
Investigation of the Membrane Fluidity Regulation of Fatty Acid Intracellular Distribution by Fluorescence Lifetime Imaging of Novel Polarity Sensitive Fluorescent Derivatives

 ,
,  , ,
, ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
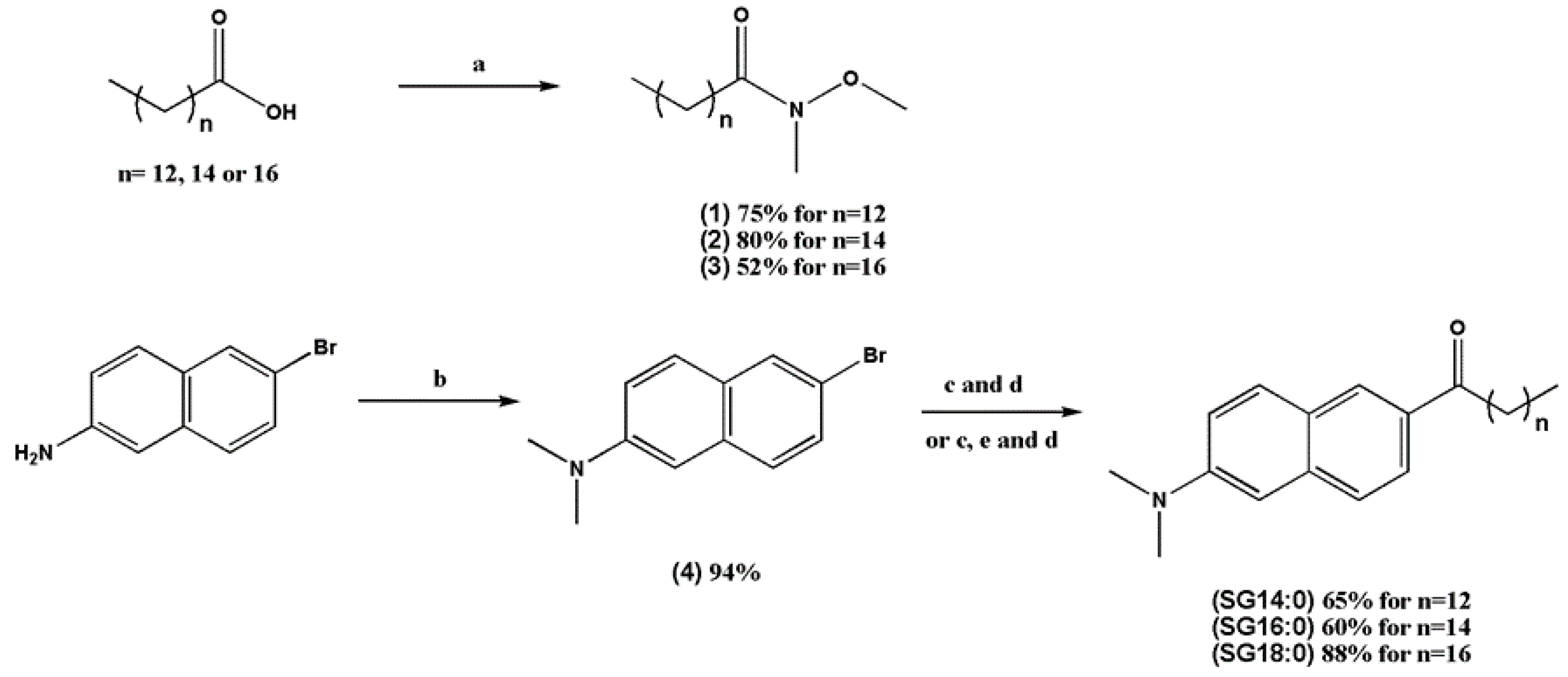
2.1. Synthesis of the Laurdan Analogs with Elongated Fatty Acid Chain
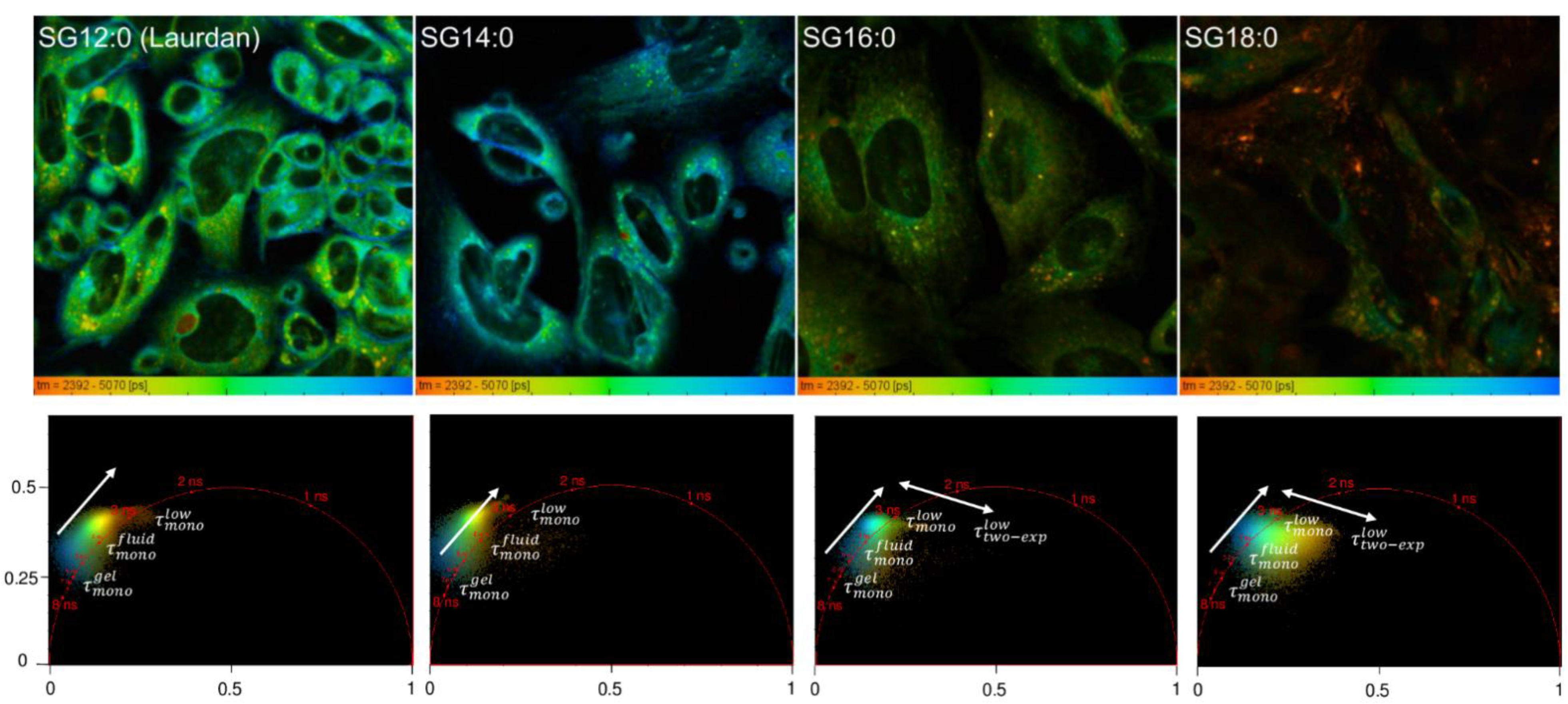
2.2. FLIM Analysis of Fluorescence Analogs: Phase Behavior
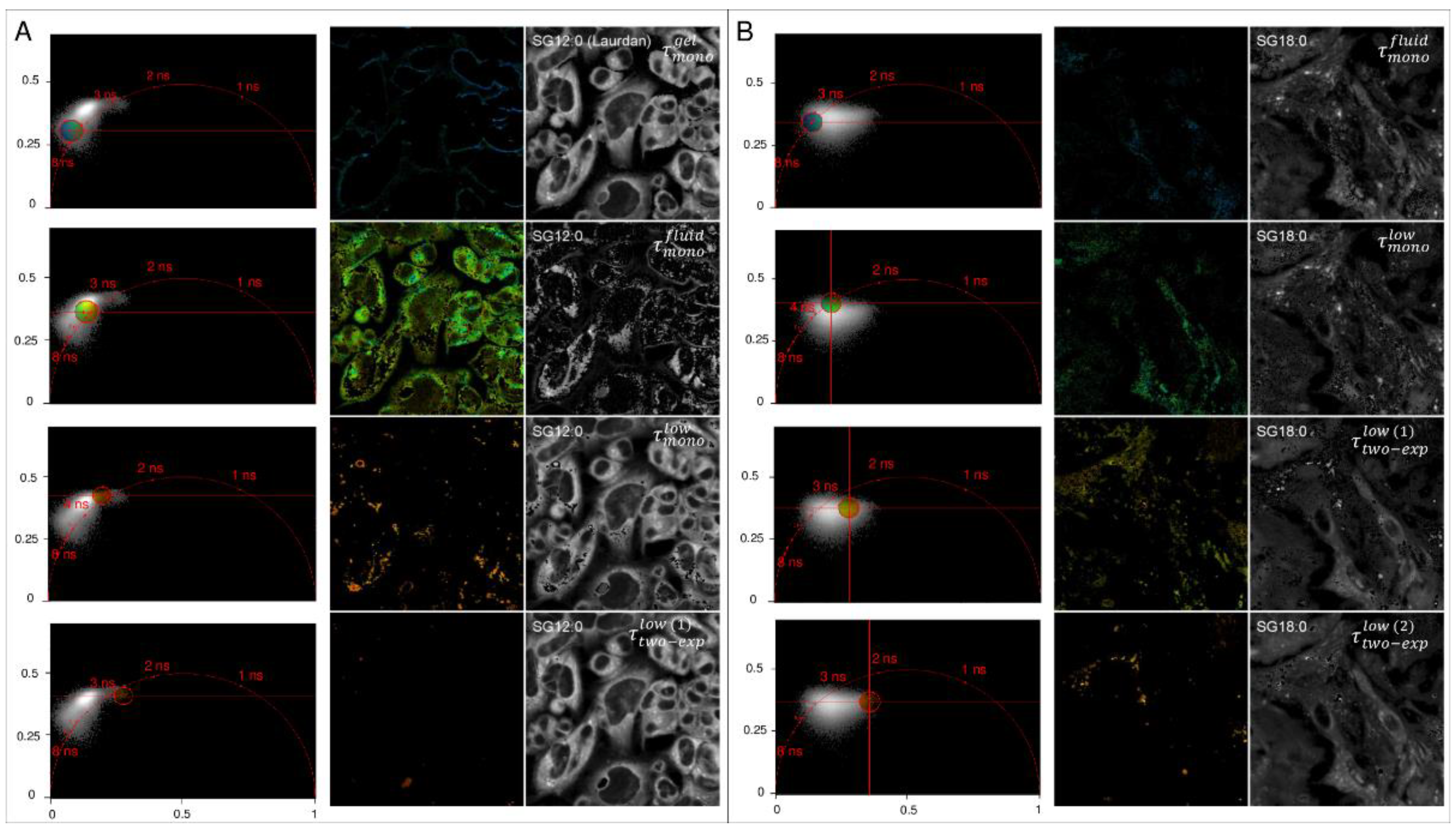
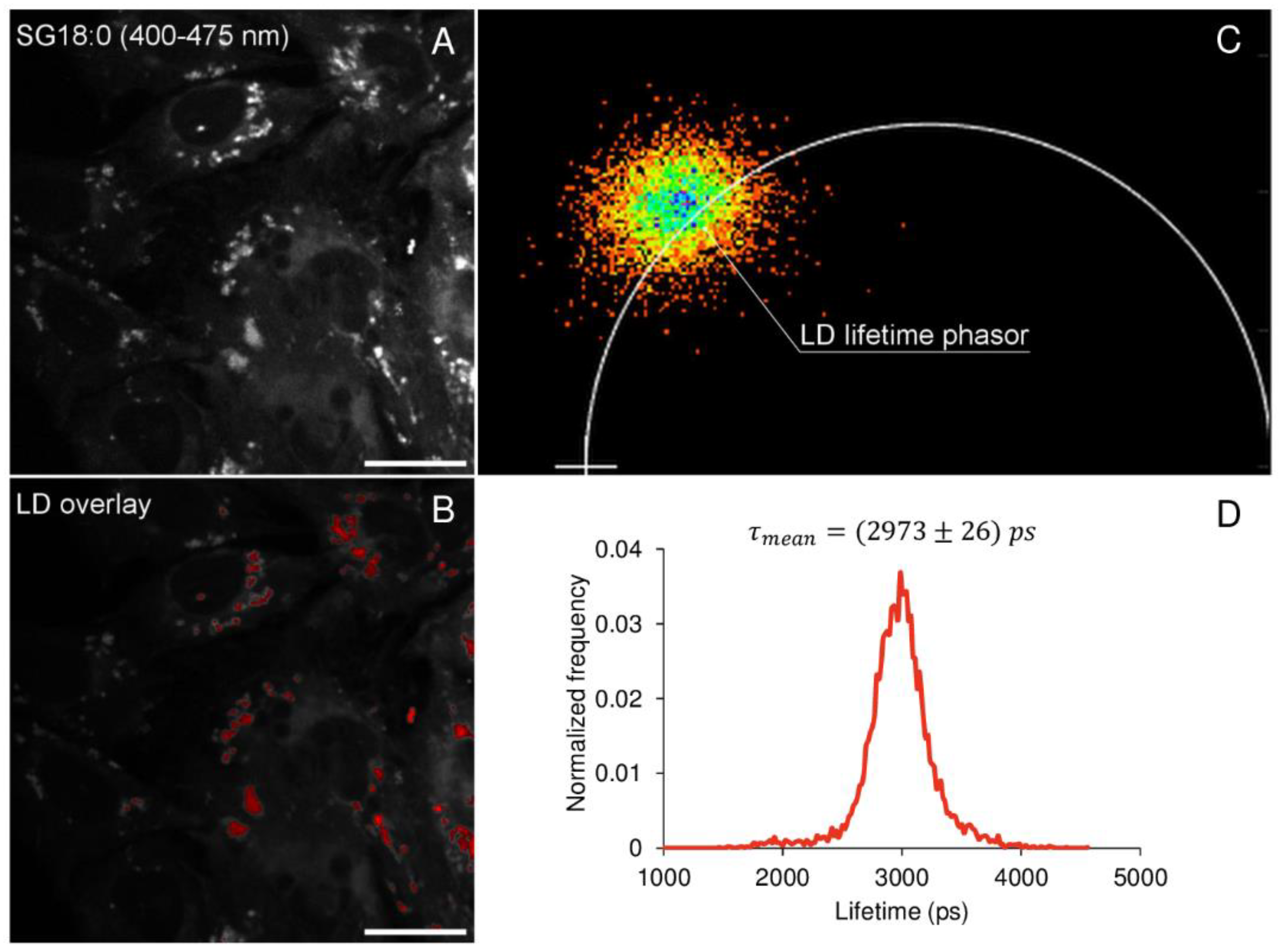
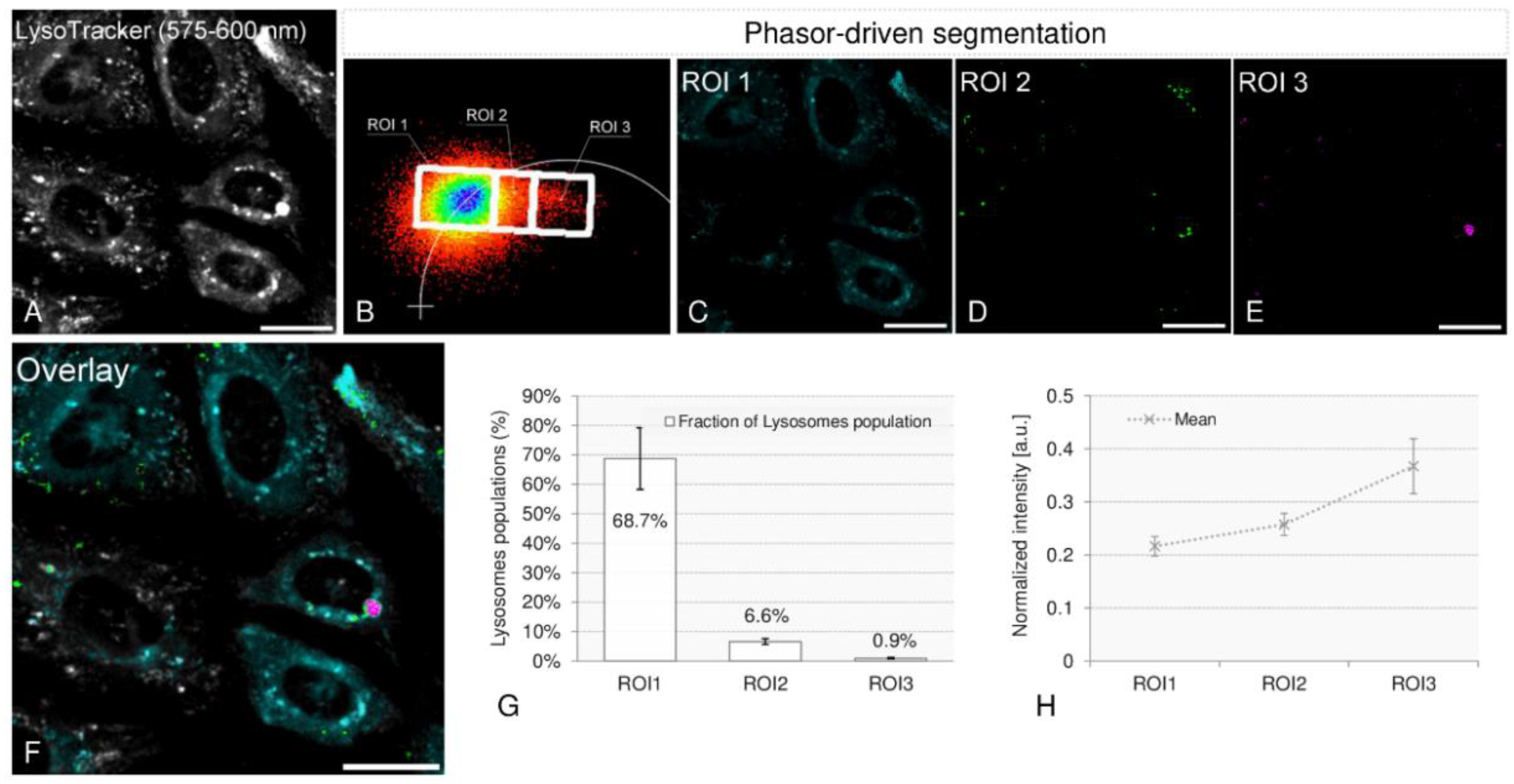
2.3. Spatial Localization of Membrane Phases through Phasor-Driven Segmentation
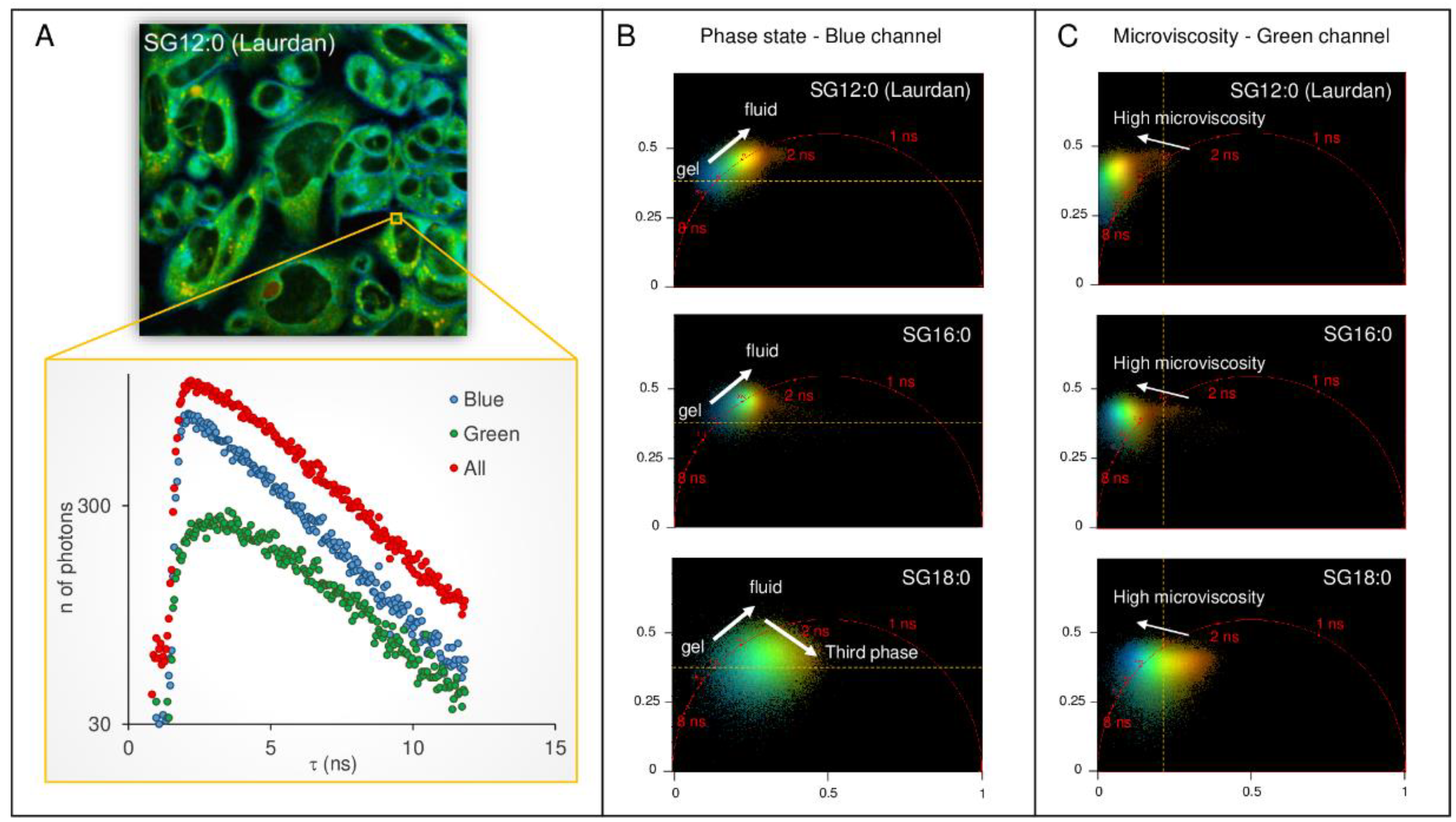
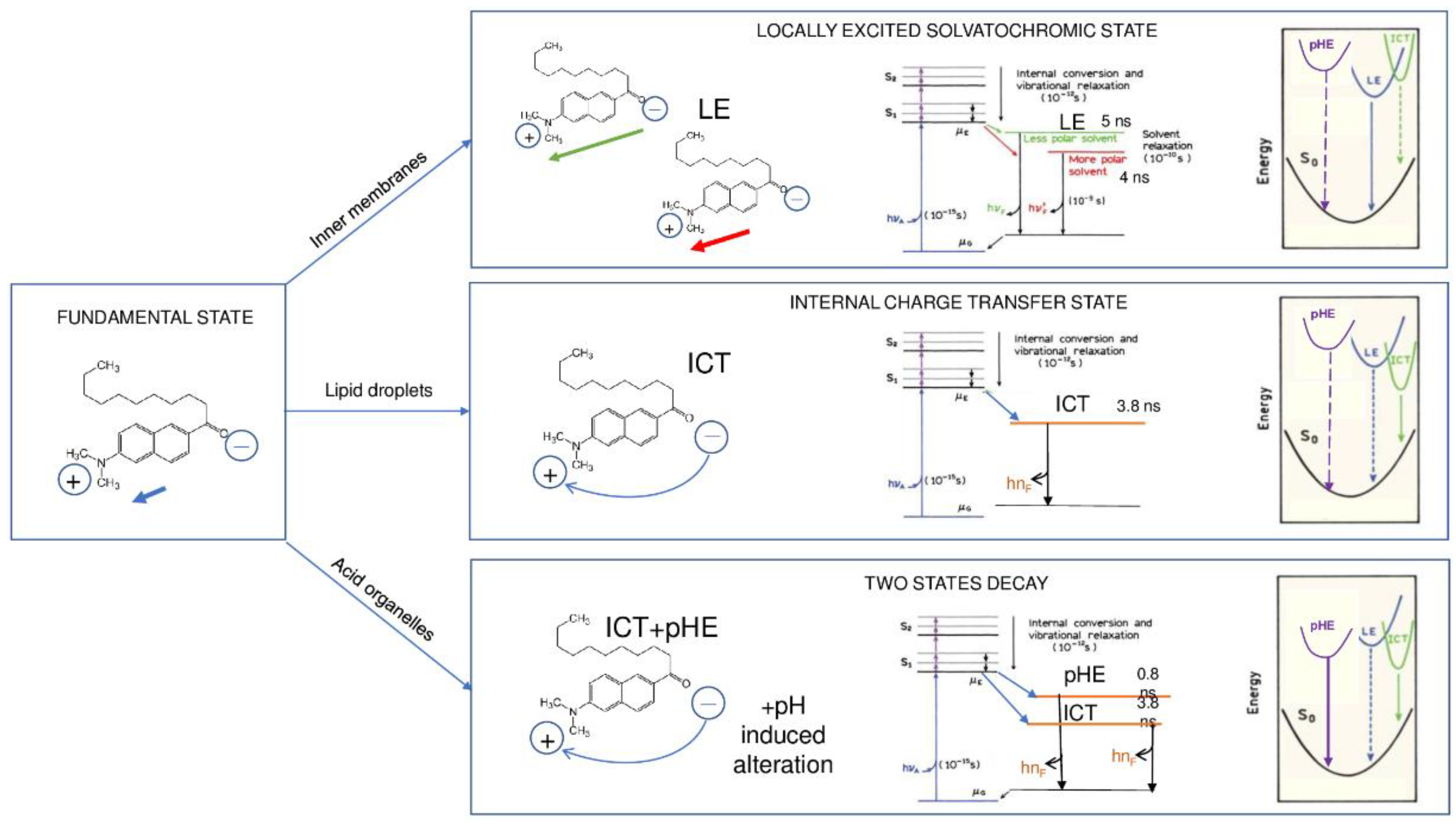
2.4. FLIM Analysis of Fluorescence Derivatives: Solvent Relaxation and Microviscosity
2.5. Colocalization Experiments
3. Discussion
4. Materials and Methods
4.1. Materials for the Synthesis of Laurdan Derivatives
4.2. Synthesis of Laurdan Analogs
4.3. Cell Labeling and Fluorescence Lifetime Imaging Microscopy Acquisitions
4.4. Phasor Analysis
4.5. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rustan, A.C.; Drevon, C.A. Fatty Acids: Structures and Properties. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd.: Chichester, UK, 2005. [Google Scholar]
- Singer, S.J.; Nicolson, G.L. The fluid mosaic model of the structure of cell membranes. Science 1972, 175, 720–731. [Google Scholar] [CrossRef]
- Simons, K.; Ikonen, E. Functional rafts in cell membranes. Nature 1997, 387, 569–572. [Google Scholar] [CrossRef]
- Falholt, K.; Dohn, K.; Lund, B.; Falholt, W. Enzyme pattern in hypoxic skeletal muscle. J. Mol. Cell. Cardiol. 1974, 6, 349–359. [Google Scholar] [CrossRef]
- Maulucci, G.; Cohen, O.; Daniel, B.; Sansone, A.; Petropoulou, P.I.; Filou, S.; Spyridonidis, A.; Pani, G.; De Spirito, M.; Chatgilialoglu, C.; et al. Fatty acid-related modulations of membrane fluidity in cells: Detection and implications. Free Radic. Res. 2016, 50, S40–S50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchetti, G.; Di Giacinto, F.; Pitocco, D.; Rizzi, A.; Rizzo, G.E.; De Leva, F.; Flex, A.; di Stasio, E.; Ciasca, G.; De Spirito, M.; et al. Red blood cells membrane micropolarity as a novel diagnostic indicator of type 1 and type 2 diabetes. Anal. Chim. Acta X 2019, 3, 100030. [Google Scholar] [CrossRef]
- Bianchetti, G.; Viti, L.; Scupola, A.; Di Leo, M.; Tartaglione, L.; Flex, A.; De Spirito, M.; Pitocco, D.; Maulucci, G. Erythrocyte membrane fluidity as a marker of diabetic retinopathy in type 1 diabetes mellitus. Eur. J. Clin. Investig. 2020, 1–6. [Google Scholar] [CrossRef]
- Sanchez, S.A.; Tricerri, M.A.; Gunther, G.; Gratton, E. Laurdan Generalized Polarization: From cuvette to microscope. Mod. Res. Educ. Top. Microsc. 2007, 2, 1007–1014. [Google Scholar]
- Maulucci, G.; Troiani, D.; Eramo, S.L.M.; Paciello, F.; Podda, M.V.; Paludetti, G.; Papi, M.; Maiorana, A.; Palmieri, V.; De Spirito, M.; et al. Time evolution of noise induced oxidation in outer hair cells: Role of NAD(P)H and plasma membrane fluidity. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 2192–2202. [Google Scholar] [CrossRef]
- Angelucci, C.; Maulucci, G.; Lama, G.; Proietti, G.; Colabianchi, A.; Papi, M.; Maiorana, A.; De Spirito, M.; Micera, A.; Balzamino, O.B.; et al. Epithelial-stromal interactions in human breast cancer: Effects on adhesion, plasma membrane fluidity and migration speed and directness. PLoS ONE 2012, 7, e50804. [Google Scholar] [CrossRef]
- Balogh, G.; Maulucci, G.; Gombos, I.; Horváth, I.; Török, Z.; Péter, M.; Fodor, E.; Páli, T.; Benko, S.; Parasassi, T.; et al. Heat stress causes spatially-distinct membrane re-modelling in K562 leukemia cells. PLoS ONE 2011, 6, e21182. [Google Scholar] [CrossRef] [Green Version]
- Scanferlato, R.; Bortolotti, M.; Sansone, A.; Chatgilialoglu, C.; Polito, L.; De Spirito, M.; Maulucci, G.; Bolognesi, A.; Ferreri, C. Hexadecenoic Fatty Acid Positional Isomers and De Novo PUFA Synthesis in Colon Cancer Cells. Int. J. Mol. Sci. 2019, 20, 832. [Google Scholar] [CrossRef] [Green Version]
- Di Giacinto, F.; De Spirito, M.; Maulucci, G. Low-Intensity Ultrasound Induces Thermodynamic Phase Separation of Cell Membranes through a Nucleation–Condensation Process. Ultrasound Med. Biol. 2019, 45, 1143–1150. [Google Scholar] [CrossRef]
- Golfetto, O.; Hinde, E.; Gratton, E. Laurdan fluorescence lifetime discriminates cholesterol content from changes in fluidity in living cell membranes. Biophys. J. 2013, 104, 1238–1247. [Google Scholar] [CrossRef] [Green Version]
- Bagatolli, L.A.; Gratton, E. Two photon fluorescence microscopy of coexisting lipid domains in giant unilamellar vesicles of binary phospholipid mixtures. Biophys. J. 2000, 78, 290–305. [Google Scholar] [CrossRef] [Green Version]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy; Springer Science & Business Media: Berlin, Germany, 2006; ISBN 9780387312781. [Google Scholar]
- Zanni, E.; Maulucci, G.; Pomata, D.; Buiarelli, F.; Krasnowska, E.K.; Parasassi, T.; De Spirito, M.; Heipieper, H.J.; Uccelletti, D. ER stress induced by the OCH1 mutation triggers changes in lipid homeostasis in Kluyveromyces lactis. Res. Microbiol. 2015, 166. [Google Scholar] [CrossRef] [PubMed]
- Cordelli, E.; Maulucci, G.; De Spirito, M.; Rizzi, A.; Pitocco, D.; Soda, P. A decision support system for type 1 diabetes mellitus diagnostics based on dual channel analysis of red blood cell membrane fluidity. Comput. Methods Programs Biomed. 2018, 162. [Google Scholar] [CrossRef] [PubMed]
- Maulucci, G.; Cordelli, E.; Rizzi, A.; De Leva, F.; Papi, M.; Ciasca, G.; Samengo, D.; Pani, G.; Pitocco, D.; Soda, P.; et al. Phase separation of the plasma membrane in human red blood cells as a potential tool for diagnosis and progression monitoring of type 1 diabetes mellitus. PLoS ONE 2017, 12, e0184109. [Google Scholar] [CrossRef]
- Bianchetti, G.; De Spirito, M.; Maulucci, G.; De Spirito, M.; Maulucci, G.; Maulucci, G. Unsupervised clustering of multiparametric fluorescent images extends the spectrum of detectable cell membrane phases with sub-micrometric resolution. Biomed. Opt. Express 2020, 11, 5728. [Google Scholar] [CrossRef] [PubMed]
- Gaus, K.; Zech, T.; Harder, T. Visualizing membrane microdomains by Laurdan 2-photon microscopy. Mol. Membr. Biol. 2006, 23, 41–48. [Google Scholar] [CrossRef]
- Ma, Y.; Benda, A.; Kwiatek, J.; Owen, D.M.; Gaus, K. Time-Resolved Laurdan Fluorescence Reveals Insights into Membrane Viscosity and Hydration Levels. Biophys. J. 2018, 115, 1498–1508. [Google Scholar] [CrossRef] [Green Version]
- Jurkiewicz, P.; Sýkora, J.; Olzyńska, A.; Humpolíčková, J.; Hof, M. Solvent relaxation in phospholipid bilayers: Principles and recent applications. J. Fluoresc. 2005, 15, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Bianchetti, G.; Di Giacinto, F.; De Spirito, M.; Maulucci, G. Machine-learning assisted confocal imaging of intracellular sites of triglycerides and cholesteryl esters formation and storage. Anal. Chim. Acta 2020, 1121, 57–66. [Google Scholar] [CrossRef]
- Maulucci, G.; Di Giacinto, F.; De Angelis, C.; Cohen, O.; Daniel, B.; Ferreri, C.; De Spirito, M.; Sasson, S. Real time quantitative analysis of lipid storage and lipolysis pathways by confocal spectral imaging of intracellular micropolarity. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 783–793. [Google Scholar] [CrossRef]
- Di Giacinto, F.; De Angelis, C.; De Spirito, M.; Maulucci, G. Quantitative imaging of membrane micropolarity in living cells and tissues by spectral phasors analysis. MethodsX 2018, 5, 1399–1412. [Google Scholar] [CrossRef]
- Goldman, S.D.B.; Funk, R.S.; Rajewski, R.A.; Krise, J.P. Mechanisms of amine accumulation in, and egress from, lysosomes. Bioanalysis 2009, 1, 1445–1459. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, L.F.; Pino, J.A.; Soto-Arriaza, M.A.; Cuevas, F.J.; Sánchez, S.; Sotomayor, C.P. Differential dynamic and structural behavior of lipid-cholesterol domains in model membranes. PLoS ONE 2012, 7, 40254. [Google Scholar] [CrossRef]
- Róg, T.; Pasenkiewicz-Gierula, M. Cholesterol Effects on the Phosphatidylcholine Bilayer Nonpolar Region: A Molecular Simulation Study. Biophys. J. 2001, 81, 2190–2202. [Google Scholar] [CrossRef] [Green Version]
- Soubias, O.; Jolibois, F.; Milon, A.; Réat, V. High-resolution 13C NMR of sterols in model membrane. Comptes Rendus Chim. 2006, 9, 393–400. [Google Scholar] [CrossRef]
- Róg, T.; Pasenkiewicz-Gierula, M.; Vattulainen, I.; Karttunen, M. What Happens if Cholesterol Is Made Smoother: Importance of Methyl Substituents in Cholesterol Ring Structure on Phosphatidylcholine–Sterol Interaction. Biophys. J. 2007, 92, 3346–3357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vequi-Suplicy, C.C.; Coutinho, K.; Lamy, M.T. New insights on the fluorescent emission spectra of Prodan and Laurdan. J. Fluoresc. 2015, 25, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Sezgin, E.; Schneider, F.; Zilles, V.; Urbančič, I.; Garcia, E.; Waithe, D.; Klymchenko, A.S.; Eggeling, C. Polarity-Sensitive Probes for Superresolution Stimulated Emission Depletion Microscopy. Biophys. J. 2017, 113, 1321–1330. [Google Scholar] [CrossRef] [Green Version]
- Bridges, C.R.; Guo, C.; Yan, H.; Miltenburg, M.B.; Li, P.; Li, Y.; Seferos, D.S. Conjugated Polymers with Switchable Carrier Polarity. Macromolecules 2015, 48, 5587–5595. [Google Scholar] [CrossRef]
- García, P.; Martín-Pastor, M.; De Lera, Á.R.; Álvarez, R. Determination of the geometry of acetoxyendiynes and acetoxyenynes by NMR heteronuclear 13C-1H scalar couplings and 13C NMR chemical shifts. Structural assignment of the oxylipin natural products peyssonenynes A and B. Magn. Reson. Chem. 2010, 48, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Rosini, G.; Andreotti, D.G.; D’Ambrosio, P.; Marotta, E.; Tinarelli, A.; Righi, P. Synthesis of 1-oxo-1-(3-pyridazinyl) derivatives—Potent inhibitors of Fatty Acid Amide Hydrolase (FAAH): An improved and optimized procedure. Synthesis 2007, 2007, 3051–3055. [Google Scholar] [CrossRef]
- Giumanini, A.G.; Chiavari, G.; Musiani, M.M.; Rossi, P. N-permethylation of primary and secondary aromatic amines. Synth. 1980, 1980, 743–746. [Google Scholar] [CrossRef]
- Digman, M.A.; Caiolfa, V.R.; Zamai, M.; Gratton, E. The phasor approach to fluorescence lifetime imaging analysis. Biophys. J. 2008, 94, L14-6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stringari, C.; Nourse, J.L.; Flanagan, L.A.; Gratton, E. Phasor Fluorescence Lifetime Microscopy of Free and Protein-Bound NADH Reveals Neural Stem Cell Differentiation Potential. PLoS ONE 2012, 7, e48014. [Google Scholar] [CrossRef] [Green Version]
- R Studio. Available online: https://rstudio.com/ (accessed on 15 March 2021).







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bianchetti, G.; Azoulay-Ginsburg, S.; Keshet-Levy, N.Y.; Malka, A.; Zilber, S.; Korshin, E.E.; Sasson, S.; De Spirito, M.; Gruzman, A.; Maulucci, G. Investigation of the Membrane Fluidity Regulation of Fatty Acid Intracellular Distribution by Fluorescence Lifetime Imaging of Novel Polarity Sensitive Fluorescent Derivatives. Int. J. Mol. Sci. 2021, 22, 3106. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063106
Bianchetti G, Azoulay-Ginsburg S, Keshet-Levy NY, Malka A, Zilber S, Korshin EE, Sasson S, De Spirito M, Gruzman A, Maulucci G. Investigation of the Membrane Fluidity Regulation of Fatty Acid Intracellular Distribution by Fluorescence Lifetime Imaging of Novel Polarity Sensitive Fluorescent Derivatives. International Journal of Molecular Sciences. 2021; 22(6):3106. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063106
Chicago/Turabian StyleBianchetti, Giada, Salome Azoulay-Ginsburg, Nimrod Yosef Keshet-Levy, Aviv Malka, Sofia Zilber, Edward E. Korshin, Shlomo Sasson, Marco De Spirito, Arie Gruzman, and Giuseppe Maulucci. 2021. "Investigation of the Membrane Fluidity Regulation of Fatty Acid Intracellular Distribution by Fluorescence Lifetime Imaging of Novel Polarity Sensitive Fluorescent Derivatives" International Journal of Molecular Sciences 22, no. 6: 3106. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063106