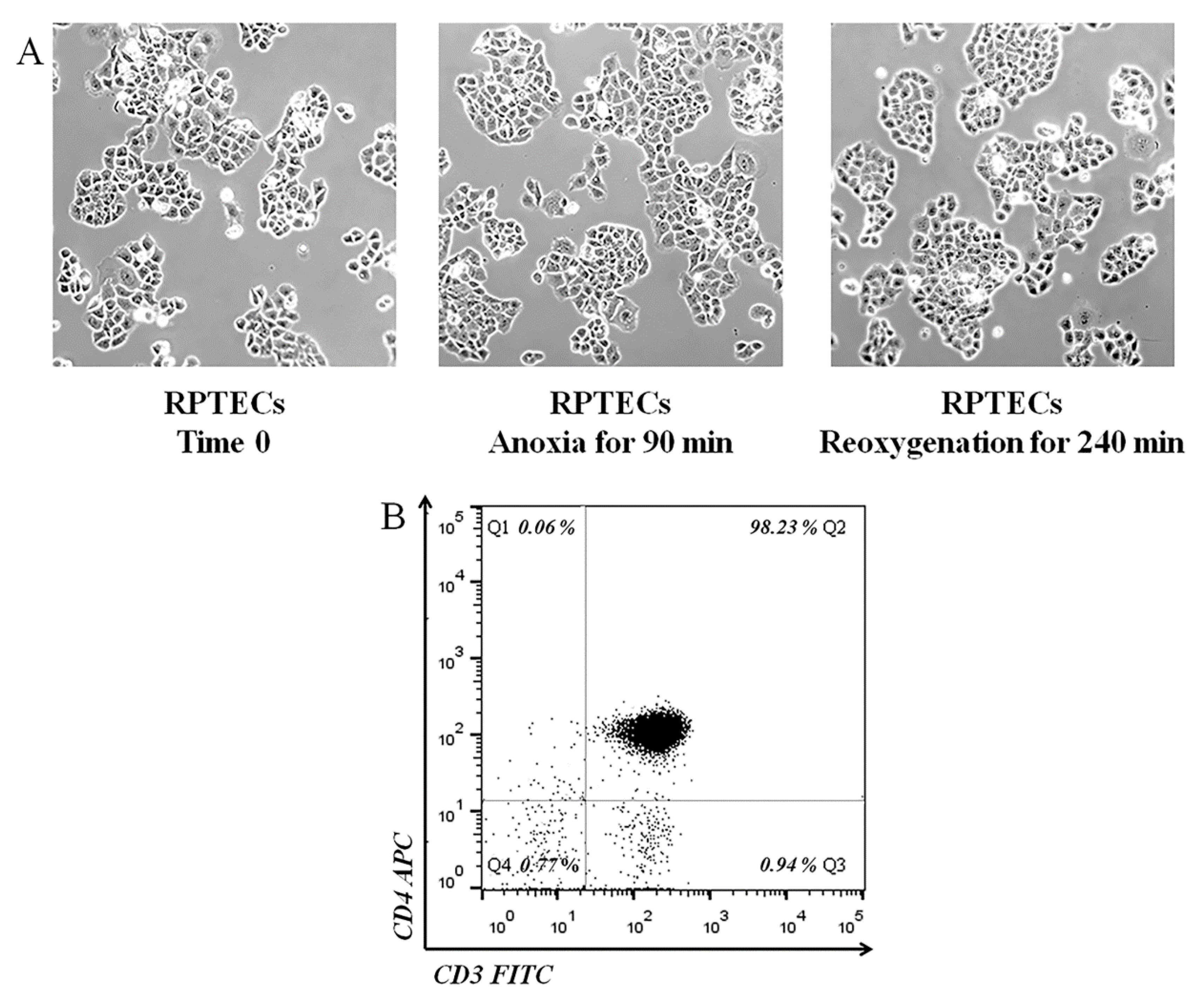
4.1. Cells and Culture Conditions
4.2. Assessment of the Proteins of Interest
4.3. Measurement of Monocyte Chemoattractant Protein-1
4.4. Assessment of Cell Proliferation
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abramowicz, D.; Oberbauer, R.; Heemann, U.; Viklicky, O.; Peruzzi, L.; Mariat, C.; Crespo, M.; Budde, K.; Oniscu, G.C. Recent advances in kidney transplantation: A viewpoint from the Descartes advisory board. Nephrol. Dial. Transplant. 2018, 33, 1699–1707. [Google Scholar] [CrossRef] [Green Version]
- Sellarés, J.; De Freitas, D.G.; Mengel, M.; Reeve, J.; Einecke, G.; Sis, B.; Hidalgo, L.G.; Famulski, K.; Matas, A.; Halloran, P.F. Understanding the Causes of Kidney Transplant Failure: The Dominant Role of Antibody-Mediated Rejection and Nonadherence. Am. J. Transplant. 2012, 12, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.S.; Schaubel, D.E.; Guidinger, M.K.; Andreoni, K.A.; Wolfe, R.A.; Merion, R.M.; Port, F.K.; Sung, R.S. A Comprehensive Risk Quantification Score for Deceased Donor Kidneys: The Kidney Donor Risk Index. Transplantation 2009, 88, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Mikhalski, D.; Wissing, K.M.; Ghisdal, L.; Broeders, E.N.; Touly, M.A.; Hoang, A.-D.; Loi, P.; Mboti, F.; Donckier, V.; Vereerstraeten, P.; et al. Cold Ischemia is a Major Determinant of Acute Rejection and Renal Graft Survival in the Modern Era of Immunosuppression. Transplantation 2008, 85, S3–S9. [Google Scholar] [CrossRef] [PubMed]
- Miglinas, M.; Supranaviciene, L.; Mateikaite, K.; Skebas, K.; Kubiliene, A. Delayed Graft Function: Risk Factors and the Effects of Early Function and Graft Survival. Transplant. Proc. 2013, 45, 1363–1367. [Google Scholar] [CrossRef] [PubMed]
- Ditonno, P.; Impedovo, S.; Palazzo, S.; Bettocchi, C.; Gesualdo, L.; Grandaliano, G.; Selvaggi, F.; Battaglia, M. Effects of Ischemia-Reperfusion Injury in Kidney Transplantation: Risk Factors and Early and Long-Term Outcomes in a Single Center. Transplant. Proc. 2013, 45, 2641–2644. [Google Scholar] [CrossRef] [PubMed]
- Helfer, M.; Vicari, A.; Spuldaro, F.; Gonçalves, L.; Manfro, R.C. Incidence, Risk Factors, and Outcomes of Delayed Graft Function in Deceased Donor Kidney Transplantation in a Brazilian Center. Transplant. Proc. 2014, 46, 1727–1729. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.K.; Famure, O.; Li, Y.; Kim, S.J. Delayed graft function and the risk of acute rejection in the modern era of kidney transplantation. Kidney Int. 2015, 88, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Sert, I.; Colak, H.; Tugmen, C.; Dogan, S.M.; Karaca, C. The effect of cold ischemia time on delayed graft function and acute rejection in kidney transplantation. Saudi J. Kidney Dis. Transplant. 2014, 25, 960. [Google Scholar] [CrossRef]
