
Identification and Characterization of the Most Common Genetic Variant Responsible for Acephalic Spermatozoa Syndrome in Men Originating from North Africa

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Results
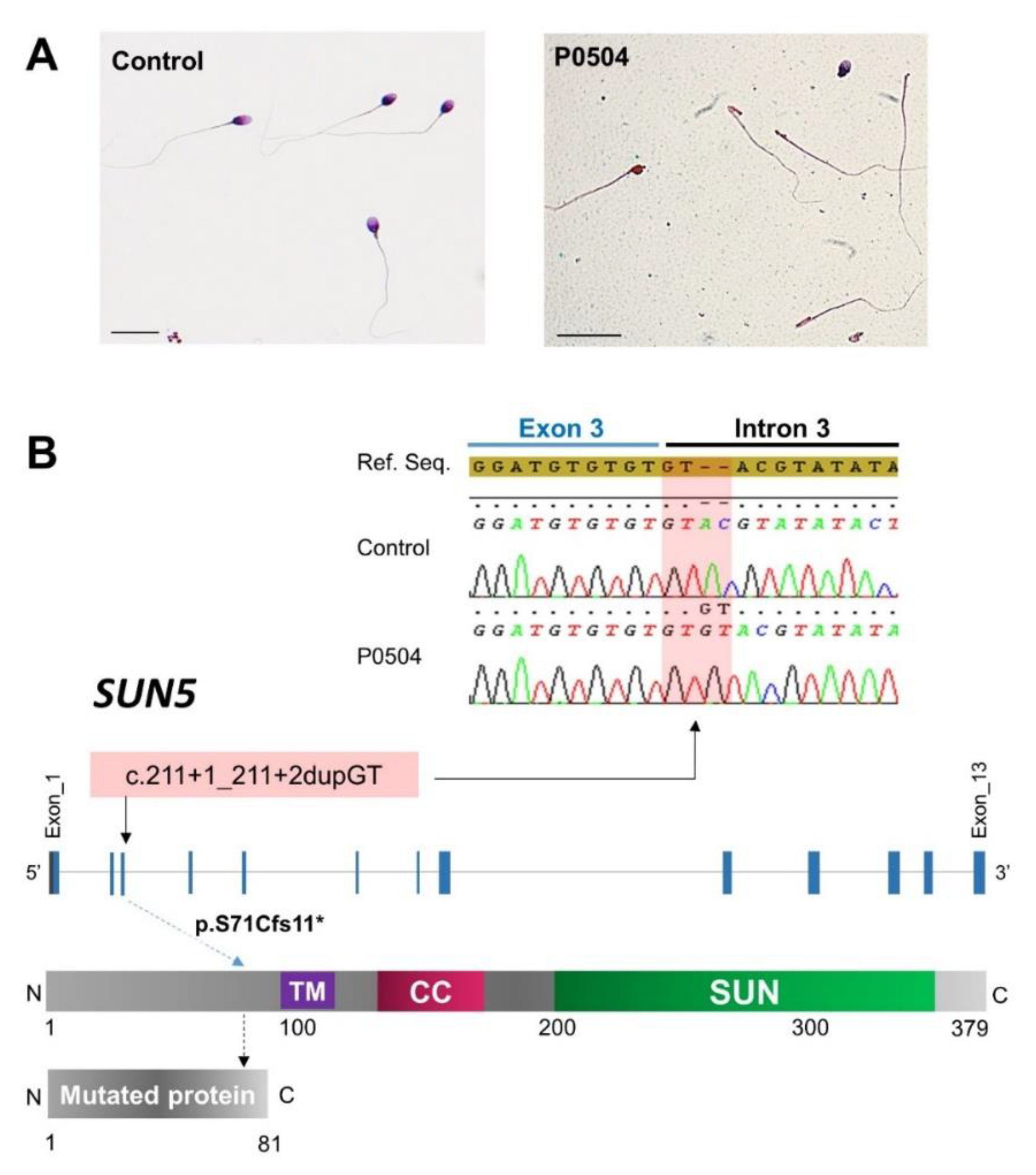
2.1. Identification of a Rare Homozygous SUN5 Variant in Seven ASS Subjects
2.2. Dating of the SUN5 c.211+1_211+2dupGT Variant
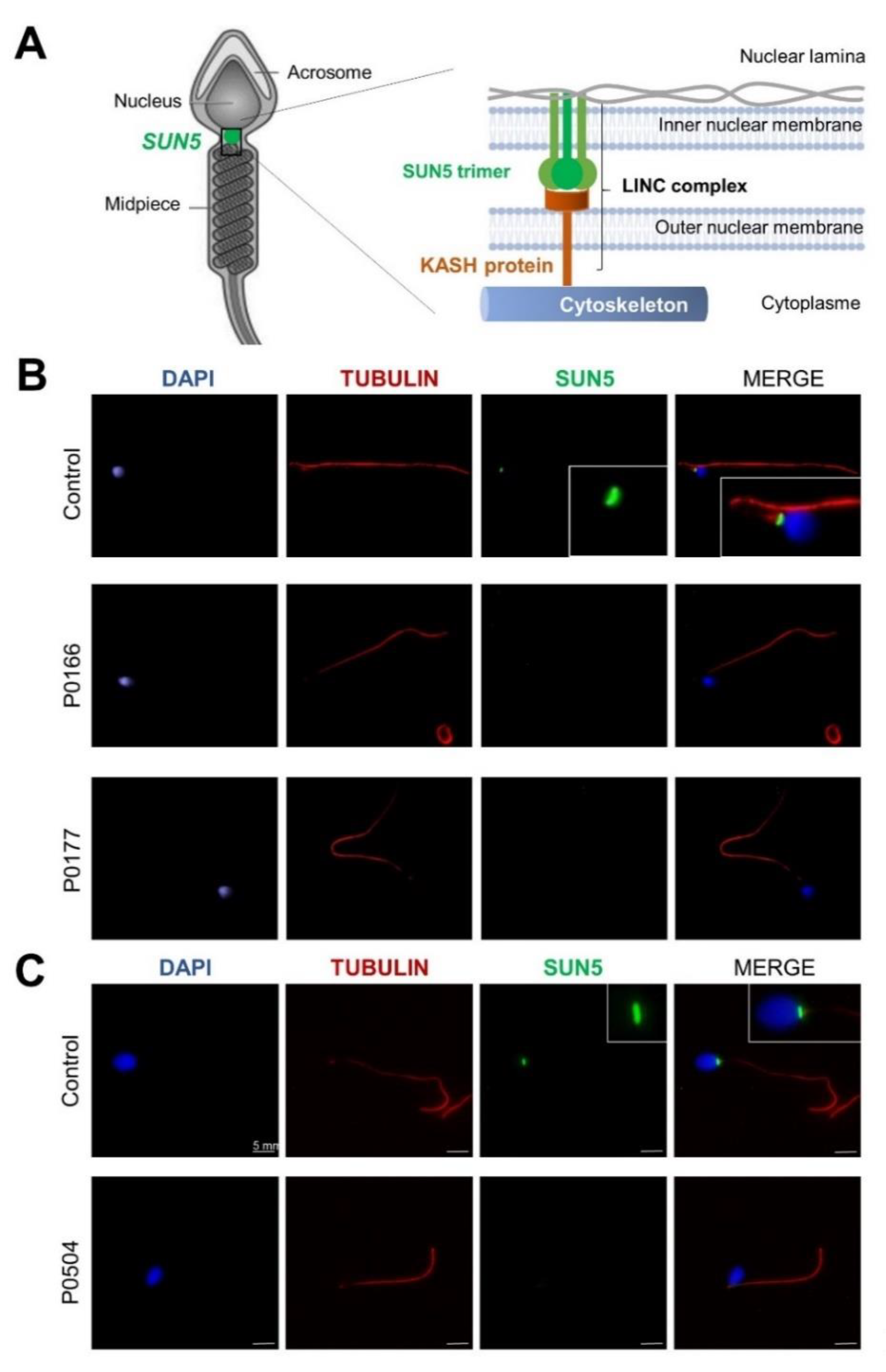
2.3. Demonstration of the Pathogenicity of the c.211+1_211+2dupGT SUN5 Variant
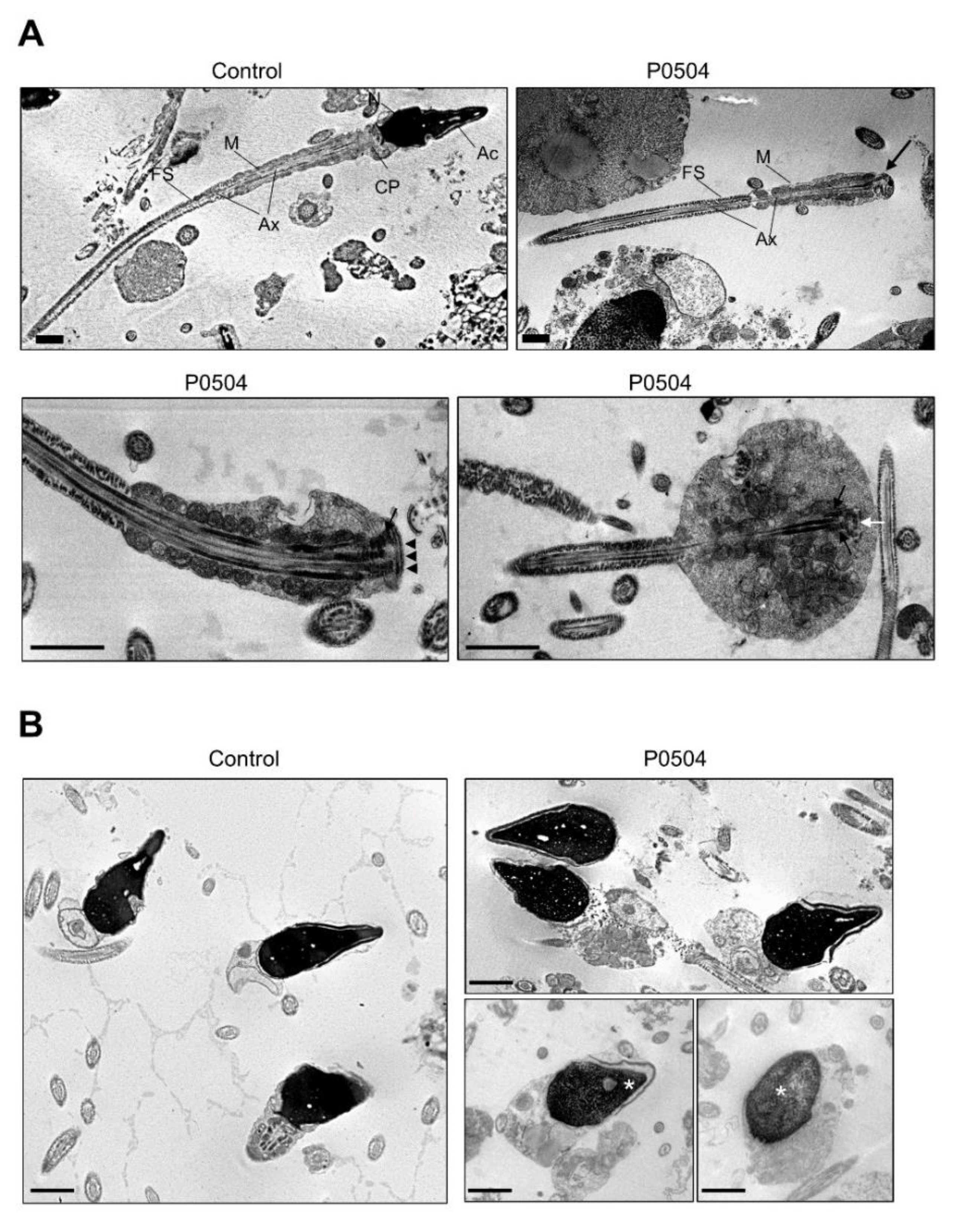
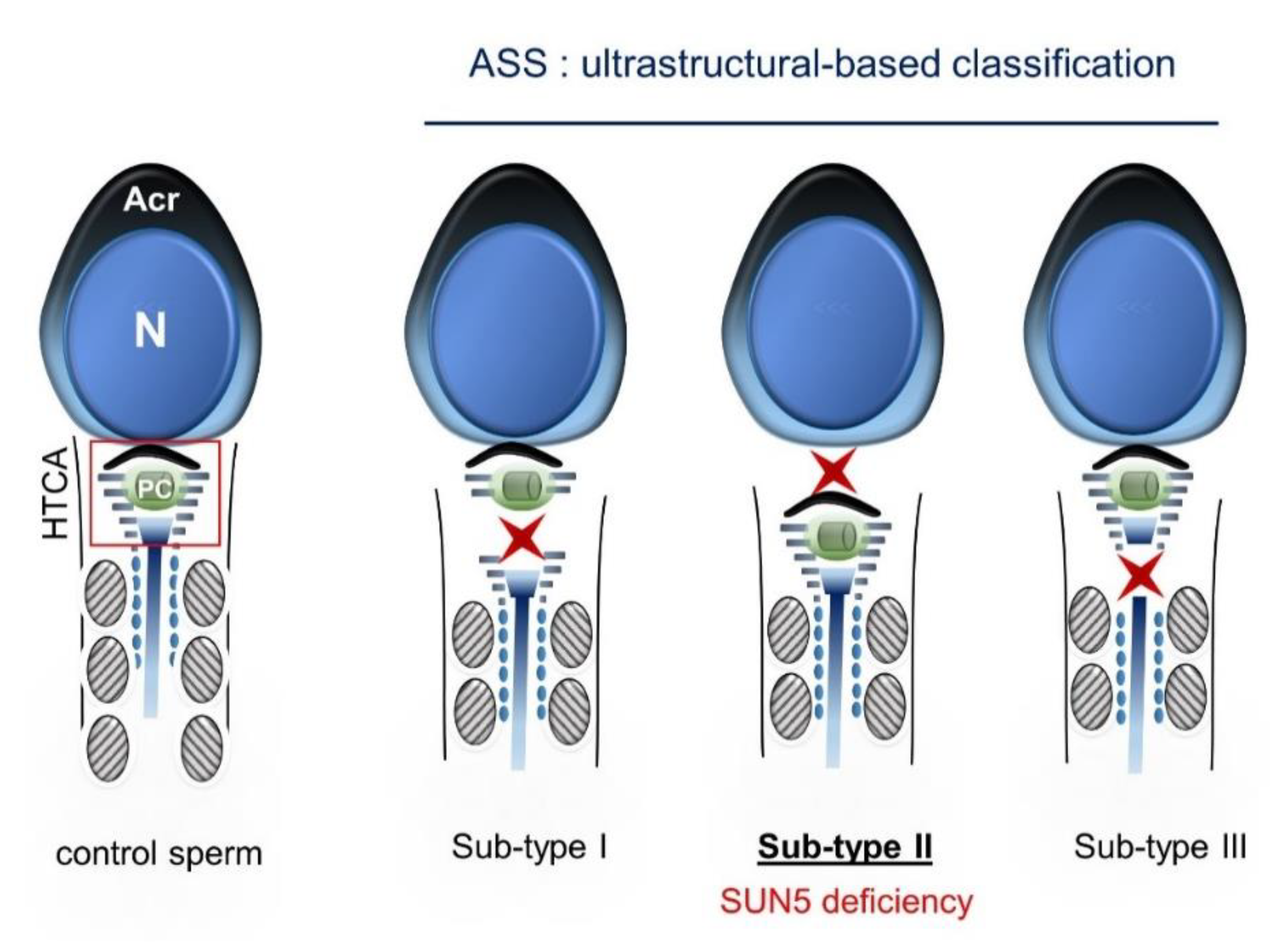
2.4. Ultrastructural Sperm Defects Associated with the Identified Homozygous SUN5 Variant
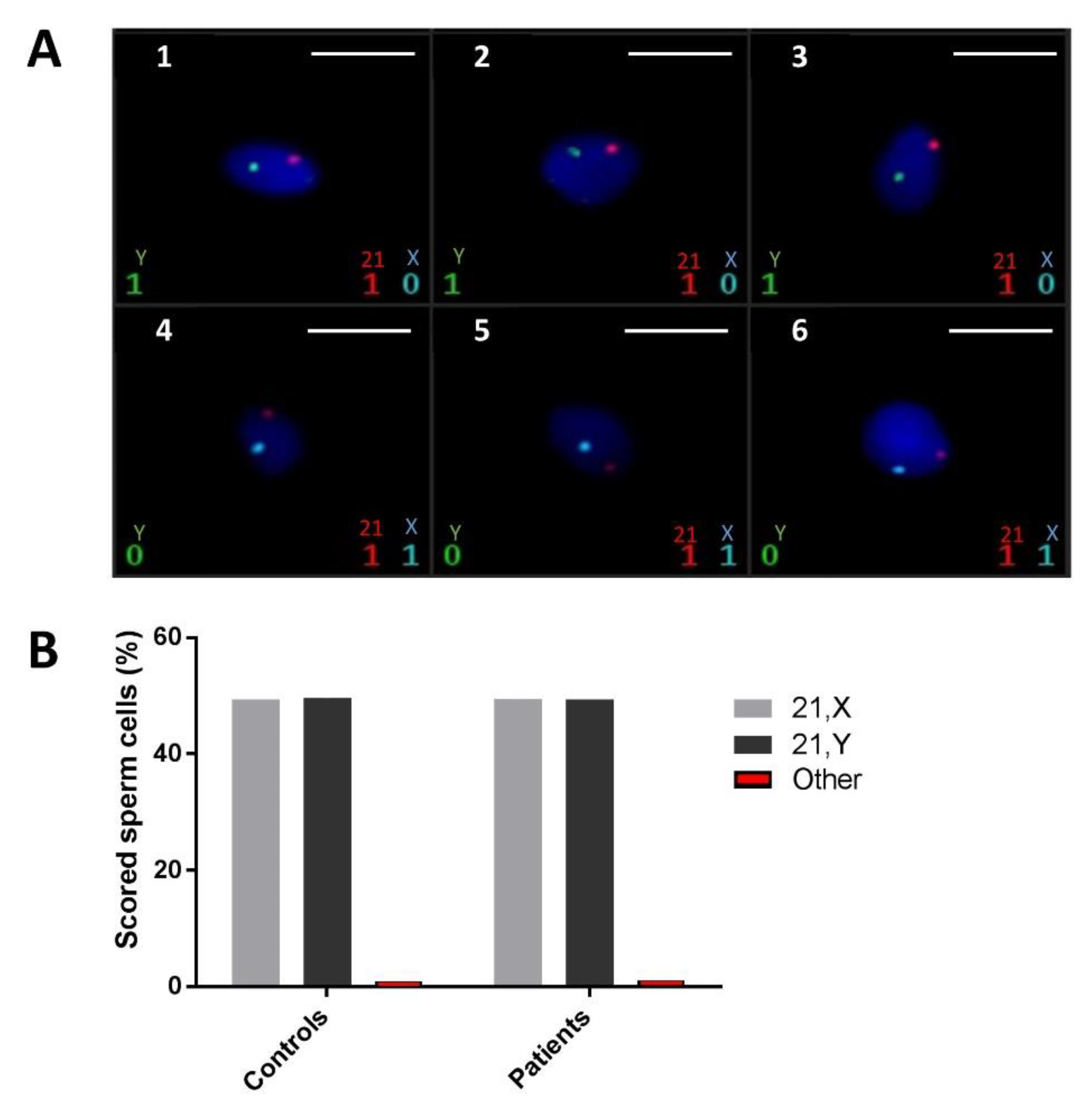
2.5. FISH Did Not Demonstrate a Higher Rate of Sperm Chromosomal Aneuploidies in ASS Men Carrying the Identified SUN5 Variant
3. Discussion
4. Material and Methods
4.1. Subject Selection and Recruitment
4.2. Semen Parameters and Sperm Morphology
4.3. Whole Exome Sequencing and Variant Filtering
4.4. Sanger Verification of the Variant
4.5. Immunofluorescence Analysis of Sperm Cells
4.6. Transmission Electron Microscopy
4.7. Sperm Fluorescent In Situ Hybridization (FISH)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mascarenhas, M.N.; Flaxman, S.R.; Boerma, T.; Vanderpoel, S.; Stevens, G.A. National, Regional, and Global Trends in Infertility Prevalence since 1990: A Systematic Analysis of 277 Health Surveys. PLoS Med. 2012, 9, e1001356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, A.; Mulgund, A.; Hamada, A.; Chyatte, M.R. A Unique View on Male Infertility around the Globe. Reprod. Biol. Endocrinol. RBE 2015, 13. [Google Scholar] [CrossRef] [Green Version]
