
Genome-Wide Characterization and Expression Analysis of KH Family Genes Response to ABA and SA in Arabidopsis thaliana


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification and Characterization of KH Gene Family in Arabidopsis thaliana
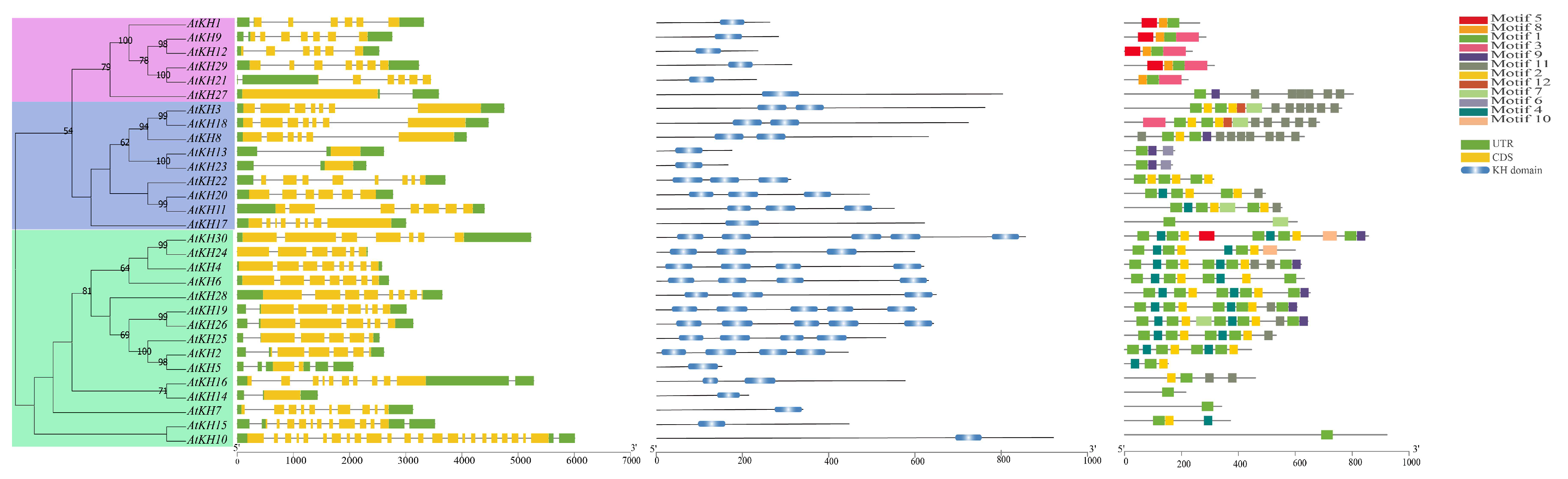
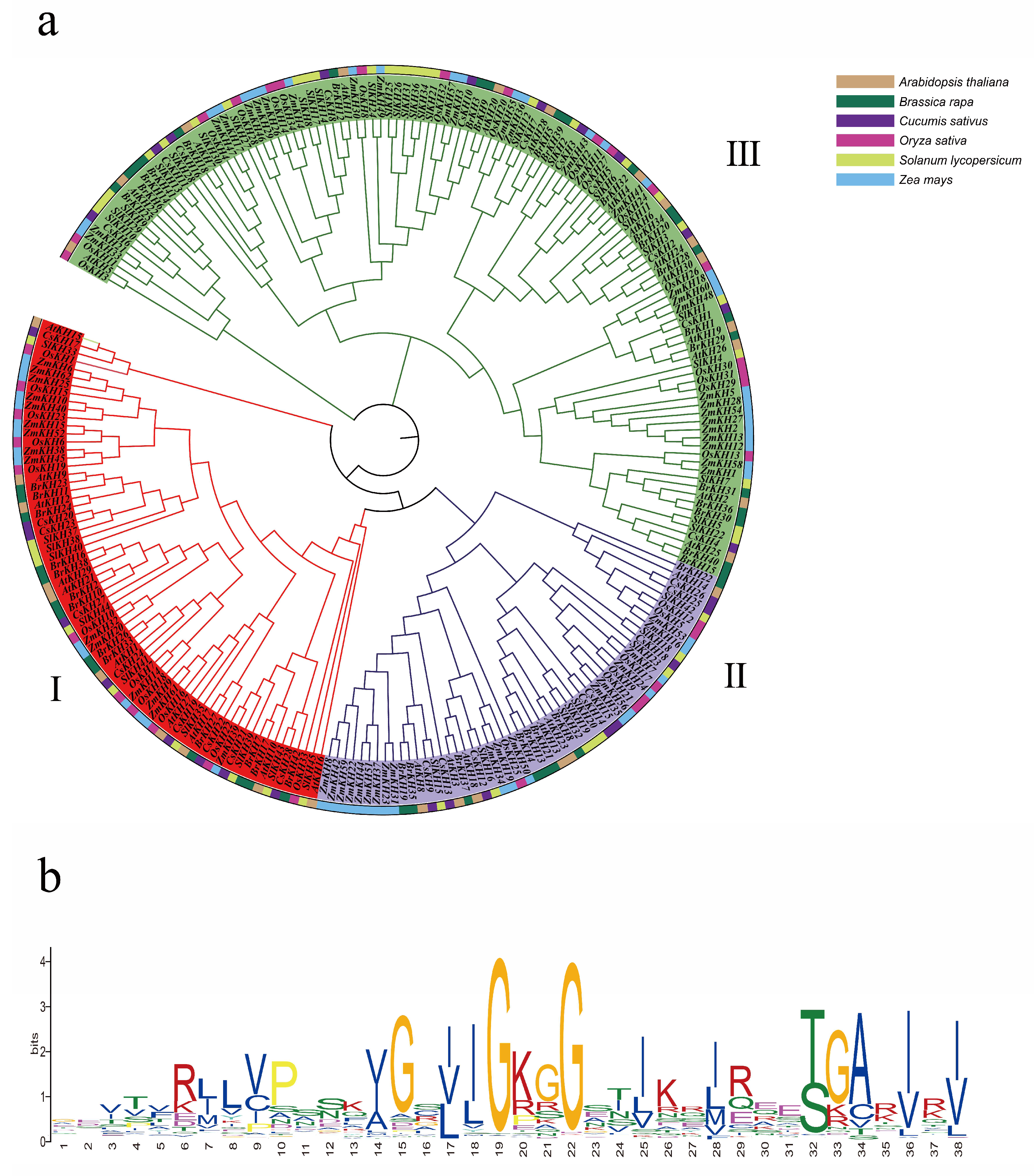
2.2. Phylogenetic Analysis of KH Family
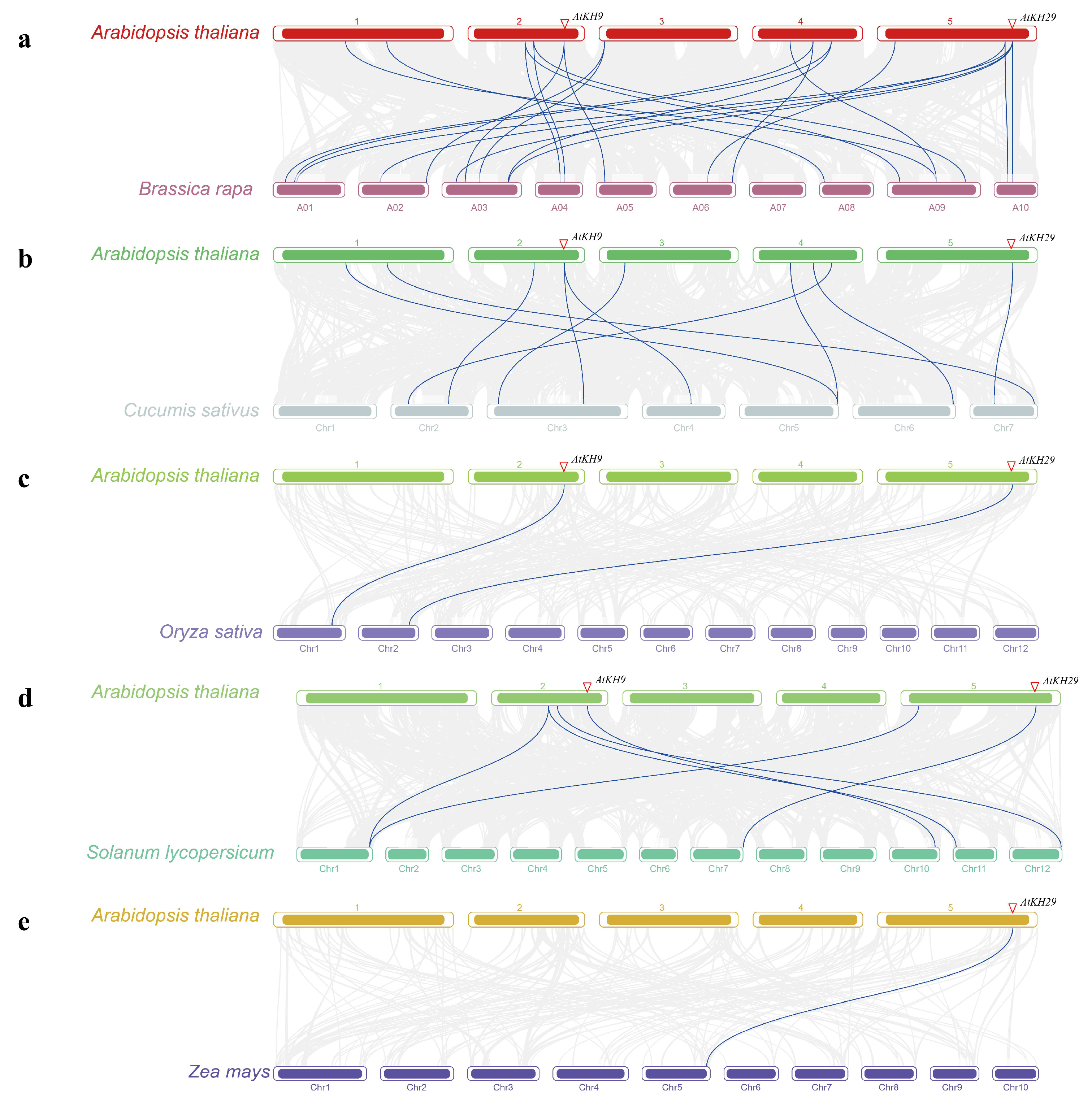
2.3. Synteny Analysis of KH Family
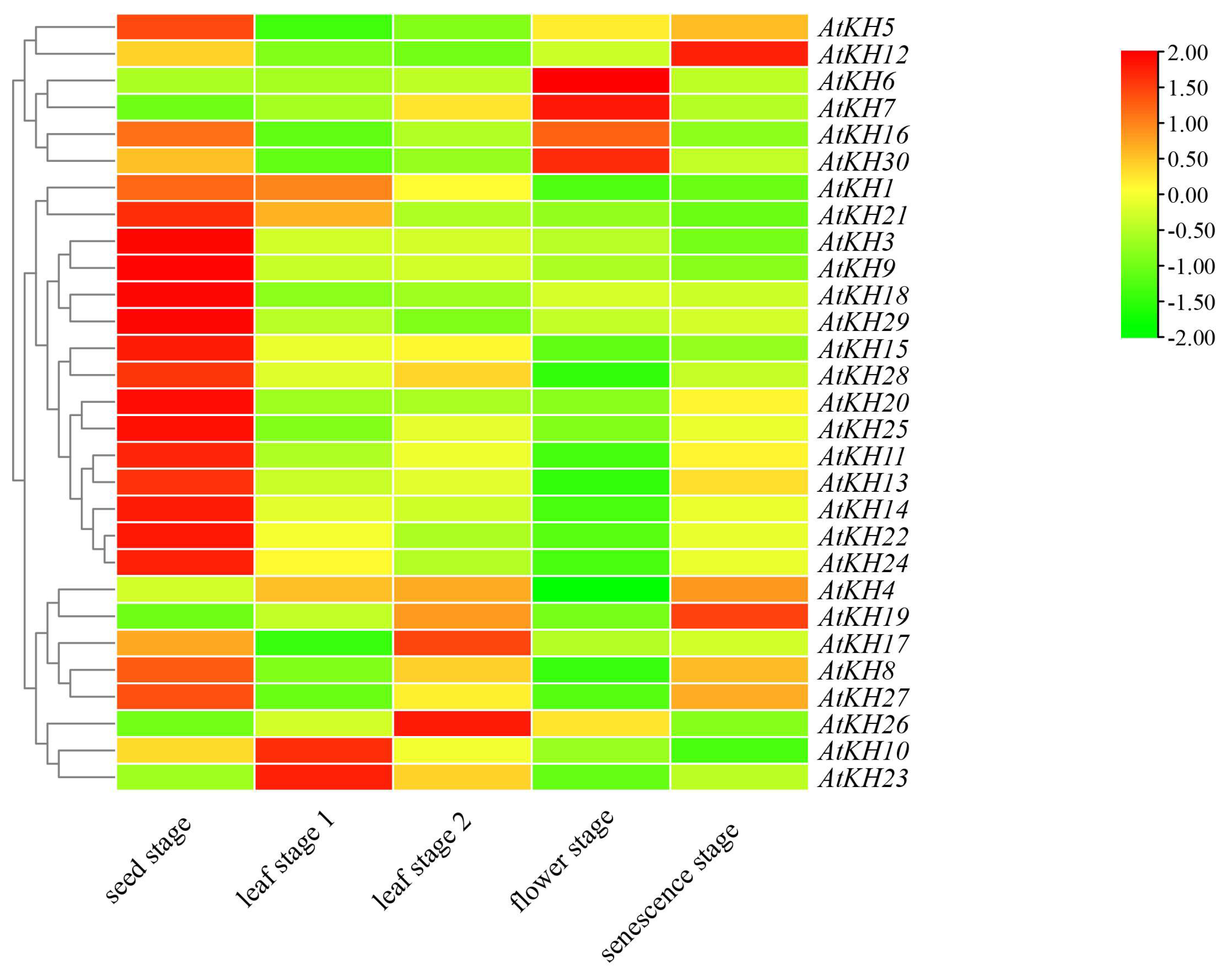
2.4. Expression of KH Family Genes in Different Stages during Growth and Development
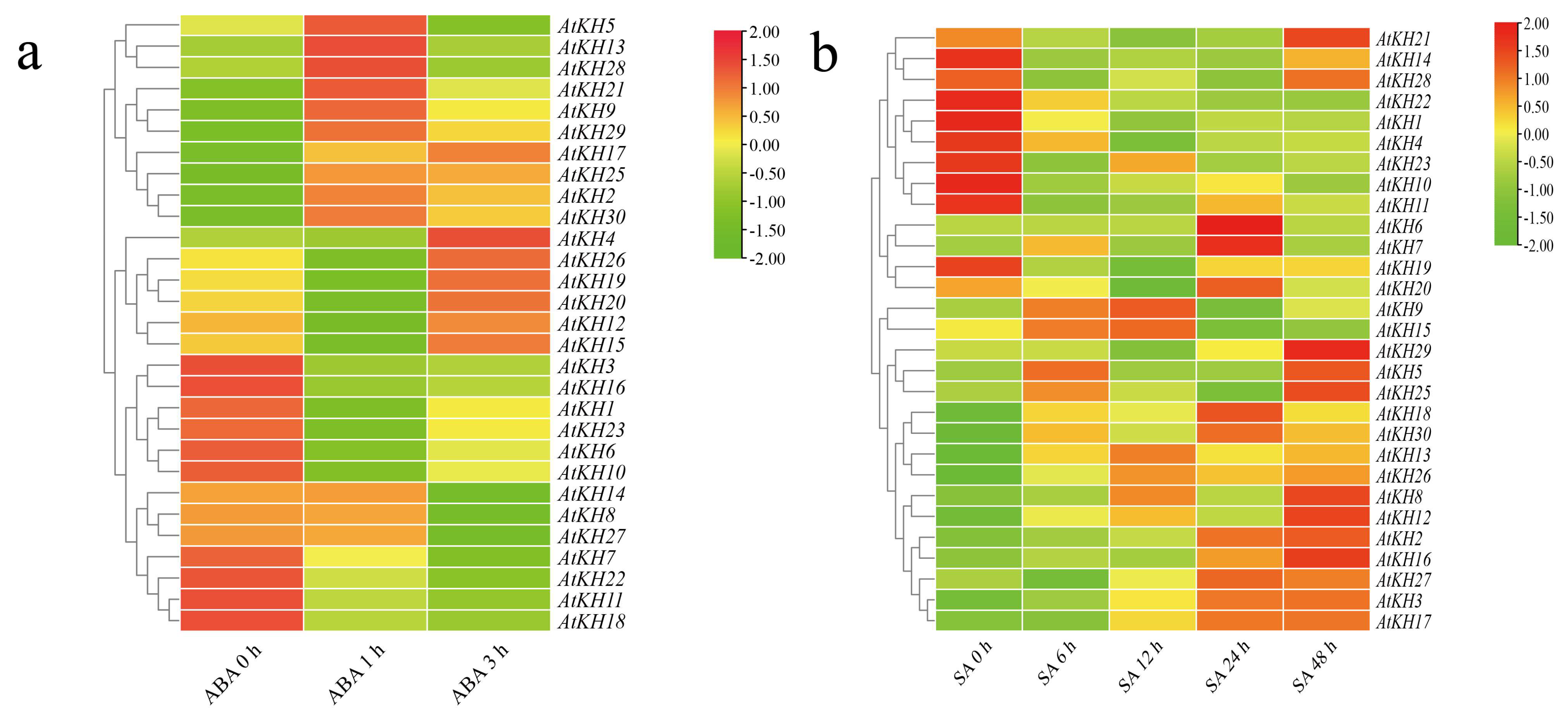
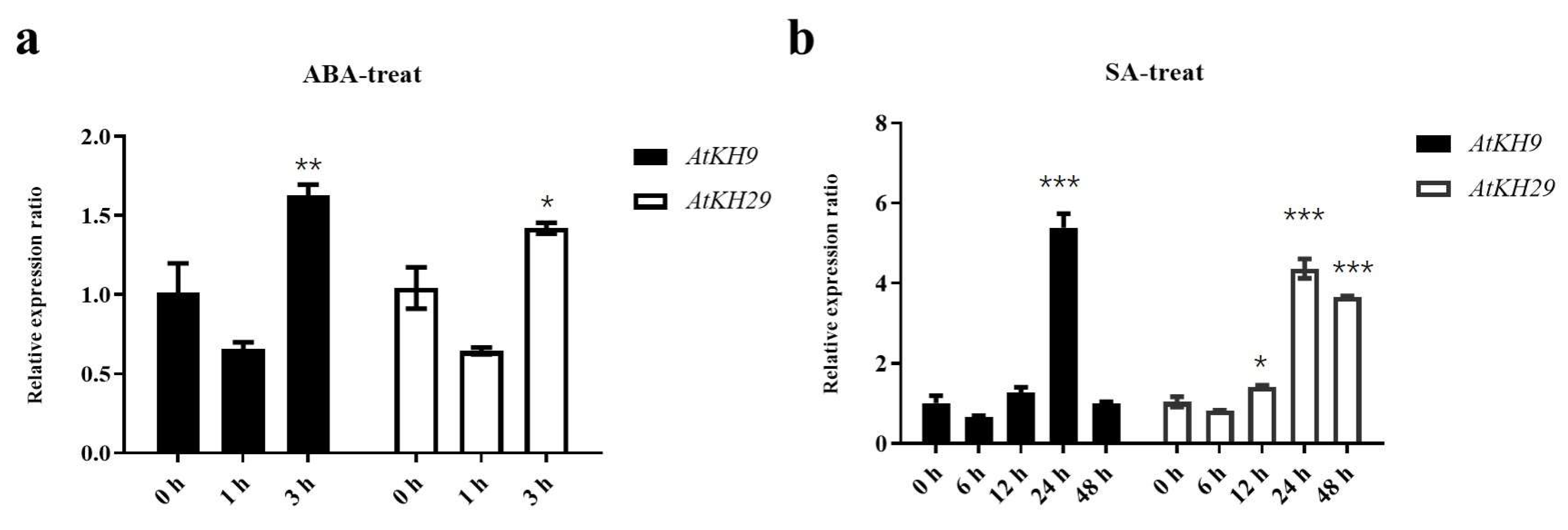
2.5. Expression of KH Family Genes under ABA and SA Treatments
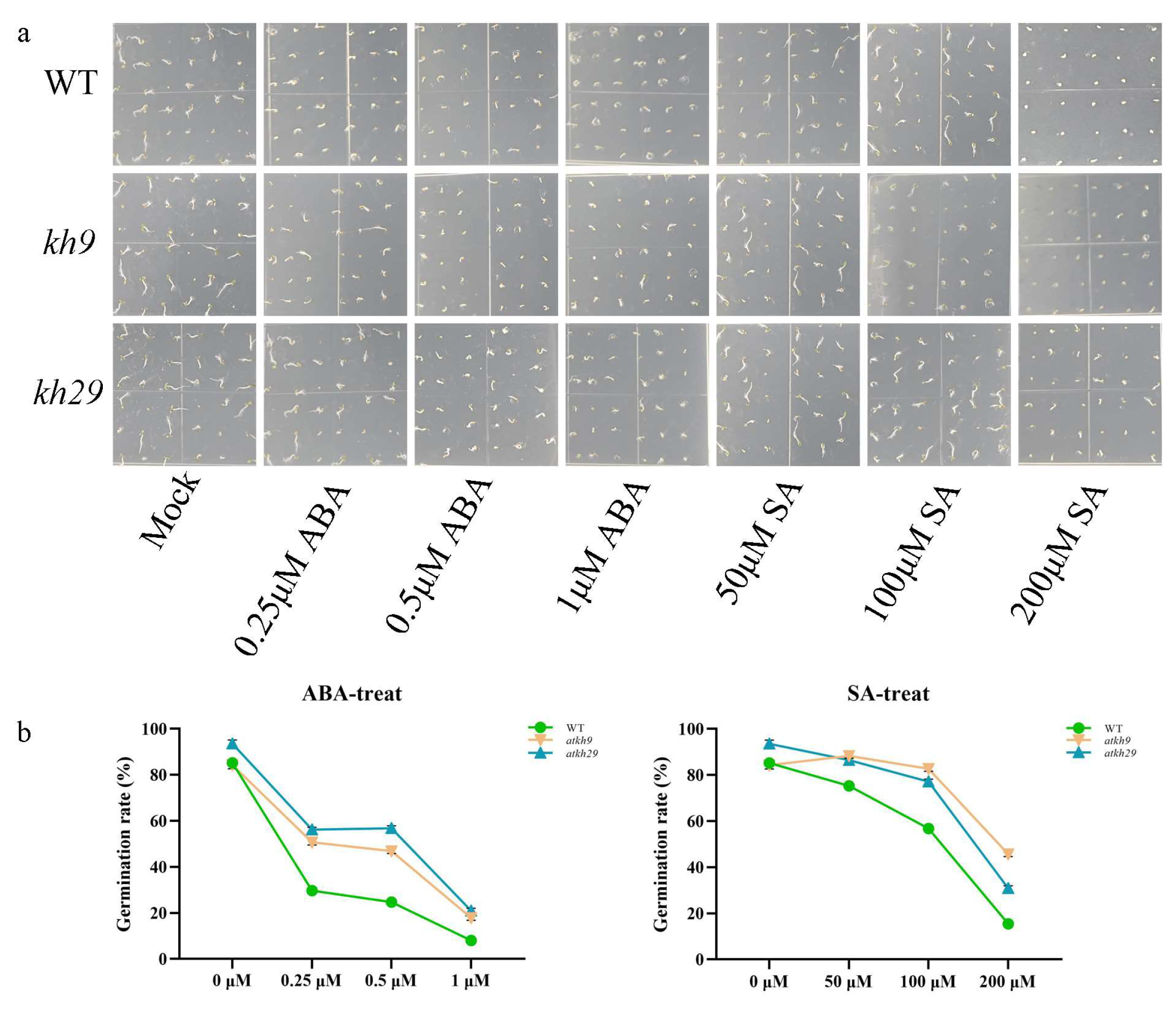
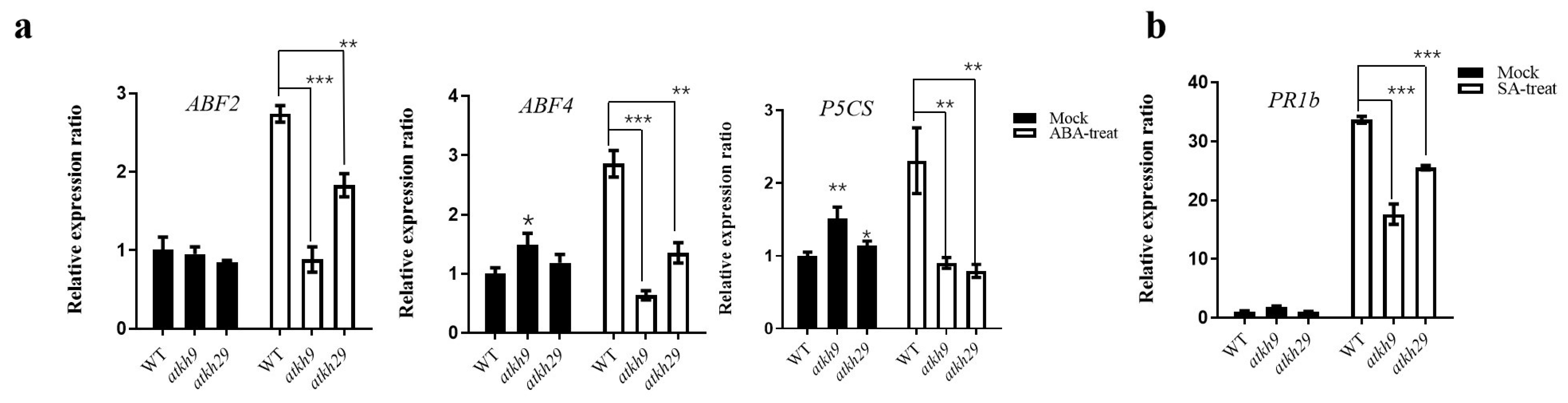
2.6. Function Analysis of AtKH9 and AtKH29
3. Discussion
4. Materials and Methods
4.1. Identification and Characterization of KH Gene Family in Arabidopsis thaliana
4.2. Multiple Sequence Alignment and Phylogeny of KH Family
4.3. Synteny Analysis and Expression Analysis Based on RNA-Seq Data
4.4. Plant Materials and Growing Conditions
4.5. Subcellular Localization
4.6. Materials of AtKH9/29 Genes Expression Analysis and Seed Germination
4.7. ABA and SA-Related Gene Expression Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siomi, H.; Matunis, M.J.; Michael, W.M.; Dreyfuss, G. The pre-mRNA binding K protein contains a novel evolutionarily conserved motif. Nucleic Acids Res. 1993, 21, 1193–1198. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Qiu, H.; He, J.; Liu, L.; Xue, W.; Fox, A.; Tickner, J.; Xu, J. The emerging roles of hnRNPK. J. Cell. Physiol. 2020, 235, 1995–2008. [Google Scholar] [CrossRef] [PubMed]
- Grishin, N.V. KH domain: One motif, two folds. Nucleic Acids Res. 2001, 29, 638–643. [Google Scholar] [CrossRef]
- Athar, Y.M.; Joseph, S. RNA-Binding Specificity of the Human Fragile X Mental Retardation Protein. J. Mol. Biol. 2020, 432, 3851–3868. [Google Scholar] [CrossRef]
- Yadav, M.; Singh, R.S.; Hogan, D.; Vidhyasagar, V.; Yang, S.; Chung, I.Y.W.; Kusalik, A.; Dmitriev, O.Y.; Cygler, M.; Wu, Y. The KH domain facilitates the substrate specificity and unwinding processivity of DDX43 helicase. J. Biol. Chem. 2021, 296, 100085. [Google Scholar] [CrossRef]
- Dreyfuss, G.; Matunis, M.J.; Piñol-Roma, S.; Burd, C.G. HnRNP proteins and the biogenesis of mRNA. Annu. Rev. Biochem. 1993, 62, 289–321. [Google Scholar] [CrossRef]
- Stains, J.P.; Lecanda, F.; Towler, D.A.; Civitelli, R. Heterogeneous nuclear ribonucleoprotein K represses transcription from a cytosine/thymidine-rich element in the osteocalcin promoter. Biochem. J. 2005, 385 Pt 2, 613–623. [Google Scholar] [CrossRef] [Green Version]
