
Early Covert Appearance of Marginal Zone B Cells in Salivary Glands of Sjögren′s Syndrome-Susceptible Mice: Initiators of Subsequent Overt Clinical Disease


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Lymphocyte and Autoimmune Profiles of Salivary Gland Lymphocytic Infiltration (LF) during SS Development
2.1.1. Lymphocytic Foci (LF)
2.1.2. Early Temporal Transcriptome Expressions in Salivary Glands of SSS Mice Identify Important Bioprocesses That Define the Pro-SS Autoimmune Response
2.1.3. MZB Cells Are Multifunctional and Therefore Express a Broad Array of Receptors That Regulate Specific Activities
2.1.4. The Appearance of Infiltrating Immune Cells in the Salivary Glands Is Regulated by Chemotactic Factors
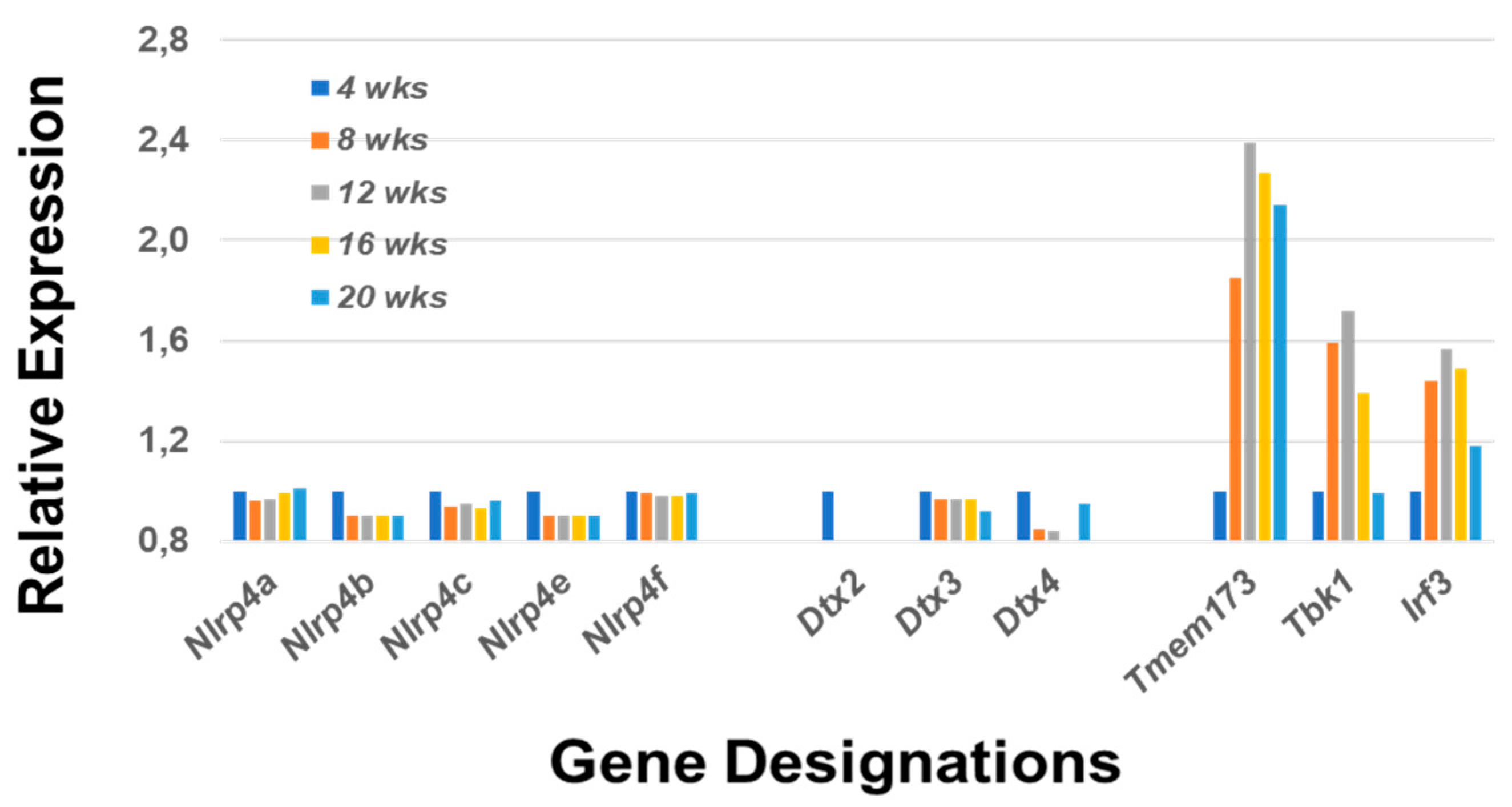
2.1.5. A Major Function of the Notch2 Signaling Pathway Is to Activate Mitogen-Activated Protein Kinase 14 (p38Mapk14α)
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Histology
4.3. RNA Preparation
4.4. Generation of Transcriptome Data
4.5. Gene expression Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| SS | Sjögren’s syndrome |
| SSS | Sjögren’s syndrome-susceptible |
| SSNS | Sjögren’s syndrome-non-susceptible |
| TG | Transgenic |
| SLE | Systemic lupus erythematosus |
| MZ | Marginal zone |
| MZB | Marginal zone B (cell) |
| NK | Natural killer (cell) |
| iNK | Invariant natural killer (cell) |
| DC | Dendritic cell |
| GC | Granulocyte |
| GC | Germinal center |
| CD | Cell differentiation antigen |
| B220 | CD45-receptor |
| TLR | Toll-like receptor |
| BCR | B cell receptor |
| S1P | Sphingosine-1-phosphate |
| S1PR | Sphingosine-1-phosphate receptor |
| C’ | Complement |
| IFN | Interferon |
| IFNR | Interferon receptor |
| IL | Interleukin |
| RNA | Ribonucleic acid |
| rt-PCR | Real-time polymerase chain reaction |
| FACS | Fluorescence-activated cell sorting |
| FITC | Fluorescein-5-isothiocynate |
References
- Dörner, T. Crossroads of B cell activation in autoimmunity: Rationale of targeting B cells. J. Rheumatol. Suppl. 2006, 77, 3–11. [Google Scholar]
- Hofmann, K.; Clauder, A.-K.; Manz, R.A. Targeting B Cells and Plasma Cells in Autoimmune Diseases. Front. Immunol. 2018, 9, 835. [Google Scholar] [CrossRef]
- Sakkas, L.I.; Daoussis, D.; Mavropoulos, A.; Liossis, S.-N.; Bogdanos, D.P. Regulatory B cells: New players in inflammatory and autoimmune rheumatic diseases. Semin. Arthritis Rheum. 2019, 48, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Du, W.; Wang, X.; Yuan, S.; Cai, X.; Liu, D.; Li, J.; Lu, L. Multiple Functions of B Cells in the Pathogenesis of Systemic Lupus Erythematosus. Int. J. Mol. Sci. 2019, 20, 6021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, J.A.; Hsu, H.; Mountz, J.D. Autoreactive B cells in SLE, villains or innocent bystanders? Immunol. Rev. 2019, 292, 120–138. [Google Scholar] [CrossRef]
