
Abiotic Stress Response of Near-Isogenic Spring Durum Wheat Lines under Different Sowing Densities

 ,
,
Abstract
:1. Introduction
2. Results
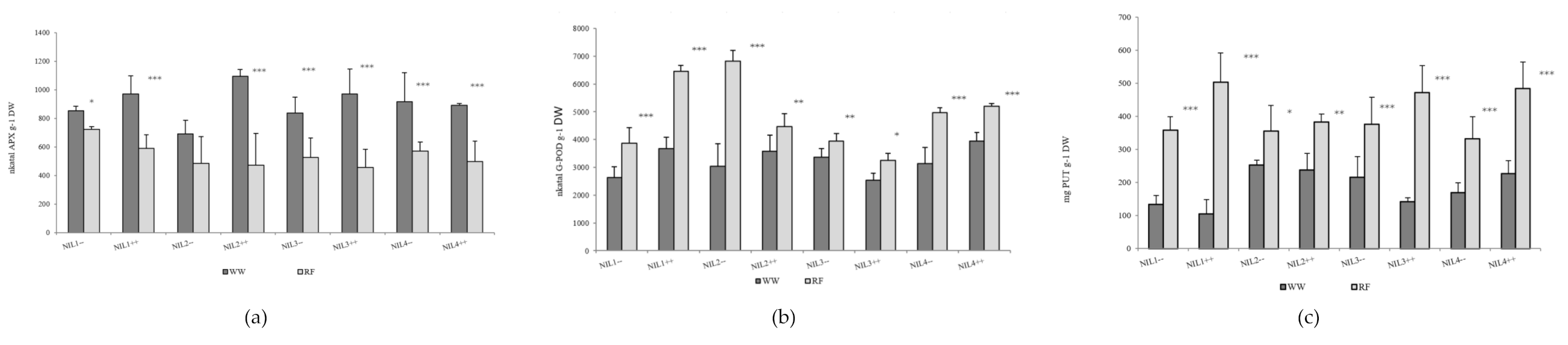
2.1. Effect of Drought Stress and Plant Sowing Densities on Morphological, Physiological, Biochemical and Yield Component Traits in Near-Isogenic Lines of Spring Durum Wheat
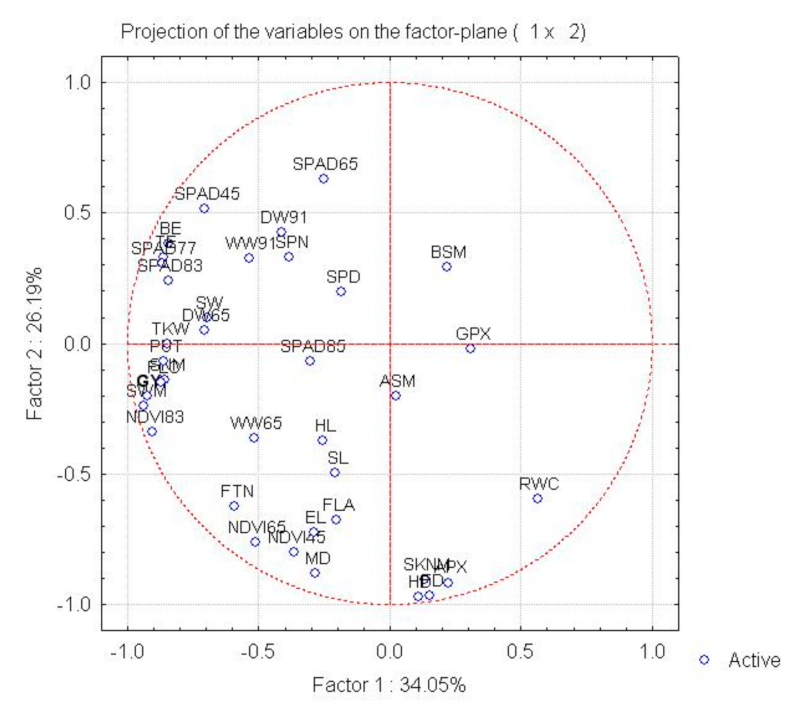
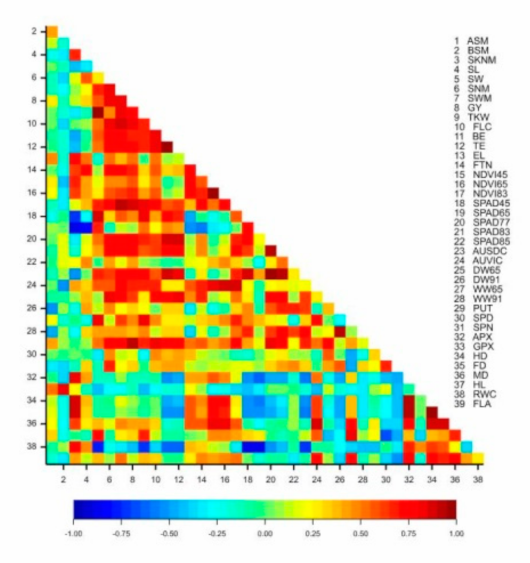
2.2. Interaction between the Morphological, Physiological, Biochemical and Yield Component Traits in Near-Isogenic Lines of Spring Durum Wheat under Drought Stress
3. Discussion
4. Materials and Methods
4.1. Plant Materials
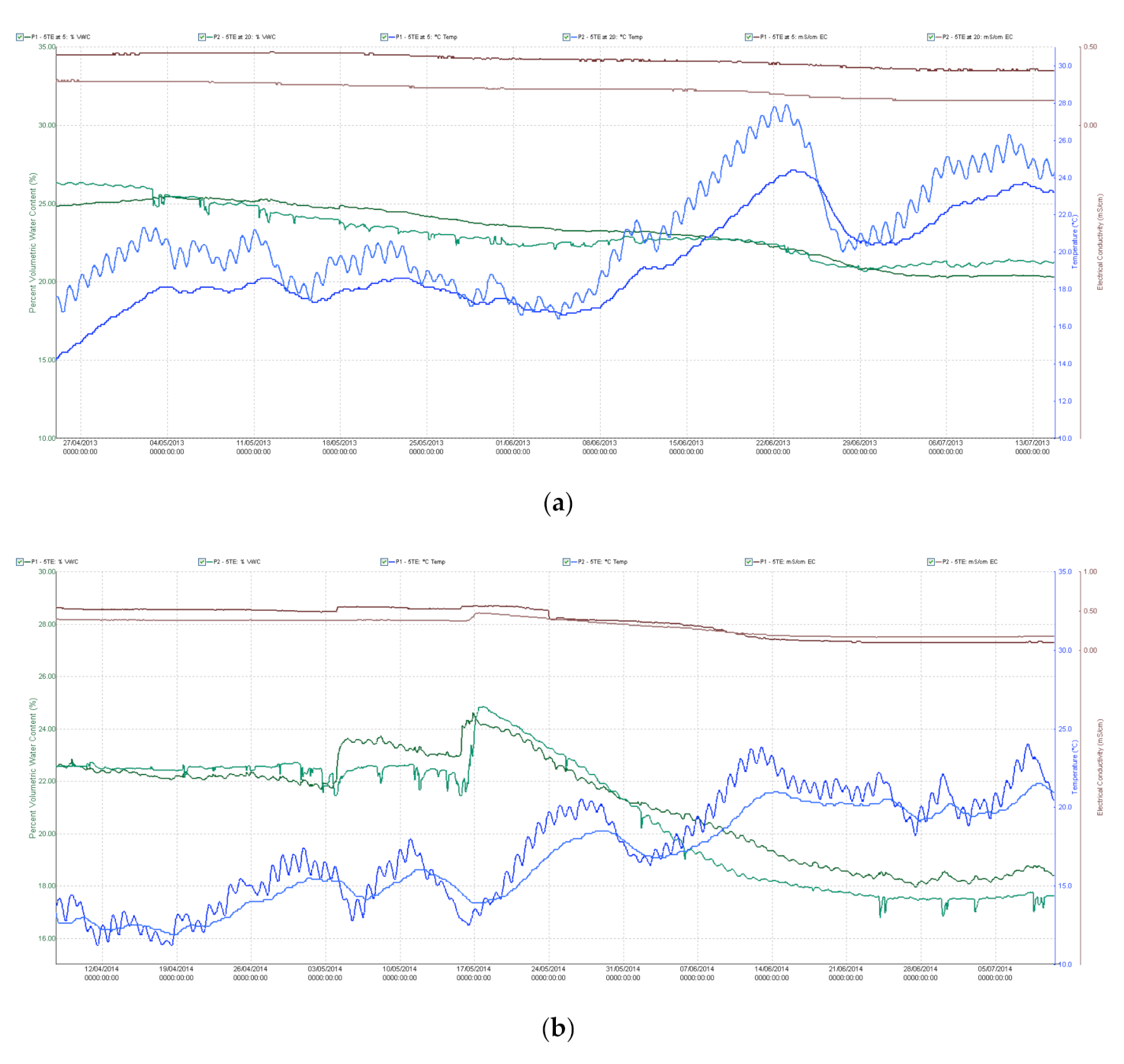
4.2. Site Description and Set-Up for Field Experiment
4.3. Data Collection
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| AUSDC | area under SPAD value decline curve |
| AUVIC | area under vegetation index curve |
| HD | heading date |
| SPAD | SPAD unit |
| NDVI | normalized difference vegetation index |
| GY | grain yield (t/ha) |
| FW | biomass fresh weight (kg) |
| DW | biomass dry weight (kg) |
| HL | test weight (HL) |
| FLC | plant height up to the flag leaf collar (cm) |
| BE | plant height up to the base of the ear (cm) |
| TE | plant height up to the tip of the ear (cm) |
| FTN | fertile tiller number (pcs./m2) |
| SNM | seed number/main spike |
| SWM | seed weight/main spike (g) |
| SW | seed width (cm) |
| SL | seed length (cm) |
| TGW | thousand-grain weight (g) |
| RWC | relative water content (%) |
| APX | ascorbate peroxidase (nkatal g-1 DW) |
| GPX | guaiacol peroxidase (nkatal g-1 DW) |
| PUT | putrescine (mg g-1 DW) |
| SPD | spermidine (mg g-1 DW) |
| SPN | spermine (mg g-1 DW) |
| Z | Zadoks scale |
Appendix A

References
- Trethowan, R.M.; Reynolds, M.; Sayre, K.; Ortiz-Monasterio, I. Adapting wheat cultivars to resource conserving farming practices and human nutritional needs. Ann. Appl. Biol. 2005, 146, 405–413. [Google Scholar] [CrossRef]
- Fukai, S.; Pantuwan, G.; Jongdee, B.; Cooper, M. Screening for drought resistance in rainfed lowland rice. Field Crops Res. 1999, 64, 61–74. [Google Scholar] [CrossRef]
- Fleury, D.; Jefferies, S.; Kuchel, H.; Langridge, P. Genetic and genomic tools to improve drought tolerance in wheat. J. Exp. Bot. 2010, 61, 3211–3222. [Google Scholar] [CrossRef] [Green Version]
- Grigorova, B.; Vaseva, I.; Demirevska, K.; Feller, U. Combined drought and heat stress in wheat: Changes in some heat shock proteins. Biol. Plant. 2011, 55, 105–111. [Google Scholar] [CrossRef]
- Keles, Y.; Oncel, I. Response of antioxidative defence system to temperature and water stress combinations in wheat seedlings. Plant Sci. 2002, 163, 783–790. [Google Scholar] [CrossRef]
- Barnabás, B.; Jäger, K.; Fehér, A. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ. 2008, 31, 11–38. [Google Scholar] [CrossRef]
- Rizhsky, L.; Liang, H.; Mittler, R. The combined effect of drought stress and heat shock on gene expression in tobacco. Plant Physiol. 2002, 130, 1143–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartels, D.; Sunkar, R. Drought and salt tolerance in plants. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Lobell, B.D.; Gourdji, M.S. The influence of climate change on global crop productivity. Plant Physiol. 2012, 160, 1686–1697. [Google Scholar] [CrossRef] [Green Version]
- Alaei, M.; Farboodi, M.; Khorshidi, M.B.; Zaeifizadeh, M. Durum wheat land race screening for drought tolerance. Middle-East J. Sci. Res. 2010, 63, 289–292. [Google Scholar]
- Dixit, S.; Singh, A.; Kumar, A. Rice breeding for high grain yield under drought: A strategic solution to a complex problem. Int. J. Agron. 2014. [Google Scholar] [CrossRef]
- Kilic, H.; Yagbasanlar, T. The effect of drought stress on grain yield, yield components and some quality traits of durum wheat (Triticum turgidum ssp. durum) cultivars. Not. Bot. Horti. Agrobot. Cluj-Napoca 2010, 38, 164–170. [Google Scholar]
- Nawaz, H.; Hussain, N.; Yasmeen, A. Growth, yield and antioxidants status of wheat (Triticum aestivum L.) cultivars under water deficit conditions. Pak. J. Agric. Sci. 2015, 52, 953–959. [Google Scholar]
- Rucker, K.S.; Kevin, C.K.; Holbrook, C.C.; Hook, J.E. Identification of peanut genotypes with improved drought avoidance traits. Peanut Sci. 1995, 22, 14–18. [Google Scholar] [CrossRef] [Green Version]
