
Kalkitoxin Reduces Osteoclast Formation and Resorption and Protects against Inflammatory Bone Loss

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
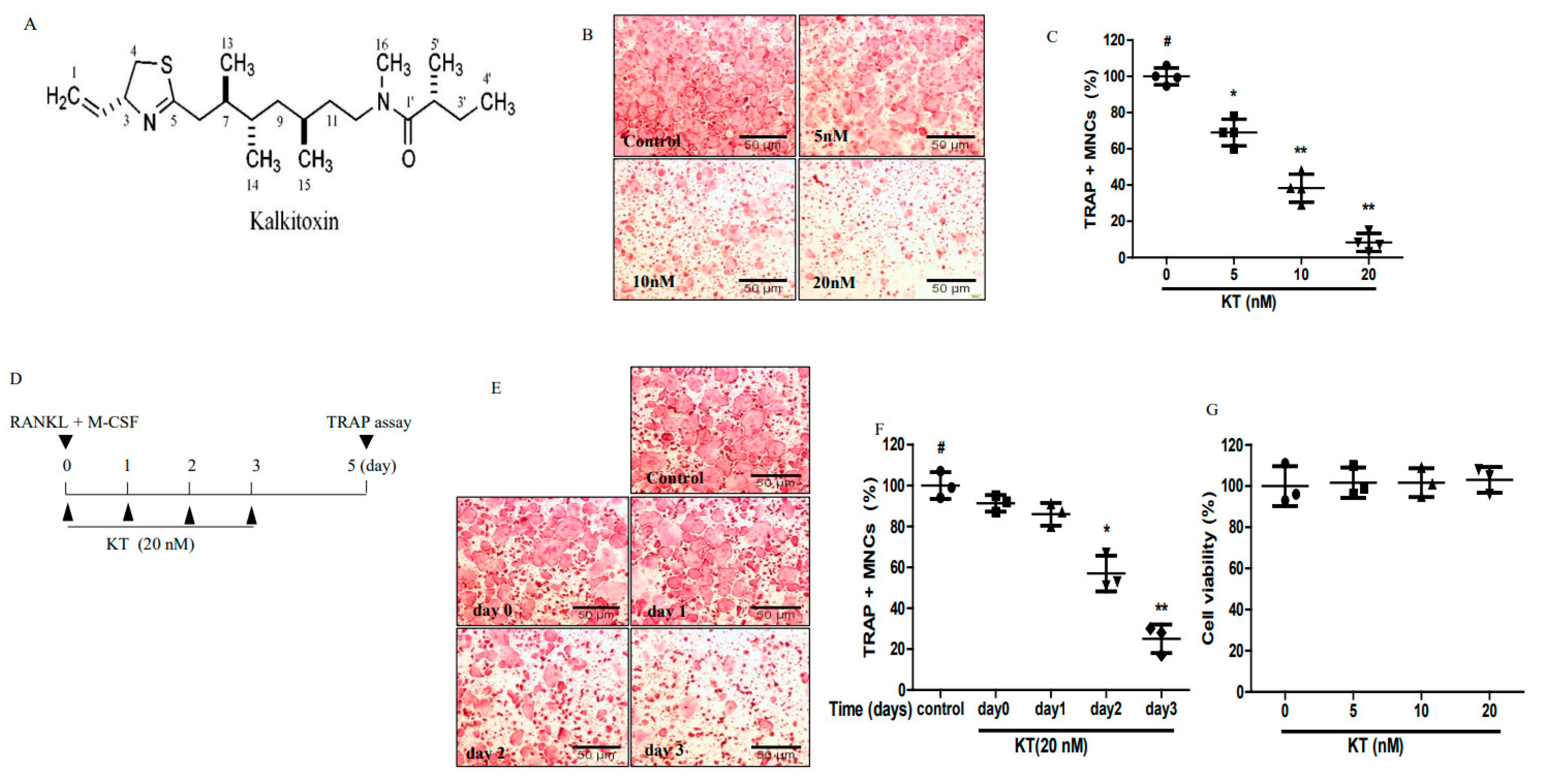
2.1. KT Reduced Osteoclastogenesis in Bone Marrow-Derived Macrophages (BMMs)
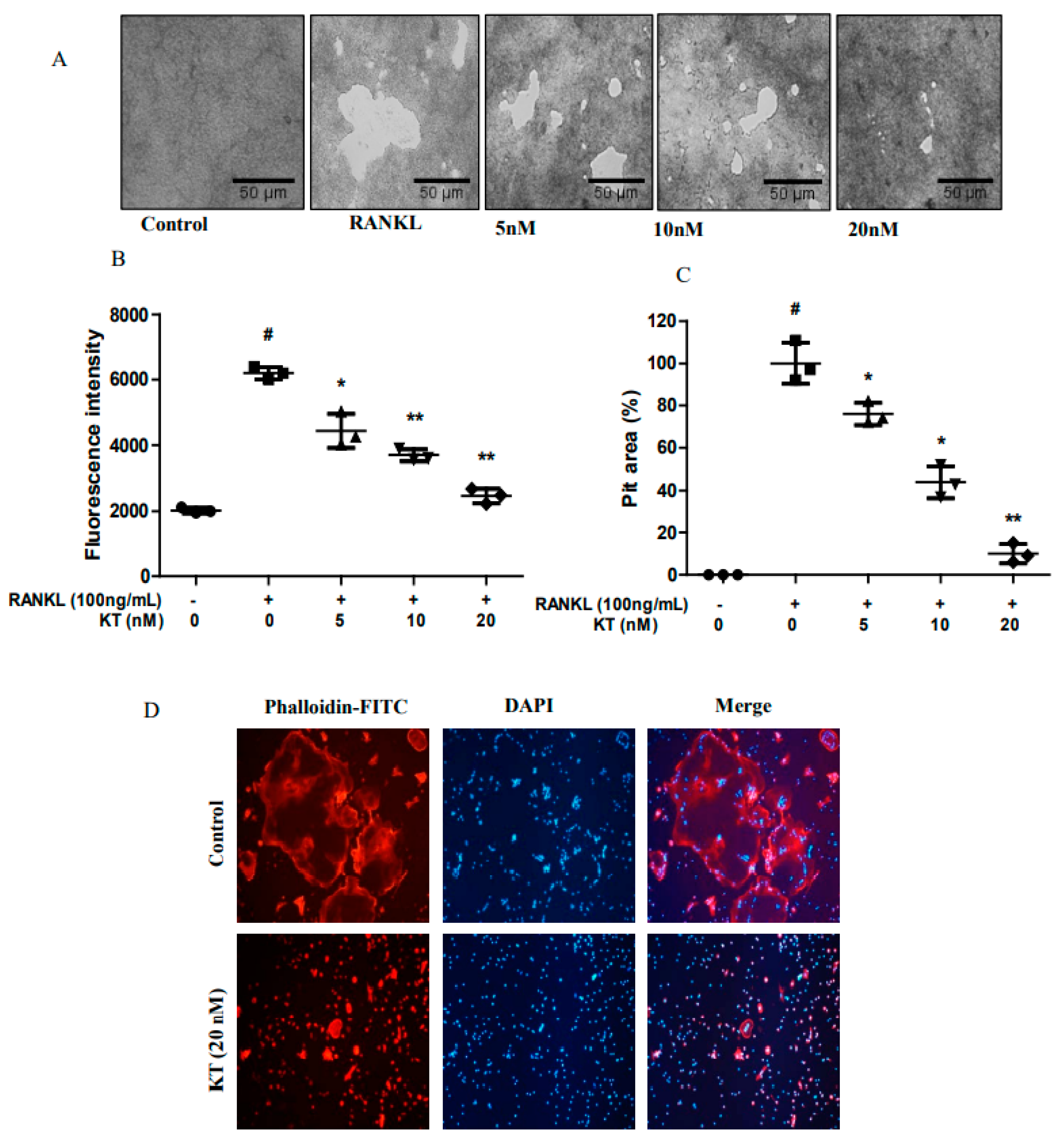
2.2. KT Impairs Bone Resorption and Actin Ring Formation in BMMs
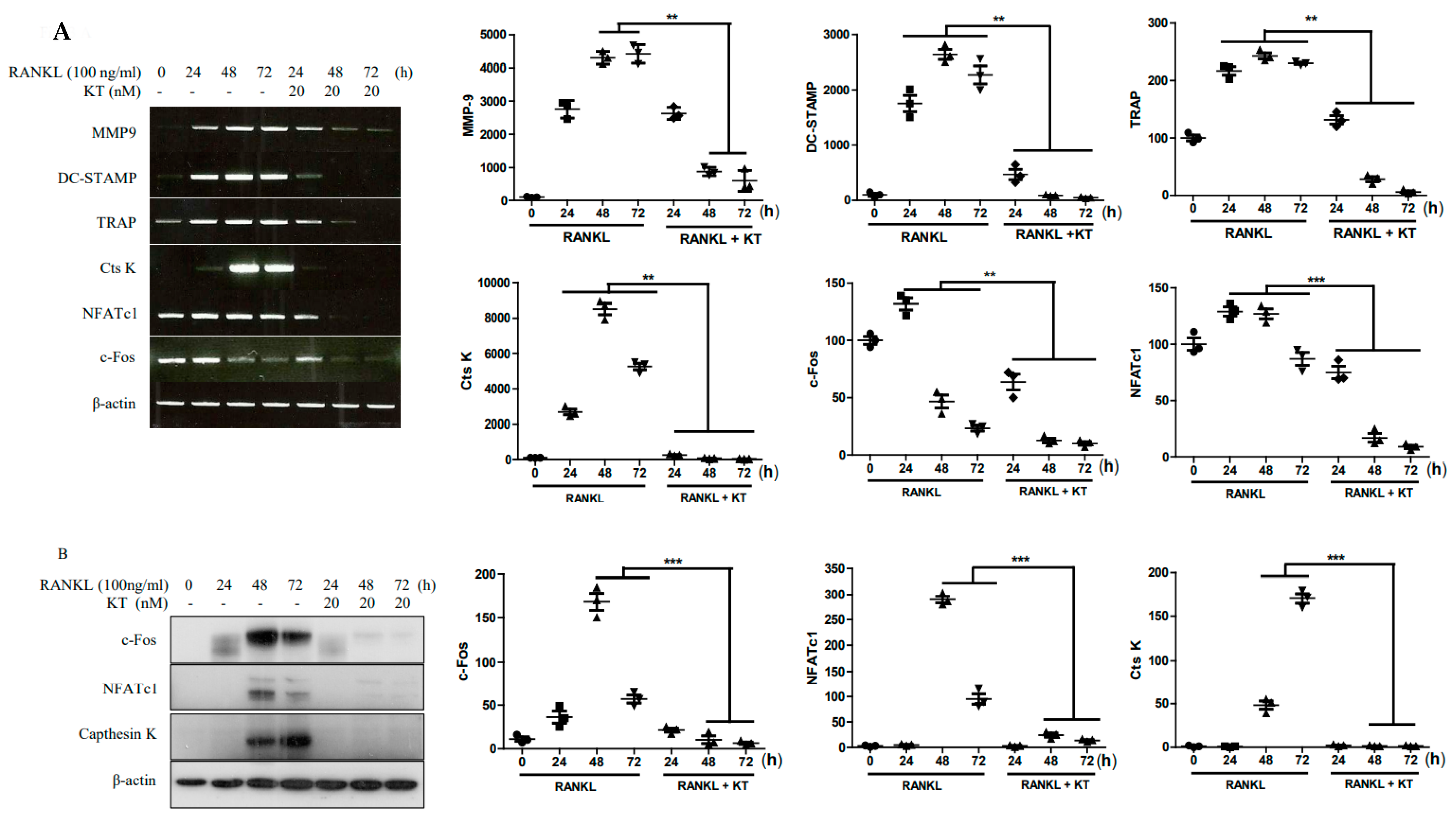
2.3. KT Negatively Regulates RANKL-Induced Gene and Protein Expression Levels
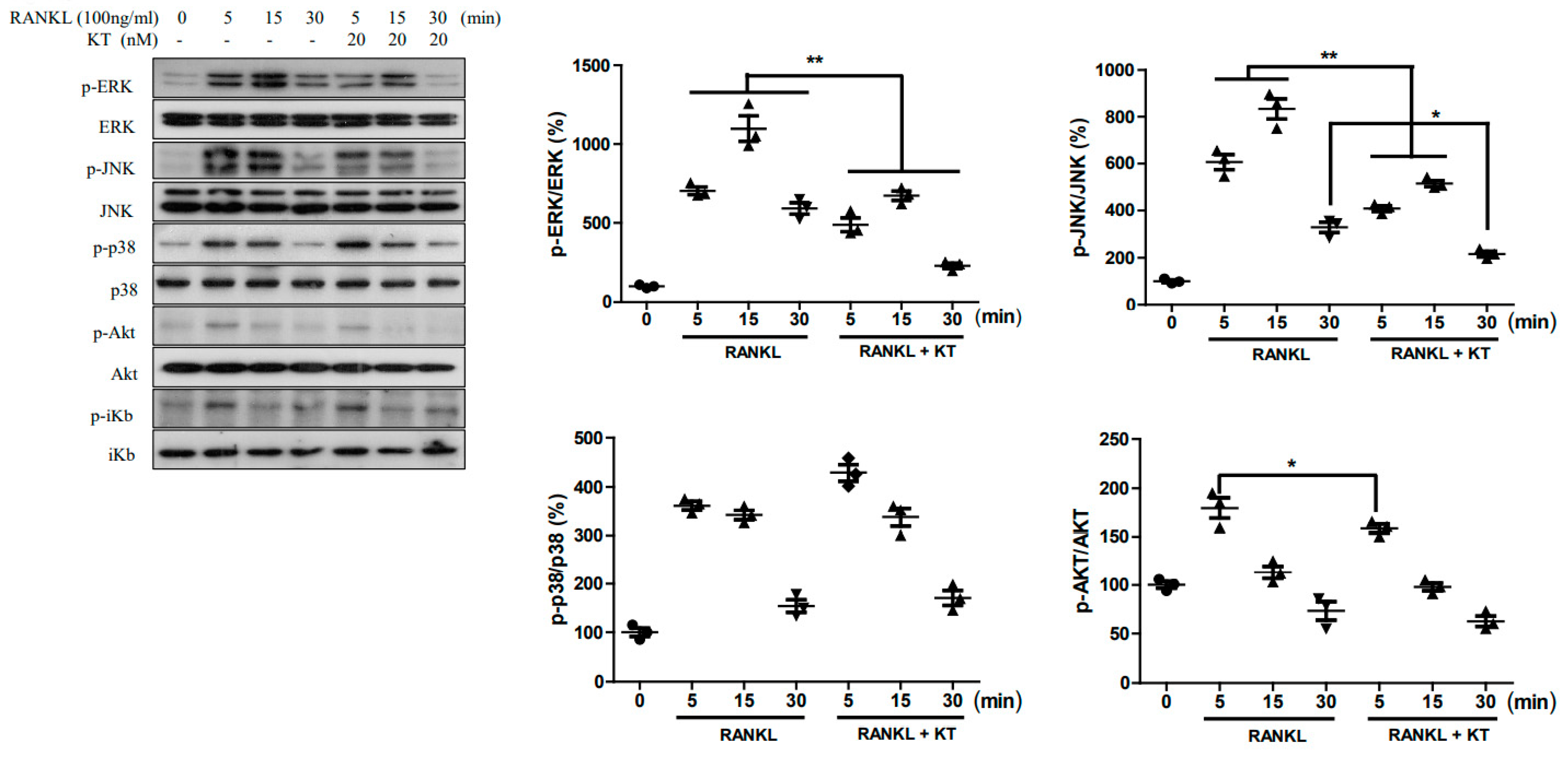
2.4. KT Repressed RANKL-Stimulated MAPK and AKT Pathways in BMMs
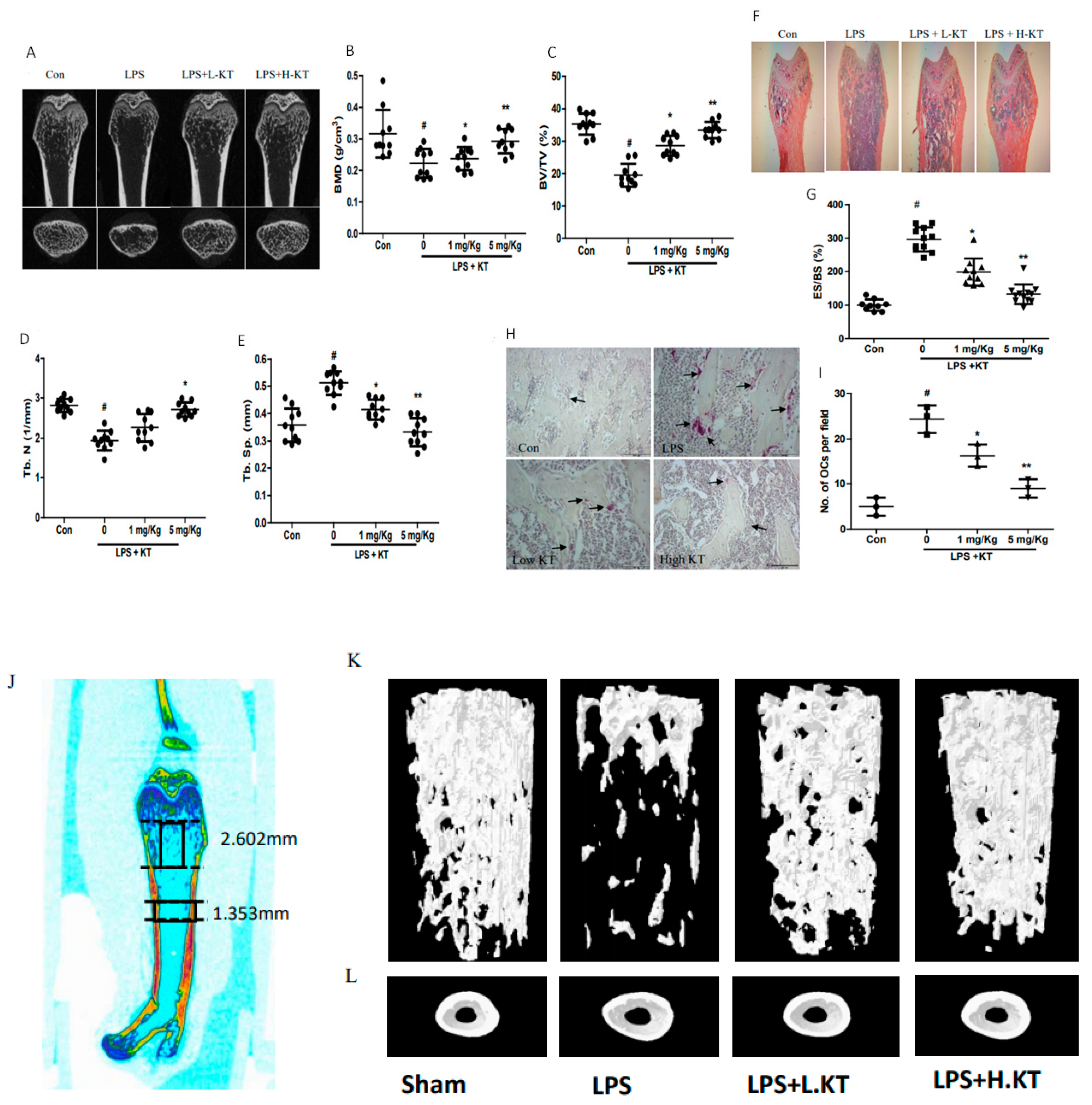
2.5. KT Administration Prevents LPS-Induced Bone Loss in Mice
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Animal Ethics and Bone Marrow Macrophage Cell Culture
4.3. Cell Viability
4.4. Osteoclast Differentiation
4.5. Resorption Pit Area
4.6. Fibrous Actin Ring Formation
4.7. RNA Extraction and Reverse-Transcribed mRNA
4.8. Immunoblot
4.9. In Vivo Experiments
4.10. U-CT and Histopathology
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation, and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef]
