
Blue Light Induces Impaired Autophagy through Nucleotide-Binding Oligomerization Domain 2 Activation on the Mouse Ocular Surface

 ,
,
Abstract
:1. Introduction
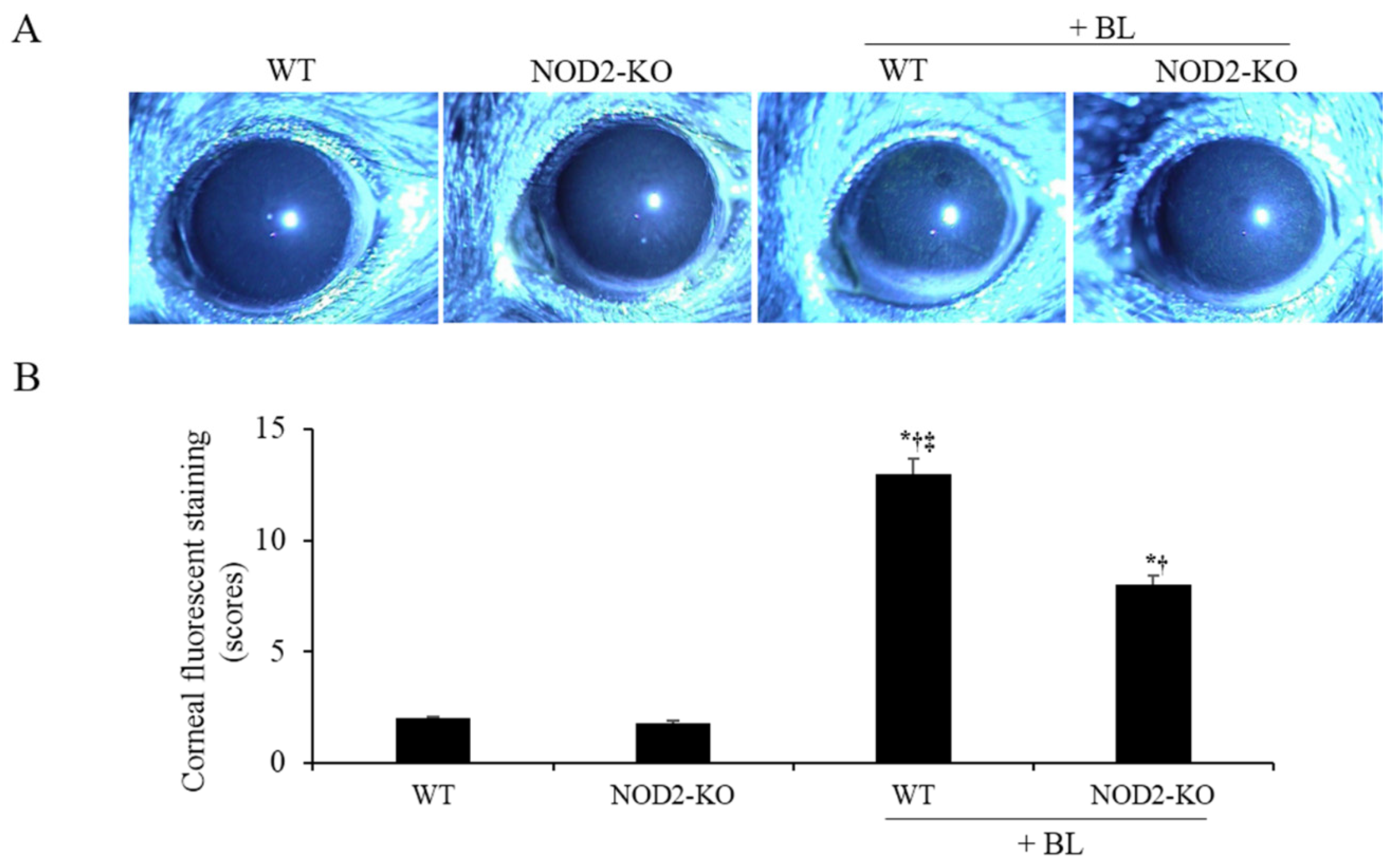
2. Results
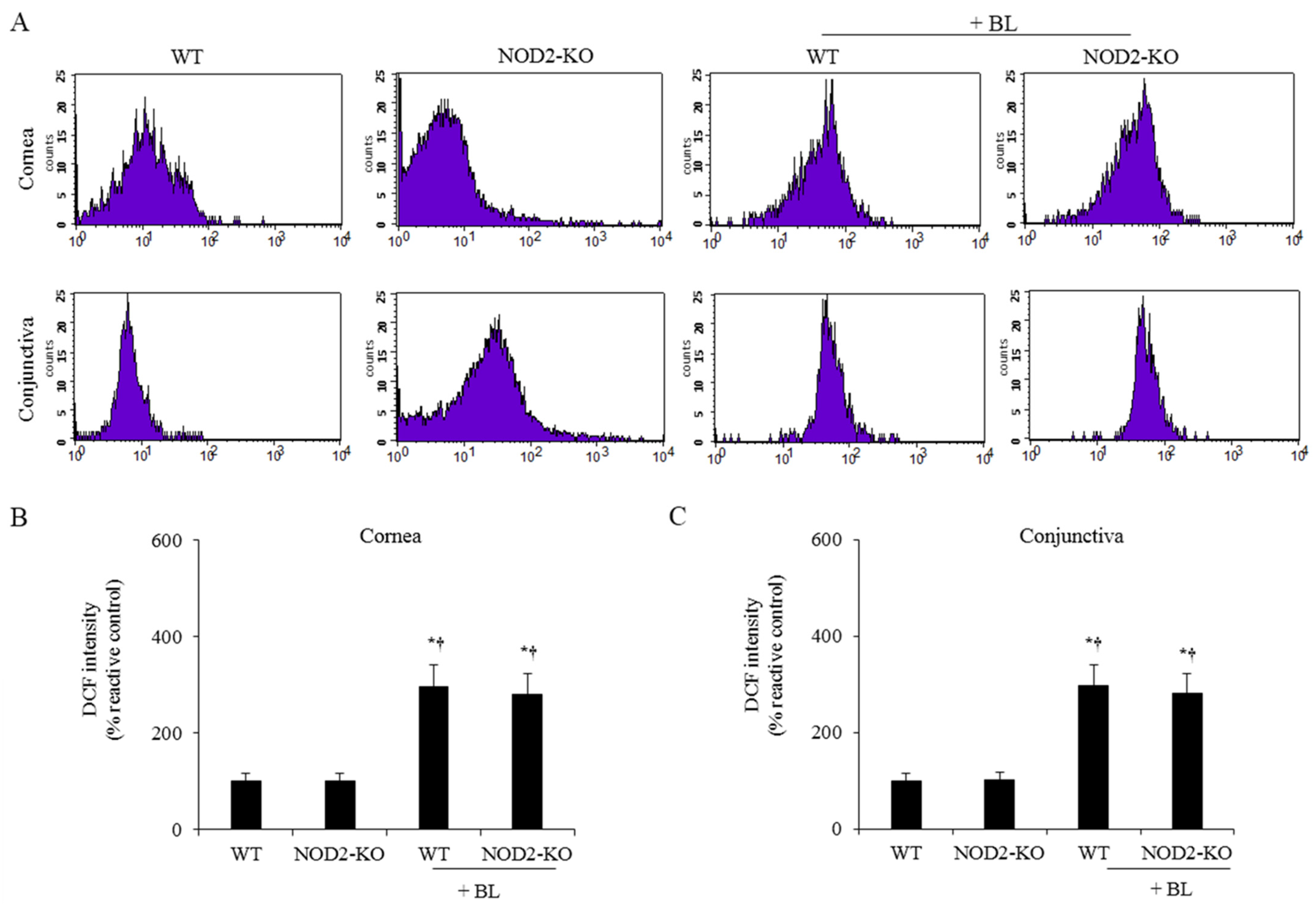
2.1. ROS Levels on the Ocular Surface
2.2. MDA Levels on the Ocular Surface
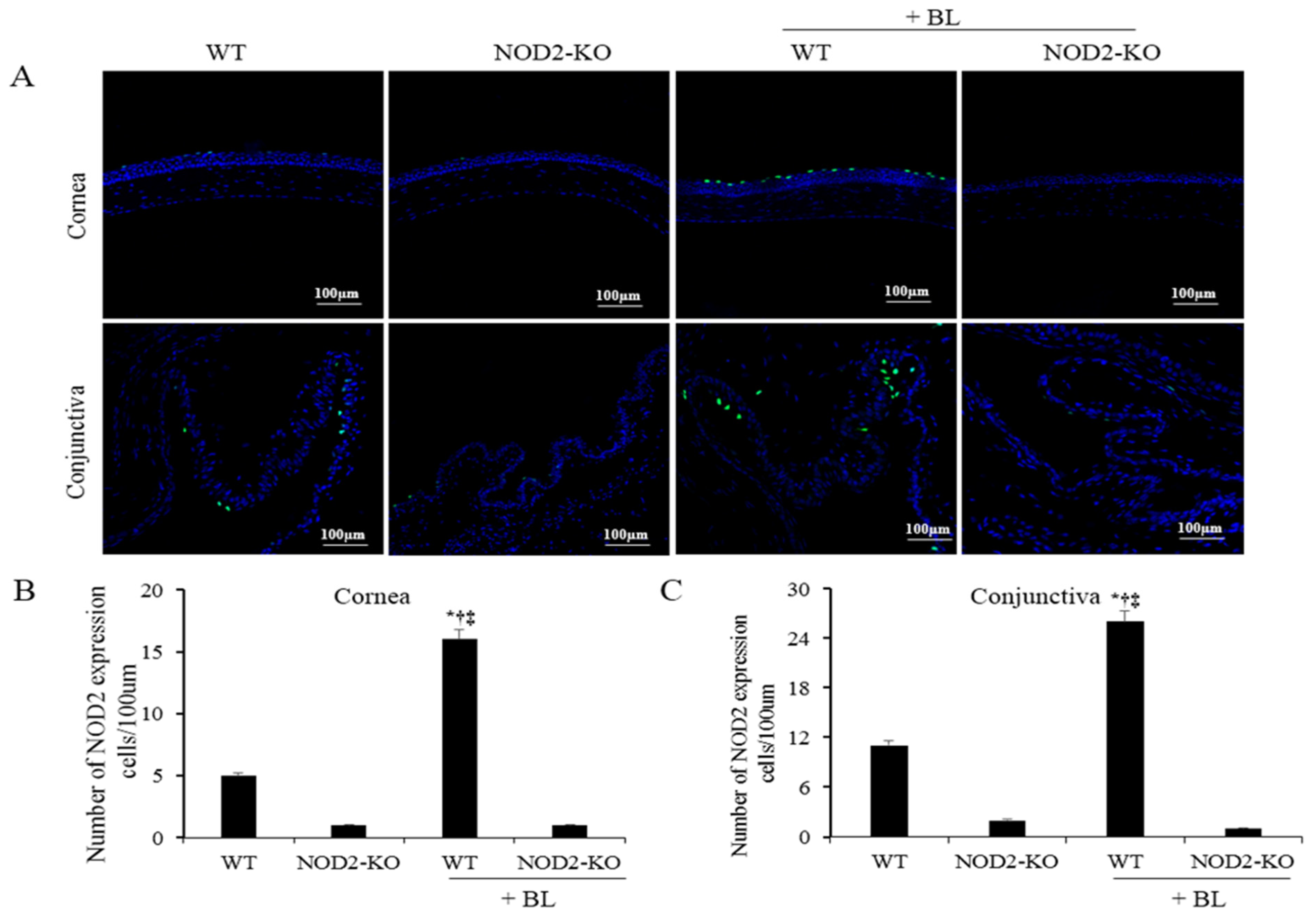
2.3. NOD2 Activation in Corneal and Conjunctival Tissues
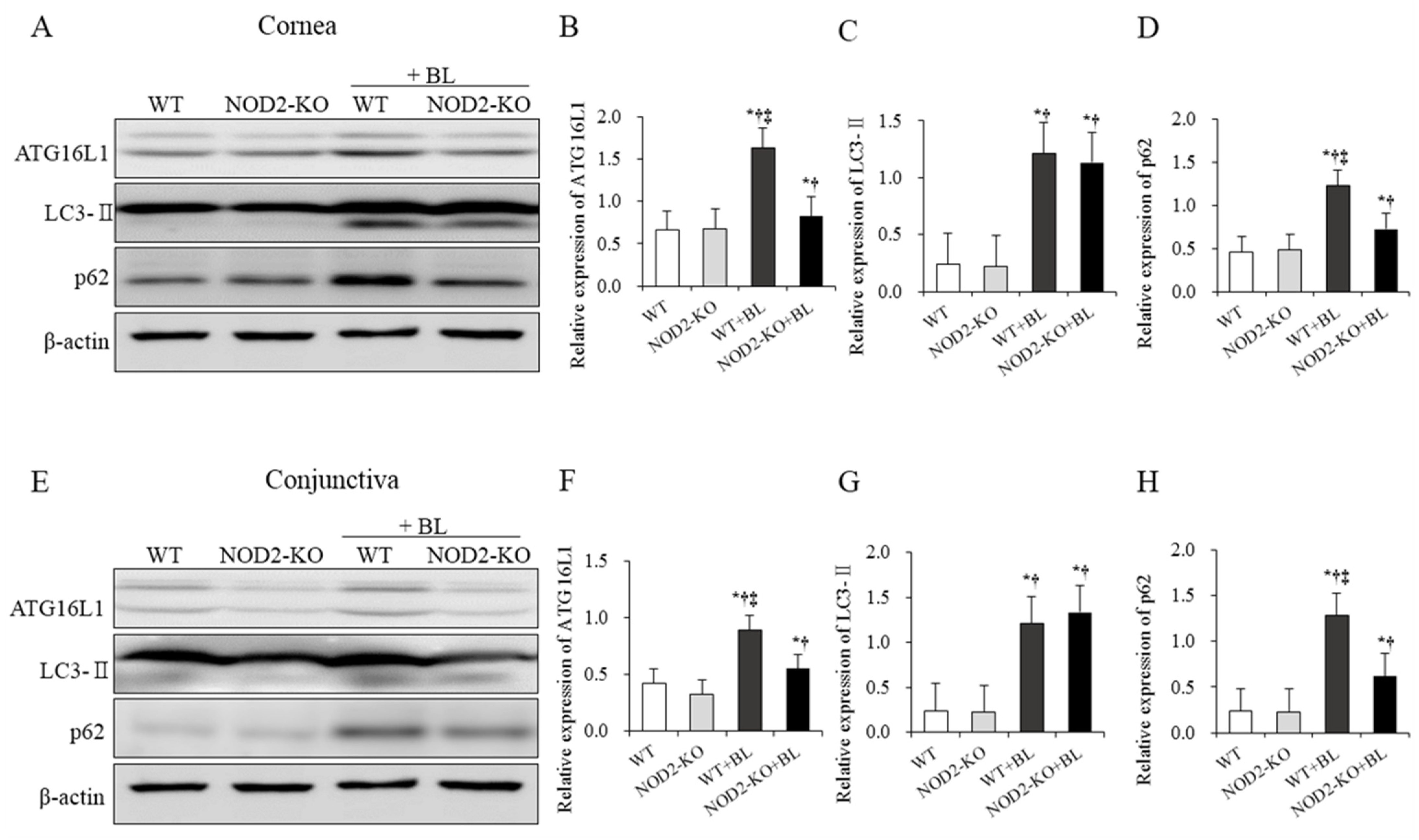
2.4. Western Blot for Autophagy in Corneal and Conjunctival Tissues
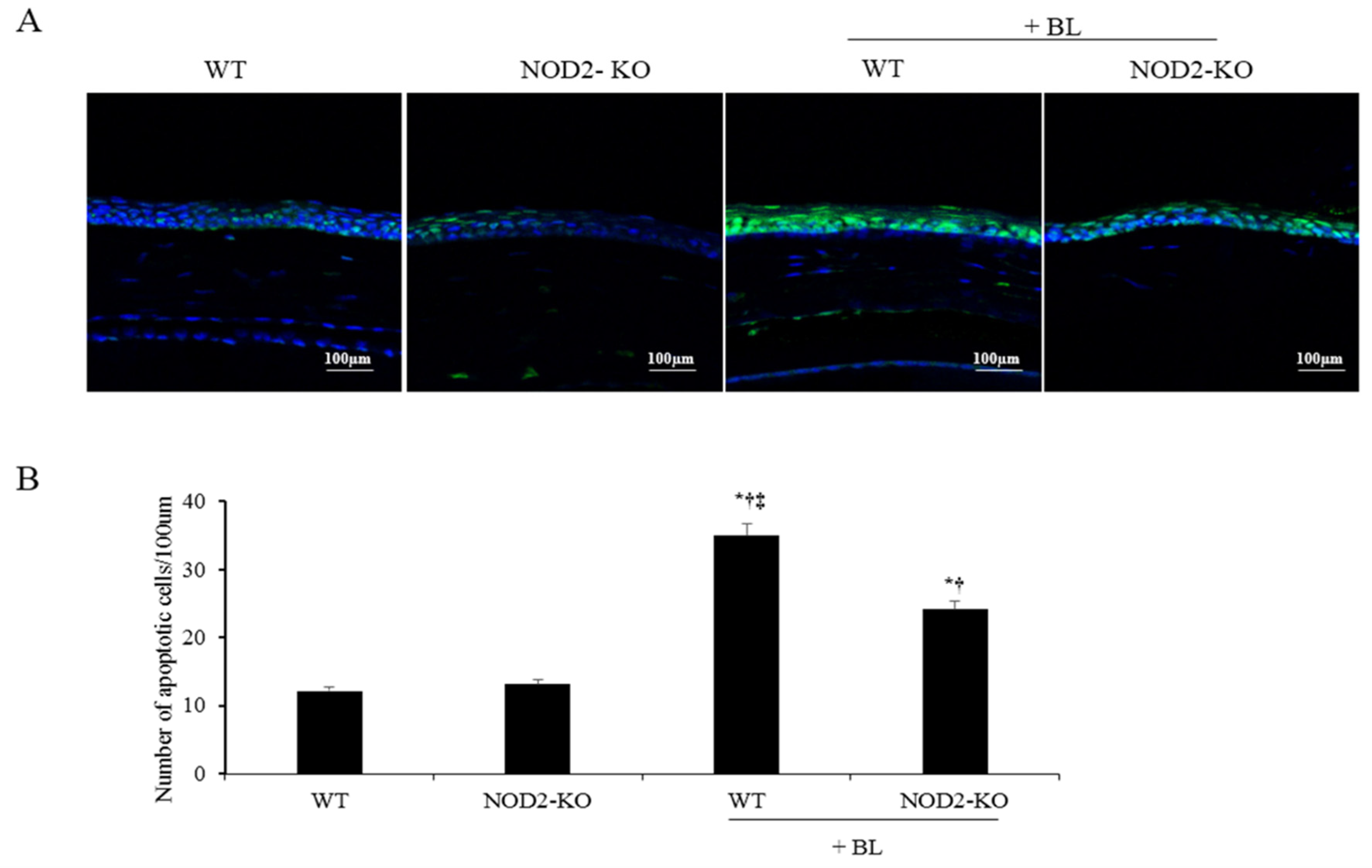
2.5. Counts of Apoptotic Cells in Corneal Tissue
2.6. Corneal Epithelial Damage
3. Discussion
4. Materials and Methods
4.1. Designs of Mouse Model and Experiments
4.2. Measurement of ROS Production in the Cornea and Conjunctiva
4.3. ELISA
4.4. Immunofluorescent Staining
4.5. Western Blot
4.6. Histology
4.7. TUNEL Staining
4.8. Ocular Surface Parameters
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moran, D.J.; Hollows, F.C. Pterygium and ultraviolet radiation: A positive correlation. Br. J. Ophthalmol. 1984, 68, 343–346. [Google Scholar] [CrossRef] [Green Version]