- Arias-Cabrales, C.E.; Pérez-Sáez, M.J.; Redondo-Pachón, D.; Buxeda, A.; Burballa, C.; Duran, X.; Mir, M.; Crespo, M.; Pascual, J. Relevance of KDPI value and acute rejection on kidney transplant outcomes in recipients with delayed graft function—A retrospective study. Transplant. Int. 2020, 33, 1071–1077. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, E.; Lerut, E.; Senev, A.; Coemans, M.; Pirenne, J.; Monbaliu, D.; Jochmans, I.; Sainz Barriga, M.; De Vusser, K.; Van Craenenbroeck, A.H.; et al. The Histological Picture of Indication Biopsies in the First 2 Weeks after Kidney Transplantation. Clin. J. Am. Soc. Nephrol. 2020, 15, 1484–1493. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Alam, A.; Soo, A.P.; George, A.J.; Ma, D. Ischemia-Reperfusion Injury Reduces Long Term Renal Graft Survival: Mechanism and Beyond. EBioMedicine 2018, 28, 31–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nankivell, B.J.; Alexander, S.I. Rejection of the kidney allograft. N. Engl. J. Med. 2010, 363, 1451–1462. [Google Scholar] [CrossRef] [Green Version]
- Safinia, N.; Afzali, B.; Atalar, K.; Lombardi, G.; Lechler, S.R. T-cell alloimmunity and chronic allograft dysfunction. Kidney Int. 2010, 78, S2–S12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hricik, D. Transplant Immunology and Immunosuppression: Core Curriculum 2015. Am. J. Kidney Dis. 2015, 65, 956–966. [Google Scholar] [CrossRef]
- Siu, J.H.Y.; Surendrakumar, V.; Richards, J.A.; Pettigrew, G.J. T cell Allorecognition Pathways in Solid Organ Transplantation. Front. Immunol. 2018, 9, 2548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaplin, D.D. Overview of the immune response. J. Allergy Clin. Immunol. 2010, 125, S3–S23. [Google Scholar] [CrossRef]
- Gaughan, A.; Wang, J.; Pelletier, R.P.; Nadasdy, T.; Brodsky, S.; Roy, S.; Lodder, M.; Bobek, D.; Mofatt-Bruce, S.; Fairchild, R.L.; et al. Key role for CD4 T cells during mixed antibody-mediated rejection of renal allografts. Am. J. Transplant. 2014, 14, 284–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, C.A.; Markovic-Lipkovski, J.; Risler, T.; Bohle, A.; Müller, G.A. Expression of HLA-DQ, -DR, and -DP antigens in normal kidney and glomerulonephritis. Kidney Int. 1989, 35, 116–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuthrich, R.P.; Glimcher, L.H.; Yui, M.A.; Jevnikar, A.M.; Dumas, S.E.; Kelley, V.E. MHC class II, antigen presentation and tumor necrosis factor in renal tubular epithelial cells. Kidney Int. 1990, 37, 783–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, D.D.H.; Welsh, K.I.; McLaren, A.J.; Roake, J.A.; Morris, P.J.; Fuggle, S.V. Cadaver versus living donor kidneys: Impact of donor factors on antigen induction before transplantation. Kidney Int. 1999, 56, 1551–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Posada, J.M.; Garcia-Castro, M.C.; Tamajon, L.P.; Torres, A.; Hernandez, D.; Losada, M.; Maceira, B.; Salido, E. HLA-DR class II and ICAM-1 expression on tubular cells taken by fine-needle aspiration biopsy in renal allograft dysfunction. Nephrol. Dial. Transplant. 1996, 11, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Dustin, M.L. The Cellular Context of T Cell Signaling. Immunity 2009, 30, 482–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couture, A.; Garnier, A.; Docagne, F.; Boyer, O.; Vivien, D.; Le-Mauff, B.; Latouche, J.-B.; Toutirais, O. HLA-Class II Artificial Antigen Presenting Cells in CD4+ T Cell-Based Immunotherapy. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Katsuma, A.; Yamakawa, T.; Nakada, Y.; Yamamoto, I.; Yokoo, T. Histopathological findings in transplanted kidneys. Ren. Replace. Ther. 2017, 3, 6. [Google Scholar] [CrossRef] [Green Version]