- Ray, P.F.; Toure, A.; Metzler-Guillemain, C.; Mitchell, M.J.; Arnoult, C.; Coutton, C. Genetic Abnormalities Leading to Qualitative Defects of Sperm Morphology or Function. Clin. Genet. 2017, 91, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Touré, A.; Martinez, G.; Kherraf, Z.-E.; Cazin, C.; Beurois, J.; Arnoult, C.; Ray, P.F.; Coutton, C. The Genetic Architecture of Morphological Abnormalities of the Sperm Tail. Hum. Genet. 2021, 140, 21–42. [Google Scholar] [CrossRef]
- Chemes, H.E.; Puigdomenech, E.T.; Carizza, C.; Olmedo, S.B.; Zanchetti, F.; Hermes, R. Acephalic Spermatozoa and Abnormal Development of the Head–neck Attachment: A Human Syndrome of Genetic Origin. Hum. Reprod. 1999, 14, 1811–1818. [Google Scholar] [CrossRef] [Green Version]
- Beurois, J.; Cazin, C.; Kherraf, Z.-E.; Martinez, G.; Celse, T.; Touré, A.; Arnoult, C.; Ray, P.F.; Coutton, C. Genetics of Teratozoospermia: Back to the Head. Best Pract. Res. Clin. Endocrinol. Metab. 2020, 101473. [Google Scholar] [CrossRef]
- Elkhatib, R.A.; Paci, M.; Longepied, G.; Saias-Magnan, J.; Courbière, B.; Guichaoua, M.-R.; Lévy, N.; Metzler-Guillemain, C.; Mitchell, M.J. Homozygous Deletion of SUN5 in Three Men with Decapitated Spermatozoa. Hum. Mol. Genet. 2017, 26, 3167–3171. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.; Zhang, J.; Zhu, F.; Yang, X.; Cui, Y.; Liu, J. Patients with Acephalic Spermatozoa Syndrome Linked to SUN5 Mutations Have a Favorable Pregnancy Outcome from ICSI. Hum. Reprod. Oxf. Engl. 2018, 33, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Wang, N.; Zhang, H.; Yin, S.; Dai, H.; Lin, G.; Li, W. Novel Mutations in PMFBP1, TSGA10 and SUN5: Expanding the Spectrum of Mutations That May Cause Acephalic Spermatozoa. Clin. Genet. 2020, 97, 938–939. [Google Scholar] [CrossRef]
- Sha, Y.-W.; Xu, X.; Ji, Z.-Y.; Lin, S.-B.; Wang, X.; Qiu, P.-P.; Zhou, Y.; Mei, L.-B.; Su, Z.-Y.; Li, L.; et al. Genetic Contribution of SUN5 Mutations to Acephalic Spermatozoa in Fujian China. Gene 2018, 647, 221–225. [Google Scholar] [CrossRef]
- Shang, Y.; Yan, J.; Tang, W.; Liu, C.; Xiao, S.; Guo, Y.; Yuan, L.; Chen, L.; Jiang, H.; Guo, X.; et al. Mechanistic Insights into Acephalic Spermatozoa Syndrome-Associated Mutations in the Human SUN5 Gene. J. Biol. Chem. 2018, 293, 2395–2407. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.; Wang, F.; Yang, X.; Zhang, J.; Wu, H.; Zhang, Z.; Zhang, Z.; He, X.; Zhou, P.; Wei, Z.; et al. Biallelic SUN5 Mutations Cause Autosomal-Recessive Acephalic Spermatozoa Syndrome. Am. J. Hum. Genet. 2016, 99, 1405. [Google Scholar] [CrossRef]
- Frohnert, C.; Schweizer, S.; Hoyer-Fender, S. SPAG4L/SPAG4L-2 Are Testis-Specific SUN Domain Proteins Restricted to the Apical Nuclear Envelope of Round Spermatids Facing the Acrosome. Mol. Hum. Reprod. 2011, 17, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.-Z.; Yang, M.-G.; Huang, L.-H.; Li, C.-Q.; Xing, X.-W. SPAG4L, a Novel Nuclear Envelope Protein Involved in the Meiotic Stage of Spermatogenesis. DNA Cell Biol. 2011, 30, 875–882. [Google Scholar] [CrossRef]