- Taylor, H.R.; West, S.K.; Rosenthal, F.S.; Munoz, B.; Newland, H.S.; Emmett, E.A. Corneal changes associated with chronic UV irradiation. Arch. Ophthalmol. 1989, 107, 1481–1484. [Google Scholar] [CrossRef] [PubMed]
- Taylor, H.R.; West, S.K.; Rosenthal, F.S.; Munoz, B.; Newland, H.S.; Abbey, H.; Emmett, E.A. Effect of ultraviolet radiation on cataract formation. N. Engl. J. Med. 1988, 319, 1429–1433. [Google Scholar] [CrossRef] [PubMed]
- Cruickshanks, K.J.; Klein, R.; Klein, B.E. Sunlight and age-related macular degeneration. The Beaver Dam Eye Study. Arch. Ophthalmol. 1993, 111, 514–518. [Google Scholar] [CrossRef]
- Zhang, W.; Ma, Y.; Zhang, Y.; Yang, J.; He, G.; Chen, S. Photo-oxidative blue-Light stimulation in retinal pigment epithelium cells promotes exosome secretion and increases the activity of the NLRP3 inflammasome. Curr. Eye. Res. 2019, 44, 67–75. [Google Scholar] [CrossRef]
- Hu, Z.; Zhang, Y.; Wang, J.; Mao, P.; Lv, X.; Yuan, S.; Huang, Z.; Ding, Y.; Xie, P.; Liu, Q. Knockout of Ccr2 alleviates photoreceptor cell death in rodent retina exposed to chronic blue light. Cell Death Dis. 2016, 7, e2468. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Cui, L.; Li, Y.; Choi, J.S.; Li, Z.; Kim, G.E.; Choi, W.; Yoon, K.C. Influence of light emitting diode-derived blue light overexposure on mouse ocular surface. PLoS ONE 2016, 11, e0161041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, H.; Park, J.K.; Lavker, R.M. Autophagy and Macropinocytosis: Keeping an Eye on the Corneal/Limbal Epithelia. Investig. Ophthalmol. Vis. Sci. 2017, 58, 416–423. [Google Scholar] [CrossRef] [Green Version]
- Barbéa, F.; Douglas, T.; Saleh, M. Advances in Nod-like receptors (NLR) biology. Cytokine Growth Factor Rev. 2014, 25, 681–697. [Google Scholar] [CrossRef]
- Akira, S.; Takeda, K.; Kaisho, T. Toll-like receptors: Critical proteins linking innate and acquired immunity. Nat. Immunol. 2001, 2, 675–680. [Google Scholar] [CrossRef]
- Brandstetter, C.; Mohr, L.K.M.; Latz, E.; Holz, F.G.; Krohne, T.U. Light induces NLRP3 inflammasome activation in retinal pigment epithelial cells via lipofuscin-mediated photooxidative damage. J. Mol. Med. (Berl.) 2015, 93, 905–916. [Google Scholar] [CrossRef] [Green Version]