- Shao, H.B.; Chu, L.Y.; Jaleel, C.A.; Zhao, C.X. Water-deficit stress-induced anatomical changes in higher plants. Comptes Rendus 2008, 331, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Tardieau, F.; Reymond, M.; Hamard, P.; Granier, C.; Muller, B. Spatial distributions of expansion rate, cell division rate and cell size in maize leaves: A synthesis of the effects of soil water status, evaporative demand and temperature. J. Exp. Bot. 2000, 51, 1505–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, L.; Wang, R.-G.; Mao, G.; Koczan, J.M. Identification of drought tolerance determinants by genetic analysis of root response to drought stress and abscisic acid. Plant Physiol. 2006, 142, 1065–1074. [Google Scholar] [CrossRef] [Green Version]
- Sharp, R.E.; Wu, Y.J.; Voetberg, G.S.; Saab, I.N.; Lenoble, M.E. Confirmation that abscisic-acid accumulation is required for maize primary root elongation at low water potentials. J. Exp. Bot. 1994, 45, 1743–1751. [Google Scholar] [CrossRef]
- Czyczyło-Mysza, I.; Marcinska, I.; Skrzypek, E.; Chrupek, M.; Grzesiak, S.; Hura, T.; Stojałowski, S.; Myskow, B.; Milczarski, P.; Quarrie, S. Mapping QTLs for yield components and chlorophyll a fluorescence parameters in wheat under three levels of water availability. Plant Genet. Resour. C. 2011, 9, 291–295. [Google Scholar] [CrossRef]
- Chachar, M.H.; Chachar, N.A.; Chachar, Q.; Mujtaba, S.M.; Chachar, S.; Chachar, Z. Physiological characterization of six wheat genotypes for drought tolerance. Int. J. Res. Granthaalayah 2016, 4, 184–196. [Google Scholar]
- Srivalli, S.; Khanna-Chopra, R. Delayed wheat flag leaf senescence due to removal of spikelets is associated with increased activities of leaf antioxidant enzymes, reduced glutathione/oxidized glutathione ratio and oxidative damage to mitochondrial proteins. Plant Physiol. Biochem. 2009, 47, 663–670. [Google Scholar] [CrossRef]
- Zhu, C.; Zhu, J.; Zeng, Q.; Liu, G.; Xie, Z.; Tang, H.; Cao, J.; Zhao, X. Elevated CO2 accelerates flag leaf senescence in wheat due to ear photosynthesis which causes greater ear nitrogen sink capacity and ear carbon sink limitation. Funct. Plant Biol. 2009, 36, 291–299. [Google Scholar] [CrossRef]
- Gregersen, P.L.; Holm, P.B.; Krupinska, K. Leaf senescence and nutrient remobilization in barley and wheat. Plant Biol. 2008, 10, 37–49. [Google Scholar] [CrossRef]
- Falqueto, A.R.; Cassol, D.; De Magalhães, J.M.A.; De Oliveira, A.C.; Bacarin, M.A. Physiological analysis of leaf senescence of two rice cultivars with different yield potential. Pesqui. Agropecu. Bras. 2009, 44, 695–700. [Google Scholar] [CrossRef]
- Saeedipour, S. Effect of drought at the post-anthesis stage on remobilization of carbon reserves in two wheat cultivars differing in senescence properties. Int. J. Plant Physiol. Biochem. 2011, 3, 15–24. [Google Scholar]
- Yang, J.; Zhang, J.; Huang, Z.; Zhu, Q.; Wang, L. Remobilization of carbon reserves is improved by controlled soil-drying during grain filling of wheat. Crop Sci. 2000, 40, 1645–1655. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Wang, Z.; Zhu, Q.; Liu, L. Water deficit-induced senescence and its relationship to the remobilization of pre-stored carbon in wheat in grain filling. Agron. J. 2001, 93, 196–206. [Google Scholar] [CrossRef]
- Waters, B.M.; Uauy, C.; Dubcovsky, J.; Grusak, M.A. Wheat (Triticum aestivum) NAM proteins regulate the translocation of iron, zinc, and nitrogen compounds from vegetative tissues to grain. J. Exp. Bot. 2009, 60, 4263–4274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedigheh, H.G.; Mortazavian, M.; Norouzian, D.; Atyabil, M.; Akbarzadeh, A.; Hasanpoor, K.; Ghorbani, M. Oxidative stress and leaf senescence. BMC Res. Notes 2011, 4, 477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishida, H.; Makino, A.; Mae, T. Fragmentation of the large subunit of ribulose-1,5-bisphosphate carboxylase by reactive oxygen species occurs near Gly-329. J. Biol. Chem. 1999, 274, 5222–5226. [Google Scholar] [CrossRef] [Green Version]
- Bray, E.A. Classification of genes differentially expressed during water-deficit stress in Arabidopsis thaliana: An analysis using microarray and differential expression data. Ann. Bot. 2002, 89, 803–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habash, D.Z.; Kehel, Z.; Nachit, M. Genomic approaches for designing durum wheat ready for climate change with a focus on drought. J. Exp. Bot. 2009, 60, 2805–2815. [Google Scholar] [CrossRef] [PubMed]
