
Advances in Sensing, Response and Regulation Mechanism of Salt Tolerance in Rice


1
State Key Laboratory for Rice Biology, China National Rice Research Institute, Hangzhou 310006, China
2
Jiangsu Key Laboratory of Crop Genetics and Physiology/Key Laboratory of Plant Functional Genomics of the Ministry of Education/Jiangsu Key Laboratory of Crop Genomics and Molecular Breeding/Jiangsu Co-Innovation Center for Modern Production Technology of Grain Crops, Agricultural College of Yangzhou University, Yangzhou 225009, China
3
CAAS-IRRI Joint Laboratory for Genomics-Assisted Germplasm Enhancement, Agricultural Genomics Institute in Shenzhen, Chinese Academy of Agricultural Sciences, Shenzhen 518120, China
4
Strategic Innovation Platform, International Rice Research Institute, DAPO BOX 7777, Metro Manila 1301, Philippines
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(5), 2254; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052254
Submission received: 31 January 2021
/
Revised: 19 February 2021
/
Accepted: 20 February 2021
/
Published: 24 February 2021
(This article belongs to the Special Issue Molecular Aspects of Plant Salinity Stress and Tolerance)
Abstract
:Soil salinity is a serious menace in rice production threatening global food security. Rice responses to salt stress involve a series of biological processes, including antioxidation, osmoregulation or osmoprotection, and ion homeostasis, which are regulated by different genes. Understanding these adaptive mechanisms and the key genes involved are crucial in developing highly salt-tolerant cultivars. In this review, we discuss the molecular mechanisms of salt tolerance in rice—from sensing to transcriptional regulation of key genes—based on the current knowledge. Furthermore, we highlight the functionally validated salt-responsive genes in rice.
Keywords:
rice; salinity; sensing; signaling; transcription factors; osmoregulation; antioxidation; ion homeostasis1. Background
Soil salinity is one of the most significant abiotic stresses hampering plant growth and development, which ultimately translates to reduced crop yield. Soil salinization is exacerbated by excessive use of chemical fertilizers and soil amendments, improper drainage, and seawater ingress. It is estimated that over 6% of the world’s total land area is salt affected, of which over 12 million hectares are irrigated lands posing a serious threat to irrigated agriculture [1].
Rice, being one of the most important staple crops in the world, is crucial for food security in many Asian countries. However, it is the most salt-sensitive cereal crop, with varying responses at different growth stages [2]. It is relatively salt-tolerant at the germination, active tillering, and maturity stages, whereas it is highly sensitive at the early seedling and reproductive stages [1]. Salt sensitivity during the seedling stage often translates to reduced stand density in salt-affected paddies [3]. Meanwhile, sensitivity during the reproductive stage results in yield reduction, as attributed to spikelet sterility [4,5]. Hence, understanding how rice responds to salt stress is crucial in developing rice cultivars that could withstand salt stress.
Salinity imposes two major stresses in rice, (i) osmotic stress, and (ii) ionic stress. Osmotic stress is characterized by hyperosmotic soil solution disrupting cell turgor, similar to drought’s effect. In contrast, ionic stress is characterized by altered Na+ and K+ concentrations inside the cell, disrupting many biological processes [1]. Both osmotic and ionic stresses are perceived by membrane-bound cytosolic sensors that relay the stress signals to secondary messengers. In turn, the secondary messengers activate the protein phosphorylation cascades required for signal transduction pathways to develop salt-tolerant adaptive traits. In general, osmotic stress triggers the plant for stomatal closure, inhibiting shoot elongation. This ultimately results in reduced overall shoot growth, and, to a lesser extent, reduced root growth [6]. Meanwhile, ionic stress inhibits enzyme activity and therefore disrupts many biological processes, such as nitrogen metabolism [7,8]. Excess uptake of Na+ ions changes the NH4+ assimilation pathway, weakens the glutamate synthase pathway, and elevates the glutamate dehydrogenase pathway, impacting leaf senescence [8]. Thus, plants develop several adaptive mechanisms—namely, Na+ efflux from the roots to the rhizosphere, Na+ sequestration into the vacuole, and Na+ loading and unloading at the xylem—to avert the deleterious effect of Na+ ions in the cytosol. These mechanisms are mediated by several ion transporters coupled with H+-pumps.
In the last decades, a large number of salt-responsive genes have been functionally validated in rice (Table 1). However, the overall gene regulatory network of rice responses to salt stress remains elusive. In this review, we aim to discuss the current research progress in gene regulatory networks involved in the development of salt tolerance adaptive mechanisms in rice. We also highlight the key genes involved in salt stress sensing, signaling, transcriptional regulation, and genes encoding downstream functional molecules.
2. Salt Stress Sensing
Stress sensing is the first event in plant response to any abiotic stresses, mounting an effective adaptive strategy. Under salt stress condition, it is presumed that osmotic and ionic stresses are perceived by membrane-bound cytosolic sensors that ultimately trigger early salt-stress signaling routes (Figure 1). However, the current knowledge of how rice sense salt stress is still limited and therefore remains an open question.
2.1. Osmosensing
Sensing salt-induced osmotic stress is crucial in early signaling cascades to develop salt tolerance adaptive traits, such as growth retardation, reduction in stomatal conductance, and high abscisic acid (ABA)accumulation. However, little is known about the genetics and physiology of how rice sense hyperosmotic stress.
The transmembrane-protein-receptors, such as histidine kinases and receptor-like kinases (RLKs), function in osmotic stress perception in rice. Histidine kinases perceive osmotic fluctuations and relay the signal to response regulators via phosphotransfer, which is mediated by histidine-containing phosphotransfer protein (HpT) [101]. The first evidence of osmosensing function of histidine kinases was reported in Arabidopsis. The AtHK1, a histidine kinase encoding gene, interacts with AtHPt1 and functions as an osmosensor during both drought and salt stress [102,103]. The ortholog of AtHK1 in rice, OsHK3b, interacts with OsHpt2 and acts as a putative osmosensor [101,104]. However, functional evidence on its osmosensing role in rice is not yet reported.
The RLKs function in drought and salt stress sensing by transmitting signals to downstream signaling pathways [105]. The rice Salt Intolerance 1 (SIT1), a lectin RLK expressed mainly in root epidermal cells, acts as an upstream mediator of salt stress via elevated kinase activity [9]. Recently, Zhao et al. [106] reported that SIT1 phosphorylates B’κ at Ser402, which in turn promotes the assembly of B’κ-protein phosphatase 2A (B’κ-PP2A) holoenzyme. The B’κ-PP2A subunit positively regulates salt tolerance by deactivating the activity of SIT1 via dephosphorylation at the Thr515/516. SIT1 kinase activity in turn activates the mitogen-activated protein kinase (MAPK) 3 and MAPK 6 [9]. Thus, it could be pointed out that RLKs are important in MAPK cascade activation during osmotic stress. However, the relationship between the RLKs and MAPKs needs to be further elucidated.
Ca2+ permeable stress-gated cation channels (OSCA) also act as hyperosmotic stress sensors. The first evidence of the role of OSCA in osmosensing was reported in Arabidopsis with the characterization of OSCA1. The OSCA1 gene forms a hyperosmolality-gated Ca2+ permeable channel during osmotic stress, thereby increasing the cytosolic free Ca2+ concentration [107]. The rice genome consists of 11 OSCA genes, of which seven (OsOSCA1.1, OsOSCA1.2, OsOSCA2.1, OsOSCA2.4, OsOSCA2.5, OsOSCA3.1, and OsOSCA4.1) were upregulated during salt-induced osmotic stress and may function as an osmosensor [108]. However, the Ca2+ conducting function of the rice OSCA genes in response to hyperosmotic stress remains an open question.
2.2. Na+ Sensing
The molecular mechanism of Na+ transport in plants is well understood; however, Na+ sensing remains elusive. It has been reported that the ion transporters at the plasma membrane are potential Na+ sensors. For instance, the plasma membrane Na+/H+ antiporter SOS1 (Salt Overly Sensitive 1) is thought to be involved in Na+ sensing [109]. It was later proposed that only the long hydrophilic cytoplasmic tail of SOS1 could potentially sense Na+ ions [110]. However, no research experiments have been undertaken to support this hypothesis, and therefore it needs to be clarified. Moreover, it is unlikely that SOS1 functions as initial Na+ sensor since the SOS3/SOS2 complex regulates its activity. Na+ ions could also be sensed either extracellularly and intracellularly by membrane receptors and unknown cytosolic sensors, respectively [110]. In rice, it was suggested that the intracellular Na+ ions are sensed by an unknown cytosolic sensor based on the observed elevated levels of free cytosolic Ca2+ ions in salt stressed plants. Thus, more research is required to point out the identity of such cytosolic Na+ sensor [111].
3. Signal Transduction
During salt stress, plants transduce the early stress signals to different cellular machinery called signal transduction. In general, signal transduction starts right after stress sensing, followed by the synthesis of secondary signaling molecules, such as Ca2+ and reactive oxygen species (ROS) (Figure 1). The production of secondary signaling molecules modulates the cytosolic Ca2+ concentration that binds to different protein kinases, such as calmodulins (CaMs)/CaM-like (CML), calcium-dependent protein kinases (CDPKs), calcineurin B-like interacting protein kinases (CIPKs), and MAPKs. As these protein kinases lack enzymatic activity, they catalyze protein phosphorylation via a Ca2+-dependent manner, resulting in protein conformational change. Thus, protein phosphorylation cascades mainly depend on the cytosolic Ca2+ concentration [112,113].
3.1. CaM/CML
CaM/CML proteins are important Ca2+ transducers in plant responses to abiotic stress [114,115]. In rice, five CaM-encoding genes—namely, OsCam1-1, OsCam1-2, OsCam1-3, OsCam2, and OsCam3—were identified [10]. Among these, OsCam1-1 is highly activated during salt stress. Yuenyong et al. [116] reported that the rice plants overexpressing OsCam1-1 affected differential expression of genes involved in signaling, hormone-mediated regulation, transcription, lipid metabolism, carbohydrate metabolism, photosynthesis, glycolysis, tricarboxylic acid cycle, and glyoxylate cycle during salt stress. This further suggests that a complex network of downstream cellular processes is involved in the CaM signal transduction pathway. CaM binds with other proteins and interacts with other signaling cascades, such as plant hormone signaling, during stress conditions. For instance, it binds either with MAPK or mitogen-activated protein kinase phosphatase (MKP) to regulate the MAPK cascades [117]. Recently, six novel proteins—namely, OsLRK5a, OsDCNL2, OsWD40-139, OsGDH1, OsCIP, and OsERD2—were identified as targets of OsCML16 in responses to salt stress through yeast hybridization and bimolecular fluorescence complementation assay. These target genes are involved in plant hormone signaling processes, including auxin and ABA [118]. Interestingly, both OsCaM1 and OsCML16 could bind with OsERD2 and thus could transduce Ca2+ via both CaM and CML proteins [118]. Although the functional role of OsERD2 in response to salt stress is still unknown, it is speculated that it plays a vital role in programmed cell death during innate immunity, similar with AtERD2 [119].
3.2. CDPK
CDPKs mediate downstream components of the Ca2+ signaling cascades by directly binding Ca2+ to CaM-like domain. In rice, a total of 29 CDPK genes have been identified [120]. Four rice CDPK genes—namely, OsCPK4, OsCDPK7, OsCPK12, and OsCPK21—were functionally validated and act as positive regulators of salt tolerance (Table 1). Overexpression of rice CDPKs upregulate expression of genes involved in lipid metabolism and the active oxygen detoxification system. For instance, overexpression of OsCPK4 upregulated the genes involved in oxidative stress and redox regulation [11]. Similarly, transgenic rice plants overexpressing OsCPK12 significantly enhanced the expression of genes encoding reactive oxygen species (ROS) scavenging enzymes, such as OsAPx2 and OsAPx8 [14]. OsCDPK7 positively regulates salt tolerance by regulating salt-stress responsive gene, rab16A [12,13]. Meanwhile, OsCPK21 enhances salt tolerance via regulation of ABA- and salt stress-inducible genes, such as Rab21, OsNAC6, OsLEA3, OsP5CS, OsNHX1, and OsSOS1 [15]. Further study revealed that OsCPK21 regulates salt tolerance by phosphorylating OsGF14e/Os14-3-3 at the Tyr138 [16]. This was the first evidence of 14-3-3 protein-associated phosphorylation of CDPK in rice. Despite intensive work in studying the role of CDPKs in regulation of salt tolerance in rice, their role in different signaling cascades needs to be elucidated.
3.3. Calcineurin B-Like Protein (CBL)/CIPK
CBLs are plant-specific Ca2+ sensors that bind with CIPKs to relay perceived Ca2+ signal, thereby inducing downstream gene regulation for abiotic stress. The SOS3–SOS2 complex is the first evidence of CBL–CIPK interaction in plant responses to salt stress [121]. Homologues of SOS2 and SOS3 in rice, the OsCIPK24 and OsCBL4, have been cloned, which suggests that the SOS pathway also operates in rice responses to salt stress [122]. Further study revealed that OsCIPK24/OsSOS2, OsCBL4/OsSOS3, and OsSOS1 were highly upregulated in salt-tolerant rice cultivars when subjected to salt stress [123]. This suggests that the rice CBL4–CIPK24 complex, together with the Ca2+ signal, regulates ion homeostasis similar to Arabidopsis. Therefore, the SOS pathway is conserved in both dicots and monocots. Many other CBL and CIPK genes are involved in rice responses to salt stress based on transcriptome analysis [124,125]. However, only OsCIPK15 and OsCIPK31 have been functionally validated for their role in salt tolerance. Transgenic rice plants overexpressing OsCIPK15 showed enhanced salt tolerance with higher free proline and soluble sugar concentration [17]. Similarly, OsCIPK31 acts as a positive regulator of salt tolerance wherein the loss-of-function mutant oscipk31:Ds exhibited hypersensitive phenotype under saline condition [18].
3.4. MAPK
MAPK is considered the last component of the protein phosphorylation cascade in transducing Ca2+ ions in response to environmental stimulus. The MAPK signaling pathway activates different transcription factors (TFs) involved in the production and scavenging of ROS [126]. Three classes of MAPKs are found in plants; namely, MAPK kinase kinase (MKKK), MAPK kinase (MKK), and MAPK [127,128]. Rice has 15 MAPKs, 8 MKKs, and 75 MKKKs, of which a few are involved in salt stress response (Table 1) [129,130,131]. Overexpression and gene silencing validated the role of OsMAPK5 as a positive regulator of salt tolerance [19]. Further study showed that OsMAPK5 phosphorylates SERF1, a regulator of ROS signaling during initial response to salt stress [34]. Thus, OsMAPK5 plays an essential role in the ROS signaling pathway. In contrast, OsMAPK33 acts as a negative regulator and alters the expression of genes involved in Na+ transport [20]. OsMAPKKK63 also acts as a negative regulator of salt tolerance and interacts with OsMKK1 and OsMKK6 [23]. Both OsMKK1 and OsMKK6 are known mediators of rice responses to salt stress. Overexpression of OsMKK6 enhances salt tolerance by inducing MAPK substrate phosphorylation [22]. Similarly, OsMKK1 acts as a positive regulator with highly upregulated transcripts under saline conditions [21]. Moreover, yeast hybridization and in-vivo/vitro kinase assays revealed that OsMPK4 is the downstream target of OsMKK1. OsMPK4 is involved in the wounding signaling pathway in rice [132]. However, its functional role in salt tolerance is not well characterized.
4. Transcriptional Regulation
In the past centuries, numerous proteins were reported to play an important role in salt tolerance. Transcriptomic tools have further subdivided these proteins into two major classes, the functional and regulatory proteins. Functional proteins are those that directly function in protecting the plants from stress. These include ion transporters, antioxidant proteins, osmolytes, water channel proteins, heat shock proteins, and late embryogenesis abundant (LEA) proteins. On the contrary, regulatory proteins, such as transcription factors (TFs), are involved in regulating the complex network of signal transduction [133,134,135,136].
TFs are key proteins that bind with cis-elements in the promoter of target genes, thereby modulating the rate of gene expression in the downstream signaling cascades in response to different environmental cues. A large number of TFs have been identified in rice, with 2025 TFs in Oryza sativa spp. indica and 2384 in spp. japonica [137]. In recent years, many TFs along with their interacting proteins have been implicated in rice responses to salt stress and regulate a series of signaling pathways (Table 1). Most of these are members of APETALA2/ethylene responsive-factor (AP2/ERF), NAC (NAM, ATAF, and CUC) proteins, myeloblastosis (MYB), basic leucine-zipper (bZIP) type proteins, zinc finger (ZF) and basic helix-loop-helix (bHLH) TFs that regulate many salt stress-responsive genes either through an ABA-dependent or -independent manner (Figure 2). Thus, understanding how TFs, along with their interacting proteins, regulate a network of signaling pathways and their downstream genes is crucial in elucidating the salt tolerance mechanisms of rice.
4.1. APETALA2/Ethylene Responsive Factor (AP2/ERF) Regulation
AP2/ERF-type TFs are characterized by the presence of an AP2 DNA-binding domain of approximately 60 amino acids. In rice, at least 163 AP2/ERF TFs have been identified. This TF family is further subdivided into four subfamilies: the AP2, dehydration responsive element-binding (DREB), ERF, and related to ABI3 and VP1 (RAV) proteins [138]. Among these, DREB is widely involved in rice responses to salt stress, though a few AP2-, ERF-, and RAV-type TFs regulate salt tolerance (Table 1).
DREB binds to the dehydration-responsive element/c-repeat (DRE/CRT) cis-elements in the promoter region of stress-responsive genes. DREBs have been isolated in several crops, and their overexpression enhances tolerance to different abiotic stresses, including salinity [139]. Rice DREB1 genes enhance salinity tolerance by regulating osmoprotection, as evident in rice and Arabidopsis DREB1 overexpression plants [25,26,140,141]. For instance, OsDREB1A targets two dehydrin genes [24]. Dehydrins protect plasma membrane from damage during drought- or salt-induced osmotic stress [142]. Moreover, the level of proline and soluble sugars, which are important for osmotic adjustment, significantly increased in DREB1 overexpression plants [140,143]. DREB genes mainly work in the ABA-independent pathway; however, some also participate in the ABA-dependent pathway, as exemplified by OsDREB1F. Transcript profiling in OsDREB1F overexpression lines showed expression of ABA-dependent genes, rd29B and RAB18 [26]. DREB2-type genes also act as positive regulators of salt tolerance. Overexpression of OsDREB2A and OsDREB2B in both rice and Arabidopsis improved salt tolerance [24,27,28,29]. Another DREB gene, OsDREB6, classified as an A-6 type of DREB TF positively regulates salt tolerance. Transgenic rice plants overexpressing OsDREB6 showed high levels of proline, soluble sugars, and catalase. Conversely, the levels of these enzymes were significantly reduced in RNAi plants [33]. This suggests that DREB genes mainly enhance salt tolerance by regulating genes responsible for osmoprotection and antioxidation. Similar to DREB, other TFs in the AP2/ERF family enhance salt tolerance by regulating several downstream genes involved in osmotic stress and antioxidant defense system. For instance, SERF1 gene regulates ROS-dependent signaling as an initial response to salt stress [34]. Recently, Wang et al. [32] demonstrated that OsSTAP1, an AP2/ERF-type TF, positively regulates salt tolerance by activating genes encoding antioxidant enzymes (OsPOD1, OsPOD72, GSTT3) and aquaporin gene (NIP2-1). Unlike most of AP2/ERF-type TFs, OsERF922 and OsAP23 act as negative regulators and downregulate the expression of defense-related genes [30,35].
4.2. NAC Regulation
NAC proteins are a plant-specific gene family that regulate both ABA-independent and ABA-dependent inducible genes [144]. Several studies have been carried out to understand the role of rice NAC genes in response to abiotic stimulus, including salinity. Most functionally characterized rice NAC proteins act as positive regulators of salt tolerance (Table 1). SNAC1, the first stress-related NAC type TF characterized in rice, enhances both drought and salt tolerance [48]. Transcriptome analysis of transgenic plants overexpressing NAC proteins showed upregulation of many stress-inducible genes. For instance, OsNAC2, OsNAC5, ONAC022, and ONAC106 target OsLEA3 [38,41,43,44]; OsNAP targets several stress-related genes, including OsPP2C06/OsABI2, OsPP2C09, OsPP2C68, and OsSalT [37]; and OsNAC2 targets genes involved in osmoprotection (OsP5CS1), antioxidation (OsCOX11), K+-efflux channel genes (OsGORK and OsSKOR), and ABA-inducible genes (OsNCED1 and OsNCED3) [42,43]. NAC TFs also regulate other stress-related TFs. For instance, OsNAP induces the expression of OsDREB1A and OsMYB2 [37]. ONAC106 binds with the promoter of OsNAC5, OsDREB2A, and OsbZIP23 TF genes [41]. Similarly, ONAC022 targets OsDREB2a and OsbZIP23 (Hong et al. 2016).
4.3. MYB Regulation
MYB proteins are one of the richest TF families in plants, representing at least 155 genes in rice. It is considered as an active player in plant development, secondary metabolism, cell differentiation, organ morphogenesis, and response to both biotic and abiotic stresses [145,146]. These TFs mainly participate in the ABA-dependent pathway, upregulating a number of stress-responsive genes. For example, expression of OsMPS, an R2R3 type MYB TF, is significantly induced by ABA and regulates several expansin and glucanase genes [54]. Transcriptome analysis of transgenic rice plants overexpressing OsMYB48-1 upregulates ABA biosynthesis genes (OsNCED4 and OsNCED5), early signaling genes (OsPP2C68 and OSRK1), and late responsive genes (RAB21, OsLEA3, RAB16C, and RAB16D) [53]. Similarly, OsMYB2 targets OsLEA3 and OsRab16A [51]. MYB TFs also regulate the expression of some transporter genes. For example, OsMYBc binds with the AAANATNY motif in the promoter of OsHKT1;1, thereby upregulating its expression [56]. Other rice MYB TFs involved in the regulation of salt tolerance are presented in Table 1.
4.4. bZIP Regulation
bZIP TFs are composed of a highly conserved basic region and a leucine zipper domain of about 60 to 80 amino acids in length. Several rice bZIP TFs are involved in transcriptional activation of several stress-responsive genes, most of which participate in the ABA-dependent pathway (Table 1). Overexpression of OsbZIP71 upregulates several genes that encode ion antiporters (OsCLC-1, OsNHX1, OsHKT6 and OsVHA-B) and ROS scavenging (OsCAT). Interestingly, OsbZIP71 directly binds to the promoter of OsNHX1, an Na+/H+ antiporter gene involved in vacuolar compartmentation of Na+ ions [60]. OsbZIP23 acts as a key player in salt tolerance by upregulating osmotic stress-inducible genes, such as dehydrins and LEA proteins [59]. OsHBP1b, also categorized under the bZIP TF family, could enhance salt tolerance by activating the genes involved in antioxidant defense system [61]. It is worth noting that OsHBP1b is localized within the Saltol quantitative trait locus (QTL) region, hence an important salt tolerance gene. Moreover, comparative transcript profiling showed that OsHBP1b is highly expressed in popular salt-tolerant rice cultivar Pokkali [147]. Meanwhile, OsABI5 acts as a negative regulator changing the expression of many salt stress-responsive genes. OsABI5 significantly downregulates the expression of OsHKT1;5/SKC1 and upregulates SalT gene [58]. Transcriptomic analysis showed that many other bZIP TFs play an important role in rice responses to salt stress. However, their regulatory roles have not been functionally studied. Taken together, bZIP TFs mainly regulate salt tolerance via the active oxygen detoxification and ion homeostasis pathways.
4.5. ZF Regulation
ZF proteins are comprised of conserved motifs with cystine (Cys) and histidine (His) residues. These motifs are classified according to the number and order of Cys and His. [148]. Several studies have shown their function in transcriptional activation of several biological processes involved in plant responses to environmental stimulus. Under salt stress conditions, ZF TFs regulate the expression of genes associated with ROS scavenging via ABA-independent and ABA-dependent pathways to reduce oxidative damage. The ZFP179, ZFP182, and ZFP252 act as positive regulators of salt tolerance. These ZF TFs transcriptionally activate the OsDREB1A, OsLEA3, OsPC5CS, and OsProT genes that are involved in the synthesis of osmolytes, such as proline and soluble sugars [64,65,67]. Conversely, drought and salt tolerance (DST) and ZFP185 act as negative regulators and downregulate several ABA-inducible genes, such as Prx24 [62,66]. Meanwhile, OsLOL5, an LSD1-like-type ZF is involved in transcriptional activation of OsAPX2, OsCAT, and OsCu/Zn-SOD [68]. Thus, ZF TFs play an essential role in the ROS signaling pathway.
4.6. bHLH Regulation
bHLH TFs widely exist in eukaryotic organisms and contain a conserved basic region and a helix-loop-helix (HLH) domain [149]. These TFs play an essential role in several abiotic stress tolerance, wherein several bHLH TF genes have been functionally validated. Concerning salt tolerance, only a few were functionally validated. Three previously reported bHLH TFs enhance salt tolerance in rice by activating ion transporters genes. For instance, OsbHLH035 enhances salt tolerance by activating Na+ transporter genes, OsHKT1;3 and OsHKT1;5/SKC1, which are involved in Na+ loading and unloading [70]. OrbHLH001 enhances Na+ efflux and K+ influx under salt stress by activating OsAKT1 [69]. Meanwhile, OsbHLH062 acts as transcriptional activator of OsHAK21 in response to salt stress [150]. The bHLH TFs therefore regulate salt tolerance via the ion homeostasis pathway. Moreover, these TFs activate gene expression through their interaction with the specific E-box motif in the promoter of the target gene [69,141,151].
4.7. Other TFs Involved in Salt Tolerance
In addition to the TFs previously discussed, many other TF families play an essential role in reprogramming transcriptome during salt stress. The homeodomain-leucine zipper (HD-Zip) TF family is also important for salt tolerance, such as Oshox22 and OsTF1L. Oshox22 acts a negative regulator of salt tolerance and is upstream to OsbZIP23 [71]. OsTF1L positively regulates salt tolerance mainly by regulating genes involved in stomatal closure and lignin biosynthesis [72].
Apart from OsbZIP71, previously discussed, several TFs belonging to different families regulate the expression of the OsNHX1 transporter gene. The OsNIN-like4 and OsPCF2, a nodule inception (NIN) and teosinte branched 1/cycloidea/proliferating cell (TCP) proteins, respectively, act as transcriptional activators of OsNHX1. Conversely, OsCPP5 and OsNIN-like2 act as repressors [152]. OsMADS25, a MADS-box TF gene, acts as positive regulator by upregulating the expression of genes involved in the ROS detoxification system [73]. Meanwhile, the WRKY-type TF, OsWRKY45, negatively regulates salt tolerance [74].
5. Salt Tolerance Adaptive Mechanisms
Several adaptive mechanisms have been observed in plant responses to salt stress. In rice, osmoregulation, stomatal closure, and development of antioxidant enzymes are the immediate responses during salt stress. This is later followed by Na+ exclusion and sequestration upon uptake of toxic Na+ ions. The tissue specific localization of genes that regulate salt tolerance adaptive traits in rice is presented in Figure 3.
5.1. Osmoprotection and Osmoregulation
Cell dehydration due to low osmotic potential of soil water is the immediate effect of salt stress. Under such a situation, plants (1) synthesize compatible solutes, known as osmolytes, to maintain cell turgor and (2) activate water channel aquaporins that regulate water uptake.
5.1.1. Osmolytes
Several osmolytes, such as trehalose and glycine betaine (GB), have been proven effective in preventing cellular dehydration during salt stress [153]. Thus, exogenous application of osmolytes has been utilized to enhance salt tolerance in rice [154,155,156,157]. However, very few studies have been conducted to characterize osmolyte encoding genes for their role in salt tolerance.
The two key enzymes in trehalose biosynthesis, trehalose-6-phosphate phosphatase (TPP) and trehalose-6-phosphate synthase (TPS), are involved in rice responses to salinity. The OsTPP1, OsTPS1, and OsTPS8 positively regulate salt tolerance by increasing the accumulation of trehalose and proline in rice overexpression plants [77,78,79].
GB is also an important osmolyte under salt stress that prevents lipid peroxidation [158]. Additionally, accumulation of high GB enhances photosynthetic activity [159]. The OsBADH1, a major gene involved in converting betaine aldehyde to GB, plays an important role in salt tolerance. This gene prevents oxidative damage, protects chlorophyll degradation, and ultimately prevents leaf senescence during salt stress [75]. Moreover, RNAi-directed knockdown of OsBADH1 enhances the production of ROS, causing lipid peroxidation [76]. Thus, the gene acts as a positive regulator of salt tolerance.
5.1.2. Water Channel Aquaporins
Plant aquaporins also play a significant role in osmoregulation. Aquaporins are membrane-localized channels that are mainly involved in water transport and homeostasis [160,161]. Rice has 33 aquaporins, few of which regulate root hydraulic conductivity under saline condition [162]. Overexpression of OsPIP1;1 and OsPIP2;2, plasma membrane intrinsic proteins (PIPs) family genes in Arabidopsis, enhanced salt tolerance by maintaining water homeostasis [80]. Likewise, rice overexpressing OsPIP1;1 increased root hydraulic conductivity under salt stress [81]. Rice aquaporins might be coordinately orchestrated in maintaining water homeostasis based on their organ-specific transcript expression. Transcript of OsPIP2 genes were highly expressed in the roots; thus, it could be the predominant gene regulating water uptake in the roots (Figure 3f). Conversely, the OsPIP1 gene transcript was the highest in the leaves, suggesting its role in leaf water transport [80]. Apart from the PIP genes, several tonoplast intrinsic protein (TIP) genes also play an important role in salt-induced osmotic stress [163].
5.2. Stomatal Closure
Stomatal closure is the initial response of plants under salinity and is controlled by both ABA and ROS signaling [164]. DST mainly regulates salt tolerance via stomatal closure under salt-induced osmotic stress. Further study revealed that a leucine-rich repeat (LRR)-RLK gene, LP2, required for stomatal closure is downstream to DST [82]. Interestingly, DST interacts with DST co-activator 1 (DCA1) and regulates the expression of OsPrx24, a gene encoding H2O2 scavenger [165]. Meanwhile, OsSRO1c, expressed in the guard cells and a downstream gene target of SNAC1 TF, also regulates stomatal closure under both drought- and salt-induced osmotic stress (Figure 3a). Overexpression of OsSRO1c in rice plants showed enhanced stomatal closure and maintained H2O2 homeostasis under salt stress. Conversely, knockdown mutants showed high sensitivity to osmotic stress [83].
5.3. Antioxidation
ROS synthesis is important in different signaling and physiological processes. However, overproduction of ROS is deleterious to different cellular components, such as proteins, nucleic acids, and membrane lipids. Thus, plants synthesize ROS scavenging enzymes to maintain redox homeostasis [126,166]. In this section, we discuss genes encoding ROS scavenging enzymes that are involved in rice responses to salt stress.
5.3.1. Superoxide Dismutase (SOD)
SODs catalyze the first step in the reactive-oxygen scavenging system by dismutation of the highly toxic O2− to H2O2. Thus, it is considered the most effective intracellular antioxidant enzyme. Rice has three distinct types of SOD isoforms that are differentiated according to the metals they contain, either Cu/Zn, Mn, or Fe. The activity of these SODs is associated with specific subcellular localization: Mn-SOD is located in both mitochondria and peroxisomes; Fe-SOD is located in the chloroplasts; and Cu/Zn-SOD is located in the chloroplasts, cytosol, and peroxisome [167]. The expression of genes encoding these SOD isoforms is highly influenced by salt stress and is activated by ZF-type TFs, as discussed in Section 4.5. Mishra et al. [168] reported that the increase in SOD activity of salt-tolerant rice cultivar CSR27 exposed to salinity was directly related to the upregulation of Cu/Zn-SOD encoding genes. Similar results were reported by Rossatto et al. [169], who observed upregulation of five Cu/Zn isoforms (OsCu/Zn-SOD, OsCu/Zn-SOD2, OsCu/Zn-SOD3, OsCu/Zn-SOD4, OsCu/Zn-SODCc1) under salt stress. Moreover, the rice plants overexpressing chloroplastic OsCu/Zn-SOD showed less salt-induced oxidative damage owing to higher ROS detoxification [84]. Upregulation of OsMn-SOD was also observed in rice subjected to salt stress. Tanaka et al. [85] reported that overexpression of OsMn-SOD in the chloroplasts significantly increased SOD activity and therefore enhanced salt tolerance. Similar results were observed in other plants such as wheat and tall fescue [170,171]. Conversely, salinity downregulates the expression of OsFe-SOD, thereby reducing the total SOD activity [172]. This suggests that Cu/Zn-SOD and Mn-SOD isoforms play vital roles in ROS detoxification system during stress condition.