- Cooney, R.; Baker, J.; Brain, O.; Danis, B.; Pichulik, T.; Allan, P.; Ferguson, D.J.; Campbell, B.J.; Jewell, D.; Simmons, A. NOD2 stimulation induces autophagy in dendritic cells influencing bacterial handling and antigen presentation. Nat. Med. 2010, 16, 90–97. [Google Scholar] [CrossRef]
- Homer, C.R.; Richmond, A.L.; Rebert, N.A.; Achkar, J.P.; McDonaid, C. ATG16L1 and NOD2 interact in an autophagy-dependent antibacterial pathway implicated in Crohn’s disease pathogenesis. Gastroenterology 2010, 139, 1630–1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, A.; Lopes, F.; Poon, K.K.H.; McKay, D.M. Absence of the NOD2 protein renders epithelia more susceptible to barrier dysfunction due to mitochondrial dysfunction. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G26–G38. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Li, Z.; Cui, L.; Li, Y.; Yoon, H.J.; Choi, W.; Lee, J.B.; Liu, Z.; Yoon, K.C. Expression of nod-like receptors and clinical correlations in patients with dry eye disease. Am. J. Ophthalmol. 2019, 200, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jin, R.; Li, L.; Yoon, H.J.; Choi, J.H.; Park, J.H.; Liu, Z.; Li, W.; Li, Z.; Yoon, K.C. Expression and role of nucleotide-binding oligomerization domain 2 (NOD2) in the ocular surface of murine dry eye. Investig. Ophthalmol. Vis. Sci. 