
Myricetin as an Antivirulence Compound Interfering with a Morphological Transformation into Coccoid Forms and Potentiating Activity of Antibiotics against Helicobacter pylori

 , ,
, ,  , and
, and
Abstract
:
1. Introduction
2. Results
2.1. Antibacterial Activity of MYR
2.2. Inhibitory Effect of MYR against the Spiral-to-Coccoid Transformation
2.3. Inhibitory Effect of MYR against Biofilm Formation
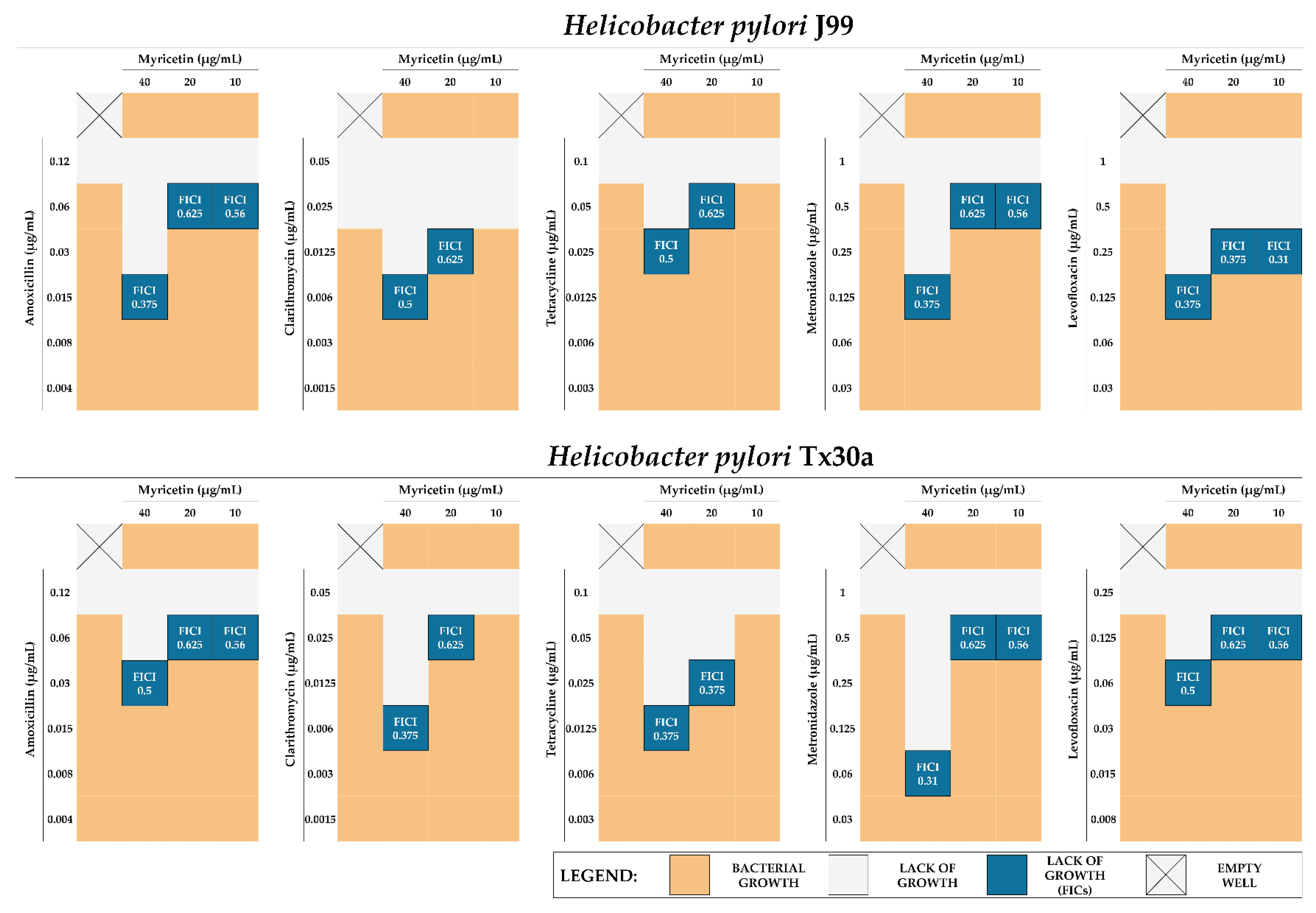
2.4. Synergistic Activity of MYR with Antibiotics
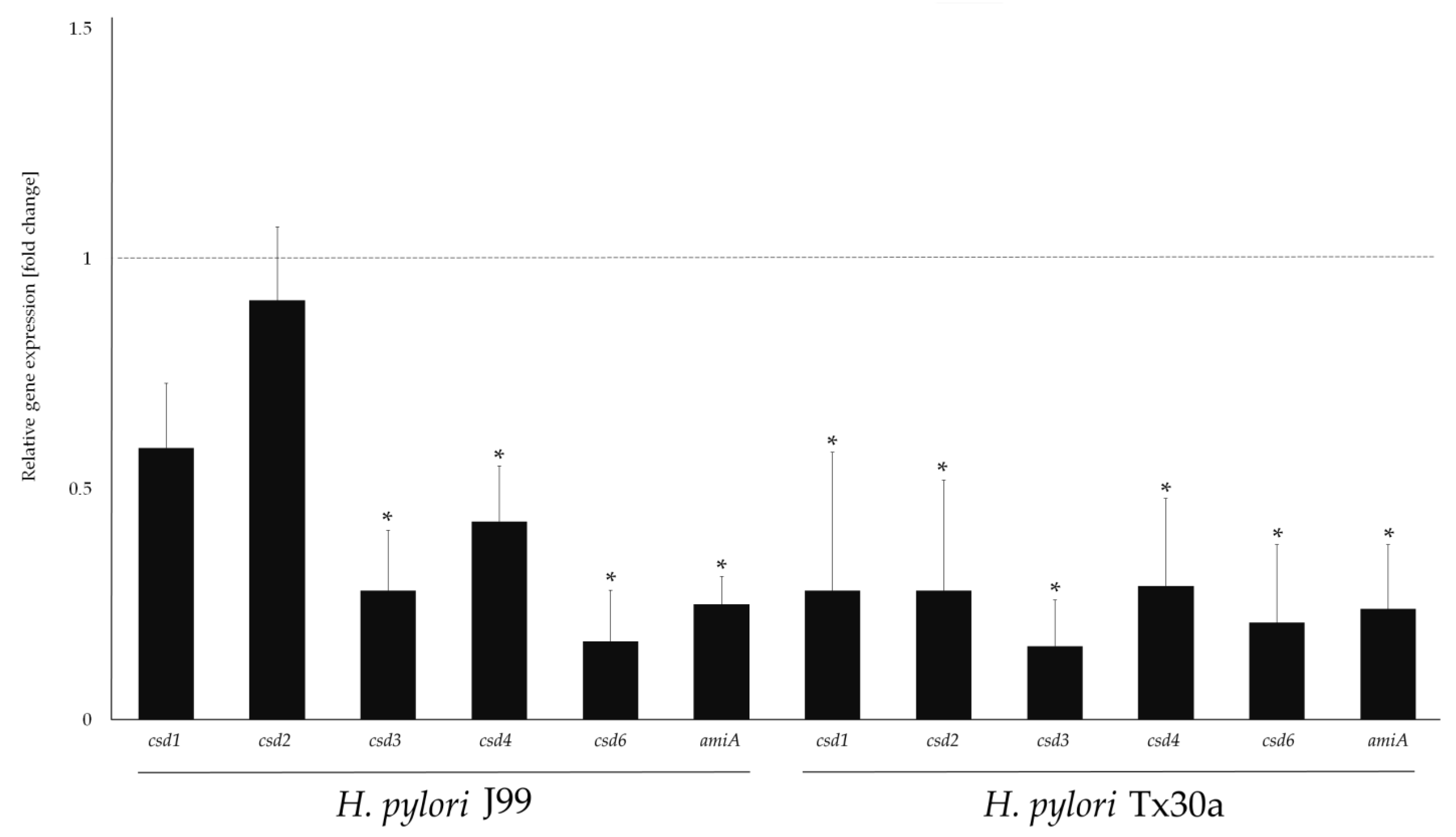
2.5. Modulatory Effect of MYR on Genes Related to Morphogenesis
3. Discussion
4. Materials and Methods
4.1. H. pylori Strains
4.2. Assessment of the Bactericidal Activity of MYR
4.3. Assessment of an Inhibitory Effect of MYR on Morphological Transformation
4.4. Assessment of the Interaction of MYR with Antibiotics
4.5. Assessment of Cell Viability
4.6. Assessment of Biofilm Formation
4.7. Assessment of Expression of Genes Encoding Morphogenesis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, C.H.; Hsieh, Y.H.; Powers, Z.M.; Kao, C.Y. Defeating Antibiotic-Resistant Bacteria: Exploring Alternative Therapies for a Post-Antibiotic Era. Int. J. Mol. Sci. 2020, 21, 1061. [Google Scholar] [CrossRef] [Green Version]
- Tagliabue, A.; Rappuoli, R. Changing Priorities in Vaccinology: Antibiotic Resistance Moving to the Top. Front. Immunol. 2018, 9, 1068. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, Research, and Development of New Antibiotics: The WHO Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Van Giau, V.; An, S.S.A.; Hulme, J. Recent Advances in the Treatment of Pathogenic Infections using Antibiotics and Nano-Drug Delivery Vehicles. Drug Des. Devel. Ther. 2019, 13, 327–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buroni, S.; Chiarelli, L.R. Antivirulence Compounds: A Future Direction to Overcome Antibiotic Resistance? Future Microbiol. 2020, 15, 299–301. [Google Scholar] [CrossRef]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, L.N.; Zimmer, K.R.; Macedo, A.J.; Trentin, D.S. Plant Natural Products Targeting Bacterial Virulence Factors. Chem. Rev. 2016, 116, 9162–9236. [Google Scholar] [CrossRef]
- Taheri, Y.; Suleria, H.A.R.; Martins, N.; Sytar, O.; Beyatli, A.; Yeskaliyeva, B.; Seitimova, G.; Salehi, B.; Semwal, P.; Painuli, S.; et al. Myricetin Bioactive Effects: Moving from Preclinical Evidence to Potential Clinical Applications. BMC Complement. Med. Ther. 2020, 20, 241. [Google Scholar] [CrossRef] [PubMed]
- Semwal, D.K.; Semwal, R.B.; Combrinck, S.; Viljoen, A. Myricetin: A Dietary Molecule with Diverse Biological Activities. Nutrients 2016, 8, 90. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Tan, L.; Wang, M.; Ren, C.; Guo, C.; Yang, B.; Ren, Y.; Cao, Z.; Li, Y.; Pei, J. Myricetin: A Review of the Most Recent Research. Biomed. Pharmacother. 2021, 134, 111017. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, P.; Lv, H.; Deng, X.; Wang, J. A Natural Dietary Flavone Myricetin as an α-Hemolysin Inhibitor for Controlling Staphylococcus aureus Infection. Front. Cell. Infect. Microbiol. 2020, 10, 330. [Google Scholar] [CrossRef]
- Silva, L.N.; Da Hora, G.C.A.; Soares, T.A.; Bojer, M.S.; Ingmer, H.; Macedo, A.J.; Trentin, D.S. Myricetin Protects Galleria mellonella against Staphylococcus aureus Infection and Inhibits Multiple Virulence Factors. Sci. Rep. 2017, 7, 2823. [Google Scholar] [CrossRef]
- Grenier, D.; Chen, H.; Lagha, A.B.; Fournier-Larente, J.; Morin, M.P. Dual Action of Myricetin on Porphyromonas gingivalis and the Inflammatory Response of Host Cells: A Promising Therapeutic Molecule for Periodontal Diseases. PLoS ONE 2015, 10, e0131758. [Google Scholar] [CrossRef]
- Li, G.; Wang, G.; Si, X.; Zhang, X.; Liu, W.; Li, L.; Wang, J. Inhibition of Suilysin Activity and Inflammation by Myricetin Attenuates Streptococcus suis Virulence. Life Sci. 2019, 223, 62–68. [Google Scholar] [CrossRef]
- Lv, Q.; Lv, Y.; Dou, X.; Wassy, S.L.; Jia, G.; Wei, L.; Yu, Q.; Deng, X.; Zhang, C.; Wang, J. Myricetin Inhibits the Type III Secretion System of Salmonella enterica serovar Typhimurium by Downregulating the Salmonella Pathogenic Island I Gene Regulatory Pathway. Microb. Pathog. 2021, 150, 104695. [Google Scholar] [CrossRef]
- Rhee, K.H.; Park, J.S.; Cho, M.J. Helicobacter pylori: Bacterial Strategy for Incipient Stage and Persistent Colonization in Human Gastric Niches. Yonsei Med. J. 2014, 55, 1453–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonaitis, L.; Pellicano, R.; Kupcinskas, L. Helicobacter pylori and Nonmalignant Upper Gastrointestinal Diseases. Helicobacter 2018, 23, e12522. [Google Scholar] [CrossRef] [Green Version]
- Ribaldone, D.G.; Simondi, D.; Petrini, E.; Astegiano, M.; Durazzo, M. Non-Invasive Biomarkers for Gastric Cancer Diagnosis: Ready for Prime Time? Minerva Biotecnol. 2019, 31, 3–10. [Google Scholar] [CrossRef]
- Sgouras, D.N.; Trang, T.T.H.; Yamaoka, Y. Pathogenesis of Helicobacter pylori Infection. Helicobacter 2015, 20, 8–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivera-Severo, D.; Uberti, A.F.; Marques, M.S.; Pinto, M.T.; Gomez-Lazaro, M.; Figueiredo, C.; Leite, M.; Carlini, C.R. A New Role for Helicobacter pylori Urease: Contributions to Angiogenesis. Front. Microbiol. 2017, 8, 1883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansari, S.; Yamaoka, Y. Role of Vacuolating Cytotoxin A in Helicobacter pylori Infection and Its Impact on Gastric Pathogenesis. Expert Rev. Anti. Infect. Ther. 2020, 18, 987–996. [Google Scholar] [CrossRef]
- Ansari, S.; Yamaoka, Y. Helicobacter pylori Virulence Factor Cytotoxin-Associated Gene A (CagA)-Mediated Gastric Pathogenicity. Int. J. Mol. Sci. 2020, 21, 7430. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Soyfoo, D.M.; Wu, Y.; Xu, S. Virulence of Helicobacter pylori Outer Membrane Proteins: An Updated Review. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1821–1830. [Google Scholar] [CrossRef]
- Krzyżek, P.; Grande, R. Transformation of Helicobacter pylori into Coccoid Forms as a Challenge for Research Determining Activity of Antimicrobial Substances. Pathogens 2020, 9, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzyżek, P.; Gościniak, G. Morphology of Helicobacter pylori as a Result of Peptidoglycan and Cytoskeleton Rearrangements. Prz. Gastroenterol. 2018, 13, 182–195. [Google Scholar] [CrossRef] [Green Version]
- Figura, N.; Moretti, E.; Vaglio, L.; Langone, F.; Vernillo, R.; Vindigni, C.; Giordano, N. Factors Modulating the Outcome of Treatment for the Eradication of Helicobacter pylori Infection. New Microbiol. 2012, 35, 335–340. [Google Scholar]
- Kadkhodaei, S.; Siavoshi, F.; Akbari Noghabi, K. Mucoid and Coccoid Helicobacter pylori with Fast Growth and Antibiotic Resistance. Helicobacter 2020, 25, e12678. [Google Scholar] [CrossRef]
- Sisto, F.; Brenciaglia, M.I.; Scaltrito, M.M.; Dubini, F. Helicobacter pylori: ureA, cagA and vacA Expression during Conversion to the Coccoid Form. Int. J. Antimicrob. Agents 2000, 15, 277–282. [Google Scholar] [CrossRef]
- Mizoguchi, H.; Fujioka, T.; Kishi, K.; Nishizono, A.; Kodama, R.; Nasu, M. Diversity in Protein Synthesis and Viability of Helicobacter pylori Coccoid Forms in Response to Various Stimuli. Infect. Immun. 1998, 66, 5555–5560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, C.; Sun, Y.; Wang, N.; Yu, H.; Zhou, Y.; Chen, C.; Jia, J. Changes of Proteome Components of Helicobacter pylori Biofilms Induced by Serum Starvation. Mol. Med. Rep. 2013, 8, 1761–1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzyżek, P.; Biernat, M.M.; Gościniak, G. Intensive Formation of Coccoid Forms as a Feature Strongly Associated with Highly Pathogenic Helicobacter pylori Strains. Folia Microbiol. 2019, 64, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Krzyżek, P.; Grande, R.; Migdał, P.; Paluch, E.; Gościniak, G. Biofilm Formation as a Complex Result of Virulence and Adaptive Responses of Helicobacter pylori. Pathogens 2020, 9, 1062. [Google Scholar] [CrossRef]