5.3.2. Catalase (CAT)
CATs are strong antioxidant enzymes primarily located in the peroxisome that directly catalyze the conversion of H2O2 to water and oxygen [173,174]. Thus, it is indispensable in the ROS detoxification system. Cloning and characterization of the rice CAT genes predicted three isoforms; namely, OsCatA, OsCatB, and OsCatC [175]. These genes are transcriptionally activated by bZIP- and ZF-type TFs, as described in Section 4.4 and Section 4.5. RLK is also involved in transcriptional activation of CAT genes. For instance, the salt tolerance receptor-like cytoplasmic kinase 1 (STRK1) activates OsCatC via phosphorylation at the Tyr120 [176]. Several environmental factors, such as salinity, affect expression of CAT genes. Under saline condition, elevated levels of CAT activity were observed in salt-tolerant rice cultivars [177]. Interestingly, high OsCatB and OsCatC activity was observed in salt-tolerant plants grown under salt stress [178]. A similar result was reported by Wutipraditkul et al. [179], who observed an inhibitory effect of OsCatC in response to salt stress.
5.3.3. Ascorbate Peroxidase (APX)
APXs, which exist in compartment-specific isoforms, have a higher affinity for H2O2 than CATs. Thus, they detoxify even at very low H2O2 concentrations. Rice has eight APX encoding genes: the cytosolic isoforms OsAPx1 and OsAPx2; the peroxisome isoforms OsAPx3 and OsAPx4; and the chloroplastic isoforms OsAPx5, OsAPx6, OsAPx7, and OsAPx8. The OsAPx6 isoform is also localized in the mitochondria [180]. All these APX encoding genes, except OsAPx3 and OsAPx5, were upregulated in rice under salt stress [178,181]. Overexpression of OsAPx2 showed very high APX activity, thereby enhancing salt tolerance in rice [87]. Likewise, overexpression of either OsAPx1 or OsAPx2 exhibited high tolerance to salt stress in Arabidopsis; however, OsAPx2 confers better tolerance than OsAPx1 [86]. Further study revealed that silencing both OsAPx1 and OsAPx2 genes in rice resulted in normal growth and development under salt stress. This is attributed to the upregulation of CAT and APX genes [182,183]. Thus, deficiency of APXs is compensated by other antioxidant enzymes.
5.3.4. Glutathione Reductase (GR)
GRs are flavoprotein oxidoreductases and are important components of the ascorbate (AsA)-glutathione (GSH) cycle [184]. Rice has three GR isoforms: OsGR1, located in the cytosol; and OsGR2 and OsGR3, located in both mitochondria and chloroplasts [185]. These rice GRs have been implicated for their role in different abiotic stimuli, including salinity. Salt stress enhances the expression of OsGR2 and OsGR3 via the ROS detoxification system [185,186,187]. Further study demonstrated that OsGR3, primarily expressed in the roots, positively regulates salt tolerance [88].
5.3.5. Thioredoxin (TRX) and Glutaredoxin (GRX)
TRXs and glutaredoxin (GRX) are key players in redox regulation, therefore considered as redox-sensing compounds. TRX are reduced by TRX reductase, whereas GRX utilizes glutathione as a cofactor in the ROS scavenging system [188]. The rice genome has 30 and 48 genes encoding TRX and GRX, respectively. However, only a few have been functionally validated for their role in salinity tolerance [189,190]. For instance, OsTRXh1/OsTRX23 negatively regulates salt tolerance. RNAi-directed knockdown of this gene resulted in salt sensitivity, possibly due to its inhibitory activity on stress-activated MAPKs [89,191]. OsTRXh1/OsTRX23 also inhibits the kinase activity of OsMPK3 and OsMPK6 [192]. Meanwhile, OsGRX8 and OsGRX20 positively regulate salt tolerance by restraining the accumulation of O2- radicals [90,91].
5.4. Na+ Exclusion and Sequestration
Na+ ions are the major toxic element taken up by the plant during salt stress. Maintaining low levels of toxic Na+ ions in the cytosol, either through Na+ exclusion or sequestration, is the most effective strategy to avert the deleterious effects of salinity. Glycophytes, such as rice, exclude Na+ from the shoot either by (i) Na+ efflux from roots to the rhizosphere, (ii) Na+ loading and unloading at the xylem, or (iii) vacuolar Na+ compartmentation.
5.4.1. Na+ Efflux
The efflux of Na+ ions across the root plasma membrane into the external medium is poorly understood. Nevertheless, it is central to the Na+ exclusion mechanisms in plants [1]. To date, only SOS1, coupled with H+-ATPases, is the major Na+ efflux transporter that has been genetically characterized in plants [110,193]. The rice SOS1 ortholog (OsSOS1) is expressed in epidermal cells at the root cap and in cells around the xylem similar with Arabidopsis AtSOS1 [194]. The OsSOS1 activity, catalyzed by Na+/H+ exchange at the plasma membrane, could suppress Na+ sensitivity of yeast mutant lacking the Na+ efflux system, thus reducing the net cellular Na+ concentration. Similarly, OsSOS1 complementation in Arabidopsis mutant sos1-1 reduced growth defect in both saline and non-saline conditions [122]. Further study demonstrated that rice sos1 loss-of-function mutant displayed very high root Na+ uptake and impaired Na+ loading into the xylem [95]. Thus, OsSOS1 plays a critical role in Na+ efflux from root epidermal cells to the rhizosphere.
5.4.2. Na+ Loading and Unloading
Na+ loading and unloading at the xylem is regulated by high-affinity K+ transporters (HKTs). HKTs are among the most well characterized Na+ and/or K+ plant transporters identified in several plants and play a central role in salt tolerance [195,196]. Two HKTs are highlighted in a proposed two-staged Na+ exclusion mechanism, whereby the (i) OsHKT1;5/SKC1 mediates root-to-shoot Na+ transfer and (ii) OsHKT1;4 mediates leaf sheath-to-blade Na+ transfer. The Na+ ions entering the root xylem via nonselective cation channel (NSCC) are shuttled back to the parenchyma via OsHKT1;5/SKC1 (Figure 3d). Meanwhile, OsHKT1;4 not only functions in Na+ unloading to the leaf sheath, but also to the stem during the reproductive stage [197]. Further study revealed that OsHKT1;4 is involved in leaf Na+ exclusion via Na+ unloading at the xylem (Figure 3c). The mutant line overexpressing OsHKT1;4 showed salt sensitivity owing to very high root Na+ uptake [93]. Thus, a coordinated balance in root and shoot Na+ exclusion is essential to achieve salt tolerance. Another HKT1 gene, OsHKT1;1, transcriptionally activated by OsMYBc as previously discussed, is also reported to regulate Na+ exclusion, possibly through both Na+ unloading from the xylem and Na+ loading into the phloem (Figure 3c). The Na+ loaded into the phloem is hypothesized to be recirculated from shoots to roots or from young leaves to old leaves, thereby reducing salt injury in newly emerging leaf [56]. Moreover, it was demonstrated that OsHKT1;1 is a positive regulator of salt tolerance that mediates Na+ exclusion from the shoot [92]. Recent studies have shown that there are eight and four transcript variations of HKT1 genes with different lengths in O. sativa spp. indica and spp. japonica, respectively. These eight transcript variations in O. sativa spp. indica show different expression levels and transport activities under salt treatment, which suggests the existence of different transport mechanisms [198].
5.4.3. Vacuolar Na+ Sequestration
Few rice cultivars with high Na+ concentrations in the leaves were found to perform well under saline condition. This is mainly due to the active compartmentation of Na+ ions into the vacuole, also known as tissue tolerance, mediated by the tonoplast localized Na+/H+ antiporters (NHX) and energized by a proton motive force (Figure 3b) [193]. This mechanism allows the plant to use Na+ ions in maintaining cell turgor, and hence continuous plant growth under salt [199,200]. Additionally, vacuolar Na+ sequestration maintains cytosolic alkalinity and vacuolar acidity. Maintaining low vacuolar pH is essential since acidity allows the vacuole to isolate and break down misfolded proteins [201]. This phenomenon was only observed in salt-tolerant rice cultivars, such as Pokkali [111].
Four vacuolar NHX genes—namely, OsNHX1, OsNHX2, OsNHX3, and OsNHX5— were identified in rice mediating cytosolic Na+ sequestration into the vacuole [202]. Further study revealed that overexpression of OsNHX1 enhanced tissue tolerance and is regulated by OsbZIP71 TF [60,96,203]. Very high transcripts of these NHX genes in either flag leaf or panicle has also been observed [202]. This suggests their potential role in enhancing salt tolerance at the reproductive stage.
Functional characterization of vacuolar-type H+-pyrophophatase (H+-PPase) also showed enhanced salt tolerance. H+-PPase is the main driving force for Na+ transport from the cytoplasm to the vacuole (Figure 3b). Overexpression of H+-PPase encoding genes in different plants significantly enhanced salt tolerance [204,205,206]. In rice, overexpression of OsVP1, a H+-PPase encoding gene, resulted in less serious Na+ toxicity under salt stress. Moreover, double overexpression of OsNHX1 and OsVP1 conferred better salt tolerance [96]. This is possibly due to the higher electrochemical gradient brought by OsVP1 overexpression, thereby promoting higher activity of OsNHX1 (Figure 3b). Interestingly, a similar result has been found in simultaneous expression of SsNHX1 from Suaeda salsa and AVP1 from Arabidopsis in rice [206].
5.5. Suberin Deposition
Suberin deposition is essential in blocking apoplastic leakage of Na+ ions into the stele, resulting in low concentration of Na+ ions that can be transported into the shoot (Figure 3e). In rice, a few studies have reported the role of suberin in salt tolerance. Enhancing suberin in the form of silicon has significantly reduced the root-to-shoot Na+ uptake by preventing apoplastic Na+ transport across the root [207]. Interestingly, the popular salt-tolerant rice, Pokkali, showed higher suberin deposition compared with the salt-sensitive cultivar IR20 [208]. However, the gene regulatory network involved in suberin deposition and salt tolerance in rice is not well understood. The OsTPS8, involved in trehalose biosynthesis, was also reported to enhance salt tolerance, mainly by enhancing suberin deposition [79].
5.6. K+ Uptake
Cytosolic K+ concentration has emerged as an important aspect of a plant’s adaptability to salt stress, wherein high K+ concentration directly relates to salt tolerance. Four high-affinity K+ transporter (HAK) genes—namely, OsHAK1, OsHAK5, OsHAK16, and OsHAK21—play crucial roles in K+ homeostasis under stress conditions [97,98,99,100]. Interestingly, differences in spatial expression were observed among these HAK genes. β-glucoronidase (GUS) staining assay showed that OsHAK1, OsHAK5, and OsHAK16 were mainly expressed in the root epidermis [97,98,100]. Conversely, OsHAK21 was mainly expressed in the root xylem parenchyma [99]. Thus, OsHAK21 is likely the predominant gene mediating K+ influx in the xylem (Figure 3d).
6. Conclusions and Perspectives
Soil salinity, apart from drought and flooding, is a serious menace afflicting global rice production. Being the staple crop of half of the world’s population, developing salt-tolerant rice varieties is crucial, requiring a better overview on molecular and physiological responses to salt stress. Rice responds to salinity through different biological processes, starting with salt stress sensing. Sensing is mediated by different sensors. The sensors relay stress signals to secondary messengers that activate protein phosphorylation cascades and finally the transcriptional regulation of stress-responsive genes via abscisic acid (ABA)-independent/ABA-dependent pathways. Rice response to salt stress also involves several signaling components, transcription factors, and functional genes that directly mediate osmoregulation, antioxidation, and ion homeostasis. Despite the characterization of these genes, understanding the molecular mechanism of rice responses to salt stress remains a great challenge.
Over the last few decades, remarkable progress in understanding the genomics-physiology of salinity tolerance in plants has taken place. Several genes have been identified to confer salt tolerance in rice; however, most were achieved through a reverse genetics approach. Thus, a large number of genes need to be identified via forward genetics. The current understanding of the molecular responses of rice to salt stress from sensing and signaling up to the development of adaptive tolerance mechanisms is still obscure and requires further research. In particular, identification of upstream pathways and the molecular mechanisms involved in salt stress sensing is crucial to clearly disentangle the osmotic and Na+ stress responses in rice. To date, only the role of ABA signaling in rice responses to salt stress is widely studied. The crosstalk between signaling pathways and of other hormones, including auxin, gibberellic acid, jasmonic acid, and ethylene, is still not clear and needs further investigation. Studying the epigenetic regulations of salt tolerance in rice is another important field to dissect. Epigenetic mechanisms control the expression of stress-responsive genes in response to internal and environmental cues. Thus, epigenomic variations may provide a useful resource of DNA methylomes that can be used to better understand the complex salt tolerance mechanisms in rice.
Author Contributions
Y.L. and K.S.P. proposed the concept; K.S.P. drafted the manuscript; L.M. and G.Y. participated in the data collection; Y.L. and L.G. revised and finalized the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (Grant No. 31801337), Agricultural Variety Improvement Project of Shandong Province (Grant No. 2019LZGC003), and Dapeng District Industry Development Special Funds (Grant No. KY20180217).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare that they have no competing interests.