- Kroese, F.G.M.; Abdulahad, W.H.; Haacke, E.; Bos, N.A.; Vissink, A.; Bootsma, H. B-cell hyperactivity in primary Sjogren’s syndrome. Expert Rev. Clin. Immunol. 2014, 10, 483–499. [Google Scholar] [CrossRef] [PubMed]
- Brito-Zerón, P.; Theander, E.; Baldini, C.; Seror, R.; Retamozo, S.; Quartuccio, L.; Bootsma, H.; Bowman, S.J.; Dorner, T.; Gottenberg, J.-E.; et al. Early diagnosis of primary Sjögren’s syndrome: EULAR-SS task force clinical recommendations. Expert Rev. Clin. Immunol. 2016, 12, 137–156. [Google Scholar] [CrossRef]
- Nocturne, G.; Mariette, X. B cells in the pathogenesis of primary Sjögren syndrome. Nat. Rev. Rheumatol. 2018, 14, 133–145. [Google Scholar] [CrossRef]
- Nguyen, C.Q.; Cha, S.R.; Peck, A.B. Sjogren’s syndrome (SjS)-like disease of mice: The importance of B lymphocytes and autoanti-bodies. Front. Biosci. 2007, 12, 1767–1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, L.; Yu, C.; Yang, C. Different Lymphoproliferative Disorders in Different Salivary Glands of Primary Sjögren Syndrome. J. Craniofacial Surg. 2013, 24, e462–e465. [Google Scholar] [CrossRef]
- Fisher, B.A.; Jonsson, R.; Daniels, T.; Bombardieri, M.; Brown, R.M.; Morgan, P.; Bombardieri, S.; Ng, W.F.; Tzioufas, A.G.; Vitali, C.; et al. Standardisation of labial salivary gland histopathology in clinical trials in primary Sjogren’s syndrome. Ann. Rheum. Dis. 2017, 76, 1161–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaleu, N.; Nguyen, C.Q.; Peck, A.B.; Jonsson, R. Sjogren’s syndrome: Studying the disease in mice. Arthritis Res. Ther. 2011, 13, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, C.P.; Yamachika, S.; Bounous, D.I.; Brayer, J.; Jonsson, R.; Holmdahl, R.; Peck, A.B.; Humphreys-Beher, M.G. A novel NOD-derived murine model of primary Sjogren’s syndrome. Arthritis Rheum. 1998, 41, 150–156. [Google Scholar] [CrossRef]
- Cha, S.; Nagashima, H.; Brown, V.B.; Peck, A.B.; Humphreys-Beher, M.G. Two NOD Idd-associated intervals contribute synergistically to the development of autoimmune exocrinopathy (Sjogren’s syndrome) on a healthy murine background. Arthritis Rheum. 2002, 46, 1390–1398. [Google Scholar] [CrossRef]
- Shen, L.; Zhang, C.; Wang, T.; Brooks, S.; Ford, R.J.; Lin-Lee, Y.C.; Kasianowicz, A.; Kumar, V.; Martin, L.; Liang, P.; et al. Development of Autoimmunity in IL-14α-Transgenic Mice. J. Immunol. 2006, 177, 5676–5686. [Google Scholar] [CrossRef] [Green Version]