2019, 60, 2641–2649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schäffler, H.; Rohde, M.; Rohde, S.; Huth, A.; Gittel, N.; Hollborn, H.; Koczan, D.; Glass, Ä.; Lamprecht, G.; Jaster, R. NOD2- and disease-specific gene expression profiles of peripheral blood mononuclear cells from Crohn’s disease patients. World. J. Gastroenterol. 2018, 24, 1196–1205. [Google Scholar] [CrossRef]
- Le Bourhis, L.; Benko, S.; Girardin, S.E. Nod1 and Nod2 in innate immunity and human inflammatory disorders. Biochem. Soc. Trans. 2007, 35, 1479–1484. [Google Scholar] [CrossRef] [Green Version]
- Iida, T.; Yokoyama, Y.; Wagatsuma, K.; Hirayama, D.; Nakase, H. Impact of autophagy of innate immune cells on inflammatory bowel disease. Cells 2018, 8, 7. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Xu, H.; An, M. mTORC1 regulates apoptosis and cell proliferation in pterygium via targeting autophagy and FGFR3. Sci. Rep. 2017, 7, 7339. [Google Scholar] [CrossRef] [Green Version]
- Strober, W.; Watanabe, T. NOD2, an intracellular innate immune sensor involved in host defense and Crohn’s disease. Mucosal. Immunol. 2011, 4, 484–495. [Google Scholar] [CrossRef] [PubMed]