Abbreviations
| AP2/ERF | APETALA2/ethylene responsive factor |
| APX AT | Ascorbate peroxidase Adenosine-thymine |
| bHLH | Basic-helix-loop-helix |
| Bzip CAMK | Basic leucine-zipper Calcium/calmodulin-dependent protein kinase |
| CAT | Catalase |
| CBL CDA | Calcineurin B-like protein Cytidine deaminase |
| CDPK | Calcium-dependent protein kinase |
| CIPK | CBL-interacting protein kinase |
| CaM | Calmodulin |
| CML | Calmodulin-like protein |
| CPP | Cysteine-rich poly comb-like protein |
| DRE/CRT | Dehydration-responsive element/c-repeat |
| DST | Drought and salt tolerance |
| DCA1 | DST co-activator 1 |
| GB | Glycine betaine |
| GR GUS | Glutathione reductase β-glucoronidase |
| HAK | High-affinity potassium transporter |
| HD-Zip | Homeodomain-leucine zipper |
| HKT | High-affinity K+ transporter |
| HpT | Histidine-containing phosphotransfer protein |
| LEA | Late embryogenesis abundant |
| LRR-RLK | Leucine-rich repeat-receptor-like kinase |
| MAPK | Mitogen-activated protein kinase |
| MKK | MAPK kinase |
| MKKK | MKK kinase |
| MKP | Mitogen-activated protein kinase phosphatase |
| MYB | Myeloblastosis |
| NAC | NAM, ATAF and CUC |
| NAM | No apical meristem |
| NHX | Na+/H+ antiporter |
| NIN | Nodule inception |
| OSCA | Ca2+ permeable stress-gated cation channels |
| PA | Phosphatidic acid |
| PIP QTL | Plasma membrane intrinsic protein Quantitative trait locus |
| RAV | Related to ABI3 and VP1 |
| RLK | Receptor-like kinase |
| ROS | Reactive oxygen species |
| SIT | Salt intolerance |
| SOD | Superoxide dismutase |
| SOS | Salt overly sensitive |
| TCP | Teosinte branched 1/cycloidea/proliferating cell |
| TF | Transcription factor |
| TIP | Tonoplast intrinsic |
| TPS | Trehalose-6-phosphate phosphatase |
| TPP | Trehalose-6-phosphate synthase |
| TRX | Thioredoxin |
| VP | Vacuolar-type H+-pyrophosphatase |
| ZF | Zinc finger |
References
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Zeng, L.; Shannon, M.C.; Grieve, C.M. Evaluation of salt tolerance in rice genotypes by multiple agronomic parameters. Euphytica 2002, 127, 235–245. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. Effects of salt stress on growth mineral nutrition and proline accumulation in relation to osmotic adjustment in rice (Oryza sativa L.) cultivars differing in salinity resistance. Plant Growth Regul. 1996, 19, 207–218. [Google Scholar] [CrossRef]
- Cui, H.; Takeoka, Y.; Wada, T. Effect of sodium chloride on the panicle and spikelet morphogenesis in rice. Jpn. J. Crop. Sci. 1995, 64, 593–600. [Google Scholar] [CrossRef] [Green Version]
- Khatun, S.; Flowers, T.J. Effects of salinity on seed set in rice. Plant Cell. Environ. 1995, 18, 61–67. [Google Scholar] [CrossRef]
- Munns, R. Genes and salt tolerance: Bringing them together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef]
- Tester, M.; Davenport, R.J. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, M.; Guo, R.; Shi, D.; Liu, B.; Lin, X.; Yang, C. Effects of salt stress on ion balance and nitrogen metabolism of old and young leaves in rice (Oryza sativa L.). BMC Plant Biol. 2012, 12, 194. [Google Scholar] [CrossRef] [Green Version]
- Li, C.H.; Wang, G.; Zhao, J.L.; Zhang, L.Q.; Ai, L.F.; Han, Y.F.; Sun, D.Y.; Zhang, S.W.; Sun, Y. The receptor-like kinase SIT1 mediates salt sensitivity by activating MAPK3/6 and regulating ethylene homeostasis in rice. Plant Cell 2014, 26, 2538–2553. [Google Scholar] [CrossRef] [Green Version]
- Boonburapong, B.; Buaboocha, T. Genome-wide identification and analyses of the rice calmodulin and related potential calcium sensor proteins. BMC Plant Biol. 2007, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Campo, S.; Baldrich, P.; Messeguer, J.; Lalanne, E.; Coca, M.; San Segundo, B. Overexpression of a calcium-dependent protein kinase confers salt and drought tolerance in rice by preventing membrane lipid peroxidation. Plant Physiol. 2014, 165, 688–704. [Google Scholar] [CrossRef] [Green Version]
- Saijo, Y.; Hata, S.; Kyozuka, J.; Shimamoto, S.; Izui, K. Over-expression of a single Ca2+-dependent protein kinase confers both cold and salt/drought tolerance on rice plants. Plant J. 2000, 23, 319–327. [Google Scholar] [CrossRef]
- Saijo, Y.; Kinoshita, N.; Ishiyama, K.; Hata, S.; Kyozuka, J.; Hayakawa, T.; Nakamura, T.; Shimamoto, K.; Yamaya, T.; Izui, K. A Ca2+-dependent protein kinase that endows rice plants with cold- and salt-stress tolerance functions in vascular bundles. Plant Cell. Physiol. 2001, 42, 1228–1233. [Google Scholar] [CrossRef] [Green Version]
- Asano, T.; Hayashi, N.; Kobayashi, M.; Aoki, N.; Miyao, A.; Mitsuhara, I.; Ichikawa, H.; Komatsu, S.; Hirochika HKikuchi, S.; Ohsugi, R. A rice calcium-dependent protein kinase OsCPK12 oppositely modulates salt-stress tolerance and blast disease resistance. Plant J. Cell. Mol. Biol. 2011, 69, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Hakata, M.; Nakamura, H.; Aoki, N.; Ichikawa, H.; Komatsu, S.; Hirochika, H.; Kikuchi, S.; Ohsugi, R. Functional characterisation of OsCPK21 a calcium-dependent protein kinase that confers salt tolerance in rice. Plant Mol. Biol. 2011, 75, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.X.; Zhou, X.J.; Chang, S.; Chu, Z.L.; Wang, H.M.; Han, S.C.; Wang, Y.D. Calcium-dependent protein kinase 21 phosphorylates 14-3-3 proteins in response to ABA signaling and salt stress in rice. Biochem. Biophy. Res. Commun. 2017, 4, 1450–1456. [Google Scholar] [CrossRef]
- Xiang, Y.; Huang, Y.M.; Xiong, L.Z. Characterization of stress-responsive CIPK genes in rice for stress tolerance improvement. Plant Physiol. 2007, 144, 1416–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piao, H.L.; Xuan, Y.H.; Park, S.H.; Je, B.I.I.; Park, S.J.; Park, S.H.; Kim, C.H.; Huang, J.; Wang, G.K.; Kim, M.J.; et al. OsCIPK31 a CBL-interacting protein kinase is involved in germination and seedling growth under abiotic stress conditions in rice plants. Mol. Cells 2010, 30, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.Z.; Yang, Y.N. Disease resistance and abiotic stress tolerance in rice are inversely modulated by an abscisic acid–inducible mitogen-activated protein kinase. Plant Cell 2003, 15, 745–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.K.; Kim, B.G.; Kwon, T.R.; Jeong, M.J.; Park, S.R.; Lee, J.W.; Byun, M.O.; Kwon, H.B.; Matthews, B.F.; Hong, C.B.; et al. Overexpression of the mitogen-activated protein kinase gene OsMAPK33 enhances sensitivity to salt stress in rice (Oryza sativa L.). J. Biosci. 2011, 36, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.Z.; Jing, W.; Zhang, W.H. The mitogen-activated protein kinase cascade MKK1–MPK4 mediates salt signaling in rice. Plant Sci. 2014, 227, 181–189. [Google Scholar] [CrossRef]
- Kumar, K.; Sinha, A.K. Overexpression of constitutively active mitogen activated protein kinase kinase 6 enhances tolerance to salt stress in rice. Rice 2013, 6, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Na, Y.J.; Choi, H.K.; Park, M.Y.; Choi, S.W.; Vo, K.T.X.; Jeon, J.S.; Kim, S.Y. OsMAPKKK63 is involved in salt stress response and seed dormancy control. Plant Signal Behav. 2019, 14, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubouzet, J.G.; Sakuma, Y.; Ito, Y.; Kasuga, M.; Dubouzet, E.G.; Miura, S.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. OsDREB genes in rice Oryza sativa L., encode transcription activators that function in drought-, high-salt- and cold-responsive gene expression. Plant J. 2003, 33, 751–763. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, C.; Jin, X.F.; Xiong, A.S.; Peng, R.H.; Hong, Y.H.; Yao, Q.H.; Chen, J.M.; Chen, J.M. Expression of a rice DREB1 gene, OsDREB1D, enhances cold and high-salt tolerance in transgenic Arabidopsis. BMB Rep. 2009, 42, 486–492. [Google Scholar] [CrossRef]
- Wang, Q.; Guan, Y.; Wu, Y.; Chen, H.; Chen, F.; Chu, C. Overexpression of a rice OsDREB1F gene increases salt drought and low temperature tolerance in both Arabidopsis and rice. Plant Mol. Biol. 2008, 67, 589–602. [Google Scholar] [CrossRef]
- Cui, M.; Zhang, W.; Zhang, Q.; Xu, Z.; Zhu, Z.; Duan, F.; Wu, R. Induced over-expression of the transcription factor OsDREB2A improves drought tolerance in rice. Plant Physiol. Biochem. 2011, 49, 1384–1391. [Google Scholar] [CrossRef]
- Mallikarjuna, G.; Mallikarjuna, K.; Reddy, M.K.; Kaul, T. Expression of OsDREB2A transcription factor confers enhanced dehydration and salt stress tolerance in rice (Oryza sativa L.). Biotechnol. Lett. 2011, 33, 1689–1697. [Google Scholar] [CrossRef]
- Matsukura, S.; Mizoi, J.; Yoshida, T.; Todaka, D.; Ito, Y.; Maruyama, K.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Comprehensive analysis of rice DREB2-type genes that encode transcription factors involved in the expression of abiotic stress-responsive genes. Mol. Genet. Genom. 2010, 283, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, J.; Jiang, H.H.; Wang, F.; Peng, R.H.; Yao, Q.H.; Xiong, A.S. A Rice OsAP23, functioning as an AP2/ERF transcription factor, reduces salt tolerance in transgenic Arabidopsis. Plant Mol. Biol. Rep. 2013, 31, 1336–1345. [Google Scholar] [CrossRef]
- Oh, S.J.; Kim, Y.S.; Kwon, C.W.; Park, H.K.; Jeong, J.S.; Kim, J.K. Overexpression of the transcription factor AP37 in rice improves grain yield under drought conditions. Plant Physiol. 2009, 150, 1368–1379. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.X.; Wang, J.; Zhao, X.Q.; Yang, S.; Huang, L.Y.; Du, F.P.; Li, Z.K.; Zhao, X.Q.; Fu, B.Y.; Wang, W. Overexpression of the transcription factor gene OsSTAP1 increases salt tolerance in rice. Rice 2020, 50, 1–12. [Google Scholar] [CrossRef]
- Ke, Y.G.; Yang, Z.J.; Yu, S.W.; Li, T.F.; Wu, J.H.; Gao, H.; Fu, Y.P.; Luo, L.J. Characterization of OsDREB6 responsive to osmotic and cold stresses in rice. J. Plant Biol. 2014, 57, 150–161. [Google Scholar] [CrossRef]
- Schmidt, R.; Mieulet, D.; Hubberten, H.M.; Obata, T.; Hoefgen, R.; Fernie, A.R.; Fisahn, J.; San Segundo, B.; Guiderdoni, E.; Schippers, J.H.M.; et al. Salt-responsive ERF1 regulates reactive oxygen species-dependent signaling during the initial response to salt stress in rice. Plant Cell 2013, 25, 2115–2131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.F.; Chen, X.J.; Liu, J.Q.; Ye, J.C.; Guo, Z.J. The rice ERF transcription factor OsERF922 negatively regulates resistance to Magnaporthe oryzae and salt tolerance. J. Exp. Bot. 2012, 63, 3899–3911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Y.B.; Li, J.; Qin, R.Y.; Xu, R.F.; Li, H.; Yang, Y.C.; Ma, H.; Li, L.; Weng, P.C.; Yang, J.B. Identification of a regulatory element responsible for salt induction of rice OsRAV2 through ex situ and in situ promoter analysis. Plant Mol. Biol. 2016, 90, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, Y.F.; Lv, B.; Li, J.; Luo, L.Q.; Lu, S.C.; Zhang, X.; Ma, H.; Ming, F. The NAC family transcription factor OsNAP confers abiotic stress response through the ABA pathway. Plant Cell. Physiol. 2016, 55, 604–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Y.B.; Zhang, H.J.; Huang, L.; Li, D.Y.; Song, F.M. Overexpression of a stress-responsive NAC transcription factor gene ONAC022 improves drought and salt tolerance in rice. Front. Plant Sci. 2016, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.N.; Chen, B.; Lu, G.J.; Han, B. Overexpression of a NAC transcription factor enhances rice drought and salt tolerance. Biochem. Biophys. Res. Commun. 2009, 379, 985–989. [Google Scholar] [CrossRef]
- Yokotani, N.; Ichikawa, T.; Kondou, Y.; Matsui, M.; Hirochika, H.; Iwabuchi, M.; Oda, K. Tolerance to various environmental stresses conferred by the salt-responsive rice gene ONAC063 in transgenic Arabidopsis. Planta 2009, 229, 1065–1075. [Google Scholar] [CrossRef] [PubMed]
- Sarukaba, Y.; Piao, W.L.; Lim, J.H.; Han, S.H.; Kim, Y.S.; An, G.; Paek, N.C. Rice ONAC106 inhibits leaf senescence and increases salt tolerance and tiller angle. Plant Cell. Physiol. 2015, 56, 2325–2339. [Google Scholar] [CrossRef]
- Mao, C.J.; Ding, J.L.; Zhang, B.; Xi, D.D.; Ming, F. OsNAC2 positively affects salt-induced cell death and binds to the OsAP37 and OsCOX11 promoters. Plant J. 2018, 94, 454–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, D.G.; Zhou, L.Y.; Chen, W.T.; Ye, N.H.; Xia, J.X.; Zhuang, C.X. Overexpression of a microRNA-targeted NAC transcription factor improves drought and salt tolerance in rice via ABA-mediated pathways. Rice 2019, 12, 76. [Google Scholar] [CrossRef] [PubMed]
- Takasaki, H.; Maruyama, K.; Kidokor, S.; Ito, Y.; Fujita, Y.; Shinozaki, K.; Yamaguchi-Shinozaki, K.; Nakashima, K. The abiotic stress-responsive NAC-type transcription factor OsNAC5 regulates stress-inducible genes and stress tolerance in rice. Mol. Genet. Genom. 2010, 284, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Song, S.Y.; Chen, Y.; Chen, J.; Dai, X.Y.; Zhang, W.H. Physiological mechanisms underlying OsNAC5-dependent tolerance of rice plants to abiotic stress. Planta 2011, 234, 331–345. [Google Scholar] [CrossRef]
- Nakashima, K.; Tran, L.S.P.; Nguyen, D.V.; Fujita, M.; Maruyama, K.; Todaka, D.; Ito, Y.; Hayashi, N.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of a NAC-type transcription factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice. Plant J. 2007, 51, 617–630. [Google Scholar] [CrossRef]
- Hu, H.H.; You, J.; Fang, Y.J.; Zhu, X.Y.; Qi, Z.Y.; Xiong, L.Z. Characterization of transcription factor gene SNAC2 conferring cold and salt tolerance in rice. Plant Mol. Biol. 2010, 67, 169–181. [Google Scholar] [CrossRef]
- Hu, H.H.; Dai, M.Q.; Yao, J.L.; Xiao, B.Z.; Li, X.H.; Zhang, Q.F.; Xiong, L.Z. Overexpressing a NAM ATAF and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proc. Natl. Acad. Sci. USA 2006, 103, 12987–12992. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.S.; Kim, Y.K.; Baek, K.H.; Jung, H.; Ha, S.H.; Choi, Y.D.; Kim, M.; Reuzeau, C.; Kim, J.K. Root-specific expression of OsNAC10 improves drought tolerance and grain yield in rice under field drought conditions. Plant Physiol. 2010, 153, 185–197. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Zhong, Z.H.; Hang, H.H.; Wang, X.; Liu, B.L.; Yang, L.J.; Han, X.Y.; Yu, D.S.; Zheng, X.L.; Wang, C.G.; et al. Targeted mutagenesis of NAC transcription factor gene OsNAC041 leading to salt sensitivity in rice. Rice 2019, 26, 98–108. [Google Scholar] [CrossRef]
- Yang, A.; Dai, X.Y.; Zhang, W.H. A R2R3-type MYB gene, OsMYB2, is involved in salt, cold, and dehydration tolerance in rice. J. Exp. Bot. 2012, 63, 2541–2556. [Google Scholar] [CrossRef]