- Yassine, S.; Escoffier, J.; Abi Nahed, R.; Nahed, R.A.; Pierre, V.; Karaouzene, T.; Ray, P.F.; Arnoult, C. Dynamics of Sun5 Localization during Spermatogenesis in Wild Type and Dpy19l2 Knock-out Mice Indicates That Sun5 Is Not Involved in Acrosome Attachment to the Nuclear Envelope. PLoS ONE 2015, 10, e0118698. [Google Scholar] [CrossRef]
- Shang, Y.; Zhu, F.; Wang, L.; Ouyang, Y.-C.; Dong, M.-Z.; Liu, C.; Zhao, H.; Cui, X.; Ma, D.; Zhang, Z.; et al. Essential Role for SUN5 in Anchoring Sperm Head to the Tail. eLife 2017, 6, e28199. [Google Scholar] [CrossRef]
- Sha, Y.-W.; Wang, X.; Xu, X.; Ding, L.; Liu, W.-S.; Li, P.; Su, Z.-Y.; Chen, J.; Mei, L.-B.; Zheng, L.-K.; et al. Biallelic Mutations in PMFBP1 Cause Acephalic Spermatozoa. Clin. Genet. 2019, 95, 277–286. [Google Scholar] [CrossRef]
- Sha, Y.-W.; Sha, Y.-K.; Ji, Z.-Y.; Mei, L.-B.; Ding, L.; Zhang, Q.; Qiu, P.-P.; Lin, S.-B.; Wang, X.; Li, P.; et al. TSGA10 Is a Novel Candidate Gene Associated with Acephalic Spermatozoa. Clin. Genet. 2018, 93, 776–783. [Google Scholar] [CrossRef]
- Ye, Y.; Wei, X.; Sha, Y.; Li, N.; Yan, X.; Cheng, L.; Qiao, D.; Zhou, W.; Wu, R.; Liu, Q.; et al. Loss-of-Function Mutation in TSGA10 Causes Acephalic Spermatozoa Phenotype in Human. Mol. Genet. Genomic Med. 2020, 8, e1284. [Google Scholar] [CrossRef]