- Fritz, T.; Niederreiter, L.; Adolph, T.; Blumberg, R.S.; Kaser, A. Crohn’s disease: NOD2, autophagy and ER stress converge. Gut 2011, 60, 1580–1588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillai, P.; Sobrin, L. Blau syndrome-associated uveitis and the NOD2 gene. Semin. Ophthalmol. 2013, 28, 327–332. [Google Scholar] [CrossRef]
- Choi, W.; Li, Z.; Oh, H.J.; Im, S.K.; Lee, S.H.; Park, S.H.; You, I.C.; Yoon, K.C. Expression of CCR5 and its ligands CCL3, -4, and -5 in the tear film and ocular surface of patients with dry eye disease. Curr. Eye Res. 2012, 37, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Li, Z.; Choi, J.S.; Oh, H.J.; Park, S.H.; Yoon, K.C. Expression of CXCL9, -10, and -11 in the aqueous humor of patients with herpetic endotheliitis. Cornea 2012, 31, 1246–1250. [Google Scholar] [CrossRef]
- Pauly, A.; Brignole-Baudouin, F.; Labbė, A.; Liang, H.; Warnet, J.M.; Baudouin, C. New tools for the evaluation of toxic ocular surface changes in the rat. Investig. Ophthalmol. Vis. Sci. 2007, 48, 5473–5483. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cornea (pmol/mL) | Conjunctiva (pmol/mL) | |
|---|---|---|
| WT | 7.6 ± 1.3 | 8.2 ± 2.3 |