- Teitelbaum, S.L.; Ross, F.P. Genetic regulation of osteoclast development and function. Nat. Rev. Genet. 2003, 4, 638–649. [Google Scholar] [CrossRef]
- Zaidi, M. Skeletal remodeling in health and disease. Nat. Med. 2007, 13, 791–801. [Google Scholar] [CrossRef]
- Teitelbaum, S.L. Osteoclasts: What do they do and how do they do it? Am. J. Pathol. 2007, 170, 427–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novack, D.V.; Teitelbaum, S.L. The osteoclast: Friend or foe? Annu. Rev. Pathol. 2008, 3, 457–484. [Google Scholar] [CrossRef] [PubMed]
- Asagiri, M.; Takayanagi, H. The molecular understanding of osteoclast differentiation. Bone 2007, 40, 251–264. [Google Scholar] [CrossRef]
- Teitelbaum, S.L. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef]
- Takayanagi, H.; Kim, S.; Koga, T.; Nishina, H.; Isshiki, M.; Yoshida, H.; Saiura, A.; Isobe, M.; Yokochi, T.; Inoue, J.; et al. induction and activation of the transcription factor NFATc1 (NFAT2) integrate RANKL signaling in terminal differentiation of osteoclasts. Dev. Cell. 2002, 3, 889–901. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, M.; Kogawa, M.; Wada, S.; Takayanagi, H.; Tsujimoto, M.; Katayama, S.; Hisatake, K.; Nogi, Y. Essential role of p38 mitogen-activated protein kinase in cathepsin K gene expression during osteoclastogenesis through association of NFATc1 and PU.1. J. Biol. Chem. 2004, 279, 45969–45979. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Okino, T.; Nogle, L.M.; Marquez, B.L.; Williamson, R.T.; Sitachitta, N.; Berman, F.W.; Murray, T.F.; McGough, K.; Jacobs, R. Structure, synthesis, and biological properties of kalkitoxin, a novel neurotoxin from the marine cyanobacterium Lyngbya majuscula. J. Am. Chem. Soc. 2000, 122, 12041–12042. [Google Scholar] [CrossRef]
- Morgan, J.B.; Liu, Y.; Coothankandaswamy, V.; Mahdi, F.; Jekabsons, M.B.; Gerwick, W.H.; Valeriote, F.A.; Zhou, Y.D.; Nagle, D.G. Kalkitoxin inhibits angiogenesis, disrupts cellular hypoxic signaling, and blocks mitochondrial electron transport in tumor cells. Mar. Drugs 2015, 13, 1552–1568. [Google Scholar] [CrossRef]
- LePage, K.T.; Goeger, D.; Yokokawa, F.; Asano, T.; Shioiri, T.; Gerwick, W.H.; Murray, T.F. The neurotoxic lipopeptide kalkitoxin interacts with voltage-sensitive sodium channels in cerebellar granule neurons. Toxicol. Lett. 2005, 158, 133–139. [Google Scholar] [CrossRef] [PubMed]