- Peck, A.B.; Nguyen, C.Q. What can Sjogren’s syndrome-like disease in mice contribute to human Sjogren’s syndrome? Clin. Immunol. 2017, 182, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Robinson, C.P.; Brayer, J.; Yamachika, S.; Esch, T.R.; Peck, A.B.; Stewart, C.A.; Peen, E.; Jonsson, R.; Humphreys-Beher, M.G. Transfer of human serum IgG to nonobese diabetic Igmu null mice reveals a role for autoantibodies in the loss of secretory function of exocrine tissues in Sjogren’s syndrome. Proc. Natl. Acad. Sci. USA 1998, 95, 7538–7543. [Google Scholar] [CrossRef] [Green Version]
- Vincent, F.B.; Saulep-Easton, D.; Figgett, W.A.; Fairfax, K.A.; Mackay, F. The BAFF/APRIL system: Emerging functions beyond B cell biology and autoimmunity. Cytokine Growth Factor Rev. 2013, 24, 203–215. [Google Scholar] [CrossRef]
- Ding, J.; Zhang, W.; Haskett, S.; Pellerin, A.; Xu, S.; Petersen, B.; Jandreski, L.; Hamann, S.; Reynolds, T.L.; Zheng, T.S.; et al. BAFF overexpression increases lymphocytic infiltration in Sjogren’s target tissue, but only inefficiently promotes ectopic B-cell differentiation. Clin. Immunol. 2016, 169, 69–79. [Google Scholar] [CrossRef]
- Sharma, A.; Kiripolsky, J.; Klimatcheva, E.; Howell, A.; Fereidouni, F.; Levenson, R.; Rothstein, T.L.; Kramer, J.M. Early BAFF receptor blockade mitigates murine Sjogren’s syndrome: Concomitant targeting of CXCL13 and the BAFF receptor prevents salivary hypofunction. Clin. Immunol. 2016, 164, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Suresh, L.; Li, H.; Zhang, C.; Kumar, V.; Pankewycz, O.; Ambrus, J.L. IL-14 alpha, the nexus for primary Sjogren’s disease in mice and humans. Clin. Immunol. 2009, 130, 304–312. [Google Scholar] [CrossRef]
- Shen, L.; Suresh, L.; Wu, J.; Xuan, J.; Li, H.; Zhang, C.; Pankewycz, O.; Ambrus, J.L. A Role for Lymphotoxin in Primary Sjogren’s Disease. J. Immunol. 2010, 185, 6355–6363. [Google Scholar] [CrossRef] [Green Version]
- Martin, F.; Kearney, J.F. Marginal-zone B cells. Nat. Rev. Immunol. 2002, 2, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, A.; Cols, M.; Puga, I. Marginal zone B cells: Virtues of innate-like antibody-producing lymphocytes. Nat. Rev. Immunol. 2013, 13, 118–132. [Google Scholar] [CrossRef] [Green Version]
- Hillion, S.; Arleevskaya, M.I.; Blanco, P.; Bordron, A.; Brooks, W.H.; Cesbron, J.Y.; Kaveri, S.; Vivier, E.; Renaudineau, Y. The Innate Part of the Adaptive Immune System. Clin. Rev. Allergy Immunol. 2019, 58, 151–154. [Google Scholar] [CrossRef]
- Shen, L.; Gao, C.; Suresh, L.; Xian, Z.; Song, N.; Chaves, L.D.; Yu, M.; Ambrus, J.L., Jr. Central role for marginal zone B cells in an animal model of Sjogren’s syndrome. Clin. Immunol. 2016, 168, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Killedar, S.J.; Eckenrode, S.E.; McIndoe, R.A.; She, J.X.; Nguyen, C.Q.; Peck, A.B.; Cha, S.R. Early pathogenic events associated with Sjogren’s syndrome (SjS)-like disease of the NOD mouse using microarray analysis. Lab. Investig. 2006, 86, 1243–1260. [Google Scholar] [CrossRef]