- Dai, X.; Xu, Y.; Ma, Q.; Xu, W.; Wang, T.; Xue, Y.; Chong, K. Overexpression of an R1R2R3 MYB gene, OsMYB3R-2, increases tolerance to freezing, drought, and salt stress in transgenic Arabidopsis. Plant Physiol. 2007, 143, 1739–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, H.Y.; Li, J.J.; Liu, P.L.; Duan, J.Z.; Zhao, Y.; Guo, X.; Li, Y.; Zhang, H.L.; Ali, J.; Li, Z.C. Overexpression of OsMYB48-1 a novel MYB-related transcription factor enhances drought and salinity tolerance in rice. PLoS ONE 2014, 9, e92913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, R.; Schippers, J.H.M.; Mieulet, D.; Obata, T.; Fernie, A.R.; Guiderdoni, E.; Mueller-Roeber, B. MULTIPASS, a rice R2R3-type MYB transcription factor, regulates adaptive growth by integrating multiple hormonal pathways. Plant J. 2013, 76, 258–273. [Google Scholar] [CrossRef]
- Zhu, N.; Cheng, S.F.; Liu, X.Y.; Du, H.; Dai, M.Q.; Zhou, D.X.; Yang, W.J.; Zhao, Y. The R2R3-type MYB gene OsMYB91 has a function in coordinating plant growth and salt stress tolerance in rice. Plant Sci. 2015, 236, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Jing, W.; Xiao, L.Y.; Jin, Y.K.; Shen, L.K.; Zhang, W.H. The rice High-Affinity Potassium Transporter1;1 is involved in salt tolerance and regulated by an MYB-type transcription factor. Plant Physiol. 2015, 168, 1076–1090. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.A.; Cho, J.; Han, M.; Ahn, C.H.; Jeon, J.S.; An, G.; Park, P.B. The ABRE-binding bZIP transcription factor OsABF2 is a positive regulator of abiotic stress and ABA signaling in rice. J. Plant Physiol. 2010, 167, 1512–1520. [Google Scholar] [CrossRef]
- Zou, M.J.; Guan, Y.C.; Ren, H.B.; Zhang, F.; Chen, F. A bZIP transcription factor, OsABI5, is involved in rice fertility and stress tolerance. Plant Mol. Biol. 2008, 66, 675–683. [Google Scholar] [CrossRef]
- Xiang, Y.; Tang, N.; Du, H.; Ye, H.Y.; Xiong, L.Z. Characterization of OsbZIP23 as a key player of the basic leucine zipper transcription factor family for conferring abscisic acid sensitivity and salinity and drought tolerance in rice. Plant Physiol. 2008, 148, 1938–1952. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.T.; Mao, B.G.; Ou, S.J.; Wang, W.; Liu, L.C.; Wu, Y.B.; Chu, C.C.; Wang, X.P. OsbZIP71, a bZIP transcription factor, confers salinity and drought tolerance in rice. Plant Mol. Biol. 2014, 89, 19–36. [Google Scholar] [CrossRef]
- Das, P.; Lakra, N.; Nutan, K.K.; Singla-Pareek, S.L.; Pareek, A. A unique bZIP transcription factor imparting multiple stress tolerance in rice. Rice 2019, 12, 58. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.Y.; Chao, D.Y.; Gao, P.J.; Zhu, M.Z.; Shi, M.Z.; Lin, H.X. A previously unknown zinc finger protein, DST, regulates drought and salt tolerance in rice via stomatal aperture control. Genes Dev. 2009, 23, 1805–1817. [Google Scholar] [CrossRef] [Green Version]
- Jan, A.M.K.; Todaka, D.; Kidokoro, S.; Abo, M.; Yoshimura, E.; Shinozaki, K.; Nakashima, K.; Yamaguchi-Shinozaki, K. OsTZF1, a CCCH-tandem zinc finger protein, confers delayed senescence and stress tolerance in rice by regulating stress-related genes. Plant Physiol. 2013, 161, 1202–1216. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.J.; Guo, S.Q.; Yang, X.; Bao, Y.M.; Tang, H.J.; Sun, H.; Huang, J.; Zhang, H.S. Functional analysis of a novel Cys2/His2-type zinc finger protein involved in salt tolerance in rice. J. Exp. Bot. 2010, 61, 2807–2818. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Sun, S.J.; Xu, D.Q.; Lan, H.X.; Sun, H.; Wang, Z.F.; Bao, Y.M.; Wang, J.F.; Tang, H.J.; Zhang, H.S. A TFIIIA-type zinc finger protein confers multiple abiotic stress tolerances in transgenic rice (Oryza sativa L.). Plant Mol. Biol. 2012, 80, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lan, H.X.; Shao, Q.L.; Wang, R.Q.; Chen, H.; Tang, H.J.; Zhang, H.S.; Huang, J. An A20/AN1-type zinc finger protein modulates gibberellins and abscisic acid contents and increases sensitivity to abiotic stress in rice (Oryza sativa). J. Exp. Bot. 2016, 67, 315–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.Q.; Huang, J.; Guo, S.Q.; Yang, X.; Bao, Y.M.; Tang, H.J.; Zhang, H.S. Overexpression of a TFIIIA-type zinc finger protein gene ZFP252 enhances drought and salt tolerance in rice (Oryza sativa L.). FEBS Lett. 2008, 582, 1037–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, Q.J.; Ma, H.Y.; Zhang, Z.J.; Wang, Z.Y.; Bu, Q.Y.; Liu, S.K. A rice LSD1-like-type ZFP gene OsLOL5 enhances saline-alkaline tolerance in transgenic Arabidopsis thaliana, yeast and rice. BMC Genom. 2016, 17, 142. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Li, F.; Ma, Y.; Ching, K.; Xu, Y. Overexpression of OrbHLH001, a putative helix-loop-helix transcription factor, causes increased expression of AKT1 and maintains ionic balance under salt stress in rice. J. Plant Physiol. 2013, 170, 93–100. [Google Scholar] [CrossRef]
- Chen, H.C.; Cheng, W.H.; Hong, C.Y.; Chang, Y.S.; Chang, M.C. The transcription factor OsbHLH035 mediates seed germination and enables seedling recovery from salt stress through ABA-dependent and ABA-independent pathways respectively. Rice 2018, 11, 50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.X.; Haider, I.; Kohlen, W.; Jiang, L.; Bouwmeester, H.; Meijer, A.H.; Schluepmann, H.; Liu, C.M.; Ouwerkerk, P.B.F. Function of the HD-Zip I gene Oshox22 in ABA-mediated drought and salt tolerances in rice. Plant Mol. Biol. 2012, 80, 571–585. [Google Scholar] [CrossRef]
- Bang, S.W.; Lee, D.K.; Jung, H.; Chung, P.J.; Kim, Y.S.; Choi, Y.D.; Suh, J.W.; Kim, J.K. Overexpression of OsTF1L a rice HD-Zip transcription factor promotes lignin biosynthesis and stomatal closure that improves drought tolerance. Plant Biotechnol. J. 2019, 17, 118–131. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.Y.; Yu, C.Y.; Huang, L.L.; Wu, M.J.; Liu, B.H.; Liu, Y.H.; Song, G.; Liu, D.D.; Gan, Y.B. Overexpression of MADS-box transcription factor OsMADS25 enhances salt stress tolerance in Rice and Arabidopsis. Plant Growth Regul. 2020, 90, 163–171. [Google Scholar] [CrossRef]
- Tao, Z.; Kou, Y.J.; Liu, H.B.; Li, X.H.; Xiao, J.H.; Wang, S.P. OsWRKY45 alleles play different roles in abscisic acid signalling and salt stress tolerance but similar roles in drought and cold tolerance in rice. J. Exp. Bot. 2011, 62, 4863–4874. [Google Scholar] [CrossRef]
- Hasthanssombut, S.; Supaibulwatana, K.; Mii, M.; Nakamura, I. Genetic manipulation of Japonica rice using the OsBADH1 from Indica rice to improve salinity tolerance. Plant Cell Tissue Organ. Cult. 2011, 104, 79–89. [Google Scholar] [CrossRef]
- Tang, W.; Sun, J.Q.; Liu, J.; Liu, F.F.; Yan, J.; Guo, X.J.; Lu, B.R.; Liu, Y.S. RNAi-directed downregulation of betaine aldehyde dehydrogenase 1 (OsBADH1) results in decreased stress tolerance and increased oxidative markers without affecting glycine betaine biosynthesis in rice (Oryza sativa). Plant Mol. Biol. 2014, 86, 443–454. [Google Scholar] [CrossRef]
- Ge, L.F.; Chao, D.Y.; Shi, M.; Zhu, M.Z.; Gao, J.P.; Lin, H.X. Overexpression of the trehalose-6-phosphate phosphatase gene OsTPP1 confers stress tolerance in rice and results in the activation of stress responsive genes. Planta 2008, 228, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Li, H.W.; Zhang, B.S.; Deng, X.W.; Wang, X.P. Overexpression of the trehalose-6-phosphate synthase gene OsTPS1 enhances abiotic stress tolerance in rice. Planta 2011, 234, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Vishal, B.; Krishnamurthy, P.; Ramamoorthy, R.; Kumar, P.P. OsTPS8 controls yield-related traits and confers salt stress tolerance in rice by enhancing suberin deposition. New Phytol. 2018, 221, 1369–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Wang, Z.Y.; Lin, H.; Cui, W.E.; Chen, J.; Liu, M.; Chen, Z.L.; Qu, L.J.; Gu, H. Expression and functional analysis of the rice plasma-membrane intrinsic protein gene family. Cell Res. 2006, 16, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.W.; Fukumoto, T.; Matsumoto, T.; Gena, P.; Frascaria, D.; Kaneko, T.; Katsuhara, M.; Zhong, S.H.; Sun, X.L.; Zhu, Y.M.; et al. Aquaporin OsPIP1;1 promotes rice salt resistance and seed germination. Plant Physiol. Biochem. 2013, 63, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.Q.; Sheng, P.K.; Tan, J.J.; Chen, X.L.; Lu, G.W.; Ma, W.W.; Heng, Y.Q.; Lin, Q.B.; Zhu, S.S.; Wang, J.L.; et al. Plasma membrane receptor-like kinase leaf panicle 2 acts downstream of the DROUGHT AND SALT TOLERANCE transcription factor to regulate drought sensitivity in rice. J. Exp. Bot. 2015, 66, 271–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, J.; Zong, W.; Li, X.K.; Ning, J.; Hu, H.H.; Li, X.H.; Xiao, J.H.; Xiong, L.Z. The SNAC1-targeted gene OsSRO1c modulates stomatal closure and oxidative stress tolerance by regulating hydrogen peroxide in rice. J. Exp. Bot. 2013, 64, 569–583. [Google Scholar] [CrossRef] [Green Version]
- Guan, Q.; Liao, X.; He, M.; Li, X.; Wang, Z.; Ma, H.; Yu, S.; Liu, S. Tolerance analysis of chloroplast OsCu/Zn-SOD overexpressing rice under NaCl and NaHCO3 stress. PLoS ONE 2017, 12, e0186052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, Y.; Hibino, T.; Tanaka, A.; Kishitani, S.; Takabe, T.; Yokota, S.; Takabe, T. Salt tolerance of transgenic rice overexpressing yeast mitochondrial Mn-SOD in chloroplasts. Plant Sci. 1999, 148, 131–138. [Google Scholar] [CrossRef]
- Lu, Z.Q.; Liu, D.L.; Liu, S.K. Two rice cytosolic ascorbate peroxidases differentially improve salt tolerance in transgenic Arabidopsis. Plant Cell Rep. 2007, 26, 1909–1917. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.G.; Zhang, Q.A.; Wu, J.X.; Zheng, X.; Zheng, S.; Sun, X.H.; Qiu, Q.S.; Lu, T.G. Gene knockout study reveals that cytosolic ascorbate peroxidase 2 (OsAPX2) plays a critical role in growth and reproduction in rice under drought, salt and cold stresses. PLoS ONE 2013, 8, e57472. [Google Scholar] [CrossRef]
- Wu, T.M.; Lin, W.R.; Kao, Y.T.; Hsu, Y.T.; Yeh, C.H.; Hong, C.Y.; Kao, C.H. Identification and characterization of a novel chloroplast/mitochondria co-localized glutathione reductase 3 involved in salt stress response in rice. Plant Mol. Biol. 2013, 83, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.J.; Guo, Y. OsTRXh1 regulates the redox state of the apoplast and influences stress responses in rice. Plant Signal Behav. 2012, 7, 440–442. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Priya, P.; Jain, M. Modified expression of an auxin-responsive rice CC-type glutaredoxin gene affects multiple abiotic stress responses. Planta 2013, 238, 871–884. [Google Scholar] [CrossRef]
- Ning, X.; Sun, Y.; Wang, C.C.; Zhang, W.L.; Sun, M.H.; Hu, H.T.; Liu, J.Z.; Yang, L. A rice CPYC-type glutaredoxin OsGRX20 in protection against bacterial blight methyl viologen and salt stresses. Front. Plant Sci. 2018, 9, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, M.T.; Bandillo, N.; Razzaq, F.; Shiblawi, A.; Sharma, S.; Liu, K.; Schmitz, A.J.; Zhang, C.; Véry, A.A.; Lorenz, A.J.; et al. Allelic variants of OsHKT1;1 underlie the divergence between indica and japonica subspecies of rice (Oryza sativa) for root sodium content. PloS Genet. 2017, 13, e1006823. [Google Scholar] [CrossRef] [PubMed]
- Oda, Y.; Kobayashi, N.I.; Tanoi, K.; Ma, J.F.; Itou, Y.; Katsuhara, M.; Itou, T.; Horie, T. T-DNA tagging-based gain-of-function of OsHKT1;4 reinforces Na+ exclusion from leaves and stems but triggers Na toxicity in roots of rice under salt stress. Int. J. Mol. Sci. 2018, 19, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Z.H.; Gao, J.P.; Li, L.G.; Cai, X.L.; Huang, W.; Chao, D.Y.; Zhu, M.Z.; Wang, Z.Y.; Luan, S.; Lin, H.X. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Genet. 2005, 37, 1141–1146. [Google Scholar] [CrossRef]
- El Mahi, H.; Pérez-Hormaeche, J.; De Luca, A.; Villalta, I.; Espartero, J.; Gámez-Arjona, F.; Fernández, J.L.; Bundó, M.; Mendoza, I.; Mieulet, D.; et al. A critical role of sodium flux via the plasma membrane Na+/H+ exchanger SOS1 in the salt tolerance of rice. Plant Physiol. 2019, 180, 1046–1065. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.P.; Zheng, L.Q.; Xue, Y.H.; Zhang, Q.; Wang, L.; Shuo, H.X. Overexpression of OsVP1 and OsNHX1 increases tolerance to drought and salinity in rice. J. Plant Biol. 2010, 53, 444–452. [Google Scholar] [CrossRef]
- Chen, G.; Hu, Q.; Luo, L.; Yang, T.; Zhang, S.; Hu, Y.; Xu, G. Rice potassium transporter OsHAK1 is essential for maintaining potassium-mediated growth and functions in salt tolerance over low and high potassium concentration ranges. Plant Cell. Environ. 2009, 38, 2747–2765. [Google Scholar] [CrossRef]
- Yang, T.Y.; Zhang, S.; Hu, Y.B.; Wu, F.C.; Hu, Q.D.; Chen, G.; Cai, J.; Wu, T.; Moran, N.; Yu, L.; et al. The role of a potassium transporter OsHAK5 in potassium acquisition and transport from roots to shoots in rice at low potassium supply levels. Plant Physiol. 2014, 166, 945–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, H.M.; Tang, Q.; Cai, J.; Xu, B.C.; Xu, G.H.; Yu, L. Rice OsHAK16 functions in potassium uptake and translocation in shoot maintaining potassium homeostasis and salt tolerance. Planta 2019, 250, 549–561. [Google Scholar] [CrossRef]
- Shen, Y.; Shen, L.; Shen, Z.J.W.; Ge, H.; Zhao, J.; Zhang, W. The potassium transporter OsHAK 21 functions in the maintenance of ion homeostasis and tolerance to salt stress in rice. Plant Cell. Environ. 2015, 38, 2766–2779. [Google Scholar] [CrossRef] [PubMed]
- Nongpiur, R.; Soni, P.; Karan, R.; Singla-Pareek, S.L.; Pareek, A. Histidine kinases in plants. Plant Signal Behav. 2012, 7, 1230–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, L.S.P.; Urao, T.; Qin, F.; Maruyama, K.; Kakimoto, T.; Shonozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of AHK1/ATHK1 and cytokinin receptor histidine kinases in response to abscisic acid drought and salt stress in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 20623–20628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.N.; Jane, W.N.; Verslues, P.E. Role of the putative osmosensor Arabidopsis histidine kinase1 in dehydration avoidance and low-water-potential response. Plant Physiol. 2013, 161, 942–953. [Google Scholar] [CrossRef] [Green Version]
- Kushwaha, H.R.; Singla-Pareek, S.L.; Pareek, A. Putative osmosensor–OsHK3b–a histidine kinase protein from rice shows high structural conservation with its ortholog AtHK1 from Arabidopsis. J. Biomol. Struct. Dyn. 2014, 32, 1318–1332. [Google Scholar] [CrossRef] [Green Version]
- Ozakabe, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S.P. Sensing the environment: Key roles of membrane-localized kinases in plant perception and response to abiotic stress. J. Exp. Bot. 2013, 64, 445–458. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.L.; Zhang, L.Q.; Liu, N.; Xu, S.L.; Yue, Z.L.; Zhang, L.L.; Deng, Z.P.; Burlingame, A.L.; Sun, D.Y.; Wang, Z.Y.; et al. Mutual regulation of receptor-like kinase SIT1 and B’κ-PP2A shapes the early response of rice to salt stress. Plant Cell 2019, 31, 2131–2151. [Google Scholar] [CrossRef]
- Yuan, F.; Yang, H.; Xue, Y.; Kong, D.; Ye, R.; Li, C.; Zhang, J.; Theprungsirikul, L.; Shrift, T.; Krichilsky, B. OSCA1 mediates osmotic-stress-evoked Ca2+ increases vital for osmosensing in Arabidopsis. Nature 2014, 514, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.S.; Yuan, F.; Wen, Z.H.; Li, Y.H.; Wang, F.; Zhu, T.; Zhou, W.Q.; Jin, X.; Wang, Y.D.; Zhao, H.P.; et al. Genome-wide survey and expression analysis of the OSCA gene family in rice. BMC Plant Biol. 2015, 15, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.Z.; Ishitani, M.; Kim, C.S.; Zhu, J.K. The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc. Natl. Acad. Sci. USA 2000, 97, 6896–6901. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441–445. [Google Scholar] [CrossRef]
- Kader, M.A.; Lindberg, S.; Seidel, T.; Golldack, D.; Yemelyanov, V. Sodium sensing induces different changes in free cytosolic calcium concentration and pH in salt-tolerant and -sensitive rice (Oryza sativa) cultivars. Physiol. Plantarum. 2007, 130, 99–111. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism oxidative stress and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Batistic, O.; Kudla, J. Integration and channeling of calcium signaling through the CBL calcium sensor/CIPK protein kinase network. Planta 2004, 219, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Bouché, N.; Yellin, A.; Snedded, W.A.; Fromm, H. Plant-specific calmodulin-binding proteins. Annu. Rev. Plant Sci. 2005, 56, 435–466. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.Q.; Xu, L.Q.; Singh, A.; Wang, H.Z.; Du, L.Q.; Poovaiah, B.W. Involvement of calmodulin and calmodulin-like proteins in plant responses to abiotic stresses. Front. Plant Sci. 2015, 6, 600. [Google Scholar] [CrossRef] [Green Version]
- Yuenyong, W.; Chinpongpanich, A.; Comai, L.; Chadchawan, S.; Buaboocha, T. Downstream components of the calmodulin signaling pathway in the rice salt stress response revealed by transcriptome profiling and target identification. BMC Plant Biol. 2018, 18, 335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katou, S.; Kuroda, K.; Seo, S.; Yanagawa, Y.; Tsuge, T.; Yamazaki, M.; Miyao, A.; Hirochika, H.; Ohashi, Y. A calmodulin-binding mitogen-activated protein kinase phosphatase is induced by wounding and regulates the activities of stress-related mitogen-activated protein kinases in rice. Plant Cell. Physiol. 2007, 48, 332–344. [Google Scholar] [CrossRef]
- Yang, J.; Liu, S.; Ji, L.X.; Tang, X.Y.; Zhu, Y.S.; Xie, G.S. Identification of novel OsCML16 target proteins and differential expression analysis under abiotic stresses in rice. J. Plant Physiol. 2020, 249, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.Y.; Li, S.Z.; Xie, K.; Zhang, Q.; Wang, Y.; Tang, Y.; Hong, Y.G.; He, C.Y.; Liu, Y.L. Plant ERD2-like proteins function as endoplasmic reticulum luminal protein receptors and participate in programmed cell death during innate immunity. Plant J. 2012, 72, 57–69. [Google Scholar] [CrossRef]
- Asano, T.; Tanaka, N.; Yang, G.X.; Hayashi, N.; Komatsu, S. Genome-wide Identification of the Rice Calcium-dependent Protein Kinase and its Closely Related Kinase Gene Families: Comprehensive Analysis of the CDPKs Gene Family in Rice. Plant Cell. Physiol. 2005, 46, 356–366. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.L.; Jiang, S.F.; Liang, J.S.; Chen, M. Roles of plant CBL-CIPK systems in abiotic stress responses. Turk. J. Bot. 2019, 43, 271–280. [Google Scholar] [CrossRef]
- Martínez-Atienza, J.; Jiang, X.; Garciadeblas, B.; Mendoza, I.; Zhu, J.K.; Pardo, J.M.; Quintero, F.J. Conservation of the salt overly sensitive pathway in rice. Plant Physiol. 2007, 143, 1001–1012. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.W.; Zhang, T.Y.; Wang, H.; Zhao, N.; Liu, B. Heritable alteration in salt tolerance in rice induced by introgression from wild rice (Zizania latifolia). Rice 2012, 5, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanwar, P.; Sanyal, S.K.; Tokas, I.; Yadav, A.K.; Pandey, A.; Kapoor, S.; Pandey, G.K. Comprehensive structural interaction and expression analysis of CBL and CIPK complement during abiotic stresses and development in rice. Cell Calcium 2014, 56, 81–95. [Google Scholar] [CrossRef]
- Chen, X.; Gu, Z.; Liu, F.; Ma, B.; Zhang, H. Molecular analysis of rice CIPKs Involved in both biotic and abiotic stress responses. Rice Sci. 2016, 18, 1–9. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Rohila, J.S.; Yang, Y. Rice mitogen-activated protein kinase gene family and its role in biotic and abiotic stress response. J. Integr. Plant Biol. 2007, 49, 751–759. [Google Scholar] [CrossRef]
- Rodriguez, M.C.; Petersen, M.; Mundy, J. Mitogen-activated protein kinase signaling in plants. Annu. Rev. Plant Biol. 2010, 61, 621–649. [Google Scholar] [CrossRef]
- MAPK Group. Mitogen-activated protein kinase cascades in plants: A new nomenclature. Trends Plant Sci. 2002, 7, 301–308. [Google Scholar] [CrossRef]
- Sinha, A.K.; Jaggi, M.; Raghuram, B.; Tuteja, N. Mitogen-activated protein kinase signalling in plants under abiotic stress. Plant Signal Behav. 2011, 16, 196–203. [Google Scholar] [CrossRef] [Green Version]
- Kumar, K.; Wankhede, D.P.; Sinha, A.K. Signal convergence through the lenses of MAP kinases: Paradigms of stress and hormone signaling in plants. Front. Biol. 2013, 8, 109–118. [Google Scholar] [CrossRef]
- Yoo, S.J.; Kim, S.H.; Kim, M.J.; Ryu, C.M.; Kim, Y.C.; Cho, B.H.; Yang, K.Y. Involvement of the OsMKK4-OsMPK1 cascade and its downstream transcription factor OsWRKY53 in the wounding response in rice. Plant Path J. 2014, 30, 168–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohnert, H.J.; Ayoubi, P.; Borchert, C.; Bressan, R.A.; Burnap, R.L.; Cushman, J.C.; Deyholos MFischer RGalbraith, D.W.; Hasegawa, P.M.; Jenks, M.; Kawasaki, M.; et al. A genomics approach towards salt stress tolerance. Plant Physiol. Biochem. 2001, 39, 295–311. [Google Scholar] [CrossRef]
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Seki, M.; Narusaka, M.; Abe, H.; Kasuga, M.; Yamaguchi-Shinozaki, K.; Carninci, P.; Hayashizaki, Y.; Shinozaki, K. Monitoring the expression pattern of 1300 Arabidopsis genes under drought and cold stresses by using a full-length cDNA microarray. Plant Cell 2001, 13, 61–72. [Google Scholar] [CrossRef] [Green Version]
- Seki, M.; Ishida, J.; Narusaka, M.; Fujita, M.; Nanjo, T.; Umezawa, T.; Kamiya, A.; Nakajima, M.; Enju, A.; Sakurai, T.; et al. Monitoring the expression pattern of 7000 Arabidopsis genes under ABA treatments by using a full-length cDNA microarray. Plant J. 2020, 2, 282–291. [Google Scholar] [CrossRef]
- Gao, G.; Zhong, Y.F.; Guo, A.; Zhu, Q.; Tang, W.; Zheng, W. DRTF: A database of rice transcription factors. Bioinformatics 2006, 22, 1286–1287. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.X.; Liu, D.; Pan, Y.; Gong, W.; Ma, L.G.; Luo, J.C.; Deng, X.W.; Zhu, Y. An annotation update via cDNA sequence analysis and comprehensive profiling of developmental hormonal or environmental responsiveness of the Arabidopsis AP2/EREBP transcription factor gene family. Plant Mol. Biol. 2005, 59, 853–868. [Google Scholar] [CrossRef]
- Agarwal, P.K.; Gupta, K.; Lopato, S.; Agarwal, P. Dehydration responsive element binding transcription factors and their applications for the engineering of stress tolerance. J. Exp. Bot. 2017, 68, 2135–2148. [Google Scholar] [CrossRef]
- Ito, Y.; Katsura, K.; Maruyama, K.; Taji, T.; Kobayashi, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of rice DREB1/CBF-type transcription factors involved in cold-responsive gene expression in transgenic rice. Plant Cell. Physiol. 2006, 47, 141–153. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.Q.; Meng, X.P.; Zhang, Y.; Xia, M.; Wang, X.P. Over-expression of OsDREB genes lead to enhanced drought tolerance in rice. Biotechnol. Lett. 2008, 30, 2191–2198. [Google Scholar] [CrossRef]
- Danyluk, J.; Perron, A.; Houde, M.; Limin, A.; Fowler, B.; Benhamou, N.; Sarhan, F. Accumulation of an acidic dehydrin in the vicinity of the plasma membrane during cold acclimation of wheat. Plant Cell 1998, 10, 623–638. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.S. The role of DREB transcription factors in abiotic stress tolerance of plants. Agric. Environ. Biotechnol. 2011, 25, 2433–2442. [Google Scholar] [CrossRef] [Green Version]
- Puranik, S.; Sahu, P.P.; Srivastava, P.S.; Prasad, M. NAC proteins: Regulation and role in stress tolerance. Trends Plant Sci. 2007, 17, 369–381. [Google Scholar] [CrossRef]
- Li, C.N.; Ng, C.K.Y.; Fan, L.M. MYB transcription factors active players in abiotic stress signaling. Environ. Exp. Bot. 2015, 114, 80–91. [Google Scholar] [CrossRef]
- Li, J.; Han, G.; Sun, C.; Sui, N. Research advances of MYB transcription factors in plant stress resistance and breeding. Trends Plant Sci. 2019, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lakra, N.; Nutan, K.K.; Das, P.; Anwar KSingla-Pareek, S.L.; Pareek, A. A nuclear-localized histone-gene binding protein from rice (OsHBP1b) functions in salinity and drought stress tolerance by maintaining chlorophyll content and improving the antioxidant machinery. J. Plant Physiol. 2015, 176, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Li, W.T.; He, M.; Wang, J.; Wang, Y.P. Zinc finger protein (ZFP) in plants-A review. Plant Omics J. 2013, 6, 474–480. [Google Scholar]
- Murre, C.; Bain, G.; Dijk, M.A.V.; Engel, I.; Furnari, B.A.; Massari, M.E.; Matthews, J.R.; Quong, M.W.; Rivera, R.R.; Stuiver, M.H. Structure and function of helix–loop–helix proteins. BBA Biomembr. 1994, 1218, 129–135. [Google Scholar] [CrossRef]
- Wu, H.; Ye, H.Y.; Yao, R.F.; Zhang, T.; Xiong, L.Z. OsJAZ9 acts as a transcriptional regulator in jasmonate signaling and modulates salt stress tolerance in rice. Plant Sci. 2015, 232, 1–12. [Google Scholar] [CrossRef]
- Sun, X.; Wang, Y.; Sui, N. Transcriptional regulation of bHLH during plant response to stress. Biochem. Biophys. Res. Commun. 2018, 503, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.M.; Gregorio, G.; Oliveira, M.M.; Saibo, N.J.M. Five novel transcription factors as potential regulators of OsNHX1 gene expression in a salt tolerant rice genotype. Plant Mol. Biol. 2017, 93, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Rontein, D.; Basset, G.; Hanson, A.D. Metabolic engineering of osmoprotectant accumulation in plants. Metab. Eng. 2002, 4, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdallah, M.M.S.; Abdelgawad, Z.A.; El-Bassiuny, H.M.S. Alleviation of the adverse effects of salinity stress using trehalose in two rice varieties. South Afr. J. Bot. 2016, 103, 275–282. [Google Scholar] [CrossRef]
- Bhusan, D.; Das, D.K.; Hossain, M.; Murata, Y.; Hoque, M.A. Improvement of salt tolerance in rice (Oryza sativa L.) by increasing antioxidant defense systems using exogenous application of proline. Aus. J. Crop. Sci. 2016, 10, 50–56. [Google Scholar]
- Demiral, T.; Türkan, I. Exogenous glycine betaine affects growth and proline accumulation and retards senescence in two rice cultivars under NaCl stress. Environ. Exp. Bot. 2005, 56, 72–79. [Google Scholar] [CrossRef]
- Harinasut, P.; Tsutsui, K.; Takabe, T.; Nomura, M.; Takabe, T.; Kishitani, S. Exogenous glycinebetaine accumulation and increased salt-tolerance in rice seedlings. Biosci. Biotech. Bioch. 1996, 60, 366–368. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Ding, T.; Zuo, J.; Gao, L.; Fan, L. Amelioration of postharvest chilling injury in sweet pepper by glycine betaine. Postharvest Biol. Technol. 2016, 112, 114–120. [Google Scholar] [CrossRef]
- Chen, T.H.H.; Murata, N. Glycine betaine protects plants against abiotic stress: Mechanisms and biotechnological applications. Plant Cell. Environ. 2010, 34, 1–20. [Google Scholar] [CrossRef]
- Chaumont, F.; Tyerman, S.D. Aquaporins: Highly regulated channels controlling plant water relations. Plant Physiol. 2014, 164, 1600–1618. [Google Scholar] [CrossRef] [Green Version]
- Maurel, C.; Boursiac, Y.; Luu, D.T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, J.; Ishikawa, F.; Yamaguchi, T.; Uemura, M.; Maeshima, M. Identification of 33 rice aquaporin genes and analysis of their expression and function. Plant Cell. Physiol. 2005, 46, 1568–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawasaki, S.; Borchert, C.; Deyholos, M.; Wang, H.; Brazille, S.; Kawai, K.; Galbraith, D.; Bohnert, H.J. Gene expression profiles during the initial phase of salt stress in rice. Plant Cell 2001, 13, 889–905. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Miao, Y.; Song, C.P. Behind the scenes: The roles of reactive oxygen species in guard cells. New Phytol. 2014, 201, 1121–1140. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.G.; Shan, J.X.; Shi, M.; Gao, J.P.; Lin, H.X. DCA1 acts as a transcriptional co-activator of DST and contributes to drought and salt tolerance in rice. PLoS Genet. 2015, 11, e1005617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef]
- Mishra, P.; Bhoomika, K.; Dubey, R.S. Differential responses of antioxidative defense system to prolonged salinity stress in salt-tolerant and salt-sensitive Indica rice (Oryza sativa L.) seedlings. Protoplasma 2013, 250, 3–19. [Google Scholar] [CrossRef]
- Rossatto, T.; do Amaral, M.N.; Benitz, L.C.; Vighi, I.L.; Braga, E.J.B.; Júnior, A.M.D.; Maia, M.A.C.; Pinto, L.D. Gene expression and activity of antioxidant enzymes in rice plants, cv. BRS AG, under saline stress. Physiol. Mol. Biol. Plants 2017, 23, 865–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Ahsan, N.; Lee, K.W.; Kim, D.H.; Lee, D.G.; Kwak, S.S. Simultaneous overexpression of both Cu/Zn superoxide dismutase and ascorbate peroxidase in transgenic tall fescue plants confers increased tolerance to a wide range of abiotic stresses. J. Plant Physiol. 2007, 164, 1626–1638. [Google Scholar] [CrossRef]
