
Changes in Homogalacturonan Metabolism in Banana Peel during Fruit Development and Ripening

 , ,
, ,
Abstract
:1. Introduction
2. Results
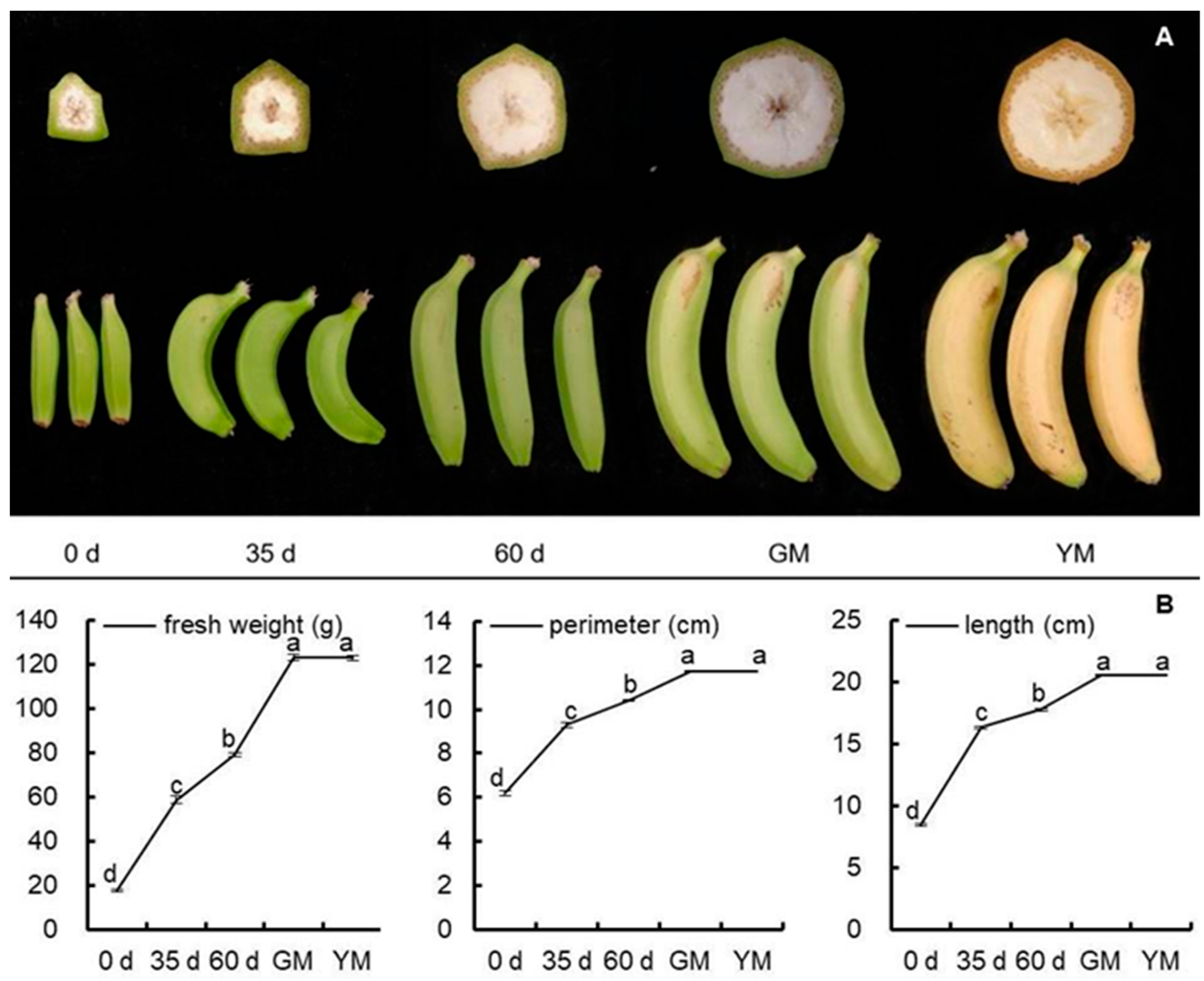
2.1. Morphological and Firmness Analyses
2.2. Systemic Analysis of the Developmental Changes in Gene Expression Levels with RNA-Seq
2.3. The Developmental Changes in the Expression Levels of HG-Modifying Genes
2.4. Enzyme Activities and Fruit Properties
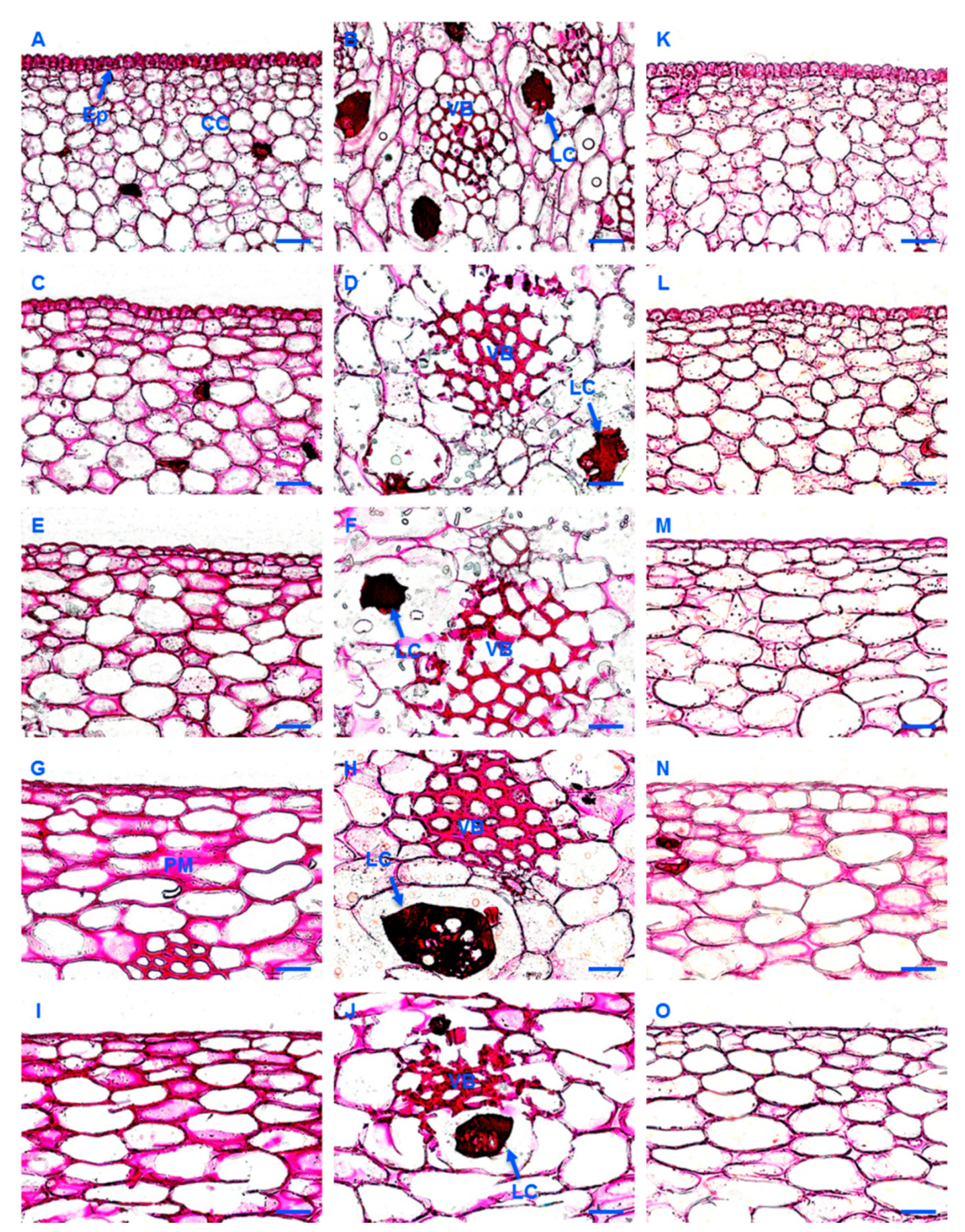
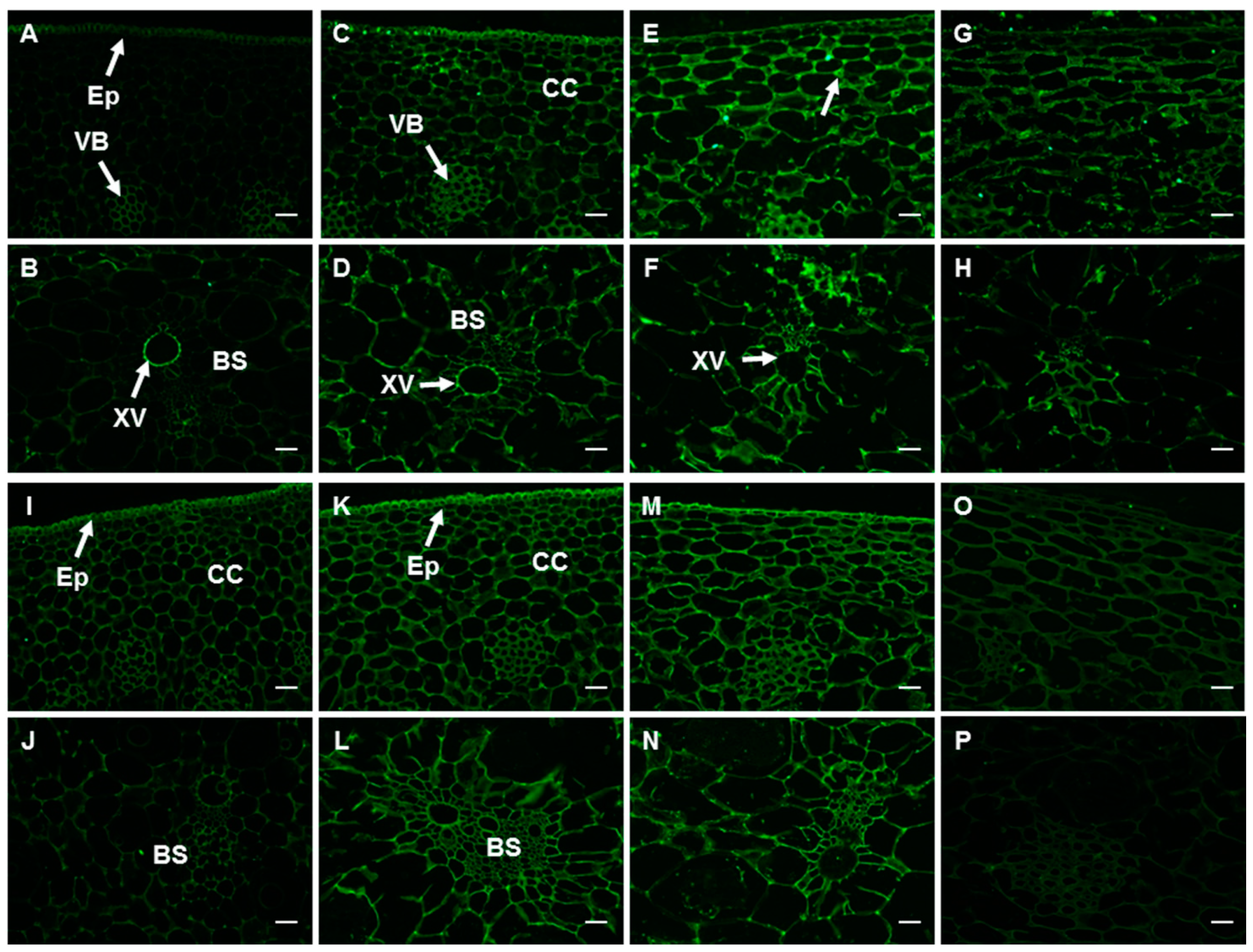
2.5. Histological Analysis
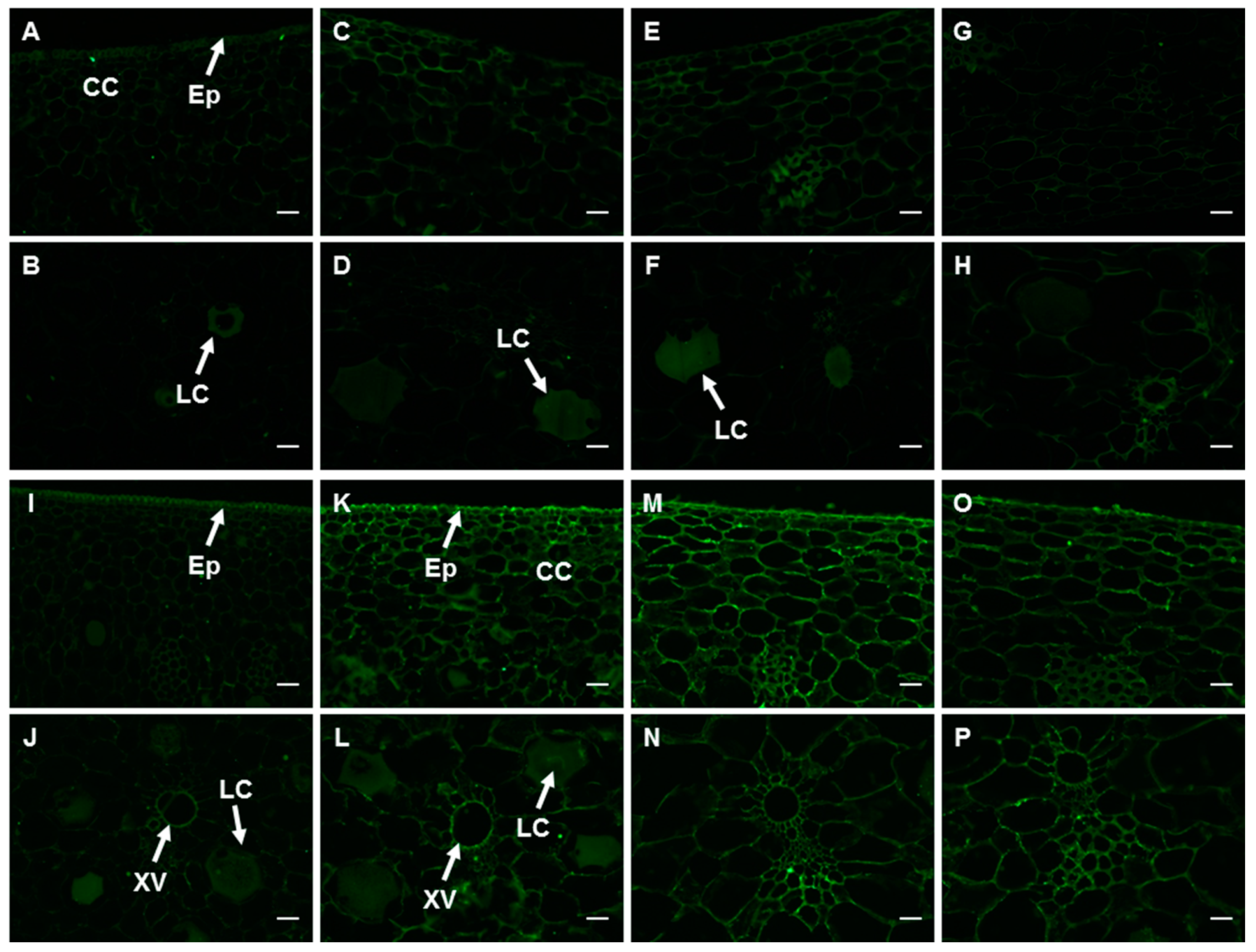
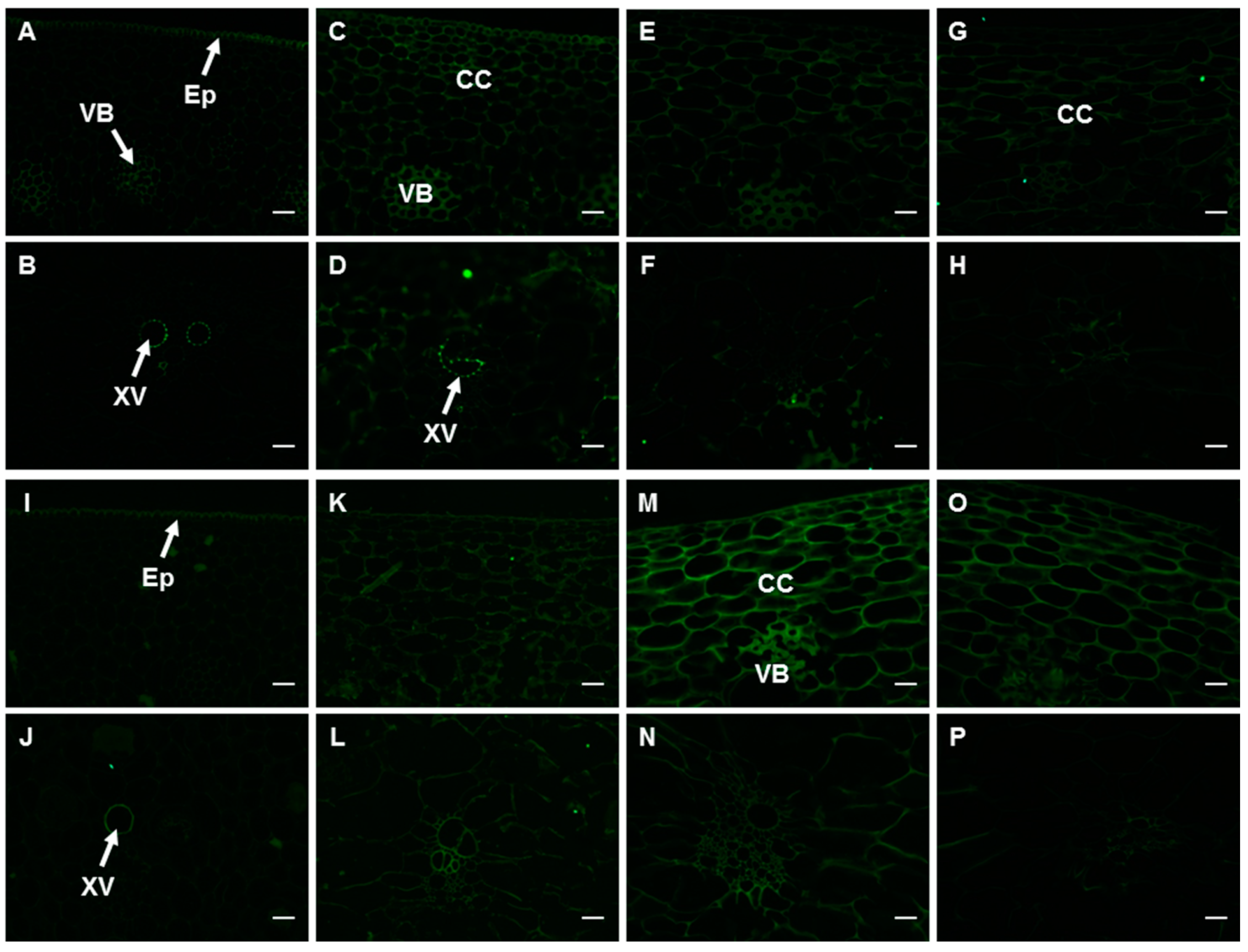
2.6. Changes in the Spatial and Temporal Distribution of HG Components in the Peel
2.6.1. PMEs and the Epitope of 2F4
2.6.2. The Epitopes of JIM5 and JIM7 Antibodies
2.6.3. The Epitopes of CCRC-M38 and LM18 Antibodies
2.6.4. The Calculation of Correlation Coefficient
3. Discussions
3.1. Changes in HG Metabolism during the Development of Banana Fruits
3.2. Changes in HG Metabolism in Banana Peel during the Fruit Ripening Process
4. Materials and Methods
4.1. Plant Materials
4.2. Firmness of Banana Peel
4.3. RNA-Seq Data Analysis
4.4. qPCR
4.5. Activity Assay of HG Degradation Enzymes
4.6. Content of Pectin, Methanol, and DM
4.7. Immuno-Labeling of Antibodies Related to Pectin HG
4.8. Histological Study
4.9. Statistical Analysis and the Calculation of Correlation Coefficient
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| DEG | Differentially expressed gene |
| DM | Degree of pectin methylesterification |
| GalA | Galacturonic acid |
| GAUT | Alpha-1:4-galacturonosyltransferase |
| GM | Green matured |
| HG | Homogalacturonan |
| PBS | Phosphate buffered saline |
| PAE | Pectin acetylesterases |
| PG | Polygalacturonase |
| PGI | Polygalacturonase inhibitor |
| PL | Pectate lyase |
| PME | Pectin methylesterase |
| PMEI | Pectin methylesterase inhibitor |
| RR | Ruthenium red |
| YM | Yellow matured |
References
- Clendennen, S.K.; May, G.D. Differential Gene Expression in Ripening Banana Fruit. Plant Physiol. 1997, 115, 463–469. [Google Scholar] [CrossRef] [Green Version]
- Giovannoni, J. Molecular Biology of Fruit Maturation and Ripening. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 725–749. [Google Scholar] [CrossRef]
- Johnson, K.L.; Gidley, M.J.; Bacic, A.; Doblin, M.S. Cell Wall Biomechanics: A Tractable Challenge in Manipulating Plant Cell Walls ‘Fit for Purpose’! Curr. Opin. Biotechnol. 2018, 49, 163–171. [Google Scholar] [CrossRef]
- Yang, L.; Huang, W.; Xiong, F.J.; Xian, Z.Q.; Su, D.D.; Ren, M.Z.; Li, Z.G. Silencing of Slpl, which Encodes a Pectate Lyase in Tomato, Confers Enhanced Fruit Firmness, Prolonged Shelf-life and Reduced Susceptibility to Grey Mould. Plant Biotechnol. J. 2017, 15, 1544–1555. [Google Scholar] [CrossRef] [Green Version]