- Nguyen, C.Q.; Sharma, A.; Lee, B.H.; She, J.X.; McIndoe, R.A.; Peck, A.B. Differential gene expression in the salivary gland during development and onset of xerostomia in Sjogren’s syn-drome-like disease of the C57BL/6.NOD-Aec1Aec2 mouse. Arthritis Res. Ther. 2009, 11, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, C.Q.; Sharma, A.; She, J.X.; McIndoe, R.A.; Peck, A.B. Differential gene expressions in the lacrimal gland during development and onset of keratoconjunctivitis sicca in Sjogren’s syndrome (SJS)-like disease of the C57BL/6.NOD-Aec1Aec2 mouse. Exp. Eye Res. 2009, 88, 398–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peck, A.B.; Nguyen, C.Q. Transcriptome analysis of the interferon-signature defining the autoimmune process of Sjogren’s syn-drome. Scand. J. Immunol. 2012, 76, 237–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaleu, N.; Nguyen, C.Q.; Tekle, K.M.; Jonsson, R.; Peck, A.B. Transcriptional landscapes of emerging autoimmunity: Transient aberrations in the targeted tissue’s extracellular milieu precede immune responses in Sjögren’s syndrome. Arthritis Res. Ther. 2013, 15, R174. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, C.; Peck, A.B. Unraveling the pathophysiology of Sjogren’s syndrome-associated dry eye disease. Ocul. Surf. 2009, 7, 11–27. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, C.; Singson, E.; Kim, J.Y.; Cornelius, J.G.; Attia, R.; Doyle, M.E.; Bulosan, M.; Cha, S.; Peck, A.B. Sjogren’s syndrome-like disease of C57BL/6.NOD-Aec1Aec2 mice: Gender differences in keratoconjunctivitis sicca defined by a cross-over in the chromosome 3 Aec1 locus. Scand. J. Immunol. 2006, 64, 295–307. [Google Scholar] [CrossRef]
- Nguyen, C.Q.; Peck, A.B. The interferon-signature of Sjogren’s syndrome: How unique biomarkers can identify underlying inflammatory and immunopathological mechanisms of specific diseases. Front. Immunol. 2013, 5, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witas, R.; Peck, A.B.; Ambrus, J.L.; Nguyen, C.Q. Sjogren’s Syndrome and TAM Receptors: A Possible Contribution to Disease Onset. J. Immunol. Res. 2019, 2019, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peck, A.B.; Saylor, B.T.; Nguyen, L.; Sharma, A.; She, J.X.; Nguyen, C.Q.; McIndoe, R.A. Gene Expression Profiling of Early-Phase Sjogren’s Syndrome in C57BL/6.NOD-Aec1Aec2 Mice Identifies Focal Adhesion Maturation Associated with Infiltrating Leukocytes. Investig. Ophthalmol. Vis. Sci. 2011, 52, 5647–5655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciccia, F.; Guggino, G.; Rizzo, A.; Bombardieri, M.; Raimondo, S.; Carubbi, F.; Cannizzaro, A.; Sireci, G.; Dieli, F.; Campisi, G.; et al. Interleukin (IL)-22 receptor 1 is over-expressed in primary Sjogren’s syndrome and Sjogren-associated non-Hodgkin lymphomas and is regulated by IL-18. Clin. Exp. Immunol. 2015, 181, 219–229. [Google Scholar] [CrossRef] [Green Version]
- Batista, F.D.; Harwood, N.E. The who, how and where of antigen presentation to B cells. Nat. Rev. Immunol. 2009, 9, 15–27. [Google Scholar] [CrossRef]
- Hammad, H.; Vanderkerken, M.; Pouliot, P.; Deswarte, K.; Toussaint, W.; Vergote, K.; Vandersarren, L.; Janssens, S.; Ramou, I.; Savvides, S.N.; et al. Transitional B cells commit to marginal zone B cell fate by Taok3-mediated surface expression of ADAM10. Nat. Immunol. 2017, 18, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Arruga, F.; Vaisitti, T.; Deaglio, S. The NOTCH Pathway and Its Mutations in Mature B Cell Malignancies. Front. Oncol. 2018, 8, 550. [Google Scholar] [CrossRef] [Green Version]
- Pillai, S.; Cariappa, A. The follicular versus marginal zone B lymphocyte cell fate decision. Nat. Rev. Immunol. 2009, 9, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.; Kearney, J.F. Positive Selection from Newly Formed to Marginal Zone B Cells Depends on the Rate of Clonal Production, CD19, and btk. Immunity 2000, 12, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Pikkarainen, T.; Elomaa, O.; Soininen, R.; Kodama, T.; Kraal, G.; Tryggvason, K. Defective Microarchitecture of the Spleen Marginal Zone and Impaired Response to a Thymus-Independent Type 2 Antigen in Mice Lacking Scavenger Receptors MARCO and SR-A. J. Immunol. 2005, 175, 8173–8180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, T.T.; Cyster, J.G. Integrin-mediated long-term B cell retention in the splenic marginal zone. Science 2002, 297, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, M.C.I.; Guinamard, R.; Bolland, S.; Sankala, M.; Steinman, R.M.; Ravetch, J.V. Macrophages Control the Retention and Trafficking of B Lymphocytes in the Splenic Marginal Zone. J. Exp. Med. 2003, 198, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Cinamon, G.; Matloubian, M.; Lesneski, M.J.; Xu, Y.; Low, C.; Lu, T.; Proia, R.L.; Cyster, J.G. Sphingosine 1-phosphate receptor 1 promotes B cell localization in the splenic marginal zone. Nat. Immunol. 2004, 5, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Cinamon, G.; Zachariah, M.A.; Lam, O.M.; Foss, F.W.; Cyster, J.G. Follicular shuttling of marginal zone B cells facilitates antigen transport. Nat. Immunol. 2008, 9, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Girkontaite, L.; Missy, K.; Sakk, V.; Harenberg, A.; Tedford, K.; Pötzel, T.; Pfeffer, K.; Fischer, K.D. Lsc is required for marginal zone B cells, regulation of lymphocyte motility and immune responses. Nat. Immunol. 