- Wang, Y.; Ying, Y.; Chen, J.; Wang, X. Transgenic Arabidopsis, overexpressing Mn-SOD enhanced salt-tolerance. Plant Sci. 2004, 167, 671–677. [Google Scholar] [CrossRef]
- Cheng, Y.W.; Kong, X.W.; Wang, N.; Wang, T.T.; Chen, J.; Shi, Z.Q. Thymol confers tolerance to salt stress by activating anti-oxidative defense and modulating Na+ homeostasis in rice root. Ecotoxicol. Environ. Saf. 2020, 188, 109894. [Google Scholar] [CrossRef] [PubMed]
- Mhamdi, A.; Queval, G.; Chaouch, S.; Vanderauwera, S.; Breusegem, F.V.; Noctor, G. Catalase function in plants: A focus on Arabidopsis mutants as stress-mimic models. J. Exp. Bot. 2010, 61, 4197–4220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Poovaiah, B.W. Hydrogen peroxide homeostasis: Activation of plant catalases by calcium/calmodulin. Proc. Natl. Acad. Sci. USA 2002, 99, 4097–4102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, J.; Lee, Y.H.; Song, S.I. Rice CatA, CatB, and CatC are involved in environmental stress response, root growth, and photorespiration, respectively. J. Plant Biol. 2002, 57, 375–382. [Google Scholar] [CrossRef]
- Zhou, Y.B.; Liu, C.; Tang, D.Y.; Yan, L.; Wang, D.; Yang, Y.Z.; Gui, J.S.; Zhao, X.Y.; Li, L.G.; Tang, X.D.; et al. The receptor-like cytoplasmic kinase STRK1 phosphorylates and activates CatC, thereby regulating H2O2 homeostasis and improving salt tolerance in rice. Plant Cell 2018, 30, 1100–1118. [Google Scholar] [CrossRef] [Green Version]
- Kibria, M.G.; Hossain, M.; Murata, Y.; Hoque, M.A. Antioxidant defense mechanisms of salinity tolerance in rice genotypes. Rice Sci. 2017, 24, 155–162. [Google Scholar] [CrossRef]
- Yamane, K.; Mitsuya, S.; Taniguchi, M.; Miyake, H. Transcription profiles of genes encoding catalase and ascorbate peroxidase in the rice leaf tissues under salinity. Plant Prod. Sci. 2010, 13, 164–168. [Google Scholar] [CrossRef]
- Wutipraditkul, N.; Boonkomrat, S.; Buaboocha, T. Cloning and characterization of catalases from rice, Oryza sativa L. Biosci Biotechnol Biochem 2011, 75, 1900–1906. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, F.K.; Menezes-Benzvente, L.; Margis, R.; Margis-Pinheiro, M. Analysis of the molecular evolutionary history of the ascorbate peroxidase gene family: Inferences from the rice genome. J. Mol. Evol. 2004, 59, 761–770. [Google Scholar] [CrossRef]
- Hong, C.Y.; Hsu, Y.T.; Tsai, Y.C.; Kao, C.H. Expression of Ascorbate Peroxidase 8 in roots of rice (Oryza sativa L) seedlings in response to NaCl. J. Exp. Bot. 2007, 58, 3273–3283. [Google Scholar] [CrossRef]
- Bonifacio, A.; Martins, M.O.; Ribeiro, C.V.; Fontenele, A.V.; Carvalho, F.E.; Margis-Pinheiro, M.; Silveira, J.G. Role of peroxidases in the compensation of cytosolic ascorbate peroxidase knockdown in rice plants under abiotic stress. Plant Cell. Environ. 2011, 34, 1705–1722. [Google Scholar] [CrossRef]
- Rosa, S.B.; Caverzan, A.; Teixeira, F.K.; Lazzarotto, F.; Silveira, J.A.G.; Ferreira-Silva, S.L.; Abreu-Neto, J.; Margis, R.; Margis-Pinheiro, M. Cytosolic APx knockdown indicates an ambiguous redox response in rice. Phytochem 2010, 71, 548–558. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. 1998, 49, 249–279. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.M.; Lin, W.R.; Kao, C.H.; Hong, C.Y. Gene knockout of glutathione reductase 3 results in increased sensitivity to salt stress in rice. Plant Mol. Biol. 2015, 87, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.Y.; Chao, Y.Y.; Yang, M.Y.; Cheng, S.Y.; Cho, S.C.; Kao, C.H. NaCl-induced expression of glutathione reductase in roots of rice (Oryza sativa L.) seedlings is mediated through hydrogen peroxide but not abscisic acid. Plant Soil 2009, 320, 103–115. [Google Scholar] [CrossRef]
- Tsai, Y.C.; Hong, Y.C.; Kiu, L.F.; Kao, C.H. Expression of ascorbate peroxidase and glutathione reductase in roots of rice seedlings in response to NaCl and H2O2. J. Plant Physiol. 2005, 162, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, A. Thioredoxin and glutaredoxin systems. J. Biol. Chem. 1989, 264, 13963–13966. [Google Scholar] [CrossRef]
- Garg, R.; Jhanwar, S.; Tyagi, A.K.; Jain, M. Genome-wide survey and expression analysis suggest diverse roles of glutaredoxin gene family members during development and response to various stimuli in rice. DNA Res. 2010, 17, 353–367. [Google Scholar] [CrossRef] [Green Version]
- Nuruzzaman, M.; Gupta, M.; Zhang, C.; Wang, L.; Xie, W.; Xiong, L.; Zhang, Q.; Lian, X. Sequence and expression analysis of the thioredoxin protein gene family in rice. Mol. Genet. Genom. 2008, 280, 139–151. [Google Scholar] [CrossRef]
- Zhang, C.J.; Zhao, B.C.; Ge, W.N.; Zhang, F.Y.; Song, Y.; Sun, D.Y.; Guo, Y. An apoplastic H-type thioredoxin is involved in the stress response through regulation of the apoplastic reactive oxygen species in rice. Plant Physiol. 2011, 157, 1884–1899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, G.S.; Kato, H.; Sasaki, K.; Imai, R. A cold-induced thioredoxin h of rice OsTrx23 negatively regulates kinase activities of OsMPK3 and OsMPK6 in vitro. Febs Lett. 2009, 583, 2734–2738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blumwald, E.; Aharon, G.S.; Apse, M.P. Sodium transport in plant cells. Biochem. Biophys. Acta 2000, 1465, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.J.; Ding, L.; Zhu, J.K. SOS1, a genetic locus essential for salt tolerance and potassium acquisition. Plant Cell. 1996, 8, 617–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garciadeblas, B.; Senn, M.E.; Banuelos, M.A.; Rodriguez-Navarro, A. Sodium transport and HKT transporters: The rice model. Plant J. 2003, 34, 788–801. [Google Scholar] [CrossRef]
- Horie, T.; Yoshida, K.; Nakayama, H.; Yamada, K.; Oiki, S.; Shinmyo, A. Two types of HKT transporters with different properties of Na+ and K+ transport in Oryza sativa. Plant J. 2001, 27, 129–138. [Google Scholar] [CrossRef]
- Suzuki, K.; Yamaji, N.; Costa, A.; Okuma, E.; Kobayashi, N.I.; Kashiwagi, T.; Katsuhara, M.; Wang, C.; Tanoi, K.; Murata, Y.; et al. OsHKT1;4-mediated Na+ transport in stems contributes to Na+ exclusion from leaf blades of rice at the reproductive growth stage upon salt stress. BMC Plant Biol. 2016, 16, 22. [Google Scholar] [CrossRef] [Green Version]
- Imran, S.; Horie, T.; Katsuhara, M. Expression and ion transport activity of rice OsHKT1;1 variants. Plants 2020, 9, 16. [Google Scholar] [CrossRef] [Green Version]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Glenn, E.P.; Brown, J.J.; Blumwald, E. Salt tolerance and crop potential of halophytes. Crit. Rev. Plant Sci. 1999, 18, 227–255. [Google Scholar] [CrossRef]
- Kader, M.A.; Lindberg, S. Cytosolic calcium and pH signaling in plants under salinity stress. Plant Signal Behav. 2010, 5, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, A.; Nakamura, A.; Hara, N.; Toki, S.; Tanaka, Y. Molecular and functional analyses of rice NHX-type Na+/H+ antiporter genes. Planta 2011, 233, 175–188. [Google Scholar] [CrossRef]
- Amin, U.S.M.; Biswas, S.; Elias, S.M.; Razzaque, S.; Haque, T.; Malo, R.; Seraj, Z.I. Enhanced salt tolerance conferred by the complete 23 kb cDNA of the rice vacuolar Na+/H+ antiporter gene compared to 19 kb coding region with 5’ UTR in transgenic lines of rice. Front. Plant Sci. 2016, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brini, F.; Hanin, M.; Mezghani, I.; Berkowitz, G.A.; Masmou, K. Overexpression of wheat Na+/H+ antiporter TNHX1 and H+-pyrophosphatase TVP1 improve salt- and drought-stress tolerance in Arabidopsis thaliana plants. J. Exp. Bot. 2007, 58, 301–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaxiola, R.A.; Li, J.S.; Undurraga, S.; Dang, L.M.; Allen, G.J.; Alper, S.L.; Fink, G.R. Drought- and salt-tolerant plants result from overexpression of the AVP1 H+-pump. Proc. Natl. Acad. Sci. USA 2005, 98, 11444–11449. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.Y.; Zhang, X.J.; Li, P.H.; Zhao, Y.X.; Zhang, H. Co-expression of the Suaeda salsa SsNHX1 and Arabidopsis AVP1 confer greater salt tolerance to transgenic rice than the single SsNHX1. Mol. Breed. 2006, 17, 341–353. [Google Scholar] [CrossRef]
- Gong, H.J.; Randall, D.P.; Flowers, T.J. Silicon deposition in the root reduces sodium uptake in rice (Oryza sativa L.) seedlings by reducing bypass flow. Plant Cell. Environ. 2006, 1273, 433. [Google Scholar] [CrossRef]
- Krishnamurthy, P.; Ranathunge, K.; Franke, R.; Prakash, H.S.; Schreiber, L.; Mathew, M.K. The role of root apoplastic transport barriers in salt tolerance of rice (Oryza sativa L.). Planta 2009, 230, 119–134. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Salt sensing and signaling involved in rice responses to salt stress. Under high salinity, salt-induced osmotic stress begins, which is sensed by putative osmosensor OsHK3b, activated by OsHpt2. SIT1 also acts as a sensor via elevated kinase activity and induces reactive oxygen species (ROS) production and mitogen-activated protein kinase (MAPK) signaling. The activity of SIT1 is deactivated by the B’κ-PP2A subunit. Later, ionic stress occurs and is sensed by an unknown Na+ sensor. The Na+ enters the mature epidermal cell through nonselective cation channel (NSCC), causing membrane depolarization, and is polarized by P-type ATPases. Excess salt triggers a spike in the concentration of cytosolic secondary messengers, including Ca2+, reactive oxygen species (ROS), and phosphatidic acid (PA). ROS triggers Ca2+ influx through the cyclic nucleotide-gated ion channel (CNGC), activated by an unknown molecule. Ca2+ not only decreases K+ efflux but also induces further ROS accumulation; thus, a positive feedback loop exists between Ca2+ and ROS. The cytosolic Ca2+ also induces vacuolar Ca2+ release and activates Ca2+-binding proteins, such as OsCIPK24-OsCBL4 complex. This complex, together with MAPK, activated by phosphatidic acid, upregulates the OsSOS1 to remove cytosolic Na+. The vacuolar OsNHX1 gene is activated by OsCPK21, whereas the V-type ATPase is activated by OsCIPK24, establishing a proton gradient and driving the activity of OsNHX1.
Figure 1.
Salt sensing and signaling involved in rice responses to salt stress. Under high salinity, salt-induced osmotic stress begins, which is sensed by putative osmosensor OsHK3b, activated by OsHpt2. SIT1 also acts as a sensor via elevated kinase activity and induces reactive oxygen species (ROS) production and mitogen-activated protein kinase (MAPK) signaling. The activity of SIT1 is deactivated by the B’κ-PP2A subunit. Later, ionic stress occurs and is sensed by an unknown Na+ sensor. The Na+ enters the mature epidermal cell through nonselective cation channel (NSCC), causing membrane depolarization, and is polarized by P-type ATPases. Excess salt triggers a spike in the concentration of cytosolic secondary messengers, including Ca2+, reactive oxygen species (ROS), and phosphatidic acid (PA). ROS triggers Ca2+ influx through the cyclic nucleotide-gated ion channel (CNGC), activated by an unknown molecule. Ca2+ not only decreases K+ efflux but also induces further ROS accumulation; thus, a positive feedback loop exists between Ca2+ and ROS. The cytosolic Ca2+ also induces vacuolar Ca2+ release and activates Ca2+-binding proteins, such as OsCIPK24-OsCBL4 complex. This complex, together with MAPK, activated by phosphatidic acid, upregulates the OsSOS1 to remove cytosolic Na+. The vacuolar OsNHX1 gene is activated by OsCPK21, whereas the V-type ATPase is activated by OsCIPK24, establishing a proton gradient and driving the activity of OsNHX1.

Figure 2.
Transcriptional regulation involved in activating salt stress-responsive genes in rice. The transcriptional regulation occurs via abscisic acid (ABA)-dependent and -independent pathway, whereby transcription factors (TFs) bind with their corresponding cis-regulatory element. The APETALA2/ethylene responsive factor (AP2/ERF) and NAC (NAM, ATAF, and CUC) TFs operate in an ABA-independent pathway. NAC TFs regulate other TFs, such as dehydration responsive element-binding (DREB), myeloblastosis (MYB), and basic leucine-zipper (bZIP). The MYB, bZIP, zinc finger (ZF), basic-helix-loop-helix (bHLH), DREB, and other TFs are involved in the ABA-dependent pathway.
Figure 2.
Transcriptional regulation involved in activating salt stress-responsive genes in rice. The transcriptional regulation occurs via abscisic acid (ABA)-dependent and -independent pathway, whereby transcription factors (TFs) bind with their corresponding cis-regulatory element. The APETALA2/ethylene responsive factor (AP2/ERF) and NAC (NAM, ATAF, and CUC) TFs operate in an ABA-independent pathway. NAC TFs regulate other TFs, such as dehydration responsive element-binding (DREB), myeloblastosis (MYB), and basic leucine-zipper (bZIP). The MYB, bZIP, zinc finger (ZF), basic-helix-loop-helix (bHLH), DREB, and other TFs are involved in the ABA-dependent pathway.

Figure 3.
Rice salt tolerance adaptive mechanisms. In the leaf, (a) stomatal closure mediated either by DST or SNAC1 is the initial response of rice to salinity. Salt stress downregulates DST which interacts with DCA1 and activates OsPrx24 and LP2. Conversely, SNAC1 is upregulated, activating the OsSRO1c. These downstream genes mediate stomatal closure via H2O2 inhibition. (b,c) Na+ content in the leaf cytoplasm is controlled by vacuolar sequestration, xylem unloading, and phloem loading. Excess Na+ is sequestered into the vacuole via OsNHX1 coupled with H+-pump and OsVP1, a vacuolar-type H+-pyrophosphatase encoding gene. Na+ unloading at the xylem and Na+ loading at the phloem are mediated by OsHKT1;4 and OsHKT1;1, respectively. In the root, (d) Na+ is loaded at the xylem through nonselective cation channel (NSCC) and OsSOS1 coupled with H+-pump. Conversely, OsHKT1;5 unloads the Na+ ions from the xylem and shuttles them back to the parenchyma cells. Apart from Na+, K+ influx occurs mediated by OsHAK21, thereby increasing the K+/Na+ ratio. (e) Enhanced suberin deposition in the root exodermis and endodermis also inhibits Na+ influx to the stele. Similarly, it blocks water transport out of the stele. (f) The plasma membrane-bound OsPIP2;2 gene increases hydraulic conductivity in the root endodermis, allowing water uptake. (g) Na+ enters the root epidermis via NSCC and is shuttled back to the external medium via the OsSOS1 coupled with H+-pump.
Figure 3.