| NOD2-KO | 8.0 ± 1.6 | 7.1 ± 1.1 |
| WT+BL | 15.0 ± 2.4 *,† | 14.0 ± 2.5 *,† |
| NOD2-KO+BL | 14.2 ± 3.0 *,† | 11.8 ± 2.2 *,† |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Jin, R.; Li, L.; Choi, J.S.; Kim, J.; Yoon, H.J.; Park, J.H.; Yoon, K.C. Blue Light Induces Impaired Autophagy through Nucleotide-Binding Oligomerization Domain 2 Activation on the Mouse Ocular Surface. Int. J. Mol. Sci. 2021, 22, 2015. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042015
Li Y, Jin R, Li L, Choi JS, Kim J, Yoon HJ, Park JH, Yoon KC. Blue Light Induces Impaired Autophagy through Nucleotide-Binding Oligomerization Domain 2 Activation on the Mouse Ocular Surface. International Journal of Molecular Sciences. 2021; 22(4):2015. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042015
Chicago/Turabian StyleLi, Ying, Rujun Jin, Lan Li, Ji Suk Choi, Jonghwa Kim, Hyeon Jeong Yoon, Jong Hwan Park, and Kyung Chul Yoon. 2021. "Blue Light Induces Impaired Autophagy through Nucleotide-Binding Oligomerization Domain 2 Activation on the Mouse Ocular Surface" International Journal of Molecular Sciences 22, no. 4: 2015. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042015