- Diarte, C.; Iglesias, A.; Romero, A.; Casero, T.; Ninot, A.; Gatius, F.; Graell, J.; Lara, I. Ripening-related Cell Wall Modifications in Olive (Olea europaea L.) Fruit: A Survey of Nine Genotypes. Food Chem. 2021, 338, 127754. [Google Scholar] [CrossRef]
- Brummell, D.A.; Harpster, M.H. Cell Wall Metabolism in Fruit Softening and Quality and its Manipulation in Transgenic Plants. Plant Mol. Biol. 2001, 47, 311–339. [Google Scholar] [CrossRef]
- Jara, K.; Castro, R.I.; Ramos, P.; Parra-Palma, C.; Valenzuela-Riffo, F.; Morales-Quintana, L. Molecular Insights into FaEG1, a Strawberry Endoglucanase Enzyme Expressed during Strawberry Fruit Ripening. Plants 2019, 8, 140. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.; Tong, C.B.S. Identification of Candidate Genes Involved in Fruit Ripening and Crispness Retention through Transcriptome Analyses of a ‘Honeycrisp’ Population. Plants 2020, 9, 1335. [Google Scholar] [CrossRef]
- Toledo, T.T.; Nogueira, S.B.; Cordenunsi, B.R.; Gozzo, F.C.; Pilau, E.J.; Lajolo, F.M.; Do Nascimento, J.R.O. Proteomic Analysis of Banana Fruit Reveals Proteins that are Differentially Accumulated during Ripening. Postharvest Biol. Technol. 2012, 70, 51–58. [Google Scholar] [CrossRef]
- Du, L.; Song, J.; Forney, C.; Palmer, L.C.; Fillmore, S.; Zhang, Z. Proteome Changes in Banana Fruit Peel Tissue in Response to Ethylene and High-temperature Treatments. Hortic. Res. 2016, 3, 16012. [Google Scholar] [CrossRef] [Green Version]
- Bhuiyan, F.; Campos, N.A.; Swennen, R.; Carpentier, S. Characterizing Fruit Ripening in Plantain and Cavendish Bananas: A Proteomics Approach. J. Proteom. 2020, 214, 103632. [Google Scholar] [CrossRef]
- Li, T.; Yun, Z.; Wu, Q.; Qu, H.; Duan, X.; Jiang, Y. Combination of Transcriptomic, Proteomic, and Metabolomic Analysis Reveals the Ripening Mechanism of Banana Pulp. Biomolecules 2019, 9, 523. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.; Li, T.T.; Jiang, G.X.; Jiang, Y.M.; Duan, X.W. Cell Wall Proteome Analysis of Banana Fruit Softening Using iTRAQ Technology. J. Proteom. 2019, 209, 103506. [Google Scholar] [CrossRef]
- Yun, Z.; Li, T.T.; Gao, H.J.; Zhu, H.; Gupta, V.K.; Jiang, Y.M.; Duan, X.W. Integrated Transcriptomic, Proteomic, and Metabolomics Analysis Reveals Peel Ripening of Harvested Banana under Natural Condition. Biomolecules 2019, 9, 167. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.Y.; Qin, J.J.; Yao, Y.L.; Lai, X.H.; Zheng, W.; Chen, W.X.; Zhu, X.Y.; Li, X.P. A Transcriptomic Analysis Unravels Key Factors in the Regulation of Stay-green Disorder in Peel of Banana Fruit (Fenjiao) Caused by Treatment with 1-MCP. Postharvest Biol. Technol. 2020, 168, 111290. [Google Scholar] [CrossRef]