- Salcido-Ochoa, F.; Hue, S.S.-S.; Peng, S.; Fan, Z.; Li, R.L.; Iqbal, J.; Allen, J.C., Jr.; Loh, A.H.L. Histopathological analysis of infiltrating T cell subsets in acute T cell-mediated rejection in the kidney transplant. World J. Transplant. 2017, 7, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguado-Domínguez, E.; Cabrera-Pérez, R.; Suarez-Benjumea, A.; Abad-Molina, C.; Núñez-Roldán, A.; Aguilera, I. Computer-Assisted Definition of the Inflammatory Infiltrates in Patients with Different Categories of Banff Kidney Allograft Rejection. Front. Immunol. 2019, 10, 2605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telega, G.W.; Baumgart, D.C.; Carding, S.R. Uptake and presentation of antigen to T cells by primary colonic epithelial cells in normal and diseased states. Gastroenterology 2000, 119, 1548–1559. [Google Scholar] [CrossRef] [PubMed]
- Beers, C.; Burich, A.; Kleijmeer, M.J.; Griffith, J.M.; Wong, P.; Rudensky, A.Y. Cathepsin S Controls MHC Class II-Mediated Antigen Presentation by Epithelial Cells In Vivo. J. Immunol. 2005, 174, 1205–1212. [Google Scholar] [CrossRef] [Green Version]
- Arebro, J.; Tengroth, L.; Razavi, R.; Kumlien Georén, S.; Winqvist, O.; Cardell, L.O. Antigen-presenting epithelial cells can play a pivotal role in airway allergy. J. Allergy Clin. Immunol. 2016, 137, 957–960.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burghardt, S.; Claass, B.; Erhardt, A.; Karimi, K.; Tiegs, G. Hepatocytes induce Foxp3+ regulatory T cells by Notch signaling. J. Leukoc. Biol. 2014, 96, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Breda, P.C.; Wiech, T.; Meyer-Schwesinger, C.; Grahammer, F.; Huber, T.B.; Panzer, U.; Tiegs, G.; Neumann, K. Renal proximal tubular epithelial cells exert immunomodulatory function by driving inflammatory CD4+ T cell responses. Am. J. Physiol. Ren. Physiol. 2019, 317, F77–F89. [Google Scholar] [CrossRef] [PubMed]
- Schlüter, C.; Duchrow, M.; Wohlenberg, C.; Becker, M.H.; Key, G.; Flad, H.D.; Gerdes, J. The cell proliferation-associated antigen of antibody Ki-67: A very large, ubiquitous nuclear protein with numerous repeated elements, representing a new kind of cell cycle-maintaining proteins. J. Cell Biol. 1993, 123, 513–522. [Google Scholar] [CrossRef] [PubMed]
- de Teresa Alguacil, F.J.; de Gracia Guindo, C.; Fuentes, C.R.; Carrasco, R.C.; Marfil, A.P.; Sacristán, P.G.; Torres Sánchez, M.J.; Ortega, A.O. Patient’s Death with a Functioning Graft is still the Most Common Cause of Kidney Transplant Loss. Transplantation 2018, 102, S555. [Google Scholar] [CrossRef]
- Zhao, B.; Han, H.; Zhen, J.; Yang, X.; Shang, J.; Xu, L.; Wang, R. CD80 and CTLA-4 as diagnostic and prognostic markers in adult-onset minimal change disease: A retrospective study. PeerJ 2018, 6, e5400. [Google Scholar] [CrossRef] [PubMed]
