
Runx1 Messenger RNA Delivered by Polyplex Nanomicelles Alleviate Spinal Disc Hydration Loss in a Rat Disc Degeneration Model

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
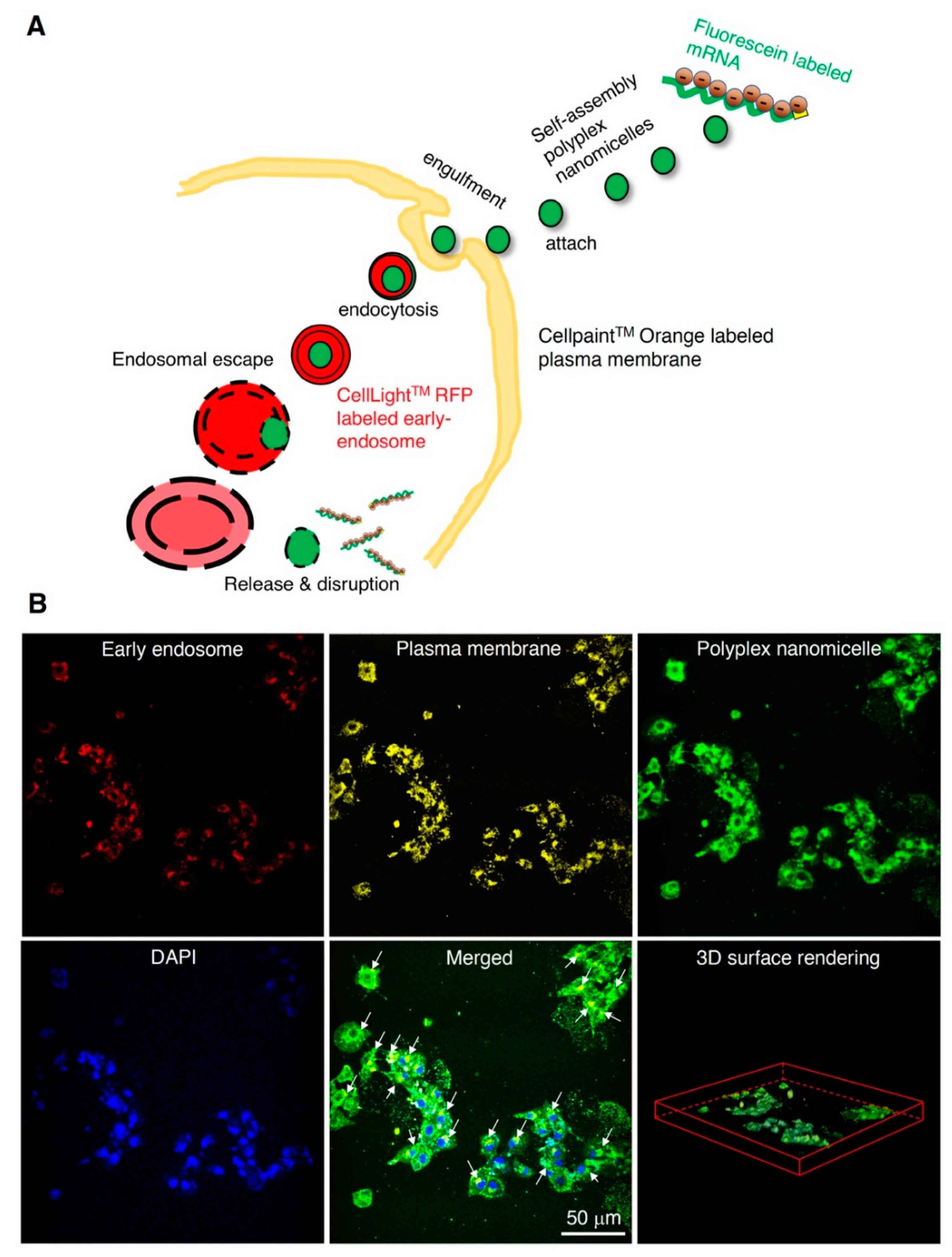
2.1. Characterization of mRNA-Loaded Polyplex Nanomicelles and Evaluation of Endocytosis
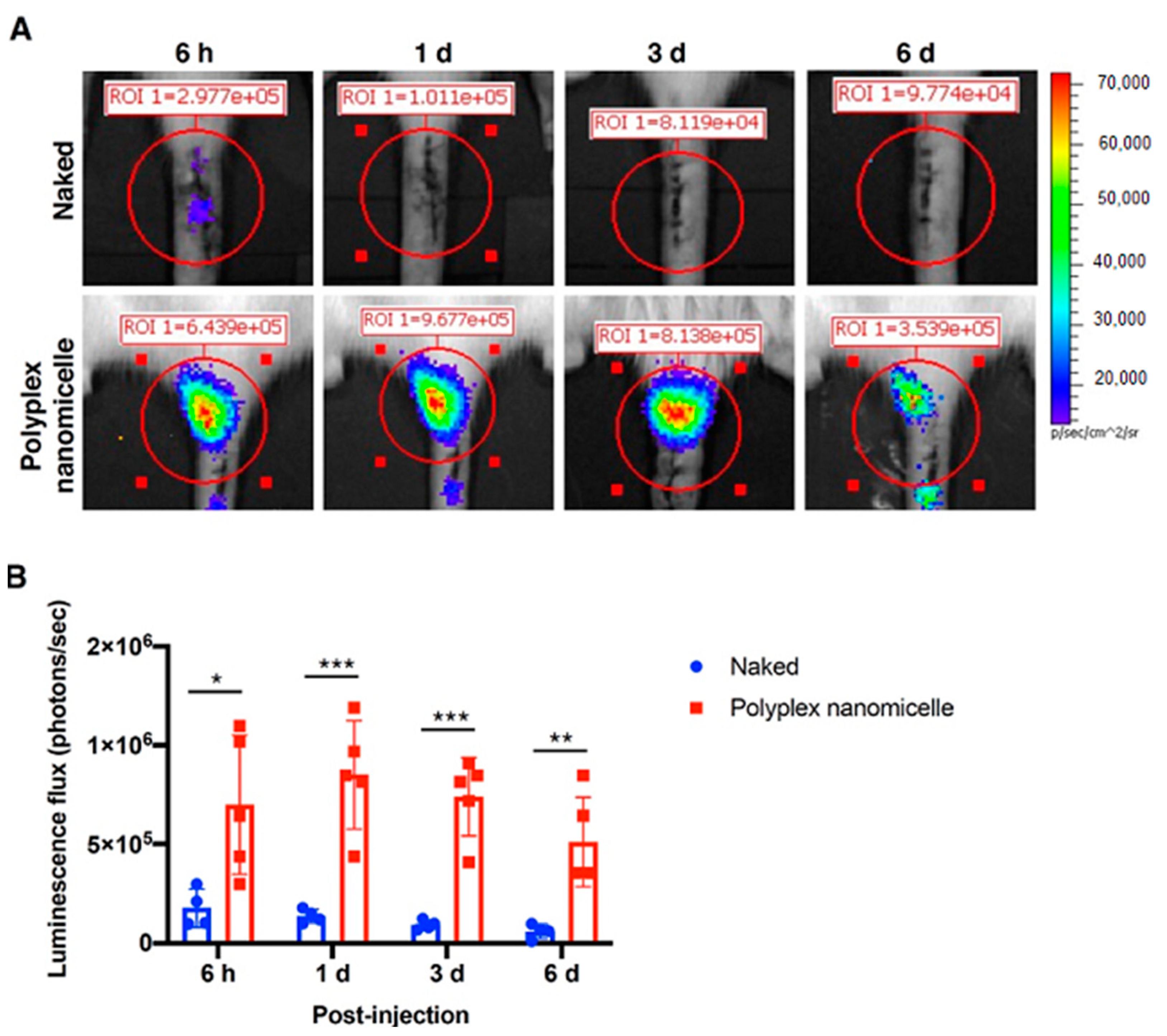
2.2. mRNA-Loaded Polyplex Nanomicelle-Mediated Gene Expression in a Rat Disc Puncture Model
2.3. Runx1 mRNA-Loaded Polyplex Nanomicelles Ameliorated the Disc Hydration Content in a Disc Degeneration Model
2.4. Disc Degeneration Evaluated by μCT and Histological Examinations
3. Discussion
4. Materials and Methods
4.1. Synthesis of PEG-PAsp(DET) Block Copolymer and Nuclear Magnetic Resonance (NMR) Analysis
4.2. Construction of Vectors for In Vitro Transcription (IVT) and Preparation of mRNA
4.3. Preparation of Polyplex Nanomicelles and Characterization of the Physicochemical Properties
4.4. Cell Culture and Endocytosis of mRNA-Loaded Polyplex Nanomicelles
4.5. Animal Coccygeal Disc Degeneration Model and Luc2 mRNA Delivery
4.6. Runx1 mRNA Transfection and Western Blot Analysis
4.7. Runx1 mRNA Polyplex Nanomicelles Delivery and 7T-MRI Examination
4.8. CT Examination
4.9. Histological Examination
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- James, S.L.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef] [Green Version]
- Martin, B.I.; Deyo, R.A.; Mirza, S.K.; Turner, J.A.; Comstock, B.A.; Hollingworth, W.; Sullivan, S.D. Expenditures and health status among adults with back and neck problems. JAMA 2008, 299, 656–664. [Google Scholar] [CrossRef] [Green Version]
- Vos, T.; Flaxman, A.D.; Naghavi, M.; Lozano, R.; Michaud, C.; Ezzati, M.; Shibuya, K.; Salomon, J.A.; Abdalla, S.; Aboyans, V.; et al. Years lived with disability (YLDs) for 1160 sequelae of 289 diseases and injuries 1990-2010: A systematic analysis for the global burden of disease study 2010. Lancet 2012, 380, 2163–2196. [Google Scholar] [CrossRef]
- Cheung, K.M.; Karppinen, J.; Chan, D.; Ho, D.W.; Song, Y.Q.; Sham, P.; Cheah, K.S.; Leong, J.C.; Luk, K.D. Prevalence and pattern of lumbar magnetic resonance imaging changes in a population study of one thousand forty-three individuals. Spine 2009, 34, 934–940. [Google Scholar] [CrossRef] [PubMed]
- Takatalo, J.; Karppinen, J.; Niinimaki, J.; Taimela, S.; Nayha, S.; Mutanen, P.; Sequeiros, R.B.; Kyllonen, E.; Tervonen, O. Does lumbar disc degeneration on magnetic resonance imaging associate with low back symptom severity in young Finnish adults? Spine 2011, 36, 2180–2189. [Google Scholar] [CrossRef]