- Dowdle, M.E.; Park, S.; Blaser Imboden, S.; Fox, C.A.; Houston, D.W.; Sheets, M.D. A single KH domain in Bicaudal-C links mRNA binding and translational repression functions to maternal development. Development 2019, 146, dev172486. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Jiménez, F.; Sánchez-Margalet, V. Role of Sam68 in post-transcriptional gene regulation. Int. J. Mol. Sci. 2013, 14, 23402–23419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mockler, T.C.; Yu, X.; Shalitin, D.; Parikh, D.; Michael, T.P.; Liou, J.; Huang, J.; Smith, Z.; Alonso, J.M.; Ecker, J.R.; et al. Regulation of flowering time in Arabidopsis by K homology domain proteins. Proc. Natl. Acad. Sci. USA 2004, 101, 12759–12764. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Cazorla, E.; Ripoll, J.J.; Andújar, A.; Bailey, L.J.; Martínez-Laborda, A.; Yanofsky, M.F.; Vera, A. K-homology nuclear ribonucleoproteins regulate floral organ identity and determinacy in arabidopsis. PLoS Genet. 2015, 11, e1004983. [Google Scholar] [CrossRef] [Green Version]
- Ripoll, J.J.; Rodríguez-Cazorla, E.; González-Reig, S.; Andújar, A.; Alonso-Cantabrana, H.; Perez-Amador, M.A.; Carbonell, J.; Martínez-Laborda, A.; Vera, A. Antagonistic interactions between Arabidopsis K-homology domain genes uncover PEPPER as a positive regulator of the central floral repressor FLOWERING LOCUS C. Dev. Biol. 2009, 333, 251–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, M.H.; Kim, J.; Kim, Y.S.; Chung, K.S.; Seo, Y.H.; Lee, I.; Kim, J.; Hong, C.B.; Kim, H.J.; Park, C.M. A new Arabidopsis gene, FLK, encodes an RNA binding protein with K homology motifs and regulates flowering time via FLOWERING LOCUS C. Plant Cell 2004, 16, 731–740. [Google Scholar] [CrossRef] [Green Version]