- Barnabás, B.; Fehér, A. Az embriófejlődés és a szemfeltöltődés molekuláris folyamatai a vízhiányban. In A búza Nemesbítésének Tudománya; Dudits, D., Ed.; MTA Szegedi Biológiai Központ-Winter Fair Kft: Szeged, Hungary, 2006; pp. 180–190. [Google Scholar]
- Jäger, K.; Fábián, A.; Barnabás, B. A tartós vízhiány és a magas hőmérséklet hatása a búza pollen életképességére. Martonvásár 2013, 55, 24–25. [Google Scholar]
- Gao, X.; Francis, D.; Ormrod, J.C.; Bennett, M.D. Changes in cell number and cell division activity during endosperm development in allohexaploid wheat, Triticum aestivum L. J. Exp. Bot. 1992, 43, 1603–1609. [Google Scholar] [CrossRef]
- Yang, J.C.; Zhang, J.H.; Wang, Z.Q.; Zhu, Q.S.; Liu, L. Involvement of abscisic acid and cytokinins in the senescence and remobilization of carbon reserves in wheat subjected to water stress during grain filling. Plant Cell Environ. 2003, 26, 1621–1631. [Google Scholar] [CrossRef]
- Jäger, K.; Fábián, A.; Barnabás, B. Effect of water deficit and elevate temperature on pollen development of drought sensitive and tolerant winter wheat (Triticum aestivum L.) genotypes. Acta Biol. Szeged. 2008, 52, 67–71. [Google Scholar]
- Bányai, J.; Maccaferri, M.; Cané, M.A.; Monostori, I.; Spitkó, T.; Kuti, C.; Mészáros, K.; Láng, L.; Pál, M.; Karsai, I. Phenotypical and physiological study of near-isogenic durum wheat lines under contrasting water regimes. S. Afr. J. Bot. 2017, 108, 248–255. [Google Scholar] [CrossRef]
- Varshney, R.K.; Graner, A.; Sorrells, M.E. Genomics-assisted breeding for crop improvement. Trends Plant Sci. 2005, 10, 621–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kooke, R.; Wijnker, E.; Keurentjes, J.J.B. Backcross populations and near isogenic lines. In Quantitative Trait Loci (QTL): Methods and Protocols; Methods in Molecular Biology; Rifkin, S.A., Ed.; Humana Press: New York, NY, USA, 2012; Volume 871, pp. 1–14. [Google Scholar]
- Sevanthi, A.M.; Prakash, C.; Shanmugavadivel, P.S. Recent progress in rice varietal development for abiotic stress tolerance. In Advances in Rice Research for Abiotic Stress Tolerance; Hasanuzzaman, M., Ed.; Woodhead Publishing: Cambridge, UK, 2019; pp. 47–68. [Google Scholar]
- Tsujimoto, H. Production of near-Isogenic lines and marked monosomic lines in common wheat (Triticum aestivum) cv. Chinese Spring. Heredity 2001, 92, 254–259. [Google Scholar] [CrossRef] [Green Version]
- Mia, M.S.; Liu, H.; Wang, X.; Yan, G. Multiple near-isogenic lines targeting a QTL hotspot of drought tolerance showed contrasting performance under post-anthesis water stress. Front. Plant Sci. 2019, 10, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naser, M.A.; Khosla, R.; Longchamps, L.; Dahal, S. Using NDVI to differentiate wheat genotypes productivity under dryland and irrigated conditions. Remote Sens. 2020, 12, 824. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.; Jiang, J.; Gao, Y.; Krienke, B.; Wang, M.; Zhong, K.; Cao, Q.; Tian, Y.; Zhu, Y.; Cao, W.; et al. Wheat growth monitoring and yield estimation based on multi-rotor unmanned aerial vehicle. Remote Sens. 2020, 12, 508. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, A.W.; Lindsey, L.E.; Harrison, S.K.; Paul, P.A. Estimating wheat yield with normalized difference vegetation index and fractional green canopy cover. Crop Forage Turfgrass Manag. 2018, 4, 180026. [Google Scholar] [CrossRef] [Green Version]
- Vannoppen, A.; Gobin, A.; Kotova, L.; Top, S.; De Cruz, L.; Viksna, A.; Aniskevich, S.; Bobylev, L.; Buntemeyer, L.; Caluwaerts, S.; et al. Wheat yield estimation from NDVI and regional climate models in Latvia. Remote Sens. 2020, 12, 2206. [Google Scholar] [CrossRef]
- Dodig, D.; Savic, J.; Kandic, V.; Zoric, M.; Radovic, B.V.; Popovic, A.; Quarrie, S. Responses of wheat plants under post-anthesis stress induced by defoliation: I. Contribution of agro-physiological traits to grain yield. Exp. Agric. 2015. [Google Scholar] [CrossRef] [Green Version]
- Cossani, C.M.; Slafer, G.A.; Savin, R. Yield and biomass in wheat and barley under a range of conditions in a Mediterranean site. Field Crops Res. 2009, 112, 205–2013. [Google Scholar] [CrossRef]