- Zhu, F.; Liu, C.; Wang, F.; Yang, X.; Zhang, J.; Wu, H.; Zhang, Z.; He, X.; Zhang, Z.; Zhou, P.; et al. Mutations in PMFBP1 Cause Acephalic Spermatozoa Syndrome. Am. J. Hum. Genet. 2018, 103, 188–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen-Seaman, M.I.; Furey, T.S.; Payseur, B.A.; Lu, Y.; Roskin, K.M.; Chen, C.-F.; Thomas, M.A.; Haussler, D.; Jacob, H.J. Comparative Recombination Rates in the Rat, Mouse, and Human Genomes. Genome Res. 2004, 14, 528–538. [Google Scholar] [CrossRef] [Green Version]
- Slatkin, M.; Rannala, B. Estimating Allele Age. Annu. Rev. Genomics Hum. Genet. 2000, 1, 225–249. [Google Scholar] [CrossRef] [Green Version]
- Labuda, M.; Labuda, D.; Korab-Laskowska, M.; Cole, D.E.; Zietkiewicz, E.; Weissenbach, J.; Popowska, E.; Pronicka, E.; Root, A.W.; Glorieux, F.H. Linkage Disequilibrium Analysis in Young Populations: Pseudo-Vitamin D-Deficiency Rickets and the Founder Effect in French Canadians. Am. J. Hum. Genet. 1996, 59, 633–643. [Google Scholar]
- Risch, N.; de Leon, D.; Ozelius, L.; Kramer, P.; Almasy, L.; Singer, B.; Fahn, S.; Breakefield, X.; Bressman, S. Genetic Analysis of Idiopathic Torsion Dystonia in Ashkenazi Jews and Their Recent Descent from a Small Founder Population. Nat. Genet. 1995, 9, 152–159. [Google Scholar] [CrossRef]
- Cain, N.E.; Jahed, Z.; Schoenhofen, A.; Valdez, V.A.; Elkin, B.; Hao, H.; Harris, N.J.; Herrera, L.A.; Woolums, B.M.; Mofrad, M.R.K.; et al. Conserved SUN-KASH Interfaces Mediate LINC Complex-Dependent Nuclear Movement and Positioning. Curr. Biol. CB 2018, 28, 3086–3097.e4. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Adham, I.M.; Meinhardt, A.; Hoyer-Fender, S. Ultra-Structure of the Sperm Head-to-Tail Linkage Complex in the Absence of the Spermatid-Specific LINC Component SPAG4. Histochem. Cell Biol. 2018, 150, 49–59. [Google Scholar] [CrossRef]
