
Overexpression of the Aldehyde Dehydrogenase Gene ZmALDH Confers Aluminum Tolerance in Arabidopsis thaliana

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Cloning and Sequence Analysis of ZmALDH
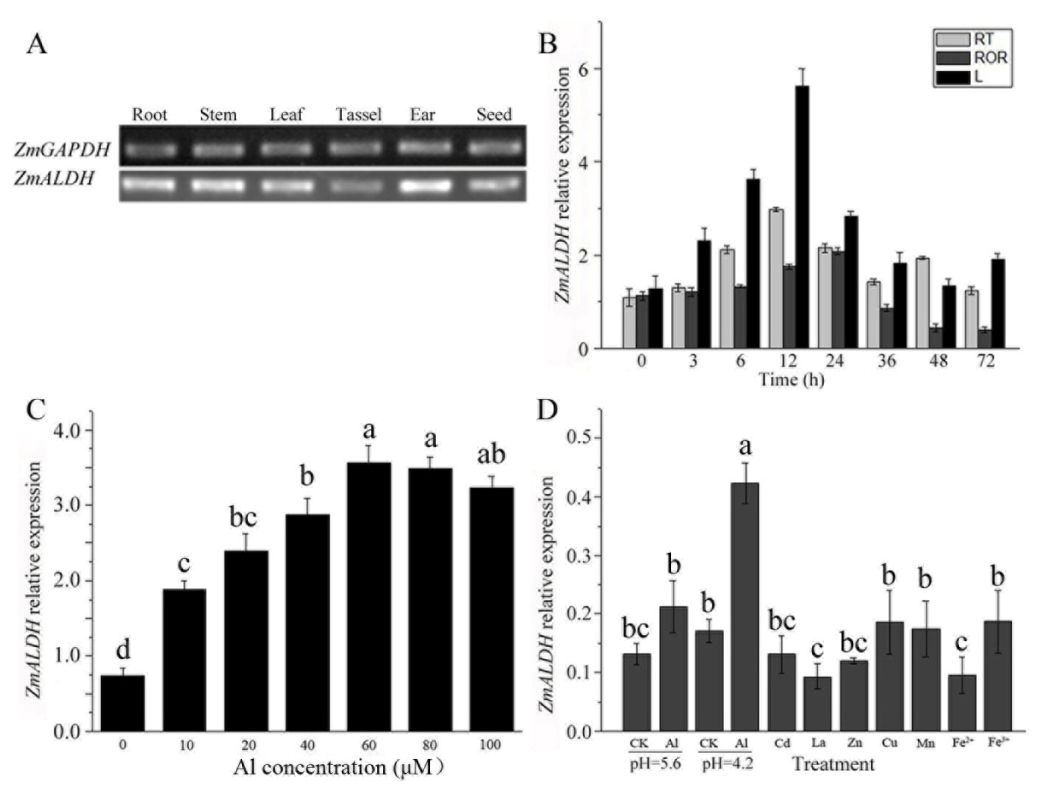
2.2. Expression Analysis of ZmALDH
2.3. Subcellular Localization
2.4. Root Growth Analysis of ZmALDH Transgenic Arabidopsis
2.5. ZmALDH Overexpression Reduces Al Accumulation
2.6. Comparative Analysis of the Oxidative Stress Response under Al Treatment
2.7. ZmALDH Enhanced the Tolerance to Oxidative Stress Induced by Al Toxicity
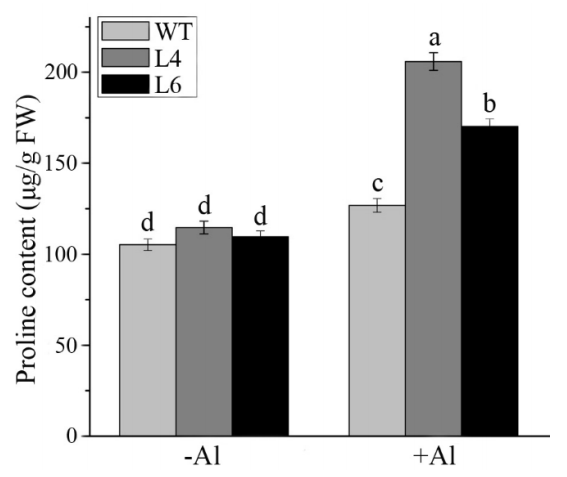
2.8. ZmALDH Promoted the Accumulation of Cellular Osmolytes
3. Discussion
4. Materials and Methods
4.1. Plant Materials, Growth Conditions, and Treatment
4.2. Cloning and Sequence Analysis of ZmALDH
4.3. RNA Isolation and Gene Expression Analysis
4.4. Subcellular Localization
4.5. Generation of Transgenic Arabidopsis Plants
4.6. Measurement of Oxidative Damage
4.7. Determination of Reactive Oxygen Species (ROS)
4.8. Determination of Glutathione and Ascorbic Acid
4.9. Enzymatic Activity and Gene Expression Analyses
4.10. Measurement of Relative Root Growth
4.11. Al Content Assay
4.12. Statistical Analysis and Reproducibility
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kochian, L.V.; Hoekenga, O.A.; Pineros, M.A. How do crop plants tolerate acid soils? Mechanisms of aluminum tolerance and phosphorous efficiency. Annu. Rev. Plant Biol. 2004, 55, 459–493. [Google Scholar] [CrossRef] [PubMed]
- Kochian, L.V. Cellular mechanisms of aluminum toxicity and resistance in plants. Annu. Rev. Plant Biol. 1995, 46, 237–260. [Google Scholar] [CrossRef]
- Ma, J.F. Role of organic acids in detoxification of aluminum in higher plants. Plant Cell Physiol. 2000, 41, 383–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, S.J.; Yang, J.L. Target sites of aluminum phytotoxicity. Biol. Plantarum 2005, 49, 321–331. [Google Scholar] [CrossRef]
- Shen, R.; Ma, J.F.; Kyo, M.; Iwashita, T. Compartmentation of aluminium in leaves of an Al-accumulator, Fagopyrum esculentum Moench. Planta 2002, 215, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Ryan, P.R.; Delhaize, E. Aluminium tolerance in plants and the complexing role of organic acids. Trends Plant Sci. 2001, 6, 273–278. [Google Scholar] [CrossRef]