- Cruz-Aguado, J.A.; Rodés, R.; Pérez, J.P.; Dorado, M. Morphological characteristics and yield components associated with accumulation and loss of dry matter in internodes of wheat. Field Crops Res. 2000, 66, 129–139. [Google Scholar] [CrossRef]
- Ehdaie, B.; Alloush, G.A.; Madore, M.A.; Waines, J.G. Genotypic variation for stem reserves and mobilization in wheat. I. Post anthesis changes in internode dry matter. Crop Sci. 2006, 46, 735–746. [Google Scholar] [CrossRef]
- Majer, P.; Sass, L.; Lelley, T.; Cseuz, L.; Vass, I.; Dudits, D.; Pauk, J. Testing drought tolerance of wheat by a complex stress diagnostic system installed in greenhouse. Acta Biol. Szeged. 2008, 52, 97–100. [Google Scholar]
- Korohou, T.; Okinda, C.; Li, H.; Cao, Y.; Nyalala, I.; Huo, L.; Potcho, M.; Li, X.; Ding, Q. Wheat grain yield estimation based on image morphological properties and wheat biomass. J. Sens. 2020, 2020, 1571936. [Google Scholar] [CrossRef]
- Bennett, D.; Reynolds, M.; Mullan, D.; Izanloo, A.; Kuchel, H.; Langridge, P.; Schnurbusch, T. Detection of two major grain yield QTL in bread wheat (Triticum aestivum L.) under heat, drought and high yield potential environments. Theor. Appl. Genet. 2012, 125, 1473–1485. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Liu, D.C.; Guo, X.L.; Yang, W.L.; Sun, J.Z.; Wang, D.W.; Zhang, A. Genomic distribution of quantitative trait loci for yield and yield-related traits in common wheat. J. Integr. Plant Biol. 2010, 52, 996–1007. [Google Scholar] [CrossRef] [PubMed]
- Pinto, R.S.; Reynolds, M.P.; Mathews, K.L.; Mcintyre, C.L.; Olivares–Villegas, J.J.; Chapman, S.C. Heat and drought adaptive QTL in a wheat population designed to minimize confounding agronomic effects. Theor. Appl. Genet. 2010, 121, 1001–1021. [Google Scholar] [CrossRef] [Green Version]
- Bonneau, J.; Taylor, J.; Parent, B.; Bennett, D.; Reynolds, M.; Feuillet, C.; Langridge, P.; Mather, D. Multi-environment analysis and improved mapping of a yield-related QTL on chromosome 3B of wheat. Theor. Appl. Genet. 2013, 126, 747–761. [Google Scholar] [CrossRef]
- Shukla, S.; Mishra, B.K.; Mishra, R.; Siddiqui, A.; Pandey, R.; Rastogi, A. Comparative study for stability and adaptability through different models in developed high thebaine lines of opium poppy (Papaver somniferum L.). Ind. Crops Prod. 2015, 74, 875–886. [Google Scholar] [CrossRef]
- Dudziak, K.; Zapalska, M.; Börner, A.; Szczerba, H.; Kowalczyk, K.; Nowak, M. Analysis of wheat gene expression related to the oxidative stress response and signal transduction under short-term osmotic stress. Sci. Rep. 2019, 9, 2743. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Cui, Z.; Ni, Y.; Zheng, M.; Yang, D.; Jin, M.; Chen, J.; Wang, Z.; Yin, Y. Plant density effect on grain number and weight of two winter wheat cultivars at different spikelet and grain positions. PLoS ONE 2016. [Google Scholar] [CrossRef]
- Whaley, Y.M.; Sparkes, D.L.; Foulkes, M.J.; Spink, J.H.; Semere, T.; Scott, R.K. The physiological response of winter wheat to reductions in plant density. Ann. Appl. Biol. 2000, 137, 165–177. [Google Scholar] [CrossRef]
- Wood, G.A.; Welsh, J.P.; Godwin, R.J.; Taylor, J.C.; Earl, R.; Knight, S.M. Real-time measures of canopy size as a basis for spatially varying nitrogen applications to winter wheat sown at different seed rates. Biosyst. Eng. 2003, 84, 513–531. [Google Scholar] [CrossRef] [Green Version]
- Valério, I.P.; De Carvalho, F.I.F.; Benin, G.; Da Silveira, G.; Da Silva, J.A.G.; Nornberg, R.; Hagemann, T.; De Souza Luche, H.; De Oliveira, C.A. Seeding density in wheat: The more, the merrier? Sci. Agric. 2013, 40, 176–184. [Google Scholar] [CrossRef]
- Ozturk, A.; Caglar, O.; Bulut, S. Growth and yield response of facultative wheat to winter sowing, freezing sowing and spring sowing at different seeding rates. J. Agron. Crop Sci. 2006, 192, 10–16. [Google Scholar] [CrossRef]
- Bastos, L.M.; Carciochi, W.; Lollato, R.P.; Jaenisch, B.R.; Rezende, C.R.; Schwalbert, R.; Prasad, P.V.V.; Zhang, G.; Fritz, A.K.; Foster, C.; et al. Winter wheat yield response to plant density as a function of yield environment and tillering potential: A review and field studies. Front. Plant Sci. 2020, 11, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.J. The role of the plant antioxidant system in drought tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Pour-Benab, S.M.; Fabriki-Ourang, S.; Mehrabi, A.A. Expression of dehydrin and antioxidant genes and enzymatic antioxidant defense under drought stress in wild relatives of wheat. Biotechnol. Equip. 2019, 33, 1063–1073. [Google Scholar] [CrossRef] [Green Version]
- Salekjalali, M.; Haddad, R.; Jafari, B. Effects of soil water shortages on the activity of antioxidant enzymes and the contents of chlorophylls and proteins in barley. Am. Eurasian J. Agric. Environ. Sci. 2012, 12, 57–63. [Google Scholar]
- Huseynova, I.M.; Suleymanov, S.Y.; Rustamova, S.M. Response of photosynthetic apparatus and antioxidant defense systems in Triticum aestivum L. genotypes subjected to drought stress. Proc. ANAS (Biol. Sci.) 2010, 65, 49–59. [Google Scholar]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine function in plants: Metabolism, regulation on development, and roles in abiotic stress responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef] [PubMed]
- Aldesuquy, H.; Haroun, S.; Abo-Hamed, S.; El Saied, A.-W. Involvement of spermine and spermidine in the control of productivity and biochemical aspects of yielded grains of wheat plants irrigated with waste water. Egypt. J. Basic Appl. Sci. 2014, 1, 16–28. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Liang, H.; Lv, X.; Liu, D.; Wen, X.; Liao, Y. Effect of polyamines on the grain filling of wheat under drought stress. Plant Physiol. Biochem. 2016, 100, 113–129. [Google Scholar] [CrossRef]
- Sahitya, U.L.; Krishna, M.S.R.; Deepthi, R.S.; Prasad, G.S.; Kasim, D.P. Seed antioxidants interplay with drought stress tolerance indices in chilli (Capsicum annuum L) seedlings. Biomed. Res. Int. 2018, 2018, 1605096. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Al Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Maccaferri, M.; Sanguineti, M.C.; Corneti, S.; Ortega, J.L.; Salem, M.B.; Bort, J.; DeAmbrogio, E.; del Moral, L.F.; Demontis, A.; El-Ahmed, A.; et al. Quantitative Trait Loci for grain yield and adaptation of durum wheat (Triticum durum Desf.) across a wide range of water availability. Genetics 2008, 178, 489–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stage of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Pál, M.; Kovács, V.; Vida, G.; Szalai, G.; Janda, T. Changes induced by powdery mildew in the salicylic acid and polyamine contents and the antioxidant enzyme activities of wheat lines. Eur. J. Plant Pathol. 2013, 135, 35–47. [Google Scholar] [CrossRef]
- Rosyara, U.R.; Pant, K.; Duveiller, E.; Sharma, R.C. Variation in chlorophyll content, anatomical traits and agronomic performance of wheat genotypes differing in spot blotch resistance under natural epiphytotic conditions. Australas. Plant Pathol. 2007, 36, 245–251. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year (Y) | Genotype (G) | Treatment (T) | Density (D) | G × Y | G × T | G × D | Y × T | Y × D | T × D | Y × G × T | Y × G × D | Y × T × D | G × T × D | Y × G × T × D | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| d.f. | 1 | 7 | 1 | 1 | 7 | 7 | 7 | 1 | 1 | 1 | 7 | 7 | 1 | 7 | 7 |
| HD 1 | 2156.85 *** | 502.57 *** | 3.71 | 525.87 *** | 27.06 *** | 2.02 | 6.53 *** | 6.77 * | 0.08 | 12.71 *** | 3.14* | 5.26 *** | 1.82 | 1.27 | 0.48 |
| FD 2 | 3167.93 *** | 332.91 *** | 1.88 | 393.75 *** | 20.86 *** | 1.24 | 3.97 *** | 8.72 ** | 6.44 * | 9.22 ** | 2.53 * | 3.78 *** | 1.88 | 1.43 | 1.12 |
| MD 3 | 3798.87 *** | 157.56 *** | 377.06 *** | 119.30 *** | 25.58 *** | 4.84 *** | 3.81 *** | 21.11 *** | 27.96 *** | 20.32 *** | 3.21 ** | 4.53 *** | 26.16 *** | 1.56 | 1.03 |
| GY 4 | 72.11 *** | 30.10 *** | 247.04 *** | 109.42 *** | 8.23 *** | 1.58 | 2.62 * | 78.62 *** | 32.88 *** | 37.09 *** | 2.24 * | 1.24 | 33.14 *** | 2.12 * | 2.75 ** |
| TGW 5 | 133.71 *** | 88.35 *** | 685.52 *** | 56.52 *** | 20.26 *** | 9.06 *** | 2.87 ** | 487.73 *** | 0.08 | 0.52 | 2.46 ** | 0.83 | 1.95 | 2.81 ** | 3.02 ** |
| SNM 6 | 826.46 *** | 8.50 *** | 2.02 | 0.08 | 1.27 | 0.27 | 1.09 | 0.03 | 2.84 | 0.94 | 0.79 | 0.16 | 0.95 | 1.06 | 0.79 |
| SWM 7 | 132.34 *** | 27.68 *** | 414.71 *** | 133.16 *** | 1.76 | 1.42 | 1.73 | 0.71 | 3.46 | 0.19 | 1.03 | 0.95 | 24.78 *** | 0.51 | 0.48 |
| SKNM 8 | 644.12 *** | 53.33 *** | 7.97 | 24.60 | 0.45 | 1 | 9.62 ** | 3.14 | 0.38 | 0.04 | 1.39 | 1.43 | 0.10 | 0.55 | 0.98 |