- Nie, H.; Tang, Y.; Qin, W. Beyond Acephalic Spermatozoa: The Complexity of Intracytoplasmic Sperm Injection Outcomes. BioMed Res. Int. 2020, 2020. [Google Scholar] [CrossRef] [Green Version]
- Avidor-Reiss, T.; Mazur, M.; Fishman, E.L.; Sindhwani, P. The Role of Sperm Centrioles in Human Reproduction—The Known and the Unknown. Front. Cell Dev. Biol. 2019, 7, 188. [Google Scholar] [CrossRef]
- Cooper, T.G.; Noonan, E.; von Eckardstein, S.; Auger, J.; Baker, H.W.G.; Behre, H.M.; Haugen, T.B.; Kruger, T.; Wang, C.; Mbizvo, M.T.; et al. World Health Organization Reference Values for Human Semen Characteristics. Hum. Reprod. Update 2010, 16, 231–245. [Google Scholar] [CrossRef]
- Auger, J.; Jouannet, P.; Eustache, F. Another Look at Human Sperm Morphology. Hum. Reprod. Oxf. Engl. 2016, 31, 10–23. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and Accurate Long-Read Alignment with Burrows-Wheeler Transform. Bioinform. Oxf. Engl. 2010, 26, 589–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinform. Oxf. Engl. 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Aligning Sequence Reads, Clone Sequences and Assembly Contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Faust, G.G.; Hall, I.M. SAMBLASTER: Fast Duplicate Marking and Structural Variant Read Extraction. Bioinform. Oxf. Engl. 2014, 30, 2503–2505. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup The Sequence Alignment/Map Format and SAMtools. Bioinform. Oxf. Engl. 