- Ryan, P.R.; Delhaize, E.; Jones, D.L. Function and mechanism of organic anion exudation from plant roots. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 527–560. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kobayashi, Y.; Matsumoto, H. Lipid peroxidation is an early symptom triggered by aluminum, but not the primary cause of elongation inhibition in pea roots. Plant Physiol. 2001, 125, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Tabaldi, L.A.; Cargnelutti, D.; Gonçalves, J.F.; Pereira, L.B.; Castro, G.Y.; Maldaner, J.; Rauber, R.; Rossato, L.V.; Bisognin, D.A.; Schetinger, M.R.C. Oxidative stress is an early symptom triggered by aluminum in Al-sensitive potato plantlets. Chemosphere 2009, 76, 1402–1409. [Google Scholar] [CrossRef]
- Achary, V.M.; Jena, S.; Panda, K.K.; Panda, B.B. Aluminium induced oxidative stress and DNA damage in root cells of Allium cepa L. Ecotoxicol. Environ. Saf. 2008, 70, 300–310. [Google Scholar] [CrossRef]
- Kirch, H.H.; Bartels, D.; Wei, Y.; Schnable, P.S.; Wood, A.J. The ALDH gene superfamily of Arabidopsis. Trends Plant Sci. 2004, 9, 371–377. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Brocker, C.; Koppaka, V.; Chen, Y.; Jackson, B.C.; Matsumoto, A.; Thompson, D.C.; Vasiliou, V. Aldehyde dehydrogenases in cellular responses to oxidative/electrophilic stress. Free Radic. Biol. Med. 2013, 56, 89–101. [Google Scholar] [CrossRef] [Green Version]
- Møller, I.M.; Jensen, P.E.; Hansson, A. Oxidative modifications to cellular components in plants. Annu. Rev. Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef] [Green Version]
- Sunkar, R.; Bartels, D.; Kirch, H.H. Overexpression of a stress-inducible aldehyde dehydrogenase gene from Arabidopsis thaliana in transgenic plants improves stress tolerance. Plant. J. 2003, 35, 452–464. [Google Scholar] [CrossRef]
- Kotchoni, S.O.; Kuhns, C.; Ditzer, A.; Kirch, H.H.; Bartels, D. Over-expression of different aldehyde dehydrogenase genes in Arabidopsis thaliana confers tolerance to abiotic stress and protects plants against lipid peroxidation and oxidative stress. Plant. Cell Environ. 2006, 29, 1033–1048. [Google Scholar] [CrossRef]
- Sun, C.; Liu, L.; Yu, Y.; Liu, W.; Lu, L.; Jin, C.; Lin, X. Nitric oxide alleviates aluminum-induced oxidative damage through regulating the ascorbate-glutathione cycle in roots of wheat. J. Integr. Plant Biol. 2015, 57, 550–561. [Google Scholar] [CrossRef]
- Yu, Y.; Zhou, W.; Zhou, K.; Liu, W.; Liang, X.; Chen, Y.; Sun, D.; Lin, X. Polyamines modulate aluminum-induced oxidative stress differently by inducing or reducing H2O2 production in wheat. Chemosphere 2018, 212, 645–653. [Google Scholar] [CrossRef]
- Mittova, V.; Volokita, M.; Guy, M.; Tal, M. Activities of SOD and the ascorbate-glutathione cycle enzymes in subcellular compartments in leaves and roots of the cultivated tomato and its wild salt-tolerant relative Lycopersicon pennellii. Physiol. Plantarum 2000, 110, 42–51. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.; Zhang, J. Advances in the research on the AsA-GSH cycle in horticultural crops. Front. Agric. China 2010, 4, 84–90. [Google Scholar] [CrossRef]