| BSM 9 | 1.34 | 0.58 | 0.28 | 0.07 | 0.69 | 0.40 | 0.43 | 0.11 | 0.26 | 0.51 | 0.91 | 0.46 | 0.15 | 0.91 | 0.28 |
| ASM 10 | 0.37 | 0.59 | 1.17 | 0 | 0.89 | 0.77 | 0.68 | 1.66 | 0.03 | 0.07 | 0.56 | 0.83 | 0.03 | 0.85 | 0.75 |
| SL 11 | 4.06 * | 13.07 *** | 0.68 | 31.23 *** | 4.09 *** | 1.90 | 1.17 | 0 | 26.32 *** | 15.52 *** | 4.66 *** | 3.50 * | 4.06 * | 0.76 | 1.99 |
| SW 12 | 600.45 *** | 220.04 *** | 34.51 *** | 16.23 *** | 44.47 *** | 1.77 | 7.44 *** | 600.45 *** | 30.68 *** | 28.85 *** | 3.14 | 1.70 | 8.14 *** | 1.38 | 0.99 |
| FLC 13 | 876.31 *** | 18.48 *** | 16.41 *** | 13.94 *** | 12.82 *** | 1.77 | 1.16 | 27.49 *** | 4.27 * | 31.21 *** | 1.69 | 0.74 | 0.1 | 1.34 | 0.74 |
| BE 14 | 978.21 *** | 128.18 *** | 52.24 *** | 16.60 *** | 32.02 *** | 3.31 * | 1.20 | 12.97 *** | 0.58 | 41.13 *** | 2.88 * | 2.00 | 2.17 | 1.21 | 1.66 |
| TE 15 | 979.14 *** | 102.85 *** | 62.92 *** | 22.70 *** | 35.24 *** | 0.28 | 0.41 | 7.33 ** | 1.26 | 22.70 *** | 1.18 | 0.55 | 1.31 | 2.02 | 1.37 |
| EL 16 | 101.32 *** | 4.38 *** | 25.08 *** | 8.56 *** | 0.72 | 0.43 | 0.41 | 0.42 | 0.02 | 0.89 | 0.13 | 0.15 | 0.02 | 0.17 | 0.07 |
| FTN 17 | 0.41 | 9.97 *** | 0.35 | 7.77 ** | 8.19 *** | 2.27 * | 1.12 * | 0.29 | 1.68 | 0.11 | 1.76 | 0.38 | 0.15 | 1.04 | 0.26 |
| FLA 18 | 28.45 | 71.72 | 0 | 2.01 | 17.07 | 3.21 | 2.28 | 0.46 | 0.15 | 0.06 | 2.61 | 0.70 | 0.21 | 1.09 | 0.82 |
| SPAD45 19 | 1610.61 *** | 18.52 *** | 2 | 36.93 *** | 1.86 | 1.75 | 1.21 | 0.25 | 0.20 | 11.43 *** | 1.06 | 1.04 | 1.23 | 0.87 | 1.27 |
| SPAD65 20 | 220.84 *** | 16.70 *** | 0 | 7.10 | 3.14 | 1.02 | 0.66 | 0.11 | 1.11 | 1.83 | 0.65 | 1.06 | 1.40 | 1.44 | 0.46 |
| SPAD77 21 | 202.70 *** | 26.52 *** | 70.18 *** | 28.00 * | 10.75 *** | 2.29 * | 0.68 | 0.73 | 52.70 *** | 3.25 | 1.91 | 0.18 | 3.79 | 1.14 | 1.38 |
| SPAD83 22 | 14.41 *** | 11.60 *** | 336.60 *** | 19.00 * | 5.23 *** | 3.15 ** | 0.35 | 1.51 | 55.48 *** | 18.87 *** | 1.20 | 0.23 | 0 | 0.46 | 0.49 |
| SPAD85 23 | 2.63 | 3.70 | 23.07 | 19.43 *** | 3.66 *** | 1.20 | 1.03 | 11.20 *** | 27.94 *** | 0.56 | 1.40 | 0.62 | 10.69 ** | 0.21 | 0.96 |
| NDVI45 24 | 1.05 | 16.91 *** | 0.02 | 0.02 | 3.82 *** | 0.87 | 0.15 | 0.13 | 0.20 | 0.01 | 0.67 | 0.49 | 0.06 | 0.33 | 0.64 |
| NDVI65 25 | 1.18 | 2.13 * | 0.02 | 0.04 | 1.24 | 0.09 | 0.06 | 0.02 | 0.07 | 0.01 | 0.15 | 0.07 | 0.01 | 0.08 | 0.22 |
| NDVI83 26 | 30.83 *** | 7.45 *** | 5.54 * | 56.25 *** | 3.86 *** | 0.48 | 0.88 | 0.35 | 37.11 *** | 0.71 | 0.28 | 0.15 | 0.16 | 0.49 | 0.49 |
| RWC 27 | 858.81 *** | 0.92 | 45.38 *** | 1.47 | 1.05 | 0.48 | 0.36 | 0.56 | 0.53 | 0.08 | 0.41 | 1.47 | 3.92 * | 0.56 | 0.68 |
| WW65 28 | 1.32 | 3.60 ** | 0.18 | 0.26 | 1.74 | 0.20 | 0.24 | 0.11 | 0.02 | 0.02 | 0.60 | 0.12 | 0.01 | 0.24 | 0.36 |
| DW65 29 | 407.65 *** | 1.84 | 49.66 *** | 34.36 *** | 1.16 | 1.26 | 0.99 | 4.04 | 1.43 | 4.60 | 1.23 | 0.49 | 0.19 | 0.87 | 0.63 |
| WW91 30 | 202.57 *** | 3.50 | 47.54 *** | 26.68 *** | 1.38 | 0.42 | 0.76 | 45.45 *** | 6.93 * | 7.22 * | 0.73 | 0.54 | 3.26 | 1.19 | 0.08 |
| DW91 31 | 261.27 *** | 3.35 | 39.82 *** | 17.72 *** | 1.48 | 0.34 | 0.65 | 40.35 *** | 2.94 | 6.07 * | 0.61 | 0.75 | 4.95 | 1.08 | 0.12 |
| HD 1 | FD 2 | MD 3 | SPAD45 4 | SPAD65 5 | SPAD77 6 | SPAD83 7 | SPAD85 8 | NDVI45 9 | NDVI65 10 | NDVI83 11 | AUSDC 12 | AUVIC 13 | FLA 14 | |||||||||||||||
| W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | |
| NIL1−− | 150.56 *** | 150.83 | 156 *** | 156.36 | 194.72 ** | 193.27 | 42.10 | 41.16 | 48.30 | 48.48 | 37.41 | 30.78 | 29.32 | 21.21 | 10.92 | 9.80 | 0.3498 | 0.3546 | 0.5515 | 0.5502 | 0.3397 | 0.3279 | 1597.95 | 1441.53 | 14.86 | 15.31 | 27.65 ** | 26.22 |
| NIL++ | 149.09 | 148.94 | 154.44 | 154.39 | 194.36 | 193.14 | 42.97 | 42.91 | 48.85 | 48.89 | 43.81 | 41.57 | 34.40 *** | 26.95 ** | 10.72 | 10.73 | 0.3585 | 0.3563 | 0.5515 | 0.5532 | 0.3900 *** | 0.3800* | 1839.04 *** | 1734.17 ** | 15.57 | 15.65 | 23.55 | 24.48 |
| LSD5% | 0.45 | 2.27 | 0.66 | 2.15 | 0.19 | 1.91 | 1.06 | 3.06 | 1.37 | 2.78 | 2.41 | 3.38 | 1.44 | 3.43 | 2.68 | 2.10 | 0.0082 | 0.0247 | 0.0044 | 0.0115 | 0.0179 | 0.0429 | 72.74 | 175.27 | 0.45 | 1.69 | 2.52 | 3.67 |
| LSD1% | 0.67 | 3.36 | 0.97 | 3.18 | 0.28 | 2.82 | 1.58 | 4.53 | 2.04 | 4.11 | 3.58 | 5.01 | 2.14 | 5.08 | 3.96 | 3.12 | 0.01223 | 0.0366 | 0.0065 | 0.0170 | 0.0265 | 0.0635 | 107.65 | 259.40 | 0.67 | 2.51 | 3.74 | 5.44 |
| LSD0.1% | 1.04 | 5.19 | 1.50 | 4.91 | 0.43 | 4.37 | 2.44 | 7.01 | 3.15 | 6.36 | 5.53 | 7.74 | 3.30 | 7.85 | 6.13 | 4.82 | 0.0189 | 0.0566 | 0.0101 | 0.0263 | 0.0409 | 0.0982 | 166.36 | 400.86 | 1.03 | 3.87 | 5.78 | 8.41 |
| FTN 15 | FLC 16 | BE 17 | TE 18 | EL 19 | GY 20 | SKNM 21 | SNM 22 | SWM 23 | BSM 24 | ASM 25 | TGW 26 | SL 27 | SW 28 | |||||||||||||||
| W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | |
| NIL1−− | 370.59 | 378.34 | 40.59 | 40.90 | 55.68 | 54.14 | 61.79 | 59.85 | 5.93 | 5.65 | 3.02 | 2.67 | 13.54 | 13.51 | 35.25 | 34.83 | 1.88 | 1.44 | 0.7582 | 0.8586 | 0.5433 | 0.5419 | 35.43 | 43.63 | 7.01 | 6.99 | 2.89 | 3.06 |
| NIL++ | 400.63 ** | 392.30 ** | 43.13 ** | 43.52 * | 62.66 ** | 62.05 *** | 69.33 *** | 67.82 *** | 6.21 | 5.81 | 3.42 *** | 3.15 *** | 13.42 | 13.32 | 37.52 ** | 37.15 *** | 2.23 ** | 1.64 ** | 0.6176 | 0.6861 | 0.5433 | 0.5070 | 39.82 *** | 46.50 ** | 7.01 | 7.02 | 3.05 *** | 3.16 * |
| LSD5% | 18.59 | 6.60 | 1.15 | 2.36 | 3.16 | 2.86 | 2.91 | 2.69 | 0.10 | 0.22 | 0.15 | 0.20 | 0.22 | 0.33 | 1.02 | 0.98 | 0.15 | 0.13 | 0.4992 | 0.3313 | 0.1889 | 0.1608 | 1.12 | 1.70 | 0.03 | 0.09 | 0.05 | 0.09 |
| LSD1% | 27.52 | 9.77 | 1.71 | 3.50 | 4.67 | 4.23 | 4.31 | 3.98 | 0.16 | 0.33 | 0.23 | 0.30 | 0.32 | 0.48 | 1.51 | 1.45 | 0.23 | 0.19 | 0.7388 | 0.4902 | 0.2796 | 0.2380 | 1.66 | 2.52 | 0.05 | 0.14 | 0.07 | 0.24 |
| LSD0.1% | 42.53 | 15.11 | 2.64 | 5.41 | 7.23 | 6.54 | 6.67 | 6.15 | 0.24 | 0.52 | 0.35 | 0.47 | 0.50 | 0.75 | 2.33 | 2.24 | 0.35 | 0.30 | 1.1417 | 0.7576 | 0.4321 | 0.3678 | 2.57 | 3.90 | 0.08 | 0.22 | 0.11 | 0.37 |
| HL 29 | P 30 | G 31 | Z 32 | WW65 33 | WW91 34 | DW65 35 | DW91 36 | RWC 37 | APX 38 | GPX 39 | PUT 40 | SPD 41 | SPN 42 | |||||||||||||||
| W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | |
| NIL1−− | 76.74 | 82.54 | 17.31 *** | 16.76 | 36.18 ** | 35.09 | 62.09 | 64.11 | 0.1444 | 0.1439 | 0.0340 | 0.0339 | 0.0195 | 0.0140 | 0.0240 | 0.0161 | 91.80 | 87.62 | 824.89 | 576.57 | 3040.49 | 4899.19 | 192.87 | 355.05 | 394.47 | 524.73 | 217.78 | 296.51 |
| NIL++ | 78.93 | 82.57 | 16.16 | 17.07 | 34.01 | 35.39 | 61.34 | 65.91 | 0.1437 | 0.1442 | 0.0389 * | 0.0380 | 0.0186 | 0.0161 | 0.0281* | 0.0280 | 91.07 | 87.52 | 982.15 | 504.77 | 3438.56 | 4840.85 | 177.70 | 460.20* | 419.43 | 572.83 | 305.50 | 342.78 |
| LSD5% | 1.54 | 1.36 | 0.45 | 1.39 | 1.17 | 2.36 | 1.01 | 2.87 | 0.0039 | 0.0054 | 0.0045 | 0.0094 | 0.0014 | 0.0039 | 0.0034 | 0.0069 | 1.75 | 2.88 | 282.90 | 78.58 | 1331.83 | 3291.57 | 86.88 | 91.89 | 24.13 | 171.95 | 113.38 | 59.48 |
| LSD1% | 2.28 | 2.01 | 0.67 | 2.06 | 1.73 | 3.50 | 1.49 | 4.25 | 0.0058 | 0.0080 | 0.0067 | 0.0139 | 0.0021 | 0.0058 | 0.0051 | 0.0102 | 2.59 | 4.26 | 519.22 | 144.23 | 2444.38 | 6041.20 | 159.46 | 168.66 | 44.30 | 315.60 | 208.10 | 109.05 |
| LSD0.1% | 3.52 | 3.11 | 1.04 | 3.19 | 2.67 | 5.41 | 2.31 | 6.56 | 0.0090 | 0.0124 | 0.0104 | 0.0215 | 0.0033 | 0.0090 | 0.0078 | 0.0158 | 4.01 | 6.58 | 1148.88 | 319.14 | 5408.61 | 13367.16 | 352.84 | 373.20 | 98.03 | 698.31 | 460.47 | 241.29 |
| HD 1 | FD 2 | MD 3 | SPAD45 4 | SPAD65 5 | SPAD77 6 | SPAD83 7 | SPAD85 8 | NDVI45 9 | NDVI65 10 | NDVI83 11 | AUSDC 12 | AUVIC 13 | ||||||||||||||
| W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | |
| NIL1−− | 146.50 | 146.09 | 150.43 | 150.00 | 189.09 | 186.42 | 52.21 | 52.23 | 48.30 | 51.871875 | 46.03 | 41.92 | 29.45 | 21.49 | 12.13 | 9.19 | 0.3527 | 0.3522 | 0.5574 | 0.5557 | 0.3829 | 0.3700 | 2170.42 | 2020.28 | 24.89 | 22.14 |
| NIL++ | 146.05 | 145.71 | 150 | 149.62 | 189.70 | 188.04 | 53.67 | 53.18 | 48.85 | 54.20 * | 47.95 | 44.94 | 31.47 | 24.81 ** | 13.31 | 10.03 | 0.3527 | 0.3523 | 0.5557 | 0.5556 | 0.4000 ** | 0.3900 * | 2229.15 | 2109.61 ** | 26.28 * | 22.54 |
| LSD5% | 1.23 | 1.37 | 1.12 | 1.31 | 1.15 | 1.84 | 1.14 | 1.14 | 1.37 | 1.63 | 2.01 | 2.09 | 2.38 | 1.66 | 1.25 | 0.96 | 0.0032 | 0.0024 | 0.0047 | 0.0039 | 0.0087 | 0.0156 | 98.81 | 45.39 | 0.96 | 0.86 |
| LSD1% | 1.82 | 2.03 | 1.66 | 1.95 | 1.71 | 2.72 | 1.69 | 1.68 | 2.04 | 2.42 | 2.97 | 3.10 | 3.52 | 2.47 | 1.85 | 1.42 | 0.0048 | 0.0036 | 0.0070 | 0.0059 | 0.0129 | 0.0231 | 146.23 | 67.18 | 1.41 | 1.28 |
| LSD0.1% | 2.81 | 3.14 | 2.56 | 3.01 | 2.64 | 4.21 | 2.61 | 2.61 | 3.15 | 3.74 | 4.59 | 4.79 | 5.44 | 3.81 | 2.86 | 2.19 | 0.0074 | 0.0055 | 0.0108 | 0.0091 | 0.0199 | 0.0357 | 225.98 | 103.82 | 2.19 | 1.98 |