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Scheffler, K.; Halpern, A.L.; Bekritsky, M.A.; Noh, E.; Källberg, M.; Chen, X.; Kim, Y.; Beyter, D.; Krusche, P.; et al. Strelka2: Fast and Accurate Calling of Germline and Somatic Variants. Nat. Methods 2018, 15, 591–594. [Google Scholar] [CrossRef]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject Parameters | Lower Reference Limits (WHO, 2010) | P0166 | P0168 | P0177 | P0386 | P0504 | PS1 | PS2 | All SUN5 Mutated Subjects (n = 7) | All Non-Mutated Subjects (n = 2) | p Value |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Age | - | 37 | 52 | 37 | 42 | 31 | 49 | 38 | 40.86 ± 7.38 (n’ = 7) | 39 ± 2.82 (n’ = 2) | 0.61 |
| Sperm volume (mL) | 1.5 | 1.4 | 3 | 7 | 2.7 | 3.6 | 2 | 2.2 | 3.12 ± 1.85 (n’ = 7) | 5.23 ± 2.09 (n’ = 2) | 0.36 |
| Sperm concentration (million/mL) | 15 | 24 | 27 | 13.2 | 25.6 | 12.2 | 6.2 | 18 | 17.85 ± 8.16 (n’ = 7) | 9.76 ± 13.07 (n’ = 2) | 0.54 |
| Total motility 1 h (%) | 40 | 10 | 30 | 30 | 40 | 29 | 40 | 50 | 34.86 ± 12.65 (n’ = 7) | 15 ± 14.14 (n’ = 2) | 0.26 |
| Vitality (%) | 58 | 83 | 70 | 80 | 65 | 47 | 60 | 80 | 69.26 ± 13.09 (n’ = 7) | 51.25 ± 1.77 (n’ = 2) | 0.29 |
| Typical forms (%) | 4 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0.14 ± 0.38 (n’ = 7) | 11.25 ± 15.91 (n’ = 2) | 0.50 |
| Abnormal head (%) | - | 84 | 90 | 70 | 84 | 54 | 72 | 74 | 75.39 ± 12.05 (n’ = 7) | 51 ± 43.84 (n’ = 2) | 0.57 |
| Absent flagella (%) | - | 48 | 30 | 33 | 52 | 33 | 52 | 46 | 42.04 ± 9.60 (n’ = 7) | 21.75 ± 28.64 (n’ = 2) | 0.50 |