- White, J.D.; Xu, Q.; Lee, C.S.; Valeriote, F.A. Total synthesis and biological evaluation of +-kalkitoxin, a cytotoxic metabolite of the cyanobacterium Lyngbya majuscula. Org. Biomol. Chem. 2004, 2, 2092–2102. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.R.; Peters, C.; Saftig, P.; Bromme, D. Cathepsin K activity-dependent regulation of osteoclast actin ring formation and bone resorption. J. Biol. Chem. 2009, 284, 2584–2592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, F.L.; Xu, R.S.; Jiang, D.L.; He, X.L.; Su, Q.; Jin, C.; Li, X. Leonurine hydrochloride inhibits osteoclastogenesis and prevents osteoporosis associated with estrogen deficiency by inhibiting the NF-kappaB and PI3K/Akt signaling pathways. Bone 2015, 75, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Chang, E.J.; Ryu, J.; Lee, Z.H.; Lee, Y.; Kim, H.H. Induction of c-Fos and NFATc1 during RANKL-stimulated osteoclast differentiation is mediated by the p38 signaling pathway. Biochem. Biophys. Res. Commun. 2006, 351, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, A.; Kumagai, M.; Mishima, T.; Ito, J.; Otoki, Y.; Harada, T.; Kato, T.; Yoshida, M.; Suzuki, M.; Yoshida, I.; et al. Toddaculin, Isolated from of Toddalia asiatica (L.) Lam., Inhibited Osteoclastogenesis in RAW 264 Cells and Enhanced Osteoblastogenesis in MC3T3-E1 Cells. PLoS ONE 2015, 1, e0127158. [Google Scholar] [CrossRef]
- Leibbrandt, A.; Penninger, J.M. RANK/RANKL: Regulators of immune responses and bone physiology. Ann. N. Y. Acad. Sci. 2008, 1143, 123–150. [Google Scholar] [CrossRef]
- Gan, K.; Xu, L.; Feng, X.; Zhang, Q.; Wang, F.; Zhang, M.; Tan, W. Celastrol attenuates bone erosion in collagen-Induced arthritis mice and inhibits osteoclast differentiation and function in RANKL-induced RAW264.7. Int. Immunopharmacol. 2015, 24, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Lee, Z.H.; Kim, H.H. Signal transduction by receptor activator of nuclear factor kappa B in osteoclasts. Biochem. Biophys. Res. Commun. 2003, 305, 211–214. [Google Scholar] [CrossRef]
- Li, C.; Yang, Z.; Li, Z.; Ma, Y.; Zhang, L.; Zheng, C.; Qiu, W.; Wu, X.; Wang, X.; Li, H.; et al. Maslinic acid suppresses osteoclastogenesis and prevents ovariectomy-induced bone loss by regulating RANKL-mediated NF-kappaB and MAPK signaling pathways. J. Bone Miner. Res. 2011, 26, 644–656. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.Q.; Pope, R.M. The role of toll-like receptors in rheumatoid arthritis. Curr. Rheumatol. Rep. 2009, 11, 357–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villa, F.A.; Gerwick, L. Marine natural product drug discovery: Leads for treatment of inflammation, cancer, infections, and neurological disorders. Immunopharm. Immunot. 2010, 32, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Sapkota, M.; Kim, S.W.; Soh, Y. Herbacetin inhibits RANKL-mediated osteoclastogenesis in vitro and prevents inflammatory bone loss in vivo. European J. Pharmacol. 2016, 777, 17–25. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Genes (Accession Number) | Primer (Forward, Reverse) | Annealing Tm (°C) | PCR Cycles |
|---|---|---|---|
| TRAP (NM_007388) | 5′-ctgctgggcctacaaatcat-3′ 5′-ggtagtaagggctggggaag-3′ | 54 | 30 |
| MMP9 (NM_013599) | 5′-cgtcgtgatccccacttact-3′ 5′-agagtactgcttgcccagga-3′ | 57.5 | 36 |
| Cathepsin K NM_007802) | 5′-aggcggctatatgaccactg-3′ 5′-ccgagccaagagagcatatc-3′ | 57.5 | 26 |
| c-Fos (NM_010234) | 5′-atgggctctcctgtcaacac-3′ 5′-ggctgccaaaataaactcca-3′ | 57.5 | 30 |