2001, 2, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Tedford, K.; Steiner, M.; Koshutin, S.; Richter, K.; Tech, L.; Eggers, Y.; Jansing, I.; Schilling, K.; Hauser, A.E.; Korthals, M.; et al. The opposing forces of shear flow and sphingosine-1-phosphate control marginal zone B cell shuttling. Nat. Commun. 2017, 8, 2261. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate—A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B-Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Nguyen, C.; Cornelius, J.; Singson, E.; Killedar, S.; Cha, S.; Peck, A.B. Role of complement and B lymphocytes in Sjogren’s syndrome-like autoimmune exocrinopathy of NOD.B10-H2(B) mice. Mol. Immunol. 2006, 43, 1332–1339. [Google Scholar] [CrossRef] [PubMed]
- Brayer, J.B.; Cha, S.; Nagashima, H.; Yasunari, U.; Lindberg, A.; Diggs, S.; Martinez, J.; Goa, J.; Humphreys-Beher, M.G.; Peck, A.B. IL-4-Dependent Effector Phase in Autoimmune Exocrinopathy as Defined by the NOD.IL-4-Gene Knockout Mouse Model of Sjogren’s Syndrome. Scand. J. Immunol. 2001, 54, 133–140. [Google Scholar] [CrossRef]
- Gao, J.H.; Killedar, S.; Cornelius, J.G.; Nguyen, C.; Cha, S.; Peck, A.B. Sjogren’s syndrome in the NOD mouse model is an interleukin-4 time-dependent, antibody isotype-specific autoimmune disease. J. Autoimmun. 2006, 26, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.; Brayer, J.; Gao, J.; Brown, V.; Killedar, S.; Yasunari, U.; Peck, A.B. A dual role for interferon-gamma in the pathogenesis of Sjogren’s syndrome-like autoimmune exocrinopathy in the nonobese diabetic mouse. Scand. J. Immunol. 2004, 60, 552–565. [Google Scholar] [CrossRef]
- Wen, L.; Brill-Dashoff, J.; Shinton, S.A.; Asano, M.; Hardy, R.R.; Hayakawa, K. Evidence of Marginal-Zone B Cell- Positive Selection in Spleen. Immunity 2005, 23, 297–308. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peck, A.B.; Nguyen, C.Q.; Ambrus, J. Early Covert Appearance of Marginal Zone B Cells in Salivary Glands of Sjögren′s Syndrome-Susceptible Mice: Initiators of Subsequent Overt Clinical Disease. Int. J. Mol. Sci. 2021, 22, 1919. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041919
Peck AB, Nguyen CQ, Ambrus J. Early Covert Appearance of Marginal Zone B Cells in Salivary Glands of Sjögren′s Syndrome-Susceptible Mice: Initiators of Subsequent Overt Clinical Disease. International Journal of Molecular Sciences. 2021; 22(4):1919. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041919
Chicago/Turabian StylePeck, Ammon B., Cuong Q. Nguyen, and Julian Ambrus. 2021. "Early Covert Appearance of Marginal Zone B Cells in Salivary Glands of Sjögren′s Syndrome-Susceptible Mice: Initiators of Subsequent Overt Clinical Disease" International Journal of Molecular Sciences 22, no. 4: 1919. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041919