- Marowa, P.; Ding, A.M.; Kong, Y.Z. Expansins: Roles in Plant Growth and Potential Applications in Crop Improvement. Plant Cell Rep. 2016, 35, 949–965. [Google Scholar] [CrossRef] [Green Version]
- Mayorga-Gómez, A.; Nambeesan, S.U. Temporal Expression Patterns of Fruit-specific α- EXPANSINS during Cell Expansion in Bell Pepper (Capsicum annuum L.). BMC Plant Biol. 2020, 20, 241. [Google Scholar] [CrossRef]
- Ridley, B.L.; O’Neill, M.A.; Mohnen, D. Pectins: Structure, Biosynthesis, and Oligogalacturonide-related Signaling. Phytochemistry 2001, 57, 929–967. [Google Scholar] [CrossRef]
- Willats, W.G.T.; McCartney, L.; Mackie, W.; Knox, J.P. Pectin: Cell Biology and Prospects for Functional Analysis. Plant Mol. Biol. 2001, 47, 9–27. [Google Scholar] [CrossRef]
- Pelloux, J.; Rusterucci, C.; Mellerowicz, E. New Insights into Pectin Methylesterase Structure and Function. Trends Plant Sci. 2007, 12, 267–277. [Google Scholar] [CrossRef]
- Doong, R.L.; Mohnen, D. Solubilization and Characterization of a Galacturonosyltransferase that Synthesizes the Pectic Polysaccharide Homogalacturonan. Plant J. 1998, 13, 363–374. [Google Scholar] [CrossRef]
- Mohnen, D. Pectin Structure and Biosynthesis. Curr. Opin. Plant Biol. 2008, 11, 266–277. [Google Scholar] [CrossRef]
- Atmodjo, M.A.; Hao, Z.; Mohnen, D. Evolving Views of Pectin Biosynthesis. Annu. Rev. Plant Biol. 2013, 64, 747–779. [Google Scholar] [CrossRef] [Green Version]
- Sénéchal, F.; Wattier, C.; Rustérucci, C.; Pelloux, J. Homogalacturonan-modifying Enzymes: Structure, Expression, and Roles in Plants. J. Exp. Bot. 2014, 65, 5125–5160. [Google Scholar] [CrossRef] [Green Version]
- Wolf, S.; Mouille, G.; Pelloux, J. Homogalacturonan Methyl-esterification and Plant Development. Mol. Plant 2009, 2, 851–860. [Google Scholar] [CrossRef]
- Posé, S.; Kirby, A.R.; Paniagua, C.; Waldron, K.W.; Morris, V.J.; Quesada, M.A.; Mercado, J.A. The Nanostructural Characterization of Strawberry Pectins in Pectate Lyase or Polygalacturonase Silenced Fruits Elucidates their Role in Softening. Carbohydr. Polym. 2015, 132, 134–145. [Google Scholar] [CrossRef]
- Marín-Rodríguez, M.C.; Smith, D.L.; Manning, K.; Orchard, J.; Seymour, G.B. Pectate Lyase Gene Expression and Enzyme Activity in Ripening Banana Fruit. Plant Mol. Biol. 2003, 51, 851–857. [Google Scholar] [CrossRef]
- Mbéguié-A-Mbéguié, D.; Hubert, O.; Baurens, F.C.; Matsumoto, T.; Chillet, M.; Fils-Lycaon, B.; Sidibé-Bocs, S. Expression Patterns of Cell Wall-modifying Genes from Banana during Fruit Ripening and in Relationship with Finger Drop. J. Exp. Bot. 2009, 60, 2021–2034. [Google Scholar] [CrossRef] [Green Version]
- Amnuaysin, N.; Jones, M.L.; Seraypheap, K. Changes in Activities and Gene Expression of Enzymes Associated with Cell Wall Modification in Peels of Hot Water Treated Bananas. Sci. Hortic. 2012, 142, 98–104. [Google Scholar] [CrossRef]
- Siddiqui, N.R.; Muhammad, A.; Khan, M.R.; Ali, G.M.; Mahmood, T.; Shahzad, A.; Jabbar, S. Differential Gene Expression of Pectin Esterase and Changes in Pectin during Development and Ripening Stages of Fruit in Selected Cultivars of Banana. Food Sci. Technol. 2020, 40, 827–831. [Google Scholar] [CrossRef] [Green Version]
- Lohani, S.; Trivedi, P.K.; Nath, P. Changes in Activities of Cell Wall Hydrolases during Ethylene-induced Ripening in Banana: Effect of 1-MCP, ABA and IAA. Postharvest Biol. Technol. 2004, 31, 119–126. [Google Scholar] [CrossRef]
- Mirshekari, A.; Ding, P.; Ghazali, H.M. Enzymatic Activity and Microstructural Changes of Hot Water Treated Banana during Ripening. J. Agric. Sci. Technol. 2015, 17, 949–962. [Google Scholar]
- Gayathri, T.; Nair, A.S. Biochemical Analysis and Activity Profiling of Fruit Ripening Enzymes in Banana Cultivars from Kerala. J. Food Meas. Charact. 2017, 11, 1274–1283. [Google Scholar] [CrossRef]
- Chopsri, A.; Sekozawa, Y.; Sugaya, S. Effects of Hot Air Treatment on Cell Wall-degrading Enzymes, Pulp Softening and Ripening in Bananas. Int. Food Res. J. 2018, 25, 2195–2203. [Google Scholar]
- Duan, X.; Cheng, G.; Yang, E.; Yi, C.; Ruenroengklin, N.; Lu, W.; Luo, Y.; Jiang, Y. Modification of Pectin Polysaccharides during Ripening of Postharvest Banana Fruit. Food Chem. 2008, 111, 144–149. [Google Scholar] [CrossRef]
- Xu, J.D.; Yan, J.J.; Li, W.J.; Wang, Q.Y.; Wang, C.X.; Guo, J.X.; Geng, D.L.; Guan, Q.M.; Ma, F.W. Integrative Analyses of Widely Targeted Metabolic Profiling and Transcriptome Data Reveals Molecular Insight into Metabolomic Variations during Apple (Malus domestica) Fruit Development and Ripening. Int. J. Mol. Sci. 2020, 21, 4797. [Google Scholar] [CrossRef]
- Rodriguez, C.E.; Bustamante, C.A.; Budde, C.O.; Müller, G.L.; Drincovich, M.F.; Lara, M.V. Peach Fruit Development: A Comparative Proteomic Study between Endocarp and Mesocarp at Very Early Stages Underpins the Main Differential Biochemical Processes between these Tissues. Front. Plant Sci. 2019, 10, 715. [Google Scholar] [CrossRef] [Green Version]
- D’Hont, A.; Denoeud, F.; Aury, J.; Baurens, F.; Carreel, F.; Garsmeur, O.; Noel, B.; Bocs, S.; Droc, G.; Rouard, M.; et al. The Banana (Musa Acuminata) Genome and the Evolution of Monocotyledonous Plants. Nature 2012, 488, 213–217. [Google Scholar] [CrossRef] [Green Version]
- Asif, M.H.; Lakhwani, D.; Pathak, S.; Gupta, P.; Bag, S.K.; Nath, P.; Trivedi, P.K. Transcriptome Analysis of Ripe and Unripe Fruit Tissue of Banana Identifies Major Metabolic Networks Involved in Fruit Ripening Process. BMC Plant Biol. 2014, 14, 316. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.G.; Wang, J.X.; Hu, Y.L.; Xie, J.H. Nutritional Component Changes in Xiangfen 1 Banana at Different Developmental Stages. Food Funct. 2020, 11, 8286–8296. [Google Scholar] [CrossRef]
- Lakhwani, D.; Pandey, A.; Sharma, D.; Asif, M.H.; Trivedi, P.K. Novel Micrornas Regulating Ripening-associated Processes in Banana Fruit. Plant Growth Regul. 2020, 90, 223–235. [Google Scholar] [CrossRef]
- Inaba, A.; Liu, X.; Yokotani, N.; Yamane, M.; Lu, W.; Nakano, R.; Kubo, Y. Differential Feedback Regulation of Ethylene Biosynthesis in Pulp and Peel Tissues of Banana Fruit. J. Exp. Bot. 2006, 58, 1047–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elitzur, T.; Vrebalov, J.; Giovannoni, J.J.; Goldschmidt, E.E.; Friedman, H. The Regulation of Mads-box Gene Expression during Ripening of Banana and Their Regulatory Interaction with Ethylene. J. Exp. Bot. 2010, 61, 1523–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bargel, H.; Neinhuis, C. Tomato (Lycopersicon esculentum Mill.) Fruit Growth and Ripening as Related to the Biomechanical Properties of Fruit Skin and Isolated Cuticle. J. Exp. Bot. 2005, 56, 1049–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasoli, M.; Dell Anna, R.; Dal Santo, S.; Balestrini, R.; Sanson, A.; Pezzotti, M.; Monti, F.; Zenoni, S. Pectins, Hemicelluloses and Celluloses Show Specific Dynamics in the Internal and External Surfaces of Grape Berry Skin during Ripening. Plant Cell Physiol. 2016, 57, 1332–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, C.W.; Somerville, C.; Anderson, C.T. Polygalacturonase Involved In Expansion1 Functions in Cell Elongation and Flower Development in Arabidopsis. Plant Cell. 2014, 26, 1018–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanal, B.P.; Knoche, M. Mechanical Properties of Cuticles and Their Primary Determinants. J. Exp. Bot. 2017, 68, 5351–5367. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.H.; Holman, T.J.; SÃ Rensen, I.; Cancho-Sanchez, E.; Wells, D.M.; Swarup, R.; Knox, J.P.; Willats, W.G.T.; Ubeda-tomã, S.S.; Holdsworth, M.; et al. Multi-omics Analysis Identifies Genes Mediating the Extension of Cell Walls in the Arabidopsis thaliana Root Elongation Zone. Front. Cell Dev. Biol. 2015, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Terao, A.; Hyodo, H.; Satoh, S.; Iwai, H. Changes in the Distribution of Cell Wall Polysaccharides in Early Fruit Pericarp and Ovule, from Fruit Set to Early Fruit Development, in Tomato (Solanum lycopersicum). J. Plant Res. 2013, 126, 719–728. [Google Scholar] [CrossRef]
- Posé, S.; Marcus, S.E.; Knox, J.P. Differential Metabolism of Pectic Galactan in Tomato and Strawberry Fruit: Detection of the LM26 Branched Galactan Epitope in Ripe Strawberry Fruit. Physiol. Plant. 2018, 164, 95–105. [Google Scholar] [CrossRef]
- Wang, D.; Yeats, T.H.; Uluisik, S.; Rose, J.K.C.; Seymour, G.B. Fruit Softening: Revisiting the Role of Pectin. Trends Plant Sci. 2018, 23, 302–310. [Google Scholar] [CrossRef]
- Brummell, D.A. Cell Wall Disassembly in Ripening Fruit. Funct. Plant Biol. 2006, 33, 103–119. [Google Scholar] [CrossRef]
- Paniagua, C.; Ric-Varas, P.; Garcia-Gago, J.A.; Lopez-Casado, G.; Blanco-Portales, R.; Munoz-Blanco, J.; Schuckel, J.; Knox, J.P.; Matas, A.J.; Quesada, M.A. Elucidating the Role of Polygalacturonase Genes in Strawberry Fruit Softening. J. Exp. Bot. 2020, 71, 7103–7117. [Google Scholar] [CrossRef] [PubMed]
- Uluisik, S.; Seymour, G. Pectate Lyases: Their Role in Plants and Importance in Fruit Ripening. Food Chem. 2020, 309, 125559. [Google Scholar] [CrossRef] [PubMed]
- Dheilly, E.; Gall, S.L.; Guillou, M.; Renou, J.; Bonnin, E.; Orsel, M.; Lahaye, M. Cell Wall Dynamics during Apple Development and Storage Involves Hemicellulose Modifications and Related Expressed Genes. BMC Plant Biol. 2016, 16, 201. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.L.; Ma, M.; Zhang, H.P.; Zhang, S.L.; Qian, M.; Zhang, Z.; Luo, W.Q.; Fan, J.B.; Liu, Z.Q.; Wang, L.B. Genome-wide Analysis of Polygalacturonase Gene Family from Pear Genome and Identification of the Member Involved in Pear Softening. BMC Plant Biol. 2019, 19, 587. [Google Scholar] [CrossRef] [PubMed]
- Dautt-Castro, M.; López-Virgen, A.G.; Ochoa-Leyva, A.; Contreras-Vergara, C.A.; Sortillón-Sortillón, A.P.; Martínez-Téllez, M.A.; González-Aguilar, G.A.; Casas-Flores, J.S.; Sañudo-Barajas, A.; Kuhn, D.N.; et al. Genome-Wide Identification of Mango (Mangifera indica L.) Polygalacturonases: Expression Analysis of Family Members and Total Enzyme Activity during Fruit Ripening. Front. Plant Sci. 2019, 10, 969. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.J.; Chen, M.Y.; Zhao, T.T.; Han, F.; Zhang, Q.; Liu, X.L.; Jiang, C.Y.; Zhong, C.H. Genome-Wide Identification and Expression Analysis of Polygalacturonase Gene Family in Kiwifruit (Actinidia chinensis) during Fruit Softening. Plants 2020, 9, 327. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, W.; Wang, Y.Q.; Liu, Y.Y.; Wang, J.X.; Zhang, X.Q.; Ye, D.; Chen, L.Q. Arabidopsis Galacturonosyltransferase (GAUT) 13 and GAUT14 Have Redundant Functions in Pollen Tube Growth. Mol. Plant 2013, 6, 1131–1148. [Google Scholar] [CrossRef] [Green Version]
- Uluisik, S.; Chapman, N.H.; Smith, R.; Poole, M.; Adams, G.; Gillis, R.B.; Besong, T.M.D.; Sheldon, J.; Stiegelmeyer, S.; Perez, L.; et al. Genetic Improvement of Tomato by Targeted Control of Fruit Softening. Nat. Biotechnol. 2016, 34, 950–952. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Samsulrizal, N.H.; Yan, C.; Allcock, N.S.; Craigon, J.; Blanco-Ulate, B.; Ortega-Salazar, I.; Marcus, S.E.; Bagheri, H.M.; Perez Fons, L.; et al. Characterization of CRISPR Mutants Targeting Genes Modulating Pectin Degradation in Ripening Tomato. Plant Physiol. 2019, 179, 544–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.; Wu, F.; Zhao, Y.; Shi, L.; Zhu, X. Cloning and Expression Analysis of Polygalacturonase and Pectin Methylesterase Genes during Softening in Apricot (Prunus armeniaca L.) Fruit. Sci. Hortic. 2019, 256, 108607. [Google Scholar] [CrossRef]
- Zhou, M.; Jiang, D.; Liu, Y.X.; Wang, X.R.; Tang, H.R.; Chen, Q. amiRNA-mediated PL Gene Silencing Driven by the Fruit-specific Promoter P119 in Strawberry and Its Effect on Fruit Firmness. Bull. Bot. Res. 2019, 39, 441–449. [Google Scholar] [CrossRef]
- Xue, C.; Guan, S.C.; Chen, J.Q.; Wen, C.J.; Cai, J.F.; Chen, X. Genome Wide Identification and Functional Characterization of Strawberry Pectin Methylesterases Related to Fruit Softening. BMC Plant Biol. 2020, 20, 13. [Google Scholar] [CrossRef] [PubMed]
- Pieczywek, P.M.; Kozioł, A.; Konopacka, D.; Cybulska, J.; Zdunek, A. Changes in Cell Wall Stiffness and Microstructure in Ultrasonically Treated Apple. J. Food Eng. 2017, 197, 1–8. [Google Scholar] [CrossRef]
- Paniagua, C.; Posé, S.; Morris, V.J.; Kirby, A.R.; Quesada, M.A.; Mercado, J.A. Fruit Softening and Pectin Disassembly: An Overview of Nanostructural Pectin Modifications Assessed by Atomic Force Microscopy. Ann. Bot. 2014, 114, 1375–1383. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.L.; Fan, W.; Li, X.Q.; Chen, H.B.; Takac, T.; Samajova, O.; Fabrice, M.R.; Xie, L.; Ma, J.; Samaj, J.; et al. Expression and Distribution of Extensins and AGPs in Susceptible and Resistant Banana Cultivars in Response to Wounding and Fusarium oxysporum. Sci. Rep. 2017, 7, 42400. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based Genome Alignment and Genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; et al. Gene Set Enrichment Analysis: A Knowledge-based Approach for Interpreting Genome-wide Expression Profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.Y.; Wilkins, T.A. A Modified Hot Borate Method Significantly Enhances the Yield of High-Quality RNA from Cotton (Gossypium hirsutum L.). Anal. Biochem. 1994, 223, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhong, H.Y.; Kuang, J.F.; Lu, W.J.; Chen, J.Y. Validation of Reference Genes for RT-qPCR Studies of Gene Expression in Banana Fruit under Different Experimental Conditions. Planta 2011, 234, 377–390. [Google Scholar] [CrossRef]
- Marcus, L.; Schejter, A. Single Step Chromatographic Purification and Characterization of the Endopolygalacturonases. Physiol. Plant Pathol. 1983, 22, 1–13. [Google Scholar] [CrossRef]
- Ma, L.; Jiang, S.; Lin, G.M.; Cai, J.H.; Ye, X.Y.; Chen, H.B.; Li, M.H.; Li, H.P.; Takáč, T.; Šamaj, J.; et al. Wound-induced Pectin Methylesterases Enhance Banana (Musa spp. AAA) Susceptibility to Fusarium oxysporum f. sp. Cubense. J. Exp. Bot. 2013, 64, 2219–2229. [Google Scholar] [CrossRef] [PubMed]
- Pathak, N.; Sanwal, G.G. Multiple Forms of Polygalacturonase from Banana Fruits. Phytochemistry 1998, 48, 249–255. [Google Scholar] [CrossRef]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Louvet, R.; Rayon, C.; Domon, J.; Rusterucci, C.; Fournet, F.; Leaustic, A.; Crépeau, M.; Ralet, M.; Rihouey, C.; Bardor, M.; et al. Major Changes in the Cell Wall during Silique Development in Arabidopsis thaliana. Phytochemistry 2011, 72, 59–67. [Google Scholar] [CrossRef]
- Xu, C.X.; Takáč, T.; Burbach, C.; Menzel, D.; Šamaj, J. Developmental Localization and the Role of Hydroxyproline Rich Glycoproteins during Somatic Embryogenesis of Banana (Musa spp. AAA). BMC Plant Biol. 2011, 11, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.X.; Zhao, L.; Pan, X.; Samaj, J. Developmental Localization and Methylesterification of Pectin Epitopes during Somatic Embryogenesis of Banana (Musa spp. AAA). PLoS ONE 2011, 6, e22992. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 0 d | 35 d | 60 d | GM | YM | |
|---|---|---|---|---|---|
| PME activity (U) | 15.59 ± 2.69 ab | 11.09 ± 1.72 b | 12.09 ± 1.78 b | 13.09 ± 1.30 b | 21.10 ± 2.95 a |
| PL activity (U) | 37.29 ± 1.61 bc | 28.78 ± 7.58 c | 38.31 ± 5.85 bc | 65.07 ± 10.29 b | 146.13 ± 15.79 a |
| PG activity (U) | 55.92 ± 12.92 b | 60.76 ± 5.36 b | 46.80 ± 15.64 b | 98.57 ± 24.28 b | 650.51 ± 121.35 a |
| Pectin content mg/g(dry weight) | 62.70 ± 0.22 a | 57.91 ± 2.82 ab | 53.40 ± 2.78 ab | 49.26 ± 2.36 b | 57.83 ± 2.41 ab |
| DM (%) | 59.07 ± 3.29 a | 48.98 ± 2.45 a | 51.34 ± 5.95 a | 52.85 ± 1.51 a | 47.60 ± 4.58 a |
| Peel firmness (N) | 10.90 ± 0.58 b | 12.20 ± 0.58 a | 12.70 ± 0.26 a | 10.77 ± 0.53 b | 4.93 ± 0.24c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ning, T.; Chen, C.; Yi, G.; Chen, H.; Liu, Y.; Fan, Y.; Liu, J.; Chen, S.; Wei, S.; Li, Z.; et al. Changes in Homogalacturonan Metabolism in Banana Peel during Fruit Development and Ripening. Int. J. Mol. Sci. 2022, 23, 243. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010243
Ning T, Chen C, Yi G, Chen H, Liu Y, Fan Y, Liu J, Chen S, Wei S, Li Z, et al. Changes in Homogalacturonan Metabolism in Banana Peel during Fruit Development and Ripening. International Journal of Molecular Sciences. 2022; 23(1):243. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010243
Chicago/Turabian StyleNing, Tong, Chengjie Chen, Ganjun Yi, Houbin Chen, Yudi Liu, Yanjie Fan, Jing Liu, Shule Chen, Sixuan Wei, Zexuan Li, and et al. 2022. "Changes in Homogalacturonan Metabolism in Banana Peel during Fruit Development and Ripening" International Journal of Molecular Sciences 23, no. 1: 243. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010243