| NFATc1 (NM_198429) DC-STAMP (AY517483) | 5′-gggtcagtgtgaccgaagat-3′ 5′-aggtgggtgaagactgaagg-3′ 5′-ctaaggagaagaaacccttg-3′ 5′-cagcatagaagacaacaatcc-3′ | 55 54 | 35 35 |
| β-actin (NM_007393) | 5′-ttctacaatgagctgcgtgt-3′ 5′-ctcatagctcttctccaggg-3′ | 50 | 26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Yang, M.; Shrestha, S.K.; Kim, H.; Gerwick, W.H.; Soh, Y. Kalkitoxin Reduces Osteoclast Formation and Resorption and Protects against Inflammatory Bone Loss. Int. J. Mol. Sci. 2021, 22, 2303. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052303
Li L, Yang M, Shrestha SK, Kim H, Gerwick WH, Soh Y. Kalkitoxin Reduces Osteoclast Formation and Resorption and Protects against Inflammatory Bone Loss. International Journal of Molecular Sciences. 2021; 22(5):2303. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052303
Chicago/Turabian StyleLi, Liang, Ming Yang, Saroj Kumar Shrestha, Hyoungsu Kim, William H. Gerwick, and Yunjo Soh. 2021. "Kalkitoxin Reduces Osteoclast Formation and Resorption and Protects against Inflammatory Bone Loss" International Journal of Molecular Sciences 22, no. 5: 2303. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052303