- Pattappa, G.; Li, Z.; Peroglio, M.; Wismer, N.; Alini, M.; Grad, S. Diversity of intervertebral disc cells: Phenotype and function. J. Anat. 2012, 221, 480–496. [Google Scholar] [CrossRef]
- Miele, V.J.; Panjabi, M.M.; Benzel, E.C. Anatomy and biomechanics of the spinal column and cord. Handb. Clin. Neurol. 2012, 109, 31–43. [Google Scholar] [CrossRef]
- Kepler, C.K.; Ponnappan, R.K.; Tannoury, C.A.; Risbud, M.V.; Anderson, D.G. The molecular basis of intervertebral disc degeneration. Spine J. 2013, 13, 318–330. [Google Scholar] [CrossRef]
- Roughley, P.J.; Melching, L.I.; Heathfield, T.F.; Pearce, R.H.; Mort, J.S. The structure and degradation of aggrecan in human intervertebral disc. Eur. Spine J. 2006, 15 (Suppl. 3), S326–S332. [Google Scholar] [CrossRef] [Green Version]
- Sakai, D.; Andersson, G.B. Stem cell therapy for intervertebral disc regeneration: Obstacles and solutions. Nat. Rev. Rheumatol. 2015, 11, 243–256. [Google Scholar] [CrossRef]
- Castro, I.; Santos, D.P.; Christoph Dde, H.; Landeiro, J.A. The history of spinal surgery for disc disease: An illustrated timeline. Arq. Neuropsiquiatr. 2005, 63, 701–706. [Google Scholar] [CrossRef] [Green Version]
- Denoziere, G.; Ku, D.N. Biomechanical comparison between fusion of two vertebrae and implantation of an artificial intervertebral disc. J. Biomech. 2006, 39, 766–775. [Google Scholar] [CrossRef]
- Disch, A.C.; Schmoelz, W.; Matziolis, G.; Schneider, S.V.; Knop, C.; Putzier, M. Higher risk of adjacent segment degeneration after floating fusions: Long-term outcome after low lumbar spine fusions. J. Spinal Disord. Tech. 2008, 21, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Siepe, C.J.; Heider, F.; Wiechert, K.; Hitzl, W.; Ishak, B.; Mayer, M.H. Mid- to long-term results of total lumbar disc replacement: A prospective analysis with 5- to 10-year follow-up. Spine J. 2014, 14, 1417–1431. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Cao, J.; Wang, L.; Shen, Y. Prevalence of adjacent segment disease following cervical spine surgery: A PRISMA-compliant systematic review and meta-analysis. Medicine 2016, 95, e4171. [Google Scholar] [CrossRef] [PubMed]
- Pan, A.; Hai, Y.; Yang, J.; Zhou, L.; Chen, X.; Guo, H. Adjacent segment degeneration after lumbar spinal fusion compared with motion-preservation procedures: A meta-analysis. Eur. Spine J. 2016, 25, 1522–1532. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Song, Y.; Liu, L.; Xue, Y. Adjacent segment degeneration and disease after lumbar fusion compared with motion-preserving procedures: A meta-analysis. Eur. J. Orthop. Surg. Traumatol. 2014, 24 (Suppl. 1), S245–S253. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.P.; Chen, H.L.; Cheng, H.B. Prevalence of adjacent segment degeneration after spine surgery: A systematic review and meta-analysis. Spine 2013, 38, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Mochida, J.; Nishimura, K.; Nomura, T.; Toh, E.; Chiba, M. The importance of preserving disc structure in surgical approaches to lumbar disc herniation. Spine 1996, 21, 1556–1563. [Google Scholar] [CrossRef]
- Boden, S.D.; McCowin, P.R.; Davis, D.O.; Dina, T.S.; Mark, A.S.; Wiesel, S. Abnormal magnetic-resonance scans of the cervical spine in asymptomatic subjects. A prospective investigation. J. Bone Jt. Surg. 1990, 72, 1178–1184. [Google Scholar] [CrossRef]
- Risbud, M.V.; Shapiro, I.M. Role of cytokines in intervertebral disc degeneration: Pain and disc content. Nat. Rev. Rheumatol. 2014, 10, 44–56. [Google Scholar] [CrossRef]
- Aini, H.; Itaka, K.; Fujisawa, A.; Uchida, H.; Uchida, S.; Fukushima, S.; Kataoka, K.; Saito, T.; Chung, U.I.; Ohba, S. Messenger RNA delivery of a cartilage-anabolic transcription factor as a disease-modifying strategy for osteoarthritis treatment. Sci. Rep. 2016, 6, 18743. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.Y.; Crowley, S.T.; Uchida, S.; Komaki, Y.; Kataoka, K.; Itaka, K. Treatment of Intervertebral Disk Disease by the Administration of mRNA Encoding a Cartilage-Anabolic Transcription Factor. Mol. Ther. Nucleic Acids 2019, 16, 162–171. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.; Zhu, S.; Tremblay, M.S.; Payette, J.N.; Wang, J.; Bouchez, L.C.; Meeusen, S.; Althage, A.; Cho, C.Y.; Wu, X.; et al. A stem cell-based approach to cartilage repair. Science 2012, 336, 717–721. [Google Scholar] [CrossRef] [Green Version]
- Yano, F.; Hojo, H.; Ohba, S.; Fukai, A.; Hosaka, Y.; Ikeda, T.; Saito, T.; Hirata, M.; Chikuda, H.; Takato, T.; et al. A novel disease-modifying osteoarthritis drug candidate targeting Runx1. Ann. Rheum. Dis. 2013, 72, 748–753. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Belflower, R.M.; Dong, Y.F.; Schwarz, E.M.; O’Keefe, R.J.; Drissi, H. Runx1/AML1/Cbfa2 mediates onset of mesenchymal cell differentiation toward chondrogenesis. J. Bone Miner. Res. 2005, 20, 1624–1636. [Google Scholar] [CrossRef]
- Wotton, S.; Terry, A.; Kilbey, A.; Jenkins, A.; Herzyk, P.; Cameron, E.; Neil, J.C. Gene array analysis reveals a common Runx transcriptional programme controlling cell adhesion and survival. Oncogene 2008, 27, 5856–5866. [Google Scholar] [CrossRef] [Green Version]
- Chan, L.Y.; Khung, Y.L.; Lin, C.Y. Preparation of messenger RNA nanomicelles via non-cytotoxic PEG-polyamine nanocomplex for intracerebroventicular delivery: A proof-of-concept study in mouse models. Nanomaterials 2019, 9, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boden, S.D.; Balderston, R.A.; Heller, J.G.; Hanley, E.N., Jr.; Zigler, J.E. An AOA critical issue. Disc replacements: This time will we really cure low-back and neck pain? J. Bone Joint Surg. 2004, 86, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Auerbach, J.D.; Johannessen, W.; Borthakur, A.; Wheaton, A.J.; Dolinskas, C.A.; Balderston, R.A.; Reddy, R.; Elliott, D.M. In vivo quantification of human lumbar disc degeneration using T(1rho)-weighted magnetic resonance imaging. Eur. Spine J. 2006, 15 (Suppl 3), S338–S344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheaton, A.J.; Dodge, G.R.; Elliott, D.M.; Nicoll, S.B.; Reddy, R. Quantification of cartilage biomechanical and biochemical properties via T1rho magnetic resonance imaging. Magn. Reson. Med. 2005, 54, 1087–1093. [Google Scholar] [CrossRef]
- Grunert, P.; Hudson, K.D.; Macielak, M.R.; Aronowitz, E.; Borde, B.H.; Alimi, M.; Njoku, I.; Ballon, D.; Tsiouris, A.J.; Bonassar, L.J.; et al. Assessment of intervertebral disc degeneration based on quantitative magnetic resonance imaging analysis: An in vivo study. Spine 2014, 39, E369–E378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, S.; Huang, Y.; Che, X.; Gu, R. Faster RCNN-based detection of cervical spinal cord injury and disc degeneration. J. Appl. Clin. Med. Phys. 2020, 21, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.M.; Diaz-Hernandez, M.E.; Presciutti, S.M.; Drissi, H. Network analysis identifies gene regulatory network indicating the role of RUNX1 in human intervertebral disc degeneration. Genes 2020, 11, 771. [Google Scholar] [CrossRef] [PubMed]
- Torkki, M.; Majuri, M.L.; Wolff, H.; Koskelainen, T.; Haapea, M.; Niinimaki, J.; Alenius, H.; Lotz, J.; Karppinen, J. Osteoclast activators are elevated in intervertebral disks with Modic changes among patients operated for herniated nucleus pulposus. Eur. Spine J. 2016, 25, 207–216. [Google Scholar] [CrossRef]
- Komori, T. Regulation of skeletal development by the Runx family of transcription factors. J. Cell Biochem. 2005, 95, 445–453. [Google Scholar] [CrossRef]
- Tosteson, A.N.; Tosteson, T.D.; Lurie, J.D.; Abdu, W.; Herkowitz, H.; Andersson, G.; Albert, T.; Bridwell, K.; Zhao, W.; Grove, M.R.; et al. Comparative effectiveness evidence from the spine patient outcomes research trial: Surgical versus nonoperative care for spinal stenosis, degenerative spondylolisthesis, and intervertebral disc herniation. Spine 2011, 36, 2061–2068. [Google Scholar] [CrossRef]
- Parker, S.L.; Godil, S.S.; Mendenhall, S.K.; Zuckerman, S.L.; Shau, D.N.; McGirt, M.J. Two-year comprehensive medical management of degenerative lumbar spine disease (lumbar spondylolisthesis, stenosis, or disc herniation): A value analysis of cost, pain, disability, and quality of life: Clinical article. J. Neurosurg. Spine 2014, 21, 143–149. [Google Scholar] [CrossRef]
- Takeoka, Y.; Yurube, T.; Nishida, K. Gene therapy approach for intervertebral disc degeneration: An update. Neurospine 2020, 17, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Hilibrand, A.S.; Carlson, G.D.; Palumbo, M.A.; Jones, P.K.; Bohlman, H.H. Radiculopathy and myelopathy at segments adjacent to the site of a previous anterior cervical arthrodesis. J. Bone Jt. Surg. 1999, 81, 519–528. [Google Scholar] [CrossRef] [Green Version]
- Sears, W.R.; Sergides, I.G.; Kazemi, N.; Smith, M.; White, G.J.; Osburg, B. Incidence and prevalence of surgery at segments adjacent to a previous posterior lumbar arthrodesis. Spine J. 2011, 11, 11–20. [Google Scholar] [CrossRef]
- Masuda, K. Biological repair of the degenerated intervertebral disc by the injection of growth factors. Eur. Spine J. 2008, 17 (Suppl 4), 441–451. [Google Scholar] [CrossRef] [Green Version]
- Nishida, K.; Suzuki, T.; Kakutani, K.; Yurube, T.; Maeno, K.; Kurosaka, M.; Doita, M. Gene therapy approach for disc degeneration and associated spinal disorders. Eur Spine J. 2008, 17 (Suppl. 4), 459–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, H.; Montgomery, S.R.; Aghdasi, B.; Kaner, T.; Tan, Y.; Tian, H.; Terrell, R.; Wang, J.C.; Daubs, M.D. The effect of bone morphogenetic protein-2 injection at different time points on intervertebral disk degeneration in a rat tail model. J. Spinal Disord. Tech. 2015, 28, E35–E44. [Google Scholar] [CrossRef]
- Farhang, N.; Ginley-Hidinger, M.; Berrett, K.C.; Gertz, J.; Lawrence, B.; Bowles, R.D. Lentiviral CRISPR epigenome editing of inflammatory receptors as a gene therapy strategy for disc degeneration. Human Gene Ther. 2019, 30, 1161–1175. [Google Scholar] [CrossRef]
- Feng, G.; Zha, Z.; Huang, Y.; Li, J.; Wang, Y.; Ke, W.; Chen, H.; Liu, L.; Song, Y.; Ge, Z. Sustained and bioresponsive two-stage delivery of therapeutic miRNA via polyplex micelle-loaded injectable hydrogels for inhibition of intervertebral disc fibrosis. Adv. Healthc. Mat. 2018, 7, e1800623. [Google Scholar] [CrossRef] [PubMed]
- Seki, S.; Asanuma-Abe, Y.; Masuda, K.; Kawaguchi, Y.; Asanuma, K.; Muehleman, C.; Iwai, A.; Kimura, T. Effect of small interference RNA (siRNA) for ADAMTS5 on intervertebral disc degeneration in the rabbit anular needle-puncture model. Arthritis Res. Ther. 2009, 11, R166. [Google Scholar] [CrossRef] [Green Version]
- Jirikowski, G.F.; Sanna, P.P.; Maciejewski-Lenoir, D.; Bloom, F.E. Reversal of diabetes insipidus in Brattleboro rats: Intrahypothalamic injection of vasopressin mRNA. Science 1992, 255, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Boczkowski, D.; Nair, S.K.; Snyder, D.; Gilboa, E. Dendritic cells pulsed with RNA are potent antigen-presenting cells in vitro and in vivo. J. Exp. Med. 1996, 184, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Sahin, U.; Kariko, K.; Tureci, O. mRNA-based therapeutics–developing a new class of drugs. Nat. Rev. Drug Discov. 2014, 13, 759–780. [Google Scholar] [CrossRef]
- Zou, S.; Scarfo, K.; Nantz, M.H.; Hecker, J.G. Lipid-mediated delivery of RNA is more efficient than delivery of DNA in non-dividing cells. Int. J. of Pharm. 2010, 389, 232–243. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.Y.; Perche, F.; Ikegami, M.; Uchida, S.; Kataoka, K.; Itaka, K. Messenger RNA-based therapeutics for brain diseases: An animal study for augmenting clearance of beta-amyloid by intracerebral administration of neprilysin mRNA loaded in polyplex nanomicelles. J. Control. Release 2016, 235, 268–275. [Google Scholar] [CrossRef]
- Phua, K.K.; Leong, K.W.; Nair, S.K. Transfection efficiency and transgene expression kinetics of mRNA delivered in naked and nanoparticle format. J. Control. Release 2013, 166, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Pardi, N.; Tuyishime, S.; Muramatsu, H.; Kariko, K.; Mui, B.L.; Tam, Y.K.; Madden, T.D.; Hope, M.J.; Weissman, D. Expression kinetics of nucleoside-modified mRNA delivered in lipid nanoparticles to mice by various routes. J. Control. Release 2015, 217, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Wallach, C.J.; Kim, J.S.; Sobajima, S.; Lattermann, C.; Oxner, W.M.; McFadden, K.; Robbins, P.D.; Gilbertson, L.G.; Kang, J.D. Safety assessment of intradiscal gene transfer: A pilot study. Spine J. 2006, 6, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.H.; Ghivizzani, S.C.; Robbins, P.D. Arthritis gene therapy’s first death. Arthritis Res. Ther. 2008, 10, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, K.; Aota, Y.; Muehleman, C.; Imai, Y.; Okuma, M.; Thonar, E.J.; Andersson, G.B.; An, H.S. A novel rabbit model of mild, reproducible disc degeneration by an anulus needle puncture: Correlation between the degree of disc injury and radiological and histological appearances of disc degeneration. Spine 2005, 30, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Holtkamp, S.; Kreiter, S.; Selmