
Characterization of Differentially Expressed Genes under Salt Stress in Olive

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
2.1. Plant Growth and Visible Symptoms
2.2. Mineral Leaf Content
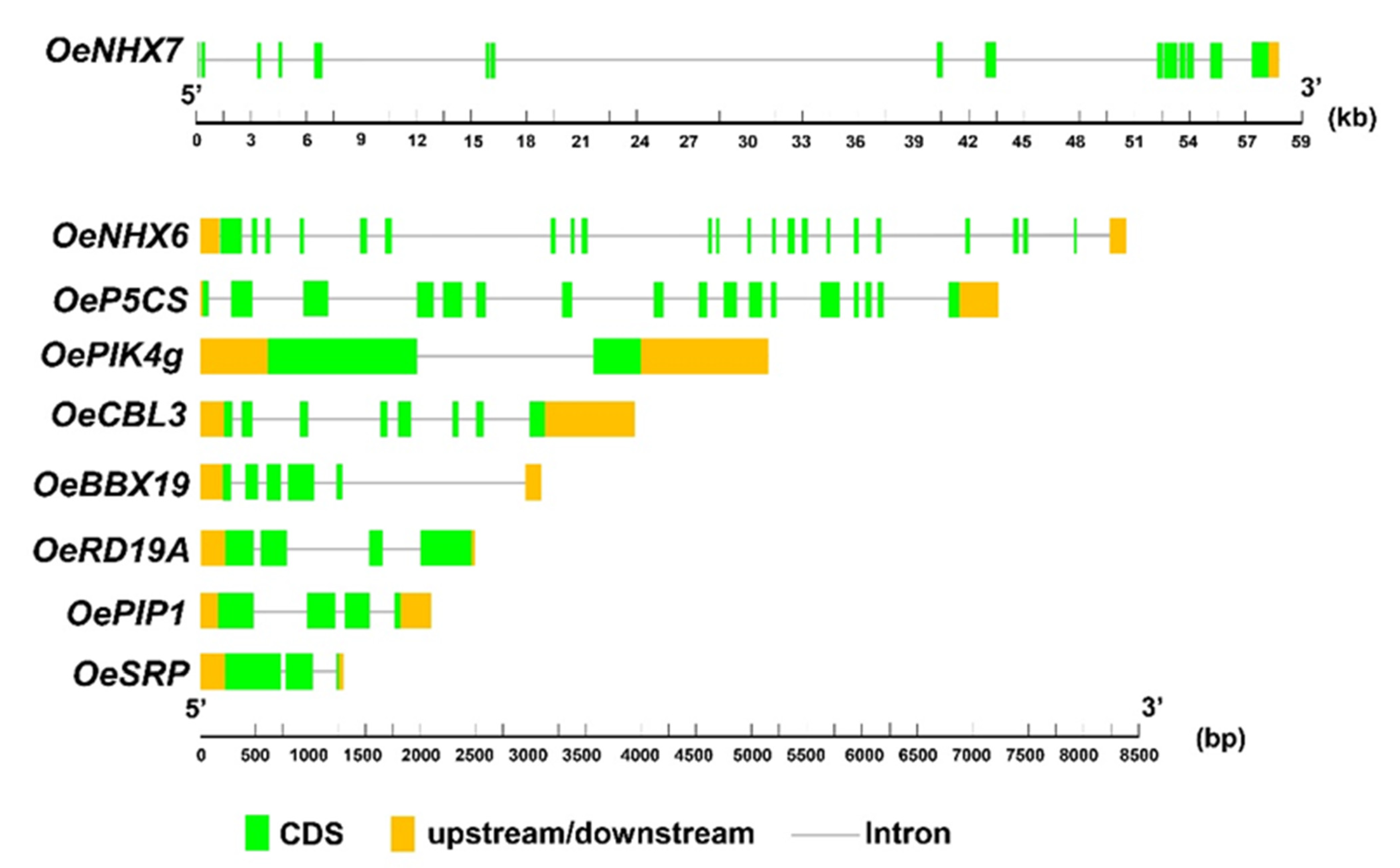
2.3. Identification and Structural Analyses of Selected Genes
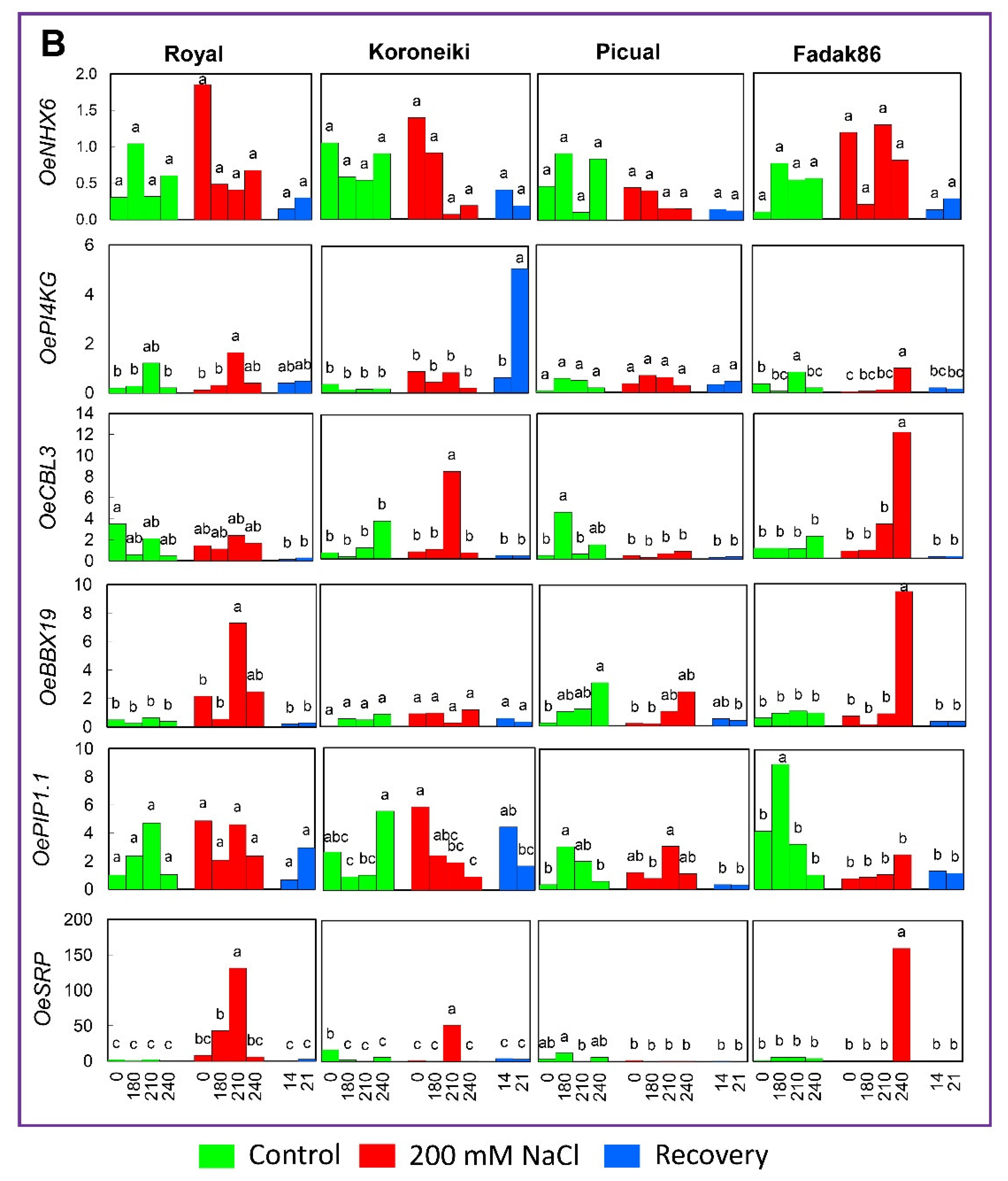
2.4. Gene Expression Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Design
4.2. Plant Growth and Chemical Measurements
4.3. Molecular Characterization of Salt Responsive Genes in Olive
4.4. RNA Extraction, cDNA Synthesis and Gene Expression Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, G.U.; Seo, K.H.; Chen, D. Climate change over the Mediterranean and current destruction of marine ecosystem. Sci. Rep. 2019, 9, 18813. [Google Scholar] [CrossRef]
- Zaied, Y.B.; Zouabi, O. Impacts of climate change on Tunisian olive oil output. Clim. Change 2016, 139, 535–549. [Google Scholar] [CrossRef]
- Mousavi, S.; de la Rosa, R.; Moukhli, A.; El Riachy, M.; Mariotti, R.; Torres, M.; Pierantozzi, P.; Stanzione, V.; Mastio, V.; Zaher, H.; et al. Plasticity of fruit and oil traits in olive among different environments. Sci. Rep. 2019, 9, 16968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mousavi, S.; Mariotti, R.; Bagnoli, F.; Costantini, L.; Cultrera, N.G.; Arzani, K.; Pandolfi, S.; Vendramin, G.G.; Torkzaban, B.; Hosseini-Mazinani, M.; et al. The eastern part of the Fertile Crescent concealed an unexpected route of olive (Olea europaea L.) differentiation. Ann. Bot. 2017, 119, 1305–1318. [Google Scholar] [CrossRef]
- Trujillo, I.; Ojeda, M.A.; Urdiroz, N.M.; Potter, D.; Barranco, D.; Rallo, L.; Diez, C.M. Identification of the Worldwide Olive Germplasm Bank of Córdoba (Spain) using SSR and morphological markers. Tree Genet. 2014, 10, 141–155. [Google Scholar] [CrossRef]
- Diez, C.M.; Trujillo, I.; Martinez-Urdiroz, N.; Barranco, D.; Rallo, L.; Marfil, P.; Gaut, B.S. Olive domestication and diversification in the Mediterranean Basin. New Phytol. 2015, 206, 436–447. [Google Scholar] [CrossRef] [PubMed]
- Kchaou, H.; Larbi, A.; Gargouri, K.; Chaieb, M.; Morales, F.; Msallem, M. Assessment of tolerance to NaCl salinity of five olive cultivars, based on growth characteristics and Na+ and Cl− exclusion mechanisms. Sci. Hortic. 2010, 124, 306–315. [Google Scholar] [CrossRef]
- Kasırğa, E.; Demiral, M. Salt stress-mineral nutrient relations in olive (Olea europaea L.) plant. Eurasian J. Soil Sci. 2016, 5, 307–313. [Google Scholar] [CrossRef] [Green Version]
- Soda, N.; Ephrath, J.E.; Dag, A.; Beiersdorf, I.; Presnov, E.; Yermiyahu, U.; Ben-Gal, A. Root growth dynamics of olive (Olea europaea L.) affected by irrigation induced salinity. Plant Soil 2017, 411, 305–318. [Google Scholar] [CrossRef]
- Mousavi, S.; Regni, L.; Bocchini, M.; Mariotti, R.; Cultrera, N.G.M.; Mancuso, S.; Googlani, J.; Chakerolhosseini, M.R.; Guerrero, C.; Albertini, E.; et al. Physiological, epigenetic and genetic regulation in some olive cultivars under salt stress. Sci. Rep. 2019, 9, 1093. [Google Scholar] [CrossRef] [Green Version]
- Regni, L.; Del Pino, A.M.; Mousavi, S.; Palmerini, C.A.; Baldoni, L.; Mariotti, R.; Mairech, H.; Gardi, T.; D’Amato, R.; Proietti, P. Behavior of four olive cultivars during salt stress. Front. Plant Sci. 2019, 10, 867. [Google Scholar] [CrossRef]
- Rossi, L.; Francini, A.; Minnocci, A.; Sebastiani, L. Salt stress modifies apoplastic barriers in olive (Olea europaea L.): A comparison between a salt-tolerant and a salt-sensitive cultivar. Sci. Hortic. 2015, 192, 38–46. [Google Scholar] [CrossRef]
- Rossi, L.; Borghi, M.; Francini, A.; Lin, X.; Xie, D.Y.; Sebastiani, L. Salt stress induces differential regulation of the phenylpropanoid pathway in Olea europaea cultivars Frantoio (salt-tolerant) and Leccino (salt-sensitive). J. Plant Physiol. 2016, 204, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Zang, W.J.; Wang, H.; Zhao, M.; Yu, X.J.; He, X.; Miao, Y.; Zhou, J. Acetylcholine promotes ROS detoxification against hypoxia/reoxygenation-induced oxidative stress through FoxO3a/PGC-1α dependent superoxide dismutase. Cell. Physiol. Biochem. 2014, 34, 1614–1625. [Google Scholar] [CrossRef] [PubMed]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The role of Na+ and K+ transporters in salt stress adaptation in glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef] [PubMed]
- Ardie, S.W.; Liu, S.; Takano, T. Expression of the AKT1-type K (+) channel gene from Puccinellia tenuiflora, PutAKT1, enhances salt tolerance in Arabidopsis. Plant Cell Rep. 2010, 29, 865–874. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, H.; Sun, L.; Jiao, Y.; Zhang, G.; Miao, C.; Hao, F. NADPH oxidase AtrbohD and AtrbohF function in ROS-dependent regulation of Na+/K+homeostasis in Arabidopsis under salt stress. J. Exp. Bot. 2012, 63, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Ben Abdallah, M.; Trupiano, D.; Polzella, A.; De Zio, E.; Sassi, M.; Scaloni, A.; Zarrouk, M.; Ben Youssef, N.; Scippa, G.S. Unraveling physiological, biochemical and molecular mechanisms involved in olive (Olea europaea L. cv. Chétoui) tolerance to drought and salt stresses. J. Plant Physiol. 2018, 220, 83–95. [Google Scholar] [CrossRef]
- Melgar, J.C.; Syvertsen, J.P.; Martínez, V.; García-Sánchez, F. Leaf gas exchange, water relations, nutrient content and growth in citrus and olive seedlings under salinity. Biol. Plant 2008, 52, 385–390. [Google Scholar] [CrossRef]
- Assimakopoulou, A.; Salmas, I.; Roussos, P.A.; Nifakos, K.; Kalogeropoulos, P.; Kostelenos, G. Salt tolerance evaluation of nine indigenous Greek olive cultivars. J. Plant Nutr. 2017, 40, 1099–1110. [Google Scholar] [CrossRef]
- Bracci, T.; Minnocci, A.; Sebastiani, L. In vitro olive (Olea europaea L.) cvs Frantoio and Moraiolo microshoot tolerance to NaCl. Plant Biosyst. 2008, 142, 563–571. [Google Scholar] [CrossRef]
- Hu, W.; Yuan, Q.; Wang, Y.; Cai, R.; Deng, X.; Wang, J.; Zhou, S.; Chen, M.; Chen, L.; Huang, C.; et al. Overexpression of a wheat aquaporin gene, TaAQP8, enhances salt stress tolerance in transgenic tobacco. Plant Cell Physiol. 2012, 53, 2127–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandhu, D.; Pudussery, M.V.; Kaundal, R.; Suarez, D.L.; Kaundal, A.; Sekhon, R.S. Molecular characterization and expression analysis of the Na+/H+ exchanger gene family in Medicago truncatula. Funct. Integr. Genom. 2018, 18, 141–153. [Google Scholar] [CrossRef]
- Brini, F.; Masmoudi, K. Ion Transporters and abiotic stress tolerance in plants. ISRN Mol. Biol. 2012, 2012, 927436. [Google Scholar] [CrossRef] [PubMed]
- Baghour, M.; Gálvez, F.J.; Sánchez, M.E.; Aranda, M.N.; Venema, K.; Rodríguez-Rosales, M.P. Overexpression of LeNHX2 and SlSOS2 increases salt tolerance and fruit production in double transgenic tomato plants. Plant Physiol. Biochem. 2019, 135, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Yarra, R.; Kirti, P.B. Expressing class I wheat NHX (TaNHX2) gene in eggplant (Solanum melongena L.) improves plant performance under saline condition. Funct. Integr. Genom. 2019, 19, 541–554. [Google Scholar] [CrossRef]
- Liu, P.; Xu, Z.S.; Pan-Pan, L.; Hu, D.; Chen, M.; Li, L.C.; Ma, Y.Z. A wheat PI4K gene whose product possesses threonine autophophorylation activity confers tolerance to drought and salt in Arabidopsis. J. Exp. Bot. 2013, 64, 2915–2927. [Google Scholar] [CrossRef] [Green Version]
- Rubilar-Hernández, C.; Osorio-Navarro, C.; Cabello, F.; Norambuena, L. PI4KIIIβ activity regulates lateral root formation driven by endocytic trafficking to the vacuole. Plant Physiol. 2019, 181, 112–126. [Google Scholar] [CrossRef]
- Adams, E.; and Shin, R. Transport, signaling, and homeostasis of potassium and sodium in plants. J. Integr. Plant Biol. 2014, 56, 231–249. [Google Scholar] [CrossRef]
- Mazars, C.; Bourque, S.; Mithöfer, A.; Pugin, A.; Ranjeva, R. Calcium homeostasis in plant cell nuclei. New Phytol. 2009, 181, 261–274. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Zhang, X.; Giraldo, J.P.; Shabala, S. It is not all about sodium: Revealing tissue specificity and signalling roles of potassium in plant responses to salt stress. Plant Soil 2018, 431, 1–17. [Google Scholar] [CrossRef]
- Luan, S.; Lan, W.; Chul Lee, S. Potassium nutrition, sodium toxicity, and calcium signaling: Connections through the CBL–CIPK network. Curr. Opin. Plant Biol. 2009, 12, 339–346. [Google Scholar] [CrossRef]
- Batistič, O.; Kudla, J. Plant calcineurin B-like proteins and their interacting protein kinases. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2009, 1793, 985–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaumont, F.; Tyerman, S.D. Aquaporins: Highly regulated channels controlling plant water relations. Plant Physiol. 2014, 164, 1600–1618. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Wang, C.; Liu, R.; Han, Q.; Vandeleur, R.K.; Du, J.; Tyerman, S.; Shou, H. Constitutive overexpression of soybean plasma membrane intrinsic protein GmPIP1; 6 confers salt tolerance. BMC Plant Biol. 2014, 14, 181. [Google Scholar] [CrossRef] [Green Version]
- Kapilan, R.; Vaziri, M.; Zwiazek, J.J. Regulation of aquaporins in plants under stress. Biol. Res. 2018, 51, 4. [Google Scholar] [CrossRef]
- Pawłowicz, I.; Masajada, K. Aquaporins as a link between water relations and photosynthetic pathway in abiotic stress tolerance in plants. Gene 2019, 687, 166–172. [Google Scholar] [CrossRef]
- Zargar, S.M.; Nagar, P.; Deshmukh, R.; Nazir, M.; Wani, A.A.; Masoodi, K.Z.; Agrawal, G.K.; Rakwal, R. Aquaporins as potential drought tolerance inducing proteins: Towards instigating stress tolerance. J. Proteom. 2017, 169, 233–238. [Google Scholar] [CrossRef]
- Zheng, L.; Chen, S.; Xie, L.; Lu, Z.; Liu, M.; Han, X.; Qiao, G.; Jiang, J.; Zhuo, R.; Qiu, W.; et al. Overexpression of cysteine protease gene from Salix matsudana enhances salt tolerance in transgenic Arabidopsis. Environ. Exp. Bot. 2018, 147, 53–62. [Google Scholar] [CrossRef]
- Qin, Y.; Tian, Y.; Liu, X. A wheat salinity-induced WRKY transcription factor TaWRKY93 confers multiple abiotic stress tolerance in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2015, 464, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Yang, L.; Jin, Y.; Lin, J.; Liu, F. Generation, annotation, and analysis of a large-scale expressed sequence tag library from Arabidopsis pumila to explore salt-responsive genes. Front. Plant Sci. 2017, 8, 955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laibach, N.; Schmidl, S.; Müller, B.; Bergmann, M.; Prüfer, D.; Schulze Gronover, C. Small rubber particle proteins from Taraxacum brevicorniculatum promote stress tolerance and influence the size and distribution of lipid droplets and artificial poly (cis -1,4-isoprene) bodies. Plant J. 2018, 93, 1045–1061. [Google Scholar] [CrossRef] [Green Version]
- Park, K.Y.; Kim, W.T.; Kim, E.Y. The proper localization of responsive to desiccation 20 in lipid droplets depends on their biogenesis induced by stress-related proteins in vegetative tissues. Biochem. Biophys. Res. Commun. 2018, 495, 1885–1889. [Google Scholar] [CrossRef]
- Kim, E.Y.; Park, K.Y.; Seo, Y.S.; Kim, W.T. Arabidopsis small rubber particle protein homolog SRPs play dual roles as positive factors for tissue growth and development and in drought stress responses. Plant Physiol. 2016, 170, 2494–2510. [Google Scholar] [CrossRef] [Green Version]
- Reddy, P.S.; Jogeswar, G.; Rasineni, G.K.; Maheswari, M.; Reddy, A.R.; Varshney, R.K.; Kishor, P.K. Proline over-accumulation alleviates salt stress and protects photosynthetic and antioxidant enzyme activities in transgenic sorghum [Sorghum bicolor (L.) Moench]. Plant Physiol. Biochem. 2015, 94, 104–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanna, R.; Kronmiller, B.; Maszle, D.R.; Coupland, G.; Holm, M.; Mizuno, T.; Wu, S.H. The Arabidopsis B-box zinc finger family. Plant Cell 2009, 21, 3416–3420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangappa, S.N.; Botto, J.F. The BBX family of plant transcription factors. Trends Plant Sci. 2014, 19, 460–470. [Google Scholar] [CrossRef]
- Xu, Y.; Zhao, X.; Aiwaili, P.; Mu, X.; Zhao, M.; Zhao, J.; Cheng, L.; Ma, C.; Gao, J.; Hong, B. A zinc finger protein BBX19 interacts with ABF3 to affect drought tolerance negatively in chrysanthemum. Plant J. 2020, 103, 1783–1795. [Google Scholar] [CrossRef]
- Wang, M.J.; Ding, L.; Liu, X.H.; Liu, J.X. Two B-box domain proteins, BBX28 and BBX29, regulate flowering time at low ambient temperature in Arabidopsis. Plant Mol. Biol. 2021, 106, 21–32. [Google Scholar] [CrossRef]
- Mariotti, R.; Cultrera, N.G.; Díez, C.M.; Baldoni, L.; Rubini, A. Identification of new polymorphic regions and differentiation of cultivated olives (Olea europaea L.) through plastome sequence comparison. BMC Plant Biol. 2010, 10, 211. [Google Scholar] [CrossRef] [Green Version]
- Bazakos, C.; Manioudaki, M.E.; Therios, I.; Voyiatzis, D.; Kafetzopoulos, D.; Awada, T.; Kalaitzis, P. Comparative transcriptome analysis of two olive cultivars in response to NaCl-stress. PLoS ONE 2012, 7, e42931. [Google Scholar] [CrossRef]
- Bazakos, C.; Manioudaki, M.E.; Sarropoulou, E.; Spano, T.; Kalaitzis, P. 454 Pyrosequencing of olive (Olea europaea L.) transcriptome in response to salinity. PLoS ONE 2015, 10, e0143000. [Google Scholar] [CrossRef] [Green Version]
- Poku, S.A.; Seçgin, Z.; Kavas, M. Overexpression of Ks-type dehydrins gene OeSRC1 from Olea europaea increases salt and drought tolerance in tobacco plants. Mol. Biol. Rep. 2019, 46, 5745–5757. [Google Scholar] [CrossRef]
- Mariotti, R.; Belaj, A.; De La Rosa, R.; Leòn, L.; Brizioli, F.; Baldoni, L.; Mousavi, S. EST–SNP study of Olea europaea L. uncovers functional polymorphisms between cultivated and wild olives. Genes 2020, 11, 916. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Ruiz, J.; Ramírez-Tejero, J.A.; Fernández-Pozo, N.; Leyva-Pérez, M.D.L.O.; Yan, H.; Rosa, R.D.L.; Belaj, A.; Montes, E.; Rodríguez-Ariza, M.O.; Navarro, F.; et al. Transposon activation is a major driver in the genome evolution of cultivated olive trees (Olea europaea L.). Plant Genome 2020, 13, e20010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oteros, J.; García-Mozo, H.; Vázquez, L.; Mestre, A.; Domínguez-Vilches, E.; Galán, C. Modelling olive phenological response to weather and topography. Agric. Ecosyst. Environ. 2013, 179, 62–68. [Google Scholar] [CrossRef]
- Torres, M.; Pierantozzi, P.; Searles, P.; Rousseaux, M.C.; García-Inza, G.; Miserere, A.; Bodoira, R.; Contreras, C.; Maestri, D. Olive cultivation in the southern hemisphere: Flowering, water requirements and oil quality responses to new crop environments. Front. Plant Sci. 2017, 8, 1830. [Google Scholar] [CrossRef] [Green Version]
- Bader, B.; Aissaoui, F.; Kmicha, I.; Salem, A.B.; Chehab, H.; Gargouri, K.; Boujnah, D.; Chaieb, M. Effects of salinity stress on water desalination, olive tree (Olea europaea L. cvs ‘Picholine’,‘Meski’and ‘Ascolana’) growth and ion accumulation. Desalination 2015, 364, 46–52. [Google Scholar] [CrossRef]
- Chartzoulakis, K.; Loupassaki, M.; Bertaki, M.; Androulakis, I. Effects of NaCl salinity on growth, ion content and CO2 assimilation rate of six olive cultivars. Sci. Hortic. 2002, 96, 235–247. [Google Scholar] [CrossRef]
- Müller, M.; Kunz, H.H.; Schroeder, J.I.; Kemp, G.; Young, H.S.; Neuhaus, H.E. Decreased capacity for sodium export out of Arabidopsis chloroplasts impairs salt tolerance, photosynthesis and plant performance. Plant J. 2014, 78, 646–658. [Google Scholar] [CrossRef] [Green Version]
- Gálvez, F.J.; Baghour, M.; Hao, G.; Cagnac, O.; Rodríguez-Rosales, M.P.; Venema, K. Expression of LeNHX isoforms in response to salt stress in salt sensitive and salt tolerant tomato species. Plant Physiol. Biochem. 2012, 51, 109–115. [Google Scholar] [CrossRef]
- Pakzad, R.; Goharrizi, K.J.; Riahi-Madvar, A.; Amirmahani, F.; Mortazavi, M.; Esmaeeli, L. Identification of Lepidium draba Δ 1-pyrroline-5-carboxylate synthetase (P5CS) and assessment of its expression under NaCl stress: P5CS identification in L. draba plant. Proc. Natl. Acad. Sci. India Sect. B Biol. Sc. 2021, 91, 195–203. [Google Scholar] [CrossRef]
- Quan, R.; Wang, J.; Yang, D.; Zhang, H.; Zhang, Z.; Huang, R. EIN3 and SOS2 synergistically modulate plant salt tolerance. Sci. Rep. 2017, 7, 44637. [Google Scholar] [CrossRef] [Green Version]
- Guan, C.; Cui, X.; Liu, H.; Li, X.; Li, M.; Zhang, Y. Proline biosynthesis enzyme genes confer salt tolerance to switchgrass (Panicum virgatum L.) in cooperation with polyamines metabolism. Front. Plant Sci. 2020, 11, 46. [Google Scholar] [CrossRef]
- Funck, D.; Baumgarten, L.; Stift, M.; von Wirén, N.; Schönemann, L. Differential contribution of P5CS isoforms to stress tolerance in Arabidopsis. Front. Plant Sci. 2020, 11, 565134. [Google Scholar] [CrossRef]
- Liu, H.; Hu, M.; Wang, Q.; Cheng, L.; Zhang, Z. Role of papain-like cysteine proteases in plant development. Front. Plant Sci. 2018, 9, 1717. [Google Scholar] [CrossRef] [Green Version]
- Joaquín-Ramos, A.; Huerta-Ocampo, J.Á.; Barrera-Pacheco, A.; De León-Rodríguez, A.; Baginsky, S.; de la Rosa, A.P.B. Comparative proteomic analysis of amaranth mesophyll and bundle sheath chloroplasts and their adaptation to salt stress. J. Plant Physiol. 2014, 171, 1423–1435. [Google Scholar] [CrossRef]
- Mirzaei, S.; Mansouri, M.; Mohammadi-Nejad, G.; Sablok, G. Comparative assessment of chloroplast transcriptional responses highlights conserved and unique patterns across Triticeae members under salt stress. Photosynth. Res. 2018, 136, 357–369. [Google Scholar] [CrossRef]
- Pandey, G.K.; Kanwar, P.; Singh, A.; Steinhorst, L.; Pandey, A.; Yadav, A.K.; Tokas, I.; Sanyal, S.K.; Kim, B.G.; Lee, S.C.; et al. Calcineurin B-like protein-interacting protein kinase CIPK21 regulates osmotic and salt stress responses in Arabidopsis. Plant Physiol. 2015, 169, 780–792. [Google Scholar] [CrossRef] [Green Version]
- Bai, M.; Sun, J.; Liu, J.; Ren, H.; Wang, K.; Wang, Y.; Wang, C.; Dehesh, K. The B-box protein BBX19 suppresses seed germination via induction of ABI5. Plant J. 2019, 99, 1192–1202. [Google Scholar] [CrossRef]
- Jia, J.; Liang, Y.; Gou, T.; Hu, Y.; Zhu, Y.; Huo, H.; Guo, J.; Gong, H. The expression response of plasma membrane aquaporins to salt stress in tomato plants. Environ. Exp. Bot. 2020, 178, 104190. [Google Scholar] [CrossRef]
- Pou, A.; Jeanguenin, L.; Milhiet, T.; Batoko, H.; Chaumont, F.; Hachez, C. Salinity-mediated transcriptional and post-translational regulation of the Arabidopsis aquaporin PIP2; 7. Plant Mol. Biol. 2016, 92, 731–744. [Google Scholar] [CrossRef]
- Katsuhara, M.; Tsuji, N.; Shibasaka, M.; Panda, S.K. Osmotic stress decreases PIP aquaporin transcripts in barley roots but H 2 O 2 is not involved in this process. J. Plant Res. 2014, 127, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Hachez, C.; Veselov, D.; Ye, Q.; Reinhardt, H.; Knipfer, T.; Fricke, W.; Chaumont, F. Short-term control of maize cell and root water permeability through plasma membrane aquaporin isoforms. Plant. Cell Environ. 2012, 35, 185–198. [Google Scholar] [CrossRef]
- Hosseini, S.A.; Gharechahi, J.; Heidari, M.; Koobaz, P.; Abdollahi, S.; Mirzaei, M.; Nakhoda, B.; Salekdeh, G.H. Comparative proteomic and physiological characterisation of two closely related rice genotypes with contrasting responses to salt stress. Funct. Plant Biol. 2015, 42, 527–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fercha, A.; Capriotti, A.L.; Caruso, G.; Cavaliere, C.; Stampachiacchiere, S.; Chiozzi, R.Z.; Laganà, A. Shotgun proteomic analysis of soybean embryonic axes during germination under salt stress. Proteomics 2016, 16, 1537–1546. [Google Scholar] [CrossRef] [PubMed]
- Weissbein, S.; Wiesman, Z.; Ephrath, Y.; Silberbush, M. Vegetative and reproductive response of olive cultivars to moderate saline water irrigation. HortScience 2008, 43, 320–327. [Google Scholar] [CrossRef] [Green Version]
- Cuin, T.A.; Shabala, S. Exogenously supplied compatible solutes rapidly ameliorate NaCl-induced potassium efflux from barley roots. Plant Cell Physiol. 2005, 46, 1924–1933. [Google Scholar] [CrossRef] [Green Version]
- Cruz, F.; Julca, I.; Gómez-Garrido, J.; Loska, D.; Marcet-Houben, M.; Cano, E.; Galán, B.; Frias, L.; Ribeca, P.; Derdak, S.; et al. Genome sequence of the olive tree, Olea europaea. Gigascience 2016, 5, s13742-016. [Google Scholar] [CrossRef] [PubMed]
- Unver, T.; Wu, Z.; Sterck, L.; Turktas, M.; Lohaus, R.; Li, Z.; Yang, M.; He, L.; Deng, T.; Escalante, F.J.; et al. Genome of wild olive and the evolution of oil biosynthesis. Proc. Natl. Acad. Sci. USA 2017, 114, E9413–E9422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solovyev, V.V.; Shahmuradov, I.A.; Salamov, A.A. Identification of promoter regions and regulatory sites. Methods Mol. Biol. 2010, 674, 57–83. [Google Scholar] [CrossRef] [PubMed]
- Onofri, A. DSAASTAT a New Excel VBA Macro to Perform Basic Statistical Analyses of Field Trials; Department of Agriculture and Environmental Sciences, University of Perugia: Perugia, Italy, 2010. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Genotype | Genomic Scaffold | mRNA/CDS | Protein | Chromosome/Linkage Group | Comment | Accession Number |
|---|---|---|---|---|---|---|---|
| OeNHX7 | Farga | Oe6_s07827 | predicted | predicted | OK637283 | ||
| var. sylvestris | NC_036248.1 | published | predicted | 12 | XM_023036083.1 | ||
| Leccino partial | tig00005075 | predicted | - | 6 | |||
| Picual partial | Oleur061Scf1390, Oleur061Scf2766 | predicted partial | - | [56] | |||
| OeNHX6 | Farga | Oe6_s09856 | predicted | predicted | OK637277 | ||
| var. sylvestris | NC_036249.1 | published | published | 13 | XM_023036540.1 | ||
| Leccino partial | scaffold75631, scaffold60078, scaffold108620 | predicted partial | predicted partial | 8 | |||
| Picual partial | Oleur061Scf1057 | predicted | predicted | Published cds do not correspond to the other three genomes | [56] | ||
| OeP5CS | Farga | Oe6_s02546 | predicted | predicted | OL310482 | ||
| var. sylvestris | CM008527.1 | published | published | 13 | XM_023036864.1 | ||
| Leccino | scaffold44052 + 94205.1 | predicted | predicted | 8 | OK637278 | ||
| Picual | Oleur061Scf2321 | predicted | predicted | Published cds are partial | [56] | ||
| OePIK4g | Farga | Oe6_s03937 | predicted | predicted | OL310483 | ||
| var. sylvestris partial | CM008529.1 | not complete | not complete | 15 | XM_023040706.1 | ||
| Leccino | scaffold20411.1 | published | published | 23 * | MF958940.1 | ||
| Picual | Oleur061Scf0433 | predicted | predicted | Published cds do not correspond to the other two genomes | [56] | ||
| OeCBL3 | Farga | Oe6_s03919 | predicted | predicted | OK637279 | ||
| var. sylvestris partial | CM008521.1 | published | published | 7 | Published protein does not correspond to the other three genomes | XM_023001857.1 | |
| Leccino partial | scaffold19373 | predicted partial | predicted partial | 8 | |||
| Picual | Oleur061Scf1775 | predicted | predicted | Published cds is partial | [56] | ||
| OeBBX19 | Farga | Oe6_s01951 | predicted | predicted | OL310484 | ||
| var. sylvestris | NC_036241.1 | published | published | 5 | XM_023020441.1 | ||
| Leccino | scaffold28640.1 | predicted | predicted | 23 * | OK637280 | ||
| Picual partial | Oleur061Scf0258 | predicted partial | predicted partial | Published cds and protein do not correspond to the three genomes | [56] | ||
| OeRD19A | Farga | Oe6_s05921 | predicted | predicted | OL310485 | ||
| not find in var. sylvestris | XM_023001197.1 | ||||||
| Leccino | scaffold33560.1 | predicted | predicted | 6 | OK637281 | ||
| not find in Picual | - | - | |||||
| OePIP1.1 | Farga | Oe6_s00807 | predicted | predicted | OL310486 | ||
| var. sylvestris | NC_036255.1 | published | published | 19 | XM_022990009.1 | ||
| Leccino | scaffold116227.1 | published | published | 14 * | MF784562 | ||
| Picual | Oleur061Scf2846 | predicted | predicted | Published cds and protein are partial | [56] | ||
| OeSRP | Farga | Oe6_s02151 | predicted | predicted | OL310487 | ||
| var. sylvestris | NC_036256.1 | published | published | 20 | XM_022991523.1 | ||
| Leccino | scaffold70781 | predicted | predicted | 17 | OK637282 | ||
| Picual | Oleur061Scf3856 | published | published | [56] | |||
| OePetD | Leccino | scaffold168451 | published | published | Chloroplast gene | GU931818.1 |
| Gene Name | Genomic Sequence Length (bp) | Transcript Sequence Length (bp) | CDS Sequence Length (bp) | Peptide Sequence Length | Molecular Function | Biological Process |
|---|---|---|---|---|---|---|
| OeNHX7 * | 58,522 | 2940 | 2646 | 882 | Potassium/proton and sodium/proton antiporter activity | Potassium ion transmembrane transport; regulation of intracellular pH; regulation of reactive oxygen species metabolic process; response to hydrogen peroxide; response to oxidative stress; response to reactive oxygen species; response to salt stress; sodium ion imports across plasma membrane sodium ion transport |
| OeNHX6 * | 7410 | 2087 | 1572 | 524 | Potassium/proton and sodium/proton antiporter activity | Potassium ion transmembrane transport; regulation of intracellular pH; sodium ion imports across plasma membrane |
| OeP5CS | 6877 | 2570 | 1893 | 631 | ATP binding; delta1-pyrroline-5-carboxylate synthetase activity; glutamate 5-kinase activity; glutamate-5-semialdehyde dehydrogenase activity | Hyperosmotic salinity response; L-proline biosynthetic process; pollen development; proline biosynthetic process; response to oxidative stress; response to salt stress; response to water deprivation; root development |
| OePIK4g | 5122 | 3576 | 1743 | 581 | 1-phosphatidylinositol 4-kinase activity; ATP binding; protein serine/threonine kinase activity | Cellular response to hypoxia; phosphatidylinositol phosphorylation; protein autophosphorylation; response to salt stress |
| OeCBL3 * | 3887 | 1661 | 675 | 225 | Calcium ion binding; kinase binding | Detection of calcium ion; potassium ion homeostasis |
| OeBBX19 | 3175 | 993 | 648 | 216 | Zinc ion binding | Negative regulation of photo-morphogenesis; photo-morphogenesis; regulation of transcription, DNA-templated |
| OeRD19A | 2471 | 1368 | 1131 | 377 | Cysteine-type endopeptidase activity | Defense response to bacterium; proteolysis involved in cellular protein catabolic process; response to osmotic stress; response to salt stress |
| OePIP1.1 | 2109 | 1465 | 858 | 286 | Water channel activity | Response to water deprivation; water transport |
| OeSRP | 1347 | 1059 | 750 | 250 | No information in uniprot | Developmental vegetative growth; lipid droplet organization; pollen development; positive regulation of growth; positive regulation of response to water deprivation |
| OePetD | 160 | - | 159 | 53 | Electron transporter, transferring electrons within cytochrome b6/f complex of photosystem II activity; electron transporter, transferring electrons within the cyclic electron transport pathway of photosynthesis activity | Photosynthetic electron transport chain |
| Gene Name | Primer 5′-3′ | Amplified Lenght (bp) | Position on the Gene Sequence |
|---|---|---|---|
| OeNHX7 | Fw-GGCGCATATTGGAATACACGA | 112 | Exon 20 |
| Rev-GCTGACTGGCCTACTGTTAAGA | |||
| OeNHX6 | Fw-CAGAAGGGCTTGGTCTCTCC | 382 | Exon 13–15 |
| Rev-CATAGCTGGTCCCATGTCGG | |||
| OeP5CS | Fw-GGGAAAGGAGGCCAGAAGAT | 202 | Exon 8–9 |
| Rev-GGGACTCATTGGACTGGTGA | |||
| OePIK4g | Fw-AGTTCTGGTTAGGTGCCTGC | 167 | Exon 1 |
| Rev-TGCGGTCTTGGATATGAGGA | |||
| OeCBL3 | Fw-TGAAACCTTGTTGCTTGAGATCA | 200 | Exon 4–5 |
| Rev-GGATGGAATACAGAAAGTGCACG | |||
| OeBBX19 | Fw-CTCAATGCCAGACCTCAACG | 260 | Exon 4–5 |
| Rev-TGGCAATCATCATGAAGGTGC | |||
| OeRD19A | Fw-TCCACAAGCTGCTGTTCACT | 82 | Exon 4 |
| Rev-CAGCGCTCCGGTTGTACTAA | |||
| OePIP1.1 | Fw-AAATCCGGCAGTGACTTTCG | 92 | Exon 2 |
| Rev-GATGCAGTGTCTTGGAGCCA | |||
| OeSRP | Fw-CCATTGGTAGAAACAGCCGG | 93 | Exon 1 |
| Rev-GCAGGTAATACGACAGCGGA | |||
| OePetD | Fw-AATGATCCTGTATTAAGAGCT | 307 | Chloroplast gene |
| Rev-CTGCGGGATTATTAACAGTA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mousavi, S.; Mariotti, R.; Valeri, M.C.; Regni, L.; Lilli, E.; Albertini, E.; Proietti, P.; Businelli, D.; Baldoni, L. Characterization of Differentially Expressed Genes under Salt Stress in Olive. Int. J. Mol. Sci. 2022, 23, 154. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010154
Mousavi S, Mariotti R, Valeri MC, Regni L, Lilli E, Albertini E, Proietti P, Businelli D, Baldoni L. Characterization of Differentially Expressed Genes under Salt Stress in Olive. International Journal of Molecular Sciences. 2022; 23(1):154. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010154
Chicago/Turabian StyleMousavi, Soraya, Roberto Mariotti, Maria Cristina Valeri, Luca Regni, Emanuele Lilli, Emidio Albertini, Primo Proietti, Daniela Businelli, and Luciana Baldoni. 2022. "Characterization of Differentially Expressed Genes under Salt Stress in Olive" International Journal of Molecular Sciences 23, no. 1: 154. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010154