- Hwang, J.Y.; Kim, C.; Kwon, Y.H.; Lee, J.E.; Jeon, S.W.; Nam, S.Y.; Seo, A.N.; Han, M.-H.; Park, J.H. Dual Clarithromycin and Metronidazole Resistance is the Main Cause of Failure in Ultimate Helicobacter pylori Eradication. Dig. Dis. 2021. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Krzyżek, P.; Pawełka, D.; Iwańczak, B.; Kempiński, R.; Leśniakowski, K.; Mégraud, F.; Łaczmański, Ł.; Biernat, M.; Gościniak, G. High Primary Antibiotic Resistance of Helicobacter pylori Strains Isolated from Pediatric and Adult Patients in Poland during 2016–2018. Antibiotics 2020, 9, 228. [Google Scholar] [CrossRef] [PubMed]
- Costa, K.; Bacher, G.; Allmaier, G.; Dominguez-Bello, M.G.; Engstrand, L.; Falk, P.; de Pedro, M.A.; García-del Portillo, F. The Morphological Transition of Helicobacter pylori Cells from Spiral to Coccoid is Preceded by a Substantial Modification of the Cell Wall. J. Bacteriol. 1999, 181, 3710–3715. [Google Scholar] [CrossRef] [Green Version]
- Chaput, C.; Ecobichon, C.; Pouradier, N.; Rousselle, J.-C.; Namane, A.; Boneca, I.G. Role of the N-acetylmuramoyl-L-alanyl Amidase, AmiA, of Helicobacter pylori in Peptidoglycan Metabolism, Daughter Cell Separation, and Virulence. Microb. Drug Resist. 2016, 22, 477–486. [Google Scholar] [CrossRef] [Green Version]
- El Mortaji, L.; Tejada-Arranz, A.; Rifflet, A.; Boneca, I.G.; Pehau-Arnaudet, G.; Radicella, J.P.; Marsin, S.; De Reuse, H. A Peptide of a Type I Toxin-Antitoxin System Induces Helicobacter pylori Morphological Transformation from Spiral Shape to Coccoids. Proc. Natl. Acad. Sci. USA 2020, 117, 31398–31409. [Google Scholar] [CrossRef] [PubMed]
- Webb, S.A.R.; Kahler, C.M. Bench-to-Bedside Review: Bacterial Virulence and Subversion of Host Defences. Crit. Care 2008, 12, 234. [Google Scholar] [CrossRef] [Green Version]
- Leitão, J.H. Microbial Virulence Factors. Int. J. Mol. Sci. 2020, 21, 5320. [Google Scholar] [CrossRef]
- Yang, D.C.; Blair, K.M.; Salama, N.R. Staying in Shape: The Impact of Cell Shape on Bacterial Survival in Diverse Environments. Microbiol. Mol. Biol. Rev. 2016, 80, 187–203. [Google Scholar] [CrossRef] [Green Version]
- Van Teeseling, M.C.F.; de Pedro, M.A.; Cava, F. Determinants of Bacterial Morphology: From Fundamentals to Possibilities for Antimicrobial Targeting. Front. Microbiol. 2017, 8, 1264. [Google Scholar] [CrossRef] [Green Version]
- Lopes, L.A.A.; dos Santos Rodrigues, J.B.; Magnani, M.; de Souza, E.L.; de Siqueira-Júnior, J.P. Inhibitory Effects of Flavonoids on Biofilm Formation by Staphylococcus aureus that Overexpresses Efflux Protein Genes. Microb. Pathog. 2017, 107, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.D.; Chin, Y.P.; Lee, M.H. Antimicrobial Activity of Antibiotics in Combination with Natural Flavonoids against Clinical Extended-Spectrum β-lactamase (ESBL)-Producing Klebsiella pneumoniae. Phyther. Res. 2005, 19, 612–617. [Google Scholar] [CrossRef] [PubMed]
- Lechner, D.; Gibbons, S.; Bucar, F. Modulation of Isoniazid Susceptibility by Flavonoids in Mycobacterium. Phytochem. Lett. 2008, 1, 71–75. [Google Scholar] [CrossRef]
- Brown, A.R.; Ettefagh, K.A.; Todd, D.; Cole, P.S.; Egan, J.M.; Foil, D.H.; Graf, T.N.; Schindler, B.D.; Kaatz, G.W.; Cech, N.B. A Mass Spectrometry-Based Assay for Improved Quantitative Measurements of Efflux Pump Inhibition. PLoS ONE 2015, 10, e0124814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitulescu, G.; Nicorescu, I.M.; Olaru, O.T.; Ungurianu, A.; Mihai, D.P.; Zanfirescu, A.; Nitulescu, G.M.; Margina, D. Molecular Docking and Screening Studies of New Natural Sortase A Inhibitors. Int. J. Mol. Sci. 2017, 18, 2217. [Google Scholar] [CrossRef] [Green Version]
- Pinto, H.B.; Brust, F.R.; Macedo, A.J.; Trentin, D.S. The Antivirulence Compound Myricetin Possesses Remarkable Synergistic Effect with Antibacterials upon Multidrug Resistant Staphylococcus aureus. Microb. Pathog. 2020, 149, 104571. [Google Scholar] [CrossRef]
- Obonyo, M.; Zhang, L.; Thamphiwatana, S.; Pornpattananangkul, D.; Fu, V.; Zhang, L. Antibacterial Activities of Liposomal Linolenic Acids against Antibiotic-Resistant Helicobacter pylori. Mol. Pharm. 2012, 9, 2677–2685. [Google Scholar] [CrossRef] [Green Version]
- Faghri, J.; Poursina, F.; Moghim, S.; Zarkesh Esfahani, H.; Nasr Esfahani, B.; Fazeli, H.; Mirzaei, N.; Jamshidian, A.; Ghasemian Safaei, H. Morphological and Bactericidal Effects of Different Antibiotics on Helicobacter pylori. Jundishapur J. Microbiol. 2014, 7, e8704. [Google Scholar] [CrossRef] [Green Version]
- Krzyżek, P.; Paluch, E.; Gościniak, G. Synergistic Therapies as a Promising Option for the Treatment of Antibiotic-Resistant Helicobacter pylori. Antibiotics 2020, 9, 658. [Google Scholar] [CrossRef]
- Griep, M.A.; Blood, S.; Larson, M.A.; Koepsell, S.A.; Hinrichs, S.H. Myricetin Inhibits Escherichia coli DnaB Helicase but Not Primase. Bioorg. Med. Chem. 2007, 15, 7203–7208. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.H.; Huang, C.Y. Characterization of Flavonol Inhibition of DnaB Helicase: Real-Time Monitoring, Structural Modeling, and Proposed Mechanism. J. Biomed. Biotechnol. 2012, 2012, 735368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinozuka, K.; Kikuchi, Y.; Nishino, C.; Mori, A.; Tawata, S. Inhibitory Effect of Flavonoids on DNA-Dependent DNA and RNA Polymerases. Experientia 1988, 44, 882–885. [Google Scholar] [CrossRef]
- Gottesman, S. Trouble is Coming: Signaling Pathways that Regulate General Stress Responses in Bacteria. J. Biol. Chem. 2019, 294, 11685–11700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive Review of Antimicrobial Activities of Plant Flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef] [Green Version]
- Chaput, C.; Ecobichon, C.; Cayet, N.; Girardin, S.E.; Werts, C.; Guadagnini, S.; Prévost, M.-C.; Mengin-Lecreulx, D.; Labigne, A.; Boneca, I.G. Role of AmiA in the Morphological Transition of Helicobacter pylori and in Immune Escape. PLoS Pathog. 2006, 2, e97. [Google Scholar] [CrossRef] [Green Version]
- Frirdich, E.; Biboy, J.; Pryjma, M.; Lee, J.; Huynh, S.; Parker, C.T.; Girardin, S.E.; Vollmer, W.; Gaynor, E.C. The Campylobacter jejuni Helical to Coccoid Transition Involves Changes to Peptidoglycan and the Ability to Elicit an Immune Response. Mol. Microbiol. 2019, 112, 280–301. [Google Scholar] [CrossRef] [Green Version]
- Frirdich, E.; Biboy, J.; Adams, C.; Lee, J.; Ellermeier, J.; Gielda, L.D.; DiRita, V.J.; Girardin, S.E.; Vollmer, W.; Gaynor, E.C. Peptidoglycan-Modifying Enzyme Pgp1 Is Required for Helical Cell Shape and Pathogenicity Traits in Campylobacter jejuni. PLoS Pathog. 2012, 8, e1002602. [Google Scholar] [CrossRef]
- Yao, Y.; Lin, G.; Xie, Y.; Ma, P.; Li, G.; Meng, Q.; Wu, T. Preformulation Studies of Myricetin: A Natural Antioxidant Flavonoid. Pharmazie 2014, 69, 19–26. [Google Scholar] [PubMed]
- Xiang, D.; Wang, C.G.; Wang, W.Q.; Shi, C.Y.; Xiong, W.; Wang, M.D.; Fang, J.G. Gastrointestinal Stability of Dihydromyricetin, Myricetin, and Myricitrin: An In Vitro Investigation. Int. J. Food Sci. Nutr. 2017, 68, 704–711. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Chen, Y.F.; Tang, Y.; Qian, J.Q. Method for Enhancing Bioavailability of Myricetin Based on Self-Assembly of Casein–Myricetin Nanomicelles. IET Nanobiotechnol. 2020, 14, 239–244. [Google Scholar] [CrossRef]
- Yao, Y.; Xie, Y.; Hong, C.; Li, G.; Shen, H.; Ji, G. Development of a Myricetin/Hydroxypropyl-β-Cyclodextrin Inclusion Complex: Preparation, Characterization, and Evaluation. Carbohydr. Polym. 2014, 110, 329–337. [Google Scholar] [CrossRef]
- Guo, R.X.; Fu, X.; Chen, J.; Zhou, L.; Chen, G. Preparation and Characterization of Microemulsions of Myricetin for Improving Its Antiproliferative and Antioxidative Activities and Oral Bioavailability. J. Agric. Food Chem. 2016, 64, 6286–6294. [Google Scholar] [CrossRef]
- Qian, J.; Meng, H.; Xin, L.; Xia, M.; Shen, H.; Li, G.; Xie, Y. Self-Nanoemulsifying Drug Delivery Systems of Myricetin: Formulation Development, Characterization, and In Vitro and In Vivo Evaluation. Colloids Surf. B Biointerfaces 2017, 160, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Junka, A.; Bartoszewicz, M.; Dziadas, M.; Szymczyk, P.; Dydak, K.; Żywicka, A.; Owczarek, A.; Bil-Lula, I.; Czajkowska, J.; Fijałkowski, K. Application of Bacterial Cellulose Experimental Dressings Saturated with Gentamycin for Management of Bone Biofilm In Vitro and Ex Vivo. J. Biomed. Mater. Res. Part B: Appl. Biomater. 2020, 108, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Ciecholewska-Juśko, D.; Żywicka, A.; Junka, A.; Drozd, R.; Sobolewski, P.; Migdał, P.; Kowalska, U.; Toporkiewicz, M.; Fijałkowski, K. Superabsorbent Crosslinked Bacterial Cellulose Biomaterials for Chronic Wound Dressings. Carbohydr. Polym. 2021, 253, 117247. [Google Scholar] [CrossRef]
- Krzyżek, P.; Gościniak, G.; Fijałkowski, K.; Migdał, P.; Dziadas, M.; Owczarek, A.; Czajkowska, J.; Aniołek, O.; Junka, A. Potential of Bacterial Cellulose Chemisorbed with Anti-Metabolites, 3-Bromopyruvate or Sertraline, to Fight against Helicobacter pylori Lawn Biofilm. Int. J. Mol. Sci. 2020, 21, 9507. [Google Scholar] [CrossRef]
- Krzyżek, P.; Franiczek, R.; Krzyżanowska, B.; Łaczmański, Ł.; Migdał, P.; Gościniak, G. In Vitro Activity of 3-Bromopyruvate, an Anti-Cancer Compound, Against Antibiotic-Susceptible and Antibiotic-Resistant Helicobacter pylori Strains. Cancers 2019, 11, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzyżek, P.; Franiczek, R.; Krzyżanowska, B.; Łaczmański, Ł.; Migdał, P.; Gościniak, G. In Vitro Activity of Sertraline, an Antidepressant, Against Antibiotic-Susceptible and Antibiotic-Resistant Helicobacter pylori Strains. Pathogens 2019, 8, 228. [Google Scholar] [CrossRef] [Green Version]