- Dai, G.Y.; Chen, D.K.; Sun, Y.P.; Liang, W.Y.; Liu, Y.; Huang, L.Q.; Li, Y.K.; He, J.F.; Yao, N. The Arabidopsis KH-domain protein FLOWERING LOCUS Y delays flowering by upregulating FLOWERING LOCUS C family members. Plant Cell Rep. 2020, 39, 1705–1717. [Google Scholar] [CrossRef] [PubMed]
- Ortuño-Miquel, S.; Rodríguez-Cazorla, E.; Zavala-Gonzalez, E.A.; Martínez-Laborda, A.; Vera, A. Arabidopsis HUA ENHANCER 4 delays flowering by upregulating the MADS-box repressor genes FLC and MAF4. Sci. Rep. 2019, 9, 1478. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Kato, N.; Wang, W.; Li, J.; Chen, X. Two RNA binding proteins, HEN4 and HUA1, act in the processing of AGAMOUS pre-mRNA in Arabidopsis thaliana. Dev. Cell 2003, 4, 53–66. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, P.; Christie, M.D.; Seymour, D.K.; Wang, H.; Wang, X.; Hagmann, J.; Kulcheski, F.; Manavella, P.A. KH domain protein RCF3 is a tissue-biased regulator of the plant miRNA biogenesis cofactor HYL1. Proc. Natl. Acad. Sci. USA 2015, 112, 14096–14101. [Google Scholar] [CrossRef] [Green Version]
- Guan, Q.; Wen, C.; Zeng, H.; Zhu, J. A KH domain-containing putative RNA-binding protein is critical for heat stress-responsive gene regulation and thermotolerance in Arabidopsis. Mol. Plant 2013, 6, 386–395. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Veksler-Lublinsky, I.; Zinovyeva, A. HRPK-1, a conserved KH-domain protein, modulates microRNA activity during Caenorhabditis elegans development. PLoS Genet. 2019, 15, e1008067. [Google Scholar] [CrossRef] [Green Version]