| FLA 14 | FTN 15 | FLC 16 | BE 17 | TE 18 | EL 19 | GY 20 | SKNM 21 | SNM 22 | SWM 23 | BSM 24 | ASM 25 | TGW 26 | ||||||||||||||
| W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | |
| NIL1−− | 26.41 | 27.37 | 361.31 | 362.78 | 51.43 | 48.33 | 65.57 | 63.97 | 73.59 | 70.55 | 6.73 | 6.31 | 3.59 | 2.58 | 15.25 | 14.98 | 44.71 | 44.26 | 2.11 | 1.71 | 0.6125 | 0.5146 | 0.4817 | 0.8330 | 37.67 | 38.83 |
| NIL++ | 28.02 | 27.49 | 423.81 ** | 407.77 *** | 52.84 | 50.47 * | 73.50 ** | 68.67 * | 80.00 ** | 76.02 ** | 7.11 ** | 6.64 *** | 4.40 ** | 3.22 *** | 15.21 | 14.89 | 46.32 ** | 45.75 ** | 2.51 *** | 1.97 ** | 0.5981 | 0.7326 | 0.4871 | 0.5387 | 40.82 ** | 40.93 *** |
| LSD5% | 2.59 | 1.08 | 32.77 | 15.15 | 3.72 | 1.54 | 3.49 | 3.78 | 4.19 | 3.46 | 0.16 | 0.14 | 0.45 | 0.22 | 0.19 | 0.31 | 1.07 | 1.00 | 0.09 | 0.13 | 0.2308 | 0.4637 | 0.1675 | 0.6190 | 1.60 | 0.52 |
| LSD1% | 3.84 | 1.60 | 48.50 | 22.41 | 5.50 | 2.28 | 5.17 | 5.60 | 6.21 | 5.13 | 0.24 | 0.20 | 0.67 | 0.33 | 0.28 | 0.46 | 1.59 | 1.48 | 0.14 | 0.20 | 0.3415 | 0.6863 | 0.2479 | 0.9161 | 2.37 | 0.77 |
| LSD0.1% | 5.94 | 2.49 | 74.96 | 34.64 | 8.50 | 3.52 | 7.99 | 8.66 | 9.59 | 7.92 | 0.38 | 0.31 | 1.04 | 0.51 | 0.44 | 0.72 | 2.46 | 2.28 | 0.22 | 0.31 | 0.5278 | 1.0606 | 0.3831 | 1.4158 | 3.67 | 1.19 |
| SL 27 | SW 28 | HL 29 | P 30 | G 31 | Z 32 | WW65 33 | WW91 34 | DW65 35 | DW91 36 | RWC 37 | ||||||||||||||||
| W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | W | NW | |||||
| NIL1−− | 7.05 | 7.02 | 2.97 | 2.87 | 78.37 | 79.64 | 18.18 | 17.75 | 38.13 | 36.98 | 65.24 * | 63.25 | 0.1399 | 0.1408 | 0.0735 | 0.0456 | 0.0358 | 0.0297 | 0.0614 | 0.0371 | 73.20 | 68.58 | ||||
| NIL++ | 7.01 | 7.02 | 2.97 | 2.88 | 78.72 | 80.24 | 17.44 | 17.82 | 36.89 | 37.16 | 64.00 | 63.67 | 0.1567 *** | 0.1525 | 0.0685 | 0.0503 | 0.03693 | 0.0283 | 0.0590 | 0.0428 | 73.07 | 67.80 | ||||
| LSD5% | 0.0601 | 0.0344 | 0.0502 | 0.0532 | 2.08 | 1.02 | 1.08 | 0.8754 | 1.58 | 1.35 | 1.19 | 1.03 | 0.0062 | 0.0084 | 0.0102 | 0.0094 | 0.0052 | 0.0036 | 0.0107 | 0.0088 | 2.30 | 4.11 | ||||
| LSD1% | 0.0890 | 0.0509 | 0.0743 | 0.0788 | 3.08 | 1.51 | 1.60 | 1.2956 | 2.34 | 1.99 | 1.76 | 1.52 | 0.0092 | 0.0124 | 0.0151 | 0.0139 | 0.0077 | 0.0053 | 0.0159 | 0.0131 | 3.40 | 6.08 | ||||
| LSD0.1% | 0.1376 | 0.0786 | 0.1149 | 0.1218 | 4.76 | 2.33 | 2.48 | 2.0022 | 3.63 | 3.08 | 2.73 | 2.35 | 0.0142 | 0.0192 | 0.0234 | 0.0216 | 0.0120 | 0.0083 | 0.0246 | 0.0203 | 5.26 | 9.40 | ||||
| Traits | Simple Regression between the Traits (R2) | ||||
|---|---|---|---|---|---|
| GY 1 | TGW 2 | SNM 3 | SWM 4 | SW 5 | |
| Grand mean for years and treatments | |||||
| NDVI45 6 | |||||
| NDVI65 7 | |||||
| NDVI83 8 | 38.16 *** | 18.29 * | 45.78 *** | ||
| SPAD45 9 | |||||
| SPAD65 10 | 13.33 * | 16.49 * | 27.78 *** | 25.26 ** | |
| SPAD77 11 | 32.73 *** | 38.41 *** | 38.98 *** | ||
| SPAD83 12 | 44.38 *** | 21.38 *** | |||
| SPAD85 13 | 16.51 * | ||||
| FLC 14 | 15.27 * | 33.11 *** | 16.00 * | ||
| BE 15 | 33.35 *** | ||||
| TE 16 | 34.04 *** | 12.81 * | |||
| EL 17 | 37.42 *** | 34.36 *** | |||
| FTN 18 | 40.41 *** | 48.87 *** | 22.79 *** | ||
| FLA 19 | 14.65 * | ||||
| WW65 20 | 17.55 * | 18.89 * | 37.34 *** | 30.37 ** | |
| DW65 21 | 12.74 * | 31.45 *** | 39.07 *** | 15.08 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bányai, J.; Maccaferri, M.; Láng, L.; Mayer, M.; Tóth, V.; Cséplő, M.; Pál, M.; Mészáros, K.; Vida, G. Abiotic Stress Response of Near-Isogenic Spring Durum Wheat Lines under Different Sowing Densities. Int. J. Mol. Sci. 2021, 22, 2053. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042053
Bányai J, Maccaferri M, Láng L, Mayer M, Tóth V, Cséplő M, Pál M, Mészáros K, Vida G. Abiotic Stress Response of Near-Isogenic Spring Durum Wheat Lines under Different Sowing Densities. International Journal of Molecular Sciences. 2021; 22(4):2053. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042053
Chicago/Turabian StyleBányai, Judit, Marco Maccaferri, László Láng, Marianna Mayer, Viola Tóth, Mónika Cséplő, Magda Pál, Klára Mészáros, and Gyula Vida. 2021. "Abiotic Stress Response of Near-Isogenic Spring Durum Wheat Lines under Different Sowing Densities" International Journal of Molecular Sciences 22, no. 4: 2053. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042053