| Bent flagella (%) | - | 34 | 62 | 52 | 22 | 25 | 40 | 48 | 40.35 ± 14.67 (n’ = 7) | 31.5 ± 4.95 (n’ = 2) | 0.23 |
| Multiple anomalies index | - | 2.8 | 3.5 | 2.2 | 2.7 | 2.4 | 2.1 | - | 2.62 ± 0.51 (n’ = 6) | 2 ± 2.1 (n’ = 2) | 0.29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cazin, C.; Boumerdassi, Y.; Martinez, G.; Fourati Ben Mustapha, S.; Whitfield, M.; Coutton, C.; Thierry-Mieg, N.; Di Pizio, P.; Rives, N.; Arnoult, C.; et al. Identification and Characterization of the Most Common Genetic Variant Responsible for Acephalic Spermatozoa Syndrome in Men Originating from North Africa. Int. J. Mol. Sci. 2021, 22, 2187. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042187
Cazin C, Boumerdassi Y, Martinez G, Fourati Ben Mustapha S, Whitfield M, Coutton C, Thierry-Mieg N, Di Pizio P, Rives N, Arnoult C, et al. Identification and Characterization of the Most Common Genetic Variant Responsible for Acephalic Spermatozoa Syndrome in Men Originating from North Africa. International Journal of Molecular Sciences. 2021; 22(4):2187. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042187
Chicago/Turabian StyleCazin, Caroline, Yasmine Boumerdassi, Guillaume Martinez, Selima Fourati Ben Mustapha, Marjorie Whitfield, Charles Coutton, Nicolas Thierry-Mieg, Pierre Di Pizio, Nathalie Rives, Christophe Arnoult, and et al. 2021. "Identification and Characterization of the Most Common Genetic Variant Responsible for Acephalic Spermatozoa Syndrome in Men Originating from North Africa" International Journal of Molecular Sciences 22, no. 4: 2187. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042187