- Hathroubi, S.; Zerebinski, J.; Ottemann, K.M. Helicobacter pylori Biofilm Involves a Multigene Stress-Biased Response, Including a Structural Role for Flagella. MBio 2018, 9, e01973-18. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, R.M.; Silva, H.; Oliveira, R.; Almeida, C.; Azevedo, N.F.; Vieira, M.J. Morphological Transition of Helicobacter pylori Adapted to Water. Future Microbiol. 2017, 12, 1167–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H. Pylori Strains | Collection Number | Myricetin Activity [µg/mL] | |
|---|---|---|---|
| MIC | MBC | ||
| J99 | ATCC 700824 | 160 ± 23.1 | 320 ± 53.3 |
| Tx30a | ATCC 51932 | 160 ± 26.7 | 320 ± 46.2 |
| Gene | Primer Sequence (5’ to 3’) | Annealing Temperature | |
|---|---|---|---|
| csd1 | Forward | TCGCATACACAGGGGTGTTA | 54 °C |
| Reverse | TGCGCCTTATCCCTAATGAC | ||
| csd2 | Forward | CCTTTCTTTGGTGGGTTTGA | 54 °C |
| Reverse | GCTCTTTATTGTGGGGCAAA | ||
| csd3 | Forward | CGCTCATTCAAGCCCTTATC | 57 °C |
| Reverse | GCTAAAAGGGGGTCATTGGT | ||
| csd4 | Forward | TTAAACCCACCAGGCTCATC | 54 °C |
| Reverse | GGCTTGTGTTCTTGGGTGTT | ||
| csd6 | Forward | GCAGAAATTAGAGCGCTTGG | 51 °C |
| Reverse | GCCCTTGGTGTTCAATTCAT | ||
| amiA | Forward | ATACGGTTTGCTTTGGATGC | 54 °C |
| Reverse | GTCCGCAAAAATTACCCTGA | ||
| ureA | Forward | TTTCACGCTAACGGCTTTTT | 54 °C |
| Reverse | AACCGGATGATGTGATGGAT | ||
| glmM | Forward | CAACCGCTTGAGAAGAAAGG | 54 °C |
| Reverse | CCAACCAATTAAGCCAGGAA | ||
| gyrB | Forward | CGTCGCTTTGGATCATTTTT | 55 °C |
| Reverse | AATGGCGTGCCACTTTTAAC | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krzyżek, P.; Migdał, P.; Paluch, E.; Karwańska, M.; Wieliczko, A.; Gościniak, G. Myricetin as an Antivirulence Compound Interfering with a Morphological Transformation into Coccoid Forms and Potentiating Activity of Antibiotics against Helicobacter pylori. Int. J. Mol. Sci. 2021, 22, 2695. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052695
Krzyżek P, Migdał P, Paluch E, Karwańska M, Wieliczko A, Gościniak G. Myricetin as an Antivirulence Compound Interfering with a Morphological Transformation into Coccoid Forms and Potentiating Activity of Antibiotics against Helicobacter pylori. International Journal of Molecular Sciences. 2021; 22(5):2695. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052695
Chicago/Turabian StyleKrzyżek, Paweł, Paweł Migdał, Emil Paluch, Magdalena Karwańska, Alina Wieliczko, and Grażyna Gościniak. 2021. "Myricetin as an Antivirulence Compound Interfering with a Morphological Transformation into Coccoid Forms and Potentiating Activity of Antibiotics against Helicobacter pylori" International Journal of Molecular Sciences 22, no. 5: 2695. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052695