
Effect of 8-Day Fasting on Leukocytes Expression of Genes and Proteins Involved in Iron Metabolism in Healthy Men

 , ,
, ,
Abstract
:1. Introduction
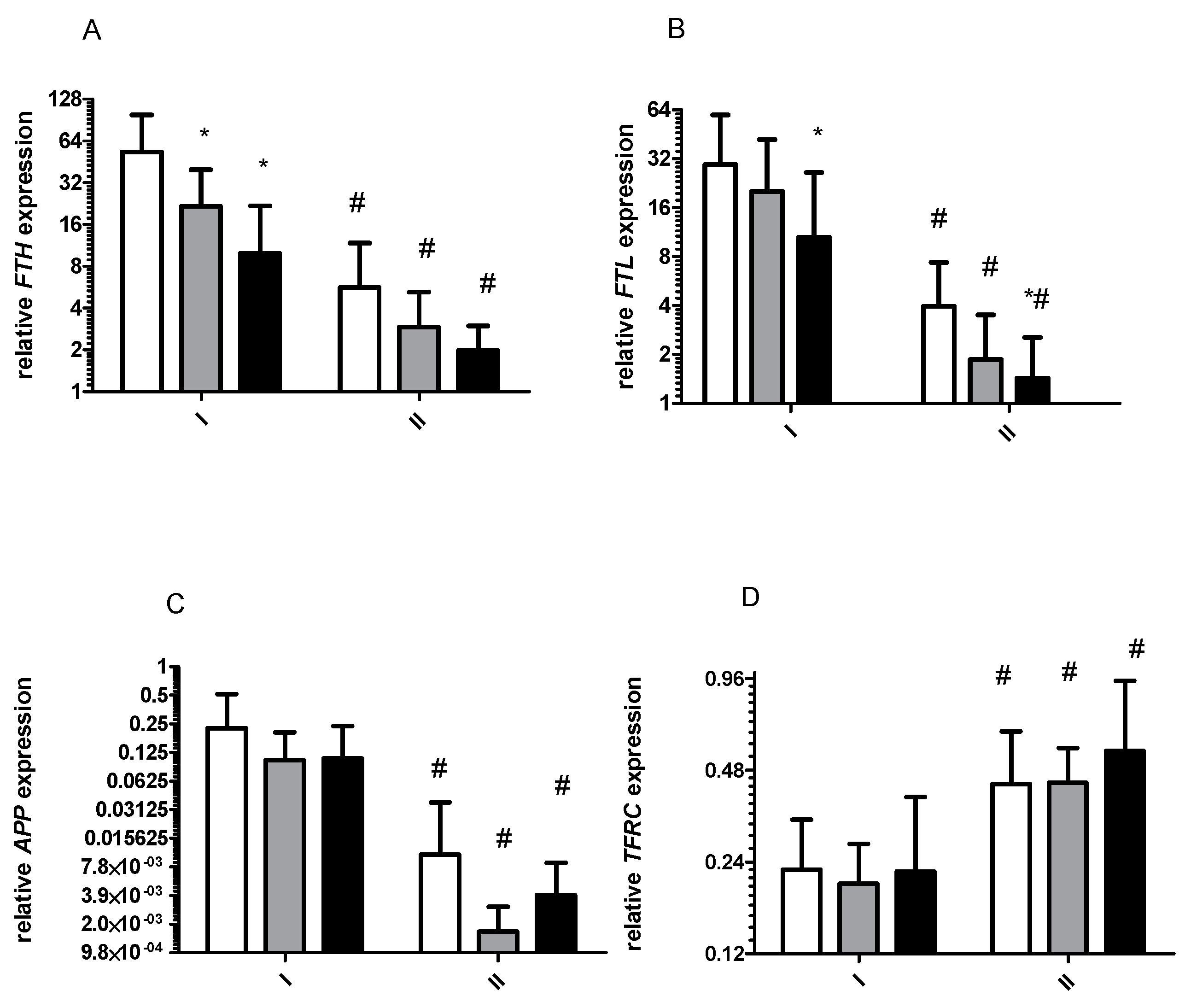
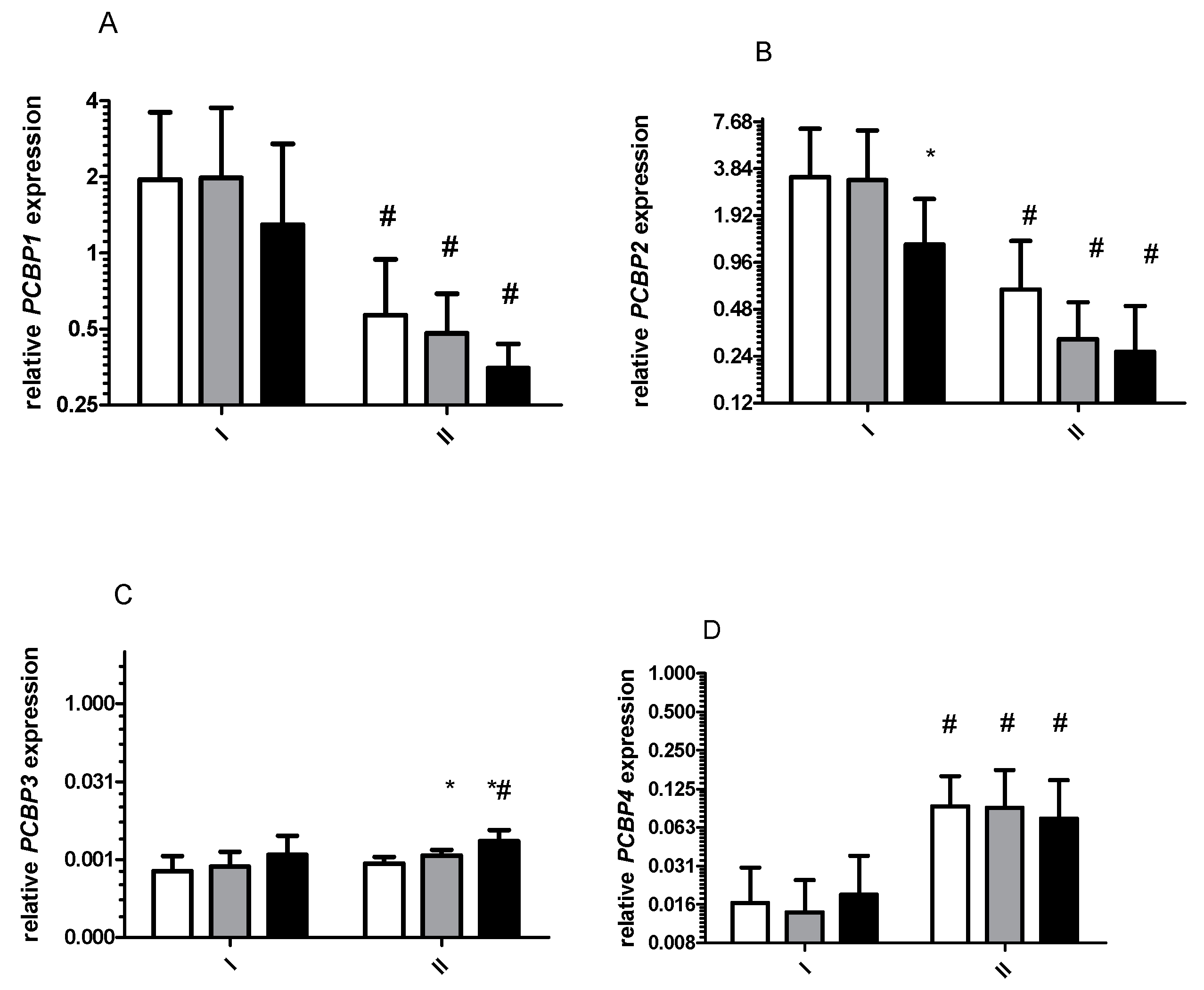
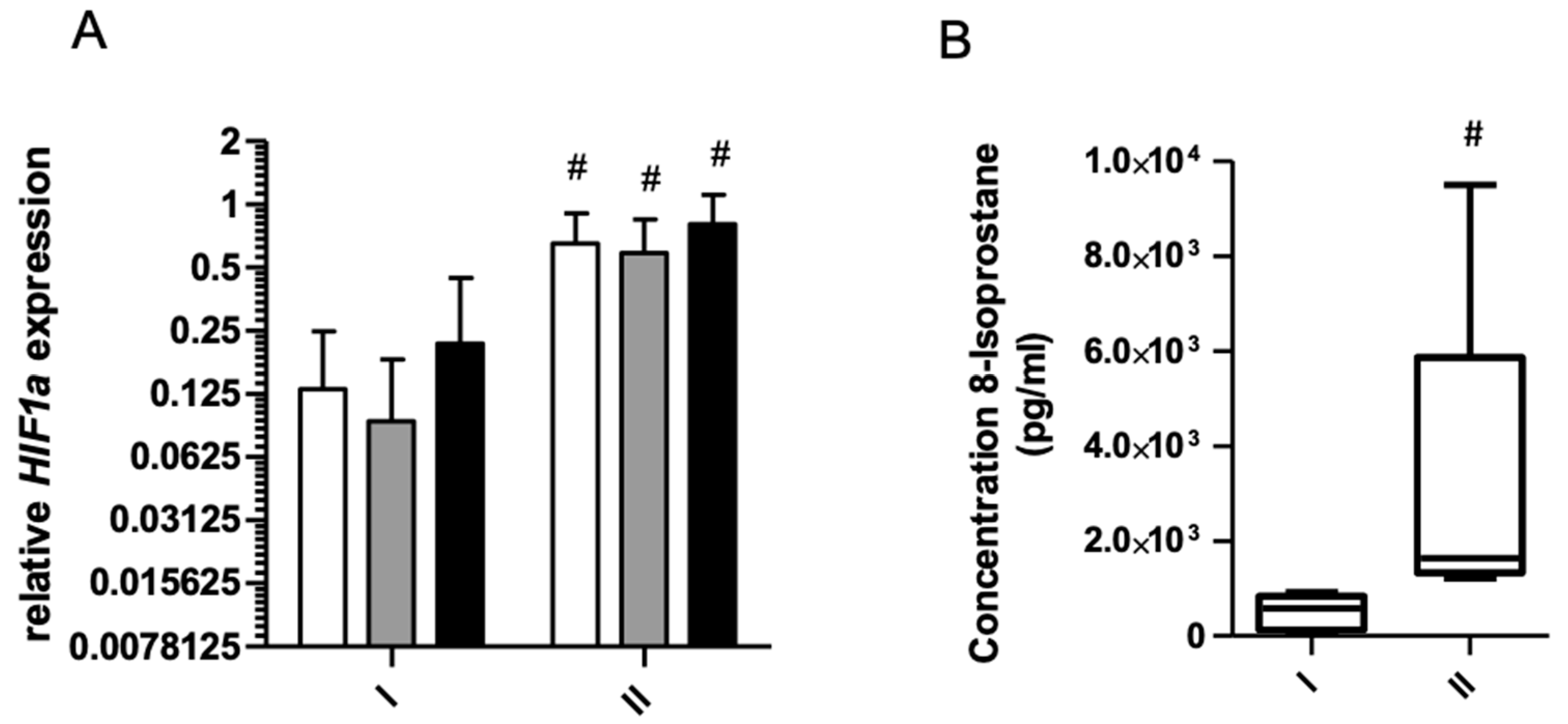
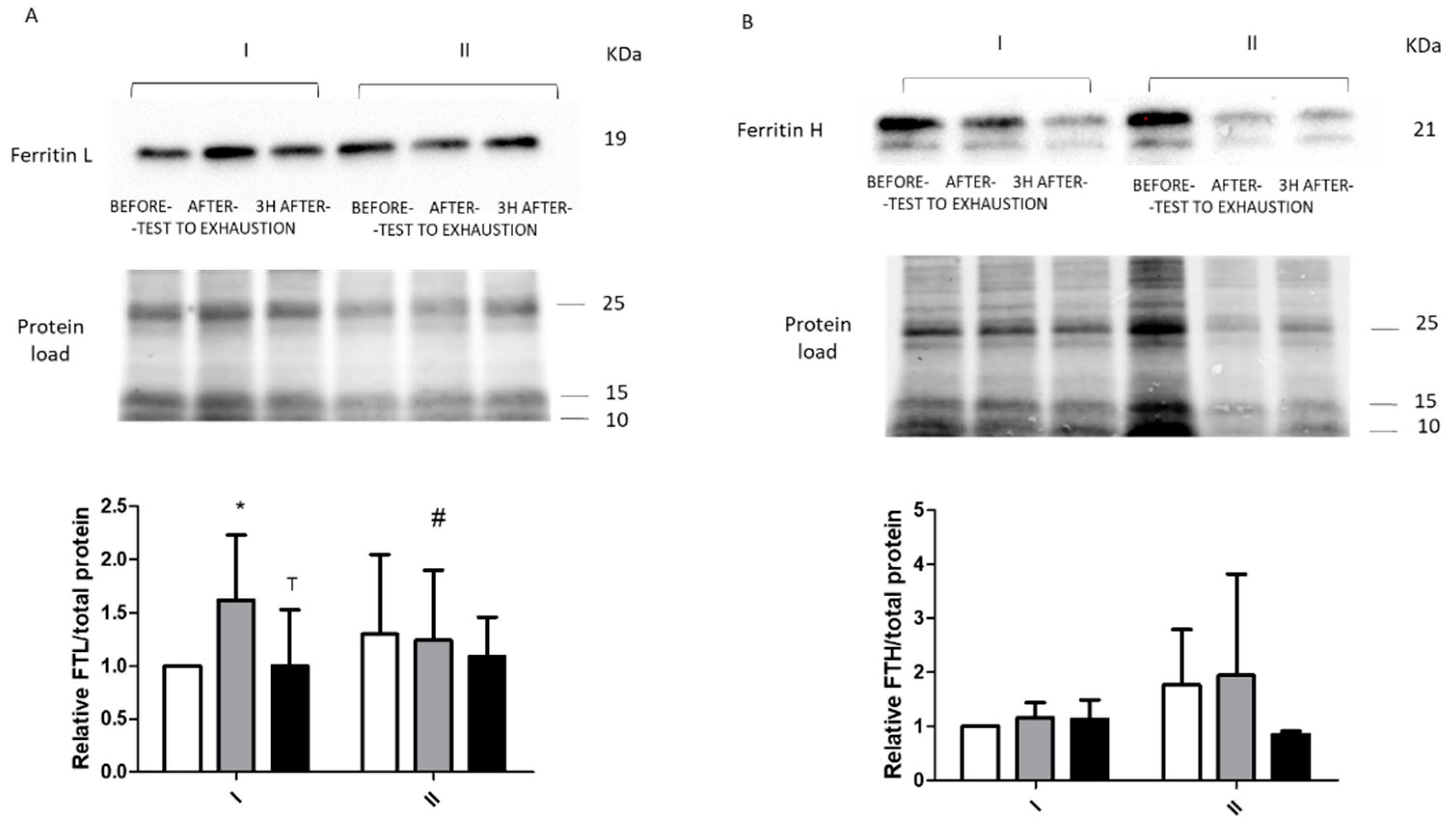
2. Results
2.1. Gene Expression and Oxidative Stress
2.1.1. Effect of 8 Days Starvation on Gene Expression
2.1.2. Effect of 8 Days starvation on 8-Isoprostane
2.1.3. Effects of Exercise on Gene Expression before and after 8 Days of Starvation
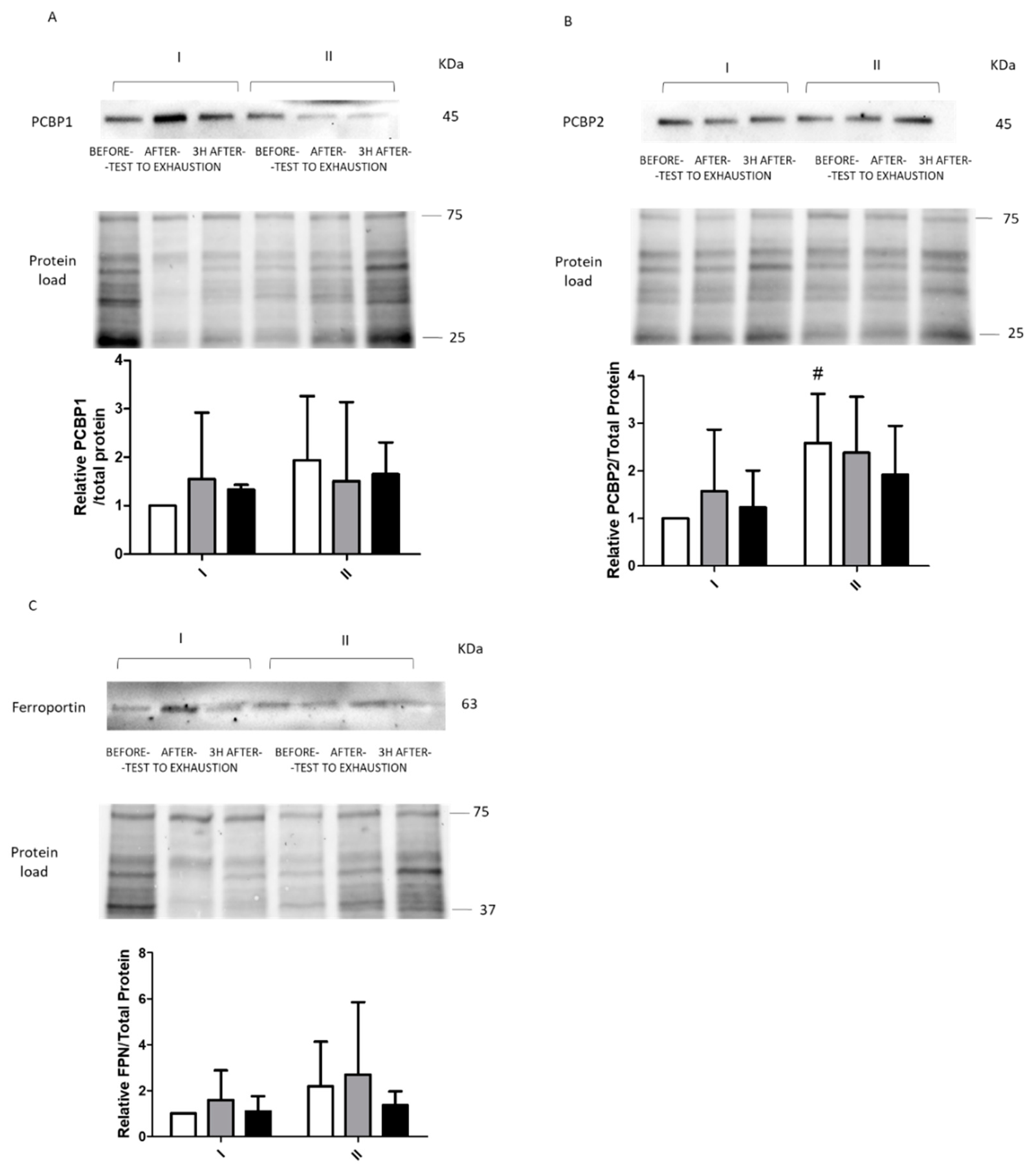
2.2. Immunoblotting
3. Discussion
4. Materials and Methods
4.1. Ethics
4.2. Study Group Characteristics
4.3. Anthropometrics
4.4. Exercise
4.5. Blood Sampling
4.6. Determination of Serum 8-Isoprostane
4.7. RNA Analysis
4.8. Reverse-Transcription and Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
4.9. Immunoblotting
4.10. Statistical Analysis
5. Conclusions
Study Limitation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| APP | amyloid precursor protein |
| BMI | body mass index |
| BV | blood volume |
| DMT1 | divalent metal transporter 1 |
| FFM | fat-free mass |
| FTH | ferritin heavy chain |
| FTL | ferritin light chain |
| FPN | ferroportin |
| Hct | hematocrit |
| Hb | hemoglobin |
| IRP 1-2 | iron regulatory proteins 1-2 |
| LIP | labile iron pool |
| PCBP 1-4 | poly(rC)-binding protein 1-4 |
| PV | plasma volume |
| TBW | total body water; |
| TFRC | transferrin receptor |
| WBC | white blood cells |
References
- Golbidi, S.; Daiber, A.; Korac, B.; Li, H.; Essop, M.F.; Laher, I. Health Benefits of Fasting and Caloric Restriction. Curr. Diabetes Rep. 2017, 17, 123. [Google Scholar] [CrossRef]
- Alaunyte, I.; Stojceska, V.; Plunkett, A. Iron and the female athlete: A review of dietary treatment methods for improving iron status and exercise performance. J. Int. Soc. Sports Nutr. 2015, 12, 38. [Google Scholar] [CrossRef] [Green Version]
- Sanvisens, N.; Bañó, M.C.; Huang, M.; Puig, S. Regulation of ribonucleotide reductase in response to iron deficiency. Mol. Cell 2011, 44, 759–769. [Google Scholar] [CrossRef] [Green Version]
- Salvail, H.; Massé, E. Regulating iron storage and metabolism with RNA: An overview of posttranscriptional controls of intracellular iron homeostasis. Wiley Interdiscip Rev. RNA 2012, 3, 26–36. [Google Scholar] [CrossRef]
- Philpott, C.C.; Ryu, M.S.; Frey, A.; Patel, S. Cytosolic iron chaperones: Proteins delivering iron cofactors in the cytosol of mammalian cells. J. Biol. Chem. 2017, 292, 12764–12771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanatori, I.; Yasui, Y.; Tabuchi, M.; Kishi, F. Chaperone protein involved in transmembrane transport of iron. Biochem. J. 2014, 462, 25–37. [Google Scholar] [CrossRef]
- Yanatori, I.; Richardson, D.R.; Imada, K.; Kishi, F. Iron Export through the Transporter Ferroportin 1 Is Modulated by the Iron Chaperone PCBP2. J. Biol. Chem. 2016, 291, 17303–17318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Bencze, K.Z.; Stemmler, T.L.; Philpott, C.C. A cytosolic iron chaperone that delivers iron to ferritin. Science 2008, 320, 1207–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leidgens, S.; Bullough, K.Z.; Shi, H.; Li, F.; Shakoury-Elizeh, M.; Yabe, T.; Subramanian, P.; Hsu, E.; Natarajan, N.; Nandal, A.; et al. Each member of the poly-r(C)-binding protein 1 (PCBP) family exhibits iron chaperone activity toward ferritin. J. Biol. Chem. 2013, 288, 17791–17802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, M.S.; Zhang, D.; Protchenko, O.; Shakoury-Elizeh, M.; Philpott, C.C. PCBP1 and NCOA4 regulate erythroid iron storage and heme biosynthesis. J. Clin. Investig. 2017, 127, 1786–1797. [Google Scholar] [CrossRef] [Green Version]
- Duce, J.A.; Tsatsanis, A.; Cater, M.A.; James, S.A.; Robb, E.; Wikhe, K.; Leong, S.L.; Perez, K.; Johanssen, T.; Greenough, M.A.; et al. Iron-export ferroxidase activity of β-amyloid precursor protein is inhibited by zinc in Alzheimer′s disease. Cell 2010, 142, 857–867. [Google Scholar] [CrossRef] [Green Version]
- Wong, B.X.; Tsatsanis, A.; Lim, L.Q.; Adlard, P.A.; Bush, A.I.; Duce, J.A. β-Amyloid precursor protein does not possess ferroxidase activity but does stabilize the cell surface ferrous iron exporter ferroportin. PLoS ONE 2014, 9, e114174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojciak, R.W. Effect of short-term food restriction on iron metabolism, relative well-being and depression symptoms in healthy women. Eat. Weight Disord. 2014, 19, 321–327. [Google Scholar] [CrossRef] [Green Version]
- Beard, J.L. Iron biology in immune function, muscle metabolism and neuronal functioning. J. Nutr. 2001, 131, 568S–579S; discussion 580S. [Google Scholar] [CrossRef]
- Lan, Y.L.; Tang, L.; Cui, R.L. Study on the changes of iron metabolism in patients with recurrent oral ulcer and its relationship with cellular immune function. Shanghai Kou Qiang Yi Xue 2017, 26, 302–304. [Google Scholar] [PubMed]
- Oppenheimer, S.J. Iron and its relation to immunity and infectious disease. J. Nutr. 2001, 131, 616S–633S; discussion 633S–635S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, S.Y.; Jeong, K.S.; Kang, B.N.; Park, S.J.; Yoon, W.K.; Kim, S.H.; Kim, T.H. Modulation of transferrin synthesis, transferrin receptor expression, iNOS expression and NO production in mouse macrophages by cytokines, either alone or in combination. Anticancer Res. 2000, 20, 3331–3338. [Google Scholar]
- Cronin, S.J.F.; Woolf, C.J.; Weiss, G.; Penninger, J.M. The Role of Iron Regulation in Immunometabolism and Immune-Related Disease. Front. Mol. Biosci. 2019, 6, 116. [Google Scholar] [CrossRef] [Green Version]
- Kemp, J.D. The Role of Iron and Iron-Binding Proteins in Lymphocyte Physiology and Pathology. J. Clin. Immunol. 1993, 13, 81–92. [Google Scholar] [CrossRef]
- Onabanjo, O.O.; Jerling, J.C.; Covic, N.; Van Graan, A.; Taljaard, C.; Mamabolo, R.L. Association between iron status and white blood cell counts in African schoolchildren of the North-West Province, South Africa. J. Epidemiol. Glob. Health 2012, 2, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Sonanez-Organis, J.G.; Vazquez-Medina, J.P.; Crocker, D.E.; Ortiz, R.M. Prolonged fasting activates hypoxia inducible factors-1 alpha,-2 alpha and-3 alpha in a tissue-specific manner in northern elephant seal pups. Gene 2013, 526, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G.L. Hypoxia-inducible factors in physiology and medicine. Cell 2012, 148, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.L.; Semenza, G.L. General Involvement of Hypoxia-Inducible Factor-I in Transcriptional Response to Hypoxia. Proc. Natl. Acad. Sci. USA 1993, 90, 4304–4308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, M.; Galy, B.; Muckenthaler, M.U.; Hentze, M.W. Iron-regulatory proteins limit hypoxia-inducible factor-2alpha expression in iron deficiency. Nat. Struct. Mol. Biol. 2007, 14, 420–426. [Google Scholar] [CrossRef]
- Borghini, A.; Giardini, G.; Tonacci, A.; Mastorci, F.; Mercuri, A.; Mrakic-Sposta, S.; Sposta, S.M.; Moretti, S.; Andreassi, M.G.; Pratali, L. Chronic and acute effects of endurance training on telomere length. Mutagenesis 2015, 30, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Dalle Carbonare, L.; Manfredi, M.; Caviglia, G.; Conte, E.; Robotti, E.; Marengo, E.; Cheri, S.; Zamboni, F.; Gabbiani, D.; Deiana, M.; et al. Can half-marathon affect overall health? The yin-yang of sport. J. Proteom. 2018, 170, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Kell, D.B.; Pretorius, E. Serum ferritin is an important inflammatory disease marker, as it is mainly a leakage product from damaged cells. Metallomics 2014, 6, 748–773. [Google Scholar] [CrossRef] [Green Version]
- Zabelinskii, S.A.; Chebotareva, M.A.; Tavrovskaya, T.V.; Skverchinskaya, E.A.; Shukolyukova, E.P.; Maslova, M.N.; Krivchenko, A.I. Effect of stress actions on some hematological and biochemical parameters of rat blood and on energetic intermolecular interactions in lipid extract under effect of light radiation. J. Evol. Biochem. Phys. 2012, 48, 500–509. [Google Scholar] [CrossRef]
- Otaki, Y.; Nakanishi, T.; Hasuike, Y.; Moriguchi, R.; Nanami, M.; Hama, Y.; Izumi, M.; Takamitsu, Y. Defective regulation of iron transporters leading to iron excess in the polymorphonuclear leukocytes of patients on maintenance hemodialysis. Am. J. Kidney Dis. 2004, 43, 1030–1039. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, K.M.; Gasche, C. Management of Iron Deficiency Anaemia in Inflammatory Bowel Disease. Acta Haematol. 2019, 142, 30–36. [Google Scholar] [CrossRef]
- Grzybkowska, A.; Anczykowska, K.; Ratkowski, W.; Aschenbrenner, P.; Antosiewicz, J.; Bonisławska, I.; Żychowska, M. Changes in Serum Iron and Leukocyte mRNA Levels of Genes Involved in Iron Metabolism in Amateur Marathon Runners-Effect of the Running Pace. Genes 2019, 10, 460. [Google Scholar] [CrossRef] [Green Version]
- Encinar del Dedo, J.; Gabrielli, N.; Carmona, M.; Ayté, J.; Hidalgo, E. A cascade of iron-containing proteins governs the genetic iron starvation response to promote iron uptake and inhibit iron storage in fission yeast. PLoS Genet. 2015, 11, e1005106. [Google Scholar] [CrossRef] [PubMed]
- Carballo, E.; Lai, W.S.; Blackshear, P.J. Feedback inhibition of macrophage tumor necrosis factor-alpha production by tristetraprolin. Science 1998, 281, 1001–1005. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Pantopoulos, K. Regulation of cellular iron metabolism. Biochem. J. 2011, 434, 365–381. [Google Scholar] [CrossRef] [Green Version]
- Casey, J.L.; Koeller, D.M.; Ramin, V.C.; Klausner, R.D.; Harford, J.B. Iron regulation of transferrin receptor mRNA levels requires iron-responsive elements and a rapid turnover determinant in the 3′ untranslated region of the mRNA. Embo J. 1989, 8, 3693–3699. [Google Scholar] [CrossRef]
- Rogers, J.T.; Randall, J.D.; Cahill, C.M.; Eder, P.S.; Huang, X.D.; Gunshin, H.; Leiter, L.; McPhee, J.; Sarang, S.S.; Utsuki, T.; et al. An iron-responsive element type II in the 5 ′-untranslated region of the Alzheimer′s amyloid precursor protein transcript. J. Biol. Chem. 2002, 277, 45518–45528. [Google Scholar] [CrossRef] [Green Version]
- Schofield, C.J.; Ratcliffe, P.J. Oxygen sensing by HIF hydroxylases. Nat. Rev. Mol. Cell Biol. 2004, 5, 343–354. [Google Scholar] [CrossRef]
- Goralska, M.; Fleisher, L.N.; McGahan, M.C. Hypoxia induced changes in expression of proteins involved in iron uptake and storage in cultured lens epithelial cells. Exp. Eye Res. 2014, 125, 135–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, B.W.; Miyazawa, M.; Tsuji, Y. Distinct regulatory mechanisms of the human ferritin gene by hypoxia and hypoxia mimetic cobalt chloride at the transcriptional and post-transcriptional levels. Cell Signal 2014, 26, 2702–2709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, P.; Chandel, N.S.; Simon, M.C. Cellular adaptation to hypoxia through hypoxia inducible factors and beyond. Nat. Rev. Mol. Cell Biol. 2020, 21, 268–283. [Google Scholar] [CrossRef]
- Han, J.S.; Lee, J.H.; Kong, J.; Ji, Y.; Kim, J.; Choe, S.S.; Kim, J.B. Hypoxia Restrains Lipid Utilization via Protein Kinase A and Adipose Triglyceride Lipase Downregulation through Hypoxia-Inducible Factor. Mol. Cell Biol. 2019, 39, e00390-18. [Google Scholar] [CrossRef] [Green Version]
- Gabryelska, A.; Karuga, F.F.; Szmyd, B.; Bialasiewicz, P. HIF-1 alpha as a Mediator of Insulin Resistance, T2DM, and Its Complications: Potential Links With Obstructive Sleep Apnea. Front. Physiol. 2020, 11, 1035. [Google Scholar] [CrossRef] [PubMed]
- Mastrogiannaki, M.; Matak, P.; Peyssonnaux, C. The gut in iron homeostasis: Role of HIF-2 under normal and pathological conditions. Blood 2013, 122, 885–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastrogiannaki, M.; Matak, P.; Keith, B.; Simon, M.C.; Vaulont, S.; Peyssonnaux, C. HIF-2alpha, but not HIF-1alpha, promotes iron absorption in mice. J. Clin. Investig. 2009, 119, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, J.R.; Heinis, M.; Zumerle, S.; Delga, S.; Le Bon, A.; Peyssonnaux, C. Investigating the real role of HIF-1 and HIF-2 in iron recycling by macrophages. Haematologica 2014, 99, e112–e114. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, W.; Ma, S.F.; Miasniakova, G.; Sergueeva, A.; Ammosova, T.; Xu, M.; Nekhai, S.; Nourai, M.; Wade, M.S.; et al. Iron deficiency modifies gene expression variation induced by augmented hypoxia sensing. Blood Cells Mol. Dis. 2014, 52, 35–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faris, M.A.I.E.; Hussein, R.N.; Al-Kurd, R.A.; Al-Fararjeh, M.A.; Bustanji, Y.K.; KhalilMohammad, A. Impact of Ramadan Intermittent Fasting on Oxidative Stress Measured by Urinary 15-F2t-Isoprostane. J. Nutr. Metab. 2012, 2012, 802924. [Google Scholar] [CrossRef] [Green Version]
- Nandal, A.; Ruiz, J.C.; Subramanian, P.; Ghimire-Rijal, S.; Sinnamon, R.A.; Stemmler, T.L.; Bruick, R.K.; Philpott, C.C. Activation of the HIF Prolyl Hydroxylase by the Iron Chaperones PCBP1 and PCBP2. Cell Metab. 2011, 14, 647–657. [Google Scholar] [CrossRef] [Green Version]
- Kurz, T.; Gustafsson, B.; Brunk, U.T. Intralysosomal iron chelation protects against oxidative stress-induced cellular damage. FEBS J. 2006, 273, 3106–3117. [Google Scholar] [CrossRef]
- Sato, T.; Chang, H.C.; Bayeva, M.; Shapiro, J.S.; Ramos-Alonso, L.; Kouzu, H.; Jiang, X.; Liu, T.; Yar, S.; Sawicki, K.T.; et al. mRNA-binding protein tristetraprolin is essential for cardiac response to iron deficiency by regulating mitochondrial function. Proc. Natl. Acad. Sci. USA 2018, 115, E6291–E6300. [Google Scholar] [CrossRef] [Green Version]
- Borkowska, A.; Popowska, U.; Spodnik, J.; Herman-Antosiewicz, A.; Wozniak, M.; Antosiewicz, J. JNK/p66Shc/ITCH Signaling Pathway Mediates Angiotensin II-induced Ferritin Degradation and Labile Iron Pool Increase. Nutrients 2020, 12, 668. [Google Scholar] [CrossRef] [Green Version]
- Antosiewicz, J.; Herman-Antosiewicz, A.; Marynowski, S.W.; Singh, S.V. c-Jun NH(2)-terminal kinase signaling axis regulates diallyl trisulfide-induced generation of reactive oxygen species and cell cycle arrest in human prostate cancer cells. Cancer Res. 2006, 66, 5379–5386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dill, D.B.; Costill, D.L. Calculation of percentage changes in volumes of blood, plasma, and red cells in dehydration. J. Appl. Physiol. 1974, 37, 247–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n = 13 | Baseline Values before Fasting | Baseline Values after Fasting | Δ(Before−after) | Cohen’s d (Effect Size) | |||
|---|---|---|---|---|---|---|---|
| Parameter | Mean | SD | Mean | SD | Mean | SD | |
| Age | 50.3 ± 14.2 | ||||||
| Body mass (kg) | 79.38 | 10.79 | 73.43 * | 10.26 | –5.96 | 0.79 | 0.57 |
| Fat mass (kg) | 15.28 | 5.42 | 13.21 * | 5.33 | –2.07 | 0.56 | 0.39 |
| FFM (kg) | 64.11 | 5.87 | 60.07 * | 5.88 | –4.04 | 0.89 | 0.69 |
| TBW (kg) | 46.94 | 4.29 | 44.08 * | 4.11 | –2.86 | 0.62 | 0.68 |
| BMI (kg/m2) | 24.89 | 3.52 | 23.02 * | 3.35 | –1.87 | 0.25 | 0.54 |
| β-hydroxybutyrate (mmol/L) | 0.32 | 0.22 | 4.93 * | 0.65 | 3.74 | 1.94 | 9.5 |
| Hb (g%) | 15.43 | 0.91 | 14.47 | 1.24 | –0.39 | 1.83 | 0.88 |
| HCT (%) | 45.4 | 2.80 | 43.5 | 3.66 | –1.09 | 5.56 | 0.58 |
| PV (%) | 100 | - | 98.54 | 7.06 | −1.46 | 7.06 | 0.28 |
| BV (%) | 100 | - | 104.00 | 12.99 | 0.84 | 7.37 | −0.42 |
| Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| TUBB NM_001293213 | TCCACGGCCTTGCTCTTGTTT | GACATCAAGGCGCATGTGAAC |
| PCBP1 NM_006196 | AGAGTCATGACCATTCCGTAC | TCCTTGAATCGAGTAGGCATC |
| PCBP2 NM_001128913 | TCCAGCTCTCCGGTCATCTTT | ACTGAATCCGGTGTTGCCATG |
| PCBP3 NM_020528.3 | TGTGAAGAAGATGCGTGAGG | RGCTGTTGCTATGGAGTTGA |
| PCBP4 NM_033010.2 | CCTGTCTAGAGACGGCCAAG | AGCAGGTGGTAAAGCCAAGA |
| FTH1 NM_002032 | TCCTACGTTTACCTGTCCATG | CTGCAGCTTCATCAGTTTCTC |
| FTL NM_000146 | GTCAATTTGTACCTGCAGGCC | CTCGGCCAATTCGCGGAA |
| TFRC NM_001128148 | TGCAGCAGTGAGTCTCTTCA | AGGCCCATCTCCTTAACGAG |
| APP NM_000484.4 | TCAGTCTCTCTCCCTGCTCT | GGTGGTTTTCGTTTCGTTTCGGTCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borkowska, A.; Tomczyk, M.; Żychowska, M.; Pilis, W.; Zych, M.; Antosiewicz, J. Effect of 8-Day Fasting on Leukocytes Expression of Genes and Proteins Involved in Iron Metabolism in Healthy Men. Int. J. Mol. Sci. 2021, 22, 3248. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063248
Borkowska A, Tomczyk M, Żychowska M, Pilis W, Zych M, Antosiewicz J. Effect of 8-Day Fasting on Leukocytes Expression of Genes and Proteins Involved in Iron Metabolism in Healthy Men. International Journal of Molecular Sciences. 2021; 22(6):3248. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063248
Chicago/Turabian StyleBorkowska, Andżelika, Maja Tomczyk, Małgorzata Żychowska, Wiesław Pilis, Michał Zych, and Jędrzej Antosiewicz. 2021. "Effect of 8-Day Fasting on Leukocytes Expression of Genes and Proteins Involved in Iron Metabolism in Healthy Men" International Journal of Molecular Sciences 22, no. 6: 3248. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063248