
The QseEF Two-Component System-GlmY Small RNA Regulatory Pathway Controls Swarming in Uropathogenic Proteus mirabilis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
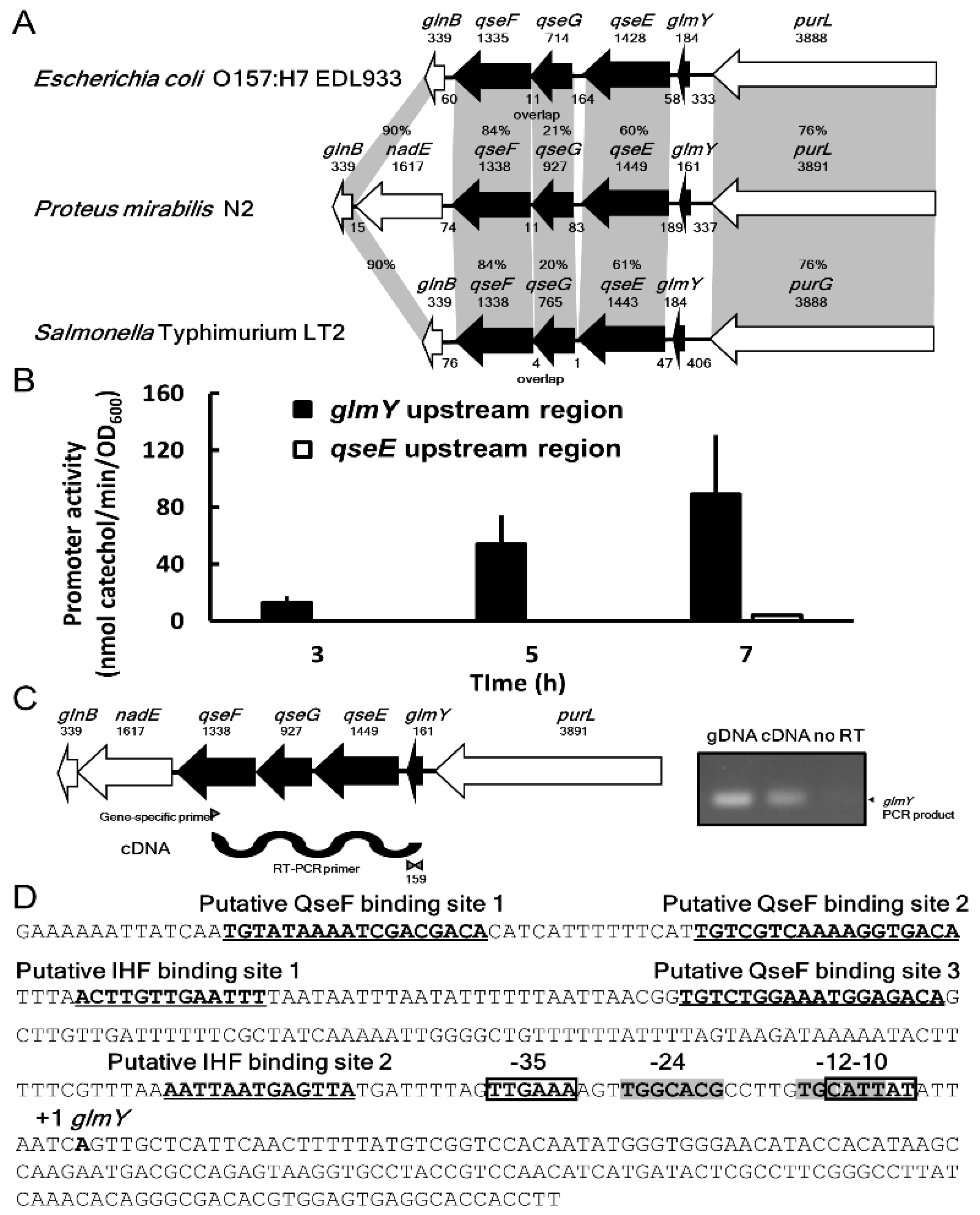
2.1. Identification of P. mirabilis qseEGF Gene Locus
2.2. Identification of P. mirabilis glmY and Co-Transcription of glmY, qseE, qseG and qseF
2.3. Phenotypic Traits of glmY and qse Mutants
2.4. Downregulation of glmYqseEGF Operon by qseF Deletion
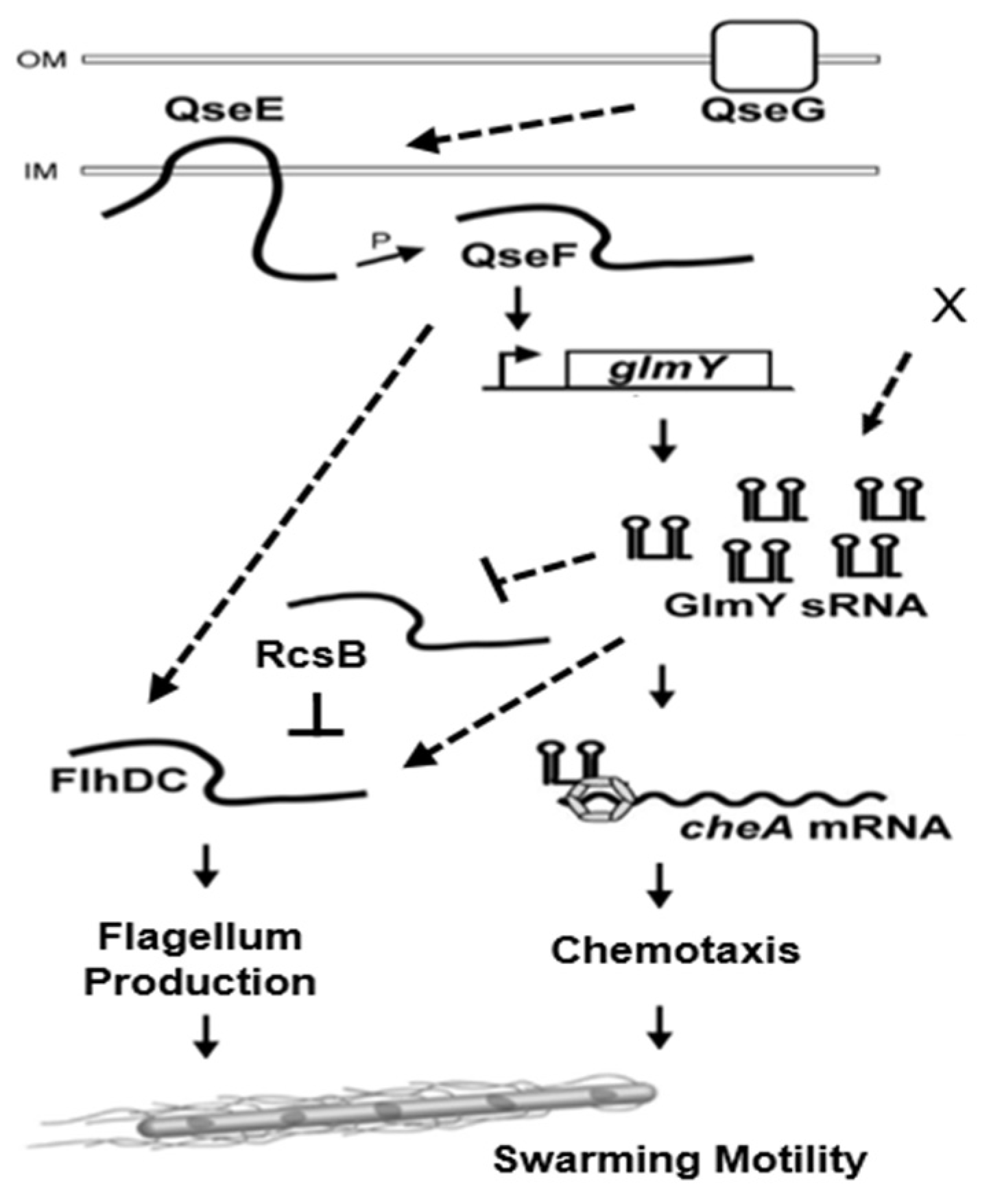
2.5. Altered Expression of flhDC, rcsB and cheA in qseF and glmY Mutants
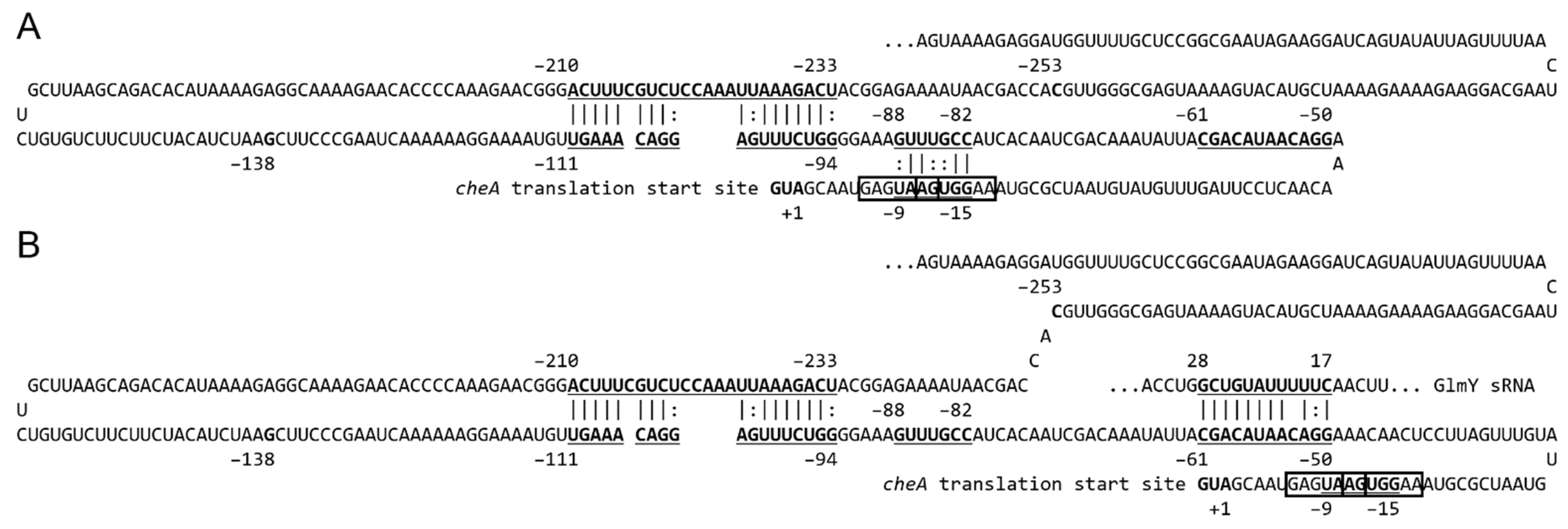
2.6. GlmY Activates cheA Expression at the Post-Transcriptional Level
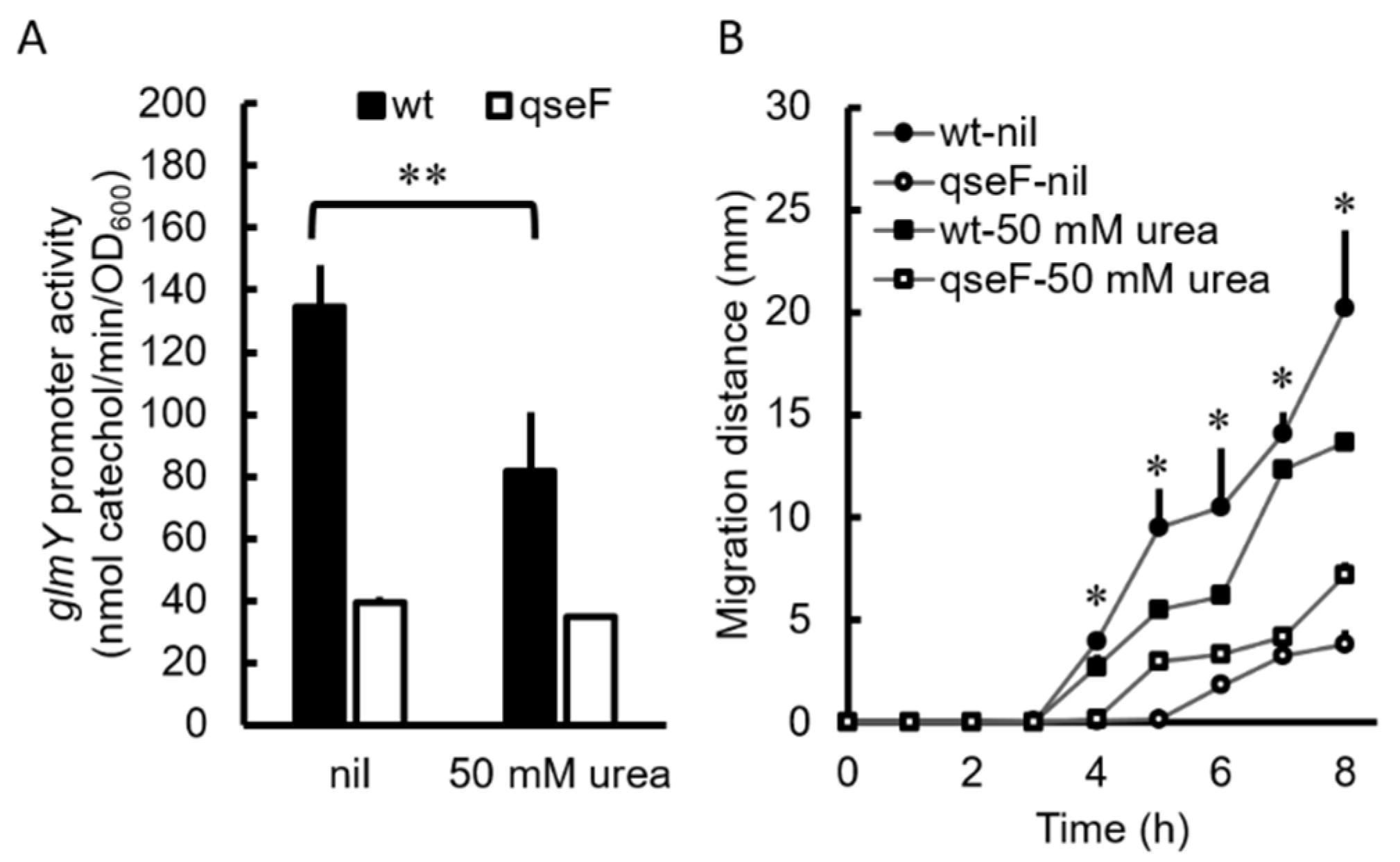
2.7. Urea Inhibited Promoter Activity of the glmYqseEGF Operon in P. mirabilis
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Plasmids, Reagents and Growth Conditions
4.2. Construction of P. mirabilis Mutants and Complemented Strains
4.3. Swarming and Swimming Assays
4.4. Measurement of the Haemolysin Activity and Cell Length
4.5. Measurement of the Flagellin Level
4.6. Transmission Electron Microscopy
4.7. Real Time Reverse Transcription PCR (RT-PCR)
4.8. Transcriptional and Translational Reporter Assays
4.9. Site-Directed Mutagenesis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Norsworthy, A.N.; Pearson, M.M. From Catheter to Kidney Stone: The Uropathogenic Lifestyle of Proteus mirabilis. Trends Microbiol. 2017, 25, 304–315. [Google Scholar] [CrossRef] [Green Version]
- Jansen, A.M.; Lockatell, V.; Johnson, D.E.; Mobley, H.L. Mannose-resistant Proteus-like fimbriae are produced by most Proteus mirabilis strains infecting the urinary tract, dictate the in vivo localization of bacteria, and contribute to biofilm formation. Infect. Immun. 2004, 72, 7294–7305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, Y.L.; Chien, H.F.; Huang, K.T.; Lin, W.Y.; Liaw, S.J. cAMP receptor protein regulates mouse colonization, motility, fimbria-mediated adhesion, and stress tolerance in uropathogenic Proteus mirabilis. Sci. Rep. 2017, 7, 7282. [Google Scholar] [CrossRef] [Green Version]
- Nielubowicz, G.R.; Mobley, H.L. Host-pathogen interactions in urinary tract infection. Nat. Rev. Urol. 2010, 7, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Armbruster, C.E.; Mobley, H.L. Merging mythology and morphology: The multifaceted lifestyle of Proteus mirabilis. Nat. Rev. Microbiol. 2012, 10, 743–754. [Google Scholar] [CrossRef]
- Jacobsen, S.M.; Stickler, D.J.; Mobley, H.L.; Shirtliff, M.E. Complicated catheter-associated urinary tract infections due to Escherichia coli and Proteus mirabilis. Clin. Microbiol. Rev. 2008, 21, 26–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser, G.M.; Hughes, C. Swarming motility. Curr. Opin. Microbiol. 1999, 2, 630–635. [Google Scholar] [CrossRef]
- Clemmer, K.M.; Rather, P.N. Regulation of flhDC expression in Proteus mirabilis. Res. Microbiol. 2007, 158, 295–302. [Google Scholar] [CrossRef]
- Gottesman, S.; Storz, G. Bacterial small RNA regulators: Versatile roles and rapidly evolving variations. Cold Spring Harb. Perspect. Biol. 2011, 3, a003798. [Google Scholar] [CrossRef] [Green Version]
- Waters, L.S.; Storz, G. Regulatory RNAs in bacteria. Cell 2009, 136, 615–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillier, M.; Gottesman, S.; Storz, G. Modulating the outer membrane with small RNAs. Genes Dev. 2006, 20, 2338–2348. [Google Scholar] [CrossRef] [Green Version]
- Bak, G.; Lee, J.; Suk, S.; Kim, D.; Young Lee, J.; Kim, K.S.; Choi, B.S.; Lee, Y. Identification of novel sRNAs involved in biofilm formation, motility, and fimbriae formation in Escherichia coli. Sci. Rep. 2015, 5, 15287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulesus, R.R.; Diaz-Perez, K.; Slechta, E.S.; Eto, D.S.; Mulvey, M.A. Impact of the RNA chaperone Hfq on the fitness and virulence potential of uropathogenic Escherichia coli. Infect. Immun. 2008, 76, 3019–3026. [Google Scholar] [CrossRef] [Green Version]
- Chao, Y.; Vogel, J. The role of Hfq in bacterial pathogens. Curr. Opin. Microbiol. 2010, 13, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.C.; Chien, H.F.; Tsai, Y.L.; Liu, M.C.; Liaw, S.J. The RNA chaperone Hfq is involved in stress tolerance and virulence in uropathogenic Proteus mirabilis. PLoS ONE 2014, 9, e85626. [Google Scholar] [CrossRef]
- Vogel, J.; Luisi, B.F. Hfq and its constellation of RNA. Nat. Rev. Microbiol. 2011, 9, 578–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valverde, C.; Haas, D. Small RNAs controlled by two-component systems. Adv. Exp. Med. Biol. 2008, 631, 54–79. [Google Scholar] [PubMed]
- Vogel, J. A rough guide to the non-coding RNA world of Salmonella. Mol. Microbiol. 2009, 71, 1–11. [Google Scholar] [CrossRef]
- Overgaard, M.; Kallipolitis, B.; Valentin-Hansen, P. Modulating the bacterial surface with small RNAs: A new twist on PhoP/Q-mediated lipopolysaccharide modification. Mol. Microbiol. 2009, 74, 1289–1294. [Google Scholar] [CrossRef]
- Coornaert, A.; Lu, A.; Mandin, P.; Springer, M.; Gottesman, S.; Guillier, M. MicA sRNA links the PhoP regulon to cell envelope stress. Mol. Microbiol. 2010, 76, 467–479. [Google Scholar] [CrossRef] [Green Version]
- Reading, N.C.; Torres, A.G.; Kendall, M.M.; Hughes, D.T.; Yamamoto, K.; Sperandio, V. A novel two-component signaling system that activates transcription of an enterohemorrhagic Escherichia coli effector involved in remodeling of host actin. J. Bacteriol. 2007, 189, 2468–2476. [Google Scholar] [CrossRef] [Green Version]
- Njoroge, J.; Sperandio, V. Enterohemorrhagic Escherichia coli virulence regulation by two bacterial adrenergic kinases, QseC and QseE. Infect. Immun. 2012, 80, 688–703. [Google Scholar] [CrossRef] [Green Version]
- Moreira, C.G.; Sperandio, V. Interplay between the QseC and QseE bacterial adrenergic sensor kinases in Salmonella enterica serovar Typhimurium pathogenesis. Infect. Immun. 2012, 80, 4344–4353. [Google Scholar] [CrossRef] [Green Version]
- Moreira, C.G.; Russell, R.; Mishra, A.A.; Narayanan, S.; Ritchie, J.M.; Waldor, M.K.; Curtis, M.M.; Winter, S.E.; Weinshenker, D.; Sperandio, V. Bacterial adrenergic sensors regulate virulence of enteric pathogens in the gut. mBio 2016, 7, e00826-16. [Google Scholar] [CrossRef] [Green Version]
- Cameron, E.A.; Gruber, C.C.; Ritchie, J.M.; Waldor, M.K.; Sperandio, V. The QseG lipoprotein impacts the virulence of enterohemorrhagic Escherichia coli and Citrobacter rodentium and regulates flagellar phase variation in Salmonella enterica serovar Typhimurium. Infect. Immun. 2018, 86, e00936-17. [Google Scholar] [CrossRef] [Green Version]
- Reading, N.C.; Rasko, D.A.; Torres, A.G.; Sperandio, V. The two-component system QseEF and the membrane protein QseG link adrenergic and stress sensing to bacterial pathogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 5889–5894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gopel, Y.; Luttmann, D.; Heroven, A.K.; Reichenbach, B.; Dersch, P.; Gorke, B. Common and divergent features in transcriptional control of the homologous small RNAs GlmY and GlmZ in Enterobacteriaceae. Nucleic Acids Res. 2011, 39, 1294–1309. [Google Scholar] [CrossRef] [Green Version]
- Lustri, B.C.; Sperandio, V.; Moreira, C.G. Bacterial chat: Intestinal metabolites and signals in host-microbiota-pathogen interactions. Infect. Immun. 2017, 85, e00476-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruber, C.C.; Sperandio, V. Posttranscriptional control of microbe-induced rearrangement of host cell actin. mBio 2014, 5, e01025-13. [Google Scholar] [CrossRef] [Green Version]
- Bennett, A.M.; Shippy, D.C.; Eakley, N.; Okwumabua, O.; Fadl, A.A. Functional characterization of glucosamine-6-phosphate synthase (GlmS) in Salmonella enterica serovar Enteritidis. Arch. Microbiol. 2016, 198, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Reichenbach, B.; Gopel, Y.; Gorke, B. Dual control by perfectly overlapping sigma 54- and sigma 70- promoters adjusts small RNA GlmY expression to different environmental signals. Mol. Microbiol. 2009, 74, 1054–1070. [Google Scholar] [CrossRef]
- Howery, K.E.; Clemmer, K.M.; Rather, P.N. The Rcs regulon in Proteus mirabilis: Implications for motility, biofilm formation, and virulence. Curr. Genet. 2016, 62, 775–789. [Google Scholar] [CrossRef]
- Liu, M.C.; Lin, S.B.; Chien, H.F.; Wang, W.B.; Yuan, Y.H.; Hsueh, P.R.; Liaw, S.J. 10′(Z),13′(E)-heptadecadienylhydroquinone inhibits swarming and virulence factors and increases polymyxin B susceptibility in Proteus mirabilis. PLoS ONE 2012, 7, e45563. [Google Scholar] [CrossRef]
- Burkart, M.; Toguchi, A.; Harshey, R.M. The chemotaxis system, but not chemotaxis, is essential for swarming motility in Escherichia coli. Proc. Natl. Acad. Sci. USA 1998, 95, 2568–2573. [Google Scholar] [CrossRef] [Green Version]
- Podkaminski, D.; Vogel, J. Small RNAs promote mRNA stability to activate the synthesis of virulence factors. Mol. Microbiol. 2010, 78, 1327–1331. [Google Scholar] [CrossRef] [PubMed]
- Verstraeten, N.; Braeken, K.; Debkumari, B.; Fauvart, M.; Fransaer, J.; Vermant, J.; Michiels, J. Living on a surface: Swarming and biofilm formation. Trends Microbiol. 2008, 16, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Valentin-Hansen, P.; Johansen, J.; Rasmussen, A.A. Small RNAs controlling outer membrane porins. Curr. Opin. Microbiol. 2007, 10, 152–155. [Google Scholar] [CrossRef]
- Gopel, Y.; Papenfort, K.; Reichenbach, B.; Vogel, J.; Gorke, B. Targeted decay of a regulatory small RNA by an adaptor protein for RNase E and counteraction by an anti-adaptor RNA. Genes Dev. 2013, 27, 552–564. [Google Scholar] [CrossRef] [Green Version]
- Gopel, Y.; Khan, M.A.; Gorke, B. Menage a trois: Post-transcriptional control of the key enzyme for cell envelope synthesis by a base-pairing small RNA, an RNase adaptor protein, and a small RNA mimic. RNA Biol. 2014, 11, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Urban, J.H.; Vogel, J. Two seemingly homologous noncoding RNAs act hierarchically to activate glmS mRNA translation. PLoS Biol. 2008, 6, e64. [Google Scholar] [CrossRef]
- Klein, G.; Raina, S. Regulated assembly of LPS, its structural alterations and cellular response to LPS defects. Int. J. Mol. Sci. 2019, 20, 356. [Google Scholar] [CrossRef] [Green Version]
- Gruber, C.C.; Sperandio, V. Global analysis of posttranscriptional regulation by GlmY and GlmZ in enterohemorrhagic Escherichia coli O157:H7. Infect. Immun. 2015, 83, 1286–1295. [Google Scholar] [CrossRef] [Green Version]
- Urban, J.H.; Papenfort, K.; Thomsen, J.; Schmitz, R.A.; Vogel, J. A conserved small RNA promotes discoordinate expression of the glmUS operon mRNA to activate GlmS synthesis. J. Mol. Biol. 2007, 373, 521–528. [Google Scholar] [CrossRef]
- De Lay, N.; Gottesman, S. A complex network of small non-coding RNAs regulate motility in Escherichia coli. Mol. Microbiol. 2012, 86, 524–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNamara, B.P.; Wolfe, A.J. Coexpression of the long and short forms of CheA, the chemotaxis histidine kinase, by members of the family Enterobacteriaceae. J. Bacteriol. 1997, 179, 1813–1818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Göpel, Y.; Görke, B. Interaction of lipoprotein QseG with sensor kinase QseE in the periplasm controls the phosphorylation state of the two-component system QseE/QseF in Escherichia coli. PLoS Genet. 2018, 14, e1007547. [Google Scholar] [CrossRef] [PubMed]
- Reading, N.C.; Rasko, D.; Torres, A.G.; Sperandio, V. A transcriptome study of the QseEF two-component system and the QseG membrane protein in enterohaemorrhagic Escherichia coli O157:H7. Microbiology 2010, 156, 1167–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liaw, S.J.; Lai, H.C.; Wang, W.B. Modulation of swarming and virulence by fatty acids through the RsbA protein in Proteus mirabilis. Infect. Immun. 2004, 72, 6836–6845. [Google Scholar] [CrossRef] [Green Version]
- Kaur, K.; Kumari, P.; Sharma, S.; Sehgal, S.; Tyagi, J.S. DevS/DosS sensor is bifunctional and its phosphatase activity precludes aerobic DevR/DosR regulon expression in Mycobacterium tuberculosis. FEBS J. 2016, 283, 2949–2962. [Google Scholar] [CrossRef] [Green Version]
- Schaffer, J.N.; Pearson, M.M. Proteus mirabilis and urinary tract infections. Microbiol. Spectr. 2015, 3, 1–39. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, W.-Y.; Lee, Y.-J.; Yu, P.-H.; Tsai, Y.-L.; She, P.-Y.; Li, T.-S.; Liaw, S.-J. The QseEF Two-Component System-GlmY Small RNA Regulatory Pathway Controls Swarming in Uropathogenic Proteus mirabilis. Int. J. Mol. Sci. 2022, 23, 487. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010487
Lin W-Y, Lee Y-J, Yu P-H, Tsai Y-L, She P-Y, Li T-S, Liaw S-J. The QseEF Two-Component System-GlmY Small RNA Regulatory Pathway Controls Swarming in Uropathogenic Proteus mirabilis. International Journal of Molecular Sciences. 2022; 23(1):487. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010487
Chicago/Turabian StyleLin, Wen-Yuan, Yuan-Ju Lee, Ping-Hung Yu, Yi-Lin Tsai, Pin-Yi She, Tzung-Shian Li, and Shwu-Jen Liaw. 2022. "The QseEF Two-Component System-GlmY Small RNA Regulatory Pathway Controls Swarming in Uropathogenic Proteus mirabilis" International Journal of Molecular Sciences 23, no. 1: 487. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010487