- Kamiński, T.W.; Pawlak, K.; Karbowska, M.; Myśliwiec, M.; Pawlak, D. Indoxyl sulfate—The uremic toxin linking hemostatic system disturbances with the prevalence of cardiovascular disease in patients with chronic kidney disease. BMC Nephrol. 2017, 18, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samelson, L.E.; Harford, J.B.; Klausner, R.D. Identification of the components of the murine T cell antigen receptor complex. Cell 1985, 43, 223–231. [Google Scholar] [CrossRef]
- Eleftheriadis, T.; Antoniadi, G.; Liakopoulos, V.; Kortsaris, A. T-Cell Zeta Chain Expression, Phosphorylation and Degradation and their Role in T-Cell Signal Transduction and Immune Response Regulation in Health and Disease. Curr. Signal Transduct. Ther. 2006, 1, 191–208. [Google Scholar] [CrossRef]
- Kunkl, M.; Sambucci, M.; Ruggieri, S.; Amormino, C.; Tortorella, C.; Gasperini, C.; Battistini, L.; Tuosto, L. CD28 Autonomous Signaling Up-Regulates C-Myc Expression and Promotes Glycolysis Enabling Inflammatory T Cell Responses in Multiple Sclerosis. Cells 2019, 8, 575. [Google Scholar] [CrossRef] [Green Version]
- Haque, M.; Song, J.; Fino, K.; Wang, Y.; Sandhu, P.; Song, X.; Norbury, C.; Ni, B.; Fang, D.; Salek-Ardakani, S.; et al. C-Myc regulation by co-stimulatory signals modulates the generation of CD8 memory T cells during viral infection. Open Biol. 2016, 6, 150208. [Google Scholar] [CrossRef] [Green Version]
- Beckermann, K.E.; Hongo, R.; Ye, X.; Young, K.; Carbonell, K.; Contreras Healey, D.C.; Siska, P.J.; Barone, S.; Roe, C.E.; Smith, C.C.; et al. CD28 co-stimulation drives tumor-infiltrating T cell glycolysis to promote inflammation. JCI Insight 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Frauwirth, K.A.; Riley, J.L.; Harris, M.H.; Parry, R.V.; Rathmell, J.C.; Plas, D.R.; Elstrom, R.L.; June, C.H.; Thompson, C.B. The CD28 Signaling Pathway Regulates Glucose Metabolism. Immunity 2002, 16, 769–777. [Google Scholar] [CrossRef] [Green Version]
- Thaventhiran, T.; Wong, W.; Alghanem, A.F.; Alhumeed, N.; Aljasir, M.A.; Ramsey, S.; Sethu, S.; Yeang, H.X.A.; Chadwick, A.E.; Cross, M.; et al. CD28 Superagonistic Activation of T Cells Induces a Tumor Cell-Like Metabolic Program. Monoclon. Antibodies Immunodiagn. Immunother. 2019, 38, 60–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, A.; Govender, L.; Hughes, J.; Mavakla, W.; De Kock, M.; Barnard, C.; Pienaar, B.; Van Rensburg, E.J.; Jacobs, G.; Khomba, G.; et al. Novel application of Ki67 to quantify antigen-specific in vitro lymphoproliferation. J. Immunol. Methods 2010, 362, 43–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taner, T.; Heimbach, J.K.; Rosen, C.B.; Nyberg, S.L.; Park, W.D.; Stegall, M.D. Decreased chronic cellular and antibody-mediated injury in the kidney following simultaneous liver-kidney transplantation. Kidney Int. 2016, 89, 909–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saravia, J.; Chapman, N.M.; Chi, H. Helper T cell differentiation. Cell. Mol. Immunol. 2019, 16, 634–643. [Google Scholar] [CrossRef]
- Bachmann, M.F.; McKall-Faienza, K.; Schmits, R.; Bouchard, D.; Beacj, J.; Speiser, D.E.; Mak, T.W.; Ohashi, P.S. Distinct roles for LFA-1 and CD28 during activation of naive T cells: Adhesion versus co-stimulation. Immunity 1997, 7, 549–557. [Google Scholar] [CrossRef] [Green Version]
- Sligh, J.E.; Ballantyne, C.M.; Rich, S.S.; Hawkins, H.K.; Smith, C.W.; Bradley, A.; Beaudet, A.L. Inflammatory and immune responses are impaired in mice deficient in intercellular adhesion molecule 1. Proc. Natl. Acad. Sci. USA 1993, 90, 8529–8533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carr, M.W.; Roth, S.J.; Luther, E.; Rose, S.S.; Springer, T.A. Monocyte chemoattractant protein 1 acts as a T-lymphocyte chemoattractant. Proc. Natl. Acad. Sci. USA 1994, 91, 3652–3656. [Google Scholar] [CrossRef] [Green Version]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte Chemoattractant Protein-1 [MCP-1[: An Overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Martin-Moreno, P.L.; Tripathi, S.; Chandraker, A. Regulatory T Cells and Kidney Transplantation. Clin. J. Am. Soc. Nephrol. 2018, 13, 1760–1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellini, M.I.; Tortorici, F.; Amabile, M.I.; D’Andrea, V. Assessing Kidney Graft Viability and Its Cells Metabolism during Machine Perfusion. Int. J. Mol. Sci. 2021, 22, 1121. [Google Scholar] [CrossRef] [PubMed]
- Semba, C.P.; Gadek, T.R. Development of lifitegrast: A novel T-cell inhibitor for the treatment of dry eye disease. Clin. Ophthalmol. 2016, 10, 1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, J.; Barker, D.J.; Georgiou, X.; Cooper, M.A.; Flicek, P.; Marsh, S.G.E. IPD-IMGT/HLA Database. Nucleic Acids Res. 2019, 48, D948–D955. [Google Scholar] [CrossRef] [PubMed]
- Eleftheriadis, T.; Pissas, G.; Antoniadi, G.; Liakopoulos, V.; Stefanidis, I. Cell Death Patterns Due to Warm Ischemia or Reperfusion in Renal Tubular Epithelial Cells Originating from Human, Mouse, or the Native Hibernator Hamster. Biology 2018, 7, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eleftheriadis, T.; Pissas, G.; Crespo, M.; Nikolaou, E.; Liakopoulos, V.; Stefanidis, I. A Role for Human Renal Tubular Epithelial Cells in Direct Allo-Recognition by CD4+ T-Cells and the Effect of Ischemia-Reperfusion. Int. J. Mol. Sci. 2021, 22, 1733. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041733
Eleftheriadis T, Pissas G, Crespo M, Nikolaou E, Liakopoulos V, Stefanidis I. A Role for Human Renal Tubular Epithelial Cells in Direct Allo-Recognition by CD4+ T-Cells and the Effect of Ischemia-Reperfusion. International Journal of Molecular Sciences. 2021; 22(4):1733. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041733
Chicago/Turabian StyleEleftheriadis, Theodoros, Georgios Pissas, Marta Crespo, Evdokia Nikolaou, Vassilios Liakopoulos, and Ioannis Stefanidis. 2021. "A Role for Human Renal Tubular Epithelial Cells in Direct Allo-Recognition by CD4+ T-Cells and the Effect of Ischemia-Reperfusion" International Journal of Molecular Sciences 22, no. 4: 1733. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041733