- Kenichi, O. Glutathione-associated regulation of plant growth and stress responses. Antioxid. Redox Signal. 2005, 7, 973–981. [Google Scholar]
- Perozich, J.; Nicholas, H.; Wang, B.C.; Lindahl, R.; Hempel, J. Relationships within the aldehyde dehydrogenase extended family. Protein Sci. 1999, 8, 137–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frommel, J.; Koncitikova, R.; Kopecny, D.; Soural, M.; Sebela, M. Oxidation of imidazole- and pyrazole-derived aldehydes by plant aldehyde dehydrogenases from the family 2 and 10. Chem. Biol. Interact. 2019, 304, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Bartels, D. Targeting detoxification pathways: An efficient approach to obtain plants with multiple stress tolerance? Trends Plant Sci. 2001, 6, 284–286. [Google Scholar] [CrossRef]
- Zhou, M.L.; Zhang, Q.; Zhou, M.; Qi, L.P.; Yang, X.B.; Zhang, K.X.; Pang, J.F.; Zhu, X.M.; Shao, J.R.; Tang, Y.X.; et al. Aldehyde dehydrogenase protein superfamily in maize. Funct. Integr. Genom. 2012, 12, 683–691. [Google Scholar] [CrossRef]
- An, X.; Duan, F.Y.; Guo, S.; Chen, F.J.; Yuan, L.X.; Gu, R.L. Transcriptional regulation of expression of the maize aldehyde dehydrogenase 7 gene (ZmALDH7B6) in response to abiotic stresses. J. Integr. Agr. 2014, 13, 1900–1908. [Google Scholar] [CrossRef] [Green Version]
- Ke, Y.G.; Yuan, M.; Liu, H.B.; Hui, S.G.; Qin, X.F.; Chen, J.; Zhang, Q.L.; LI, X.H.; Xiao, J.H.; Zhang, Q.F.; et al. The versatile functions of OsALDH2B1 provide a genic basis for growth-defense trade-offs in rice. Proc. Natl. Acad. Sci. USA 2020, 117, 3867–3873. [Google Scholar] [CrossRef]
- Inostroza-Blancheteau, C.; Reyes-Diaz, M.; Aquea, F.; Nunes-Nesi, A.; Alberdi, M.; Arce-Johnson, P. Biochemical and molecular changes in response to aluminium-stress in highbush blueberry (Vaccinium corymbosum L.). Plant Physiol. Biochem. 2011, 49, 1005–1012. [Google Scholar] [CrossRef]
- Xu, L.M.; Liu, C.; Cui, B.M.; Wang, N.; Zhao, Z.; Zhou, L.N.; Huang, K.F.; Ding, J.Z.; Du, H.M.; Jiang, W.; et al. Transcriptomic responses to aluminum (Al) stress in maize. J. Integr. Agr. 2018, 17, 1946–1958. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.M.; Liu, W.; Cui, B.M.; Wang, N.; Ding, J.Z.; Liu, C.; Gao, S.B.; Zhang, S.Z. Aluminium tolerance assessment of 141 maize germplasms in a solution culture. Univers. J. Agric. Res. 2017, 5, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Šimonovičová, M.; Huttová, J.; Mistrík, I.; Široká, B.; Tamás, L. Root growth inhibition by aluminum is probably caused by cell death due to peroxidase-mediated hydrogen peroxide production. Protoplasma 2004, 224, 91–98. [Google Scholar] [CrossRef]
- Chaudhuri, P.; Rashid, N.; Thapliyal, C. Osmolyte System and Its Biological Significance. In Cellular Osmolytes; Rajendrakumar, S.L., Dar, T., Eds.; Springer: Singapore, 2017; pp. 1–34. [Google Scholar]
- Hasanuzzaman, M.; Anee, T.I.; Bhuiyan, T.F.; Nahar, K.; Fujita, M. Emerging role of osmolytes in enhancing abiotic stress tolerance in rice. In Advances in Rice Research for Abiotic Stress Tolerance; Hasanuzzaman, M., Nahar, K., Fujita, M., Biswas, J.K., Eds.; Woodhead Publishing: New York, NY, USA, 2019; pp. 677–708. [Google Scholar]
- Gao, C.; Han, B. Evolutionary and expression study of the aldehyde dehydrogenase (ALDH) gene superfamily in rice (Oryza sativa). Gene 2009, 431, 86–94. [Google Scholar] [CrossRef]
- Jimenez-Lopez, J.C.; Gachomo, E.W.; Seufferheld, M.J.; Kotchoni, S.O. The maize ALDH protein superfamily: Linking structural features to functional specificities. BMC Struct. Biol. 2010, 10, 43. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Kobayashi, Y.; Devi, S.R.; Rikiishi, S.; Matsumoto, H. Aluminum toxicity is associated with mitochondrial dysfunction and the production of reactive oxygen species in plant cells. Plant Physiol. 2002, 128, 63–72. [Google Scholar] [CrossRef]
- Chowra, U.; Yanase, E.; Koyama, H.; Panda, S.K. Aluminium-induced excessive ROS causes cellular damage and metabolic shifts in black gram Vigna mungo (L.) Hepper. Protoplasma 2017, 254, 293–302. [Google Scholar] [CrossRef]
- Richards, K.D.; Schott, E.J.; Sharma, Y.K.; Davis, K.R.; Gardner, R.C. Aluminum induces oxidative stress genes in Arabidopsis thaliana. Plant Physiol. 1998, 116, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Liu, L.; Zhou, W.; Lu, L.; Jin, C.; Lin, X. Aluminum Induces Distinct Changes in the Metabolism of Reactive Oxygen and Nitrogen Species in the Roots of Two Wheat Genotypes with Different Aluminum Resistance. J. Age Food Chem. 2017, 65, 9419–9427. [Google Scholar] [CrossRef]
- Chen, Y.; Mehta, G.; Vasiliou, V. Antioxidant defenses in the ocular surface. Ocul. Surf. 2009, 7, 176–185. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, S.M.; Andrade, M.O.; Gomes, A.P.; Damatta, F.M.; Baracat-Pereira, M.C.; Fontes, E.P. Arabidopsis and tobacco plants ectopically expressing the soybean antiquitin-like ALDH7 gene display enhanced tolerance to drought, salinity, and oxidative stress. J. Exp. Bot. 2006, 57, 1909–1918. [Google Scholar] [CrossRef] [Green Version]
- Jing, X.Q.; Zhou, M.R.; Nie, X.M.; Zhang, L.; Shi, P.T.; Shalmani, A.; Miao, H.; Li, W.Q.; Liu, W.T.; Chen, K.M. OsGSTU6 Contributes to Cadmium Stress Tolerance in Rice by Involving in Intracellular ROS Homeostasis. J. Plant Growth Regul. 2021, 40, 945–961. [Google Scholar] [CrossRef]
- Ezaki, B.; Gardner, R.C.; Ezaki, Y.; Matsumoto, H. Expression of aluminum-induced genes in transgenic Arabidopsis plants can ameliorate aluminum stress and/or oxidative stress. Plant Physiol. 2000, 122, 657–665. [Google Scholar] [CrossRef] [Green Version]
- Panda, S.K.; Sahoo, L.; Katsuhara, M.; Matsumoto, H. Overexpression of alternative oxidase gene confers aluminum tolerance by altering the respiratory capacity and the response to oxidative stress in tobacco cells. Mol. Biotechnol. 2013, 54, 551–563. [Google Scholar] [CrossRef]
- Basu, U.; Good, A.G.; Taylor, G.J. Transgenic Brassica napus plants overexpressing aluminum-induced mitochondrial manganese superoxide dismutase cDNA are resistant to aluminum. Plant Cell Environ. 2001, 24, 1269–1278. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.S.; Yang, Z.L.; How, J.Y.; Xu, H.N.; Chen, L.M.; Li, K.Z. Overexpression of a peroxidase gene (AtPrx64) of Arabidopsis thaliana in tobacco improves plant’s tolerance to aluminum stress. Plant Mol. Biol. 2017, 95, 157–168. [Google Scholar] [CrossRef]
- Du, H.; Huang, Y.; Qu, M.; Li, Y.; Hu, X.; Yang, W.; Li, H.; He, W.; Ding, J.; Liu, C.; et al. A maize ZmAT6 gene confers aluminum tolerance via reactive oxygen species scavenging. Front. Plant Sci. 2020, 11, 1016. [Google Scholar] [CrossRef]
- Irany, R.P.; Anny, C.D.L.; Tatiane, L.P.; Raquel, A.A.; Maria, D.C.P.B. Gene expression and antioxidant enzymatic activity in passion fruit exposed to aluminum. Afr. J. Agric. Res. 2018, 13, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Patade, V.Y.; Bhargava, S.; Suprasanna, P. Salt and drought tolerance of sugarcane under iso-osmotic salt and water stress: Growth, osmolytes accumulation, and antioxidant defense. J. Plant Interact. 2011, 6, 275–282. [Google Scholar] [CrossRef] [Green Version]
- Pirzadah, T.B.; Malik, B.; Tahir, I.; Rehman, R.U.; Hakeem, K.R.; Alharby, H.F. Aluminium stress modulates the osmolytes and enzyme defense system in Fagopyrum species. Plant Physiol. Biochem. 2019, 144, 178–186. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Proline: A key player in plant abiotic stress tolerance. Biol. Plantarum 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Du, H.; Ryan, P.R.; Liu, C.; Li, H.; Hu, W.; Yan, W.; Huang, Y.; He, W.; Luo, B.; Zhang, X.; et al. ZmMATE6 from maize encodes a citrate transporter that enhances aluminum tolerance in transgenic Arabidopsis thaliana. Plant Sci. 2021, 311, 111016. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Du, H.; Huang, K.; Chen, X.; Liu, T.; Gao, S.; Liu, H.; Tang, Q.; Rong, T.; Zhang, S. Identification, and functional and expression analyses of the CorA/MRS2/MGT-type magnesium transporter family in maize. Plant Cell Physiol. 2016, 57, 1153–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clough, S.J.; Bent, A.F. Floral dip: A simplifed method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. Cell Mol. Biol. 1998, 16, 735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Wang, N.; Ding, J.; Liu, C.; Du, H.; Huang, K.; Cao, M.; Lu, Y.; Gao, S.; Zhang, S. The maize CorA/MRS2/MGT-type Mg transporter, ZmMGT10, responses to magnesium deficiency and confers low magnesium tolerance in transgenic Arabidopsis. Plant Mol. Biol. 2017, 95, 269–278. [Google Scholar] [CrossRef]
- Baker, C.J.; Mock, N.M. An improved methods for monitoring cell death in cell suspension and leaf disc assays using evans blue. Plant Cell Tissue Org. Cult. 1994, 39, 7–12. [Google Scholar] [CrossRef]
- Elstner, E.F.; Heupel, A. Inhibition of nitrite formation from hydroxylammoniumchloride: A simple assay for superoxide dismutase. Anal. Biochem. 1976, 70, 616–620. [Google Scholar] [CrossRef]
- Hodges, D.; Andrews, C.; Johnson, D.; Hamilton, R. Antioxidant compound responses to chilling stress in differentially sensitive inbred maize lines. Plant Physiol. 1996, 98, 685–692. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Foyer, C.H.; Halliwell, B. The presence of glutathione and glutathione reductase in chloroplasts: A proposed role in ascorbic acid metabolism. Planta 1976, 133, 21–25. [Google Scholar] [CrossRef]
- Ribeiro, A.P.; de Souza, W.R.; Martins, P.K.; Vinecky, F.; Duarte, K.E.; Basso, M.F.; da Cunha, B.; Campanha, R.B.; de Oliveira, P.A.; Centeno, D.C.; et al. Overexpression of BdMATE gene improves aluminum tolerance in Setaria viridis. Front. Plant Sci. 2017, 8, 865. [Google Scholar] [CrossRef] [Green Version]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, H.-M.; Liu, C.; Jin, X.-W.; Du, C.-F.; Yu, Y.; Luo, S.; He, W.-Z.; Zhang, S.-Z. Overexpression of the Aldehyde Dehydrogenase Gene ZmALDH Confers Aluminum Tolerance in Arabidopsis thaliana. Int. J. Mol. Sci. 2022, 23, 477. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010477
Du H-M, Liu C, Jin X-W, Du C-F, Yu Y, Luo S, He W-Z, Zhang S-Z. Overexpression of the Aldehyde Dehydrogenase Gene ZmALDH Confers Aluminum Tolerance in Arabidopsis thaliana. International Journal of Molecular Sciences. 2022; 23(1):477. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010477
Chicago/Turabian StyleDu, Han-Mei, Chan Liu, Xin-Wu Jin, Cheng-Feng Du, Yan Yu, Shuai Luo, Wen-Zhu He, and Su-Zhi Zhang. 2022. "Overexpression of the Aldehyde Dehydrogenase Gene ZmALDH Confers Aluminum Tolerance in Arabidopsis thaliana" International Journal of Molecular Sciences 23, no. 1: 477. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010477