Rice salt tolerance adaptive mechanisms. In the leaf, (a) stomatal closure mediated either by DST or SNAC1 is the initial response of rice to salinity. Salt stress downregulates DST which interacts with DCA1 and activates OsPrx24 and LP2. Conversely, SNAC1 is upregulated, activating the OsSRO1c. These downstream genes mediate stomatal closure via H2O2 inhibition. (b,c) Na+ content in the leaf cytoplasm is controlled by vacuolar sequestration, xylem unloading, and phloem loading. Excess Na+ is sequestered into the vacuole via OsNHX1 coupled with H+-pump and OsVP1, a vacuolar-type H+-pyrophosphatase encoding gene. Na+ unloading at the xylem and Na+ loading at the phloem are mediated by OsHKT1;4 and OsHKT1;1, respectively. In the root, (d) Na+ is loaded at the xylem through nonselective cation channel (NSCC) and OsSOS1 coupled with H+-pump. Conversely, OsHKT1;5 unloads the Na+ ions from the xylem and shuttles them back to the parenchyma cells. Apart from Na+, K+ influx occurs mediated by OsHAK21, thereby increasing the K+/Na+ ratio. (e) Enhanced suberin deposition in the root exodermis and endodermis also inhibits Na+ influx to the stele. Similarly, it blocks water transport out of the stele. (f) The plasma membrane-bound OsPIP2;2 gene increases hydraulic conductivity in the root endodermis, allowing water uptake. (g) Na+ enters the root epidermis via NSCC and is shuttled back to the external medium via the OsSOS1 coupled with H+-pump.


{kind=link}
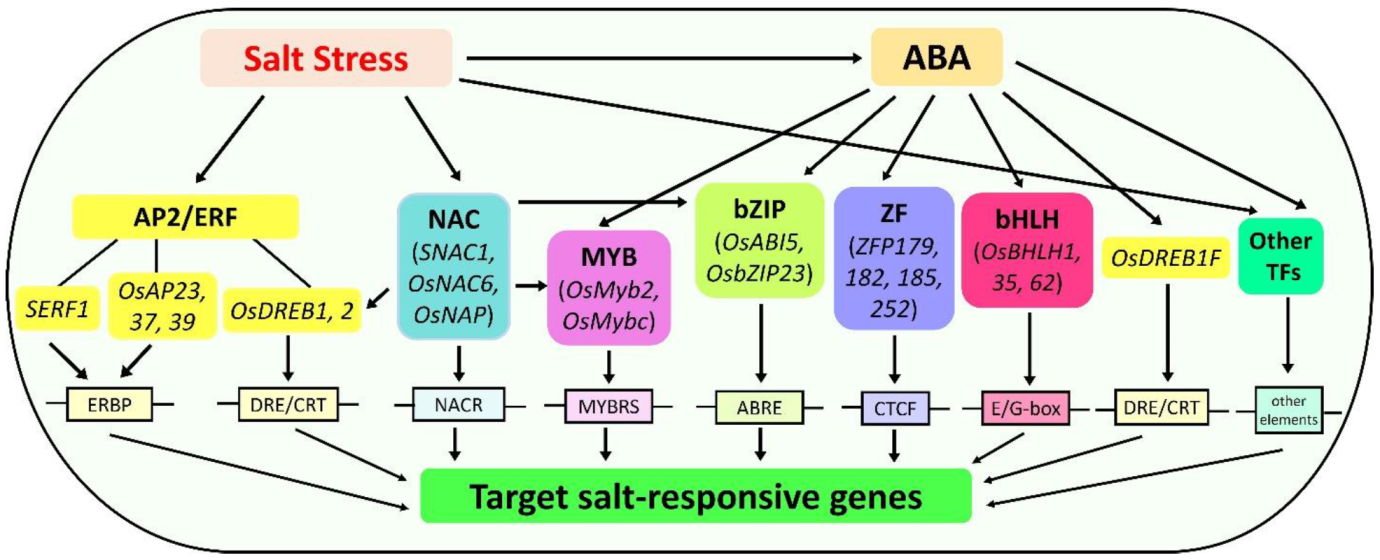{kind=link}
{kind=link}
Table 1.
List of functionally validated candidate genes involved from sensing to development of salt tolerance adaptive mechanisms in rice.
Table 1.
List of functionally validated candidate genes involved from sensing to development of salt tolerance adaptive mechanisms in rice.
| Gene Name | Gene ID | Functional Annotation | Method of Validation | * Regulation Role | References |
|---|---|---|---|---|---|
| Osmosensing | |||||
| SIT1 | LOC_Os02g42780 | lectin receptor-type protein kinase | Knockdown Overexpression | – | [9] |
| Signaling | |||||
| OsCam1-1 | LOC_Os03g20370 | Calmodulin | Overexpression | + | [10] |
| OsCPK4 | LOC_Os02g03410 | CAMK_CAMK_like.12—Ca2+/calmodulin-dependent protein kinase(CAMK) includes calcium/calmodulin dependent protein kinases | Knockdown Overexpression | + | [11] |
| OsCDPK7 | LOC_Os04g49510 | CAMK_CAMK_like.27—CAMK includes calcium/calmodulin dependent protein kinases | Overexpression | + | [12,13] |
| OsCPK12 | LOC_Os04g47300 | CAMK_CAMK_like.26—CAMK includes calcium/calmodulin dependent protein kinases | Overexpression | + | [14] |
| OsCPK21 | LOC_Os08g42750 | CAMK_CAMK_like.37—CAMK includes calcium/calmodulin dependent protein kinases | Overexpression | + | [15,16] |
| OsCIPK15 | LOC_Os11g02240 | CAMK_Nim1_like.4—CAMK includes calcium/calmodulin dependent protein kinases | Overexpression | + | [17] |
| OsCIPK31 | LOC_Os03g20380 | CAMK_Nim1_like.2—CAMK includes calcium/calmodulin dependent protein kinases | Mutant | + | [18] |
| OsMAPK5 | LOC_Os03g17700 | CGMC_MAPKCGMC_2_ERK.2—CGMC includes CDA, MAPK, GSK3, and CLKC kinases | Knockdown Overexpression | + | [19] |
| OsMAPK33 | LOC_Os02g05480 | CGMC_MAPKCMGC_2_SLT2y_ERK.1—includes cytidine deaminase (CDA), glycogen synthase kinase 3 (GSK3), mitogen-activated protein kinase (MAPK), and CLKC kinases | Knockdown Overexpression | – | [20] |
| OsMKK1 | LOC_Os06g05520 | MAPK | Knockdown | + | [21] |
| OsMKK6 | LOC_Os01g32660 | STE_MEK_ste7_MAP2K.2—STE kinases | Overexpression | + | [22] |
| OsMaPKKK63 | LOC_Os01g50370 | STE_MEKK_ste11_MAP3K.4—STE kinases | Knockdown | – | [23] |
| Transcriptional regulation | |||||
| OsDREB1A | LOC_Os09g35030 | Dehydration-responsive element (DRE)--binding protein | Overexpression | + | [24] |
| OsDREB1D | LOC_Os06g06970 | DRE--binding protein | Overexpression | + | [25] |
| OsDREB1F | LOC_Os01g73770 | DRE--binding protein | Overexpression | + | [26] |
| OsDREB2A | LOC_Os01g07120 | APETALA2 (AP2) domain containing protein | Overexpression | + | [27,28] |
| OsDREB2B | LOC_Os05g27930 | AP2 domain containing protein | Overexpression | + | [29] |
| OsAP23 | LOC_Os03g05590 | AP2 domain containing protein | Overexpression | – | [30] |
| OsAP37 | LOC_Os01g58420 | AP2 domain containing protein | Overexpression | + | [31] |
| OsSTAP1 | LOC_Os03g08470 | APETALA2/ethylene responsive factor (AP2/ERF)-type transcription factor | Overexpression | + | [32] |
| OsDREB6 | LOC_Os09g20350 | ERF transcription factor | Knockdown Overexpression | + | [33] |
| SERF1 | LOC_Os05g34730 | ERF020- transcription factor | Knockdown | + | [34] |
| OsERF922 | LOC_Os01g54890 | Ethylene-responsive transcription factor 2 | Knockdown Overexpression | – | [35] |
| OsRAV2 | LOC_Os01g04800 | B3 DNA binding domain containing protein | Mutant | + | [36] |
| OsNAP | LOC_Os03g21060 | No apical meristem (NAM)protein | Overexpression | + | [37] |
| ONAC022 | LOC_Os03g04070 | NAM protein | Overexpression | + | [38] |
| ONAC045 | LOC_Os11g03370 | NAM protein | Overexpression | + | [39] |
| ONAC063 | LOC_Os08g33910 | NAM protein | Overexpression | + | [40] |
| ONAC106 | LOC_Os08g33670 | NAM protein | Overexpression | + | [41] |
| OsNAC2 | LOC_Os04g38720 | NAM protein | Overexpression | + | [42,43] |
| OsNAC5 | LOC_Os11g08210 | NAM protein | Knockdown Overexpression | + | [44,45] |
| OsNAC6/SNAC2 | LOC_Os01g66120 | NAM protein | Overexpression | + | [46,47] |
| SNAC1 | LOC_Os03g60080 | NAM, ATAF and CUC (NAC) domain-containing protein 67 | Overexpression | + | [48] |
| OsNAC10 | LOC_Os11g03300 | NAC domain transcription factor | Overexpression | + | [49] |
| OsNAC041 | - | - | Knockdown | + | [50] |
| OsMYB2 | LOC_Os03g20090 | Myeloblastosis (MYB) family transcription factor | Overexpression | + | [51] |
| OsMYB3R-2 | LOC_Os01g62410 | MYB family transcription factor | Overexpression | + | [52] |
| OsMYB48-1 | LOC_Os01g74410 | MYB family transcription factor | Overexpression | + | [53] |
| OsMPS | LOC_Os02g40530 | MYB family transcription factor | Overexpression | + | [54] |
| OsMYB91 | LOC_Os12g38400 | MYB family transcription factor | Knockdown Overexpression | + | [55] |
| OsMYBc | LOC_Os09g12770 | Adenosine-thymine (AT) hook motif domain containing protein | Mutant | + | [56] |
| OsABF2 | LOC_Os06g10880 | Basic leucine-zipper (bZIP) transcription factor | Mutant | + | [57] |
| OsABI5 | LOC_Os01g64000 | bZIP transcription factor | Overexpression | – | [58] |
| OsbZIP23 | LOC_Os02g52780 | bZIP transcription factor | Overexpression | + | [59] |
| OsbZIP71 | LOC_Os09g13570 | CPuORF2—conserved peptide uORF-containing transcript | Knockdown Overexpression | + | [60] |
| OsHBP1b | LOC_Os01g17260 | Transcription factor | Overexpression | + | [61] |
| DST | LOC_Os03g57240 | ZOS3-19—C2H2 zinc finger (ZF) protein | Mutant | – | [62] |
| OsTZF1 | LOC_Os05g10670 | ZF CCCH type family protein | Knockdown Overexpression | + | [63] |
| ZFP179 | LOC_Os01g62190 | ZOS1-15—C2H2 ZF protein | Overexpression | + | [64] |
| ZFP182 | LOC_Os03g60560 | ZOS3-21—C2H2 ZF protein | Overexpression | + | [65] |
| ZFP185 | LOC_Os02g10200 | ZF A20 and AN1 domain-containing stress-associated protein | Knockdown Overexpression | – | [66] |
| ZFP252 | LOC_Os12g39400 | ZOS12-09—C2H2 ZF protein | Knockdown Overexpression | + | [67] |
| OsLOL5 | LOC_Os01g42710 | LSD1-like-type ZF protein | Overexpression | + | [68] |
| OrbHLH001 | LOC_Os01g70310 | Inducer of CBF expression 2 | Overexpression | + | [69] |
| OsbHLH035 | LOC_Os01g06640 | Basic helix-loop-helix (bHLH) | Mutant | + | [70] |
| Oshox22 | LOC_Os04g45810 | Homeobox associated leucine zipper | Mutant Overexpression | – | [71] |
| OsTF1L | LOC_Os08g19590 | Homeobox domain containing protein | Knockdown Overexpression | + | [72] |
| OsMADS25 | LOC_Os04g23910 | MADS-box family gene with MIKCc type-box | Knockdown Overexpression | + | [73] |
| OsWRKY45 | LOC_Os05g25770 | WRKY45 | Knockdown Overexpression | – | [74] |
| Osmoprotection | |||||
| OsBADH1 | LOC_Os04g39020 | Aldehyde dehydrogenase | Knockdown; Overexpression | + | [75,76] |
| OsTPP1 | LOC_Os02g44230 | CPuORF22—conserved peptide uORF-containing transcript | Overexpression | + | [77] |
| OsTPS1 | LOC_Os05g44210 | Trehalose-6-phosphate synthase | Overexpression | + | [78] |
| OsTPS8 | LOC_Os08g34580 | Trehalose-6-phosphate synthase | Mutant | + | [79] |
| Overexpression | |||||
| Osmoregulation | |||||
| OsPIP1;1 | LOC_Os02g44630 | Aquaporin protein | Overexpression | + | [80,81] |
| OsPIP2;2 | LOC_Os02g41860 | Aquaporin protein | Overexpression | + | [80] |
| Stomatal Closure | |||||
| LP2 | LOC_Os02g40240 | Receptor kinase | Overexpression | + | [82] |
| OsSRO1c | LOC_Os03g12820 | ATP8 | Mutant Overexpression | + | [83] |
| Antioxidation | |||||
| OsCu/Zn-SOD | LOC_Os08g44770 | Copper/zinc superoxide dismutase | Overexpression | + | [84] |
| OsMn-SOD | LOC_Os05g25850 | Manganese superoxide dismutase | Overexpression | + | [85] |
| OsAPx1 | LOC_Os03g17690 | Cytosolic Ascorbate Peroxidase encoding gene 1-8 | Overexpression | + | [86] |
| OsAPx2 | LOC_Os07g49400 | Cytosolic Ascorbate Peroxidase encoding gene 4,5,6,8 | Knockdown | + | [87] |
| Overexpression | |||||
| OsGR3 | LOC_Os10g28000 | Glutathione reductase | Knockdown | + | [88] |
| OsTRXh1/OsTrx23 | LOC_Os07g08840 | Thioredoxin | Knockdown; Overexpression | – | [89] |
| OsGRX8 | LOC_Os02g30850 | OsGrx_C8—Glutaredoxin subgroup III | Knockdown; Overexpression | + | [90] |
| OsGRX20 | LOC_Os08g44400 | Glutathione S-transferase | Knockdown; Overexpression | + | [91] |
| Na+ exclusion | |||||
| OsHKT1;1 | LOC_Os04g51820 | Na+ transporter | Natural variation | + | [92] |
| OsHKT1;4 | LOC_Os04g51830 | Na+ transporter | Mutant | – | [93] |
| OsHKT1;5/SKC1 | LOC_Os01g20160 | Na+ transporter | Natural variation | + | [94] |
| OsSOS1 | LOC_Os12g44360 | Sodium/hydrogen exchanger 7 | Mutant | + | [95] |
| Na+ compartmentation | |||||
| OsNHX1 | LOC_Os07g47100 | transporter, monovalent cation:proton antiporter-2 family | Overexpression | + | [96] |
| OsVP1 | LOC_Os01g68370 | B3 DNA binding domain containing protein | Overexpression | + | [96] |
| K+ uptake | |||||
| OsHAK1 | LOC_Os04g32920 | Potassium transporter | Mutant and overexpression | + | [97] |
| OsHAK5 | LOC_Os01g70490 | Potassium transporter | Knockdown overexpression | + | [98] |
| OsHAK16 | LOC_Os03g37840 | Potassium transporter | Overexpression | + | [99] |
| OsHAK21 | LOC_Os03g37930 | Potassium transporter | Knockdown | + | [100] |
* + positive regulation; – negative regulation.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ponce, K.S.; Guo, L.; Leng, Y.; Meng, L.; Ye, G. Advances in Sensing, Response and Regulation Mechanism of Salt Tolerance in Rice. Int. J. Mol. Sci. 2021, 22, 2254. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052254
AMA Style
Ponce KS, Guo L, Leng Y, Meng L, Ye G. Advances in Sensing, Response and Regulation Mechanism of Salt Tolerance in Rice. International Journal of Molecular Sciences. 2021; 22(5):2254. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052254
Chicago/Turabian StylePonce, Kimberly S., Longbiao Guo, Yujia Leng, Lijun Meng, and Guoyou Ye. 2021. "Advances in Sensing, Response and Regulation Mechanism of Salt Tolerance in Rice" International Journal of Molecular Sciences 22, no. 5: 2254. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052254
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.