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef]
- Finkelstein, R.; Reeves, W.; Ariizumi, T.; Steber, C. Molecular aspects of seed dormancy. Annu. Rev. Plant Biol. 2008, 59, 387–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Bally, J.; Job, C.; Job, D. Seed germination and vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senaratna, T.; Touchell, D.; Bunn, E.; Dixon, K. Acetyl salicylic acid (Aspirin) and salicylic acid induce multiple stress tolerance in bean and tomato plants. Plant Growth Regul. 2000, 30, 157–161. [Google Scholar] [CrossRef]
- Singh, B.; Usha, K. Salicylic acid induced physiological and biochemical changes in wheat seedlings under water stress. Plant Growth Regul. 2003, 39, 137–141. [Google Scholar] [CrossRef]
- Borsani, O.; Valpuesta, V.; Botella, M.A. Evidence for a role of salicylic acid in the oxidative damage generated by NaCl and osmotic stress in Arabidopsis seedlings. Plant Physiol. 2001, 126, 1024–1030. [Google Scholar] [CrossRef] [Green Version]
- El-Mergawi, R.A.; Abd El-Wahed, M.S.A. Effect of exogenous salicylic acid or indole acetic acid on their endogenous levels, germination, and growth in maize. Bull. Natl. Res. Cent. 2020, 44, 167. [Google Scholar] [CrossRef]
- Valverde, R.; Edwards, L.; Regan, L. Structure and function of KH domains. FEBS J. 2008, 275, 2712–2726. [Google Scholar] [CrossRef]
- Rushton, D.L.; Tripathi, P.; Rabara, R.C.; Lin, J.; Ringler, P.; Boken, A.K.; Langum, T.J.; Smidt, L.; Boomsma, D.D.; Emme, N.J.; et al. WRKY transcription factors: Key components in abscisic acid signalling. Plant Biotechnol. J. 2012, 10, 2–11. [Google Scholar] [CrossRef]
- Strizhov, N.; Abrahám, E.; Okrész, L.; Blickling, S.; Zilberstein, A.; Schell, J.; Koncz, C.; Szabados, L. Differential expression of two P5CS genes controlling proline accumulation during salt-stress requires ABA and is regulated by ABA1, ABI1 and AXR2 in Arabidopsis. Plant J. 1997, 12, 557–569. [Google Scholar]
- Wang, N.; Liu, M.; Guo, L.; Yang, X.; Qiu, D. A Novel Protein Elicitor (PeBA1) from Bacillus amyloliquefaciens NC6 Induces Systemic Resistance in Tobacco. Int. J. Biol. Sci. 2016, 12, 757–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mou, S.; Liu, Z.; Guan, D.; Qiu, A.; Lai, Y.; He, S. Functional analysis and expressional characterization of rice ankyrin repeat-containing protein, OsPIANK1, in basal defense against Magnaporthe oryzae attack. PLoS ONE 2013, 8, e59699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thatcher, L.F.; Kamphuis, L.G.; Hane, J.K.; Oñate-Sánchez, L.; Singh, K.B. The Arabidopsis KH-Domain RNA-Binding Protein ESR1 Functions in Components of Jasmonate Signalling, Unlinking Growth Restraint and Resistance to Stress. PLoS ONE 2015, 10, e0126978. [Google Scholar] [CrossRef] [Green Version]
- Lorković, Z.J.; Barta, A. Genome analysis: RNA recognition motif (RRM) and K homology (KH) domain RNA-binding proteins from the flowering plant Arabidopsis thaliana. Nucleic Acids Res. 2002, 30, 623–635. [Google Scholar] [CrossRef] [Green Version]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol 2004, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Wu, N.; Song, W.; Yin, G.; Qin, Y.; Yan, Y.; Hu, Y. Soybean (Glycine max) expansin gene superfamily origins: Segmental and tandem duplication events followed by divergent selection among subfamilies. BMC Plant Biol 2014, 14, 93. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res 2015, 43, W39–W49. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Waese, J.; Fan, J.; Pasha, A.; Yu, H.; Fucile, G.; Shi, R.; Cumming, M.; Kelley, L.A.; Sternberg, M.J.; Krishnakumar, V.; et al. ePlant: Visualizing and Exploring Multiple Levels of Data for Hypothesis Generation in Plant Biology. Plant Cell 2017, 29, 1806–1821. [Google Scholar] [CrossRef] [PubMed]
- Boyes, D.C.; Zayed, A.M.; Ascenzi, R.; McCaskill, A.J.; Hoffman, N.E.; Davis, K.R.; Görlach, J. Growth stage-based phenotypic analysis of Arabidopsis: A model for high throughput functional genomics in plants. Plant Cell 2001, 13, 1499–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhang, F.; Yu, Y.; Feng, L.; Jia, J.; Liu, B.; Li, B.; Guo, H.; Zhai, J. A Comprehensive Online Database for Exploring ∼20,000 Public Arabidopsis RNA-Seq Libraries. Mol. Plant 2020, 13, 1231–1233. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yu, G.; Wen, W.; Ma, X.; Xu, B.; Huang, B. Functional characterization and hormonal regulation of the PHEOPHYTINASE gene LpPPH controlling leaf senescence in perennial ryegrass. J. Exp. Bot. 2016, 67, 935–945. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Ren, Z.; Cheng, C.; Wang, T.; Ji, H.; Zhao, Y.; Deng, Z.; Zhi, L.; Lu, J.; Wu, X.; et al. Counteraction of ABA-Mediated Inhibition of Seed Germination and Seedling Establishment by ABA Signaling Terminator in Arabidopsis. Mol. Plant 2020, 13, 1284–1297. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Ma, Y.; Liu, R.; Li, G. Genome-Wide Characterization and Expression Analysis of KH Family Genes Response to ABA and SA in Arabidopsis thaliana. Int. J. Mol. Sci. 2022, 23, 511. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010511
Zhang Y, Ma Y, Liu R, Li G. Genome-Wide Characterization and Expression Analysis of KH Family Genes Response to ABA and SA in Arabidopsis thaliana. International Journal of Molecular Sciences. 2022; 23(1):511. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010511
Chicago/Turabian StyleZhang, Yanjie, Yu Ma, Ruiqi Liu, and Guanglin Li. 2022. "Genome-Wide Characterization and Expression Analysis of KH Family Genes Response to ABA and SA in Arabidopsis thaliana" International Journal of Molecular Sciences 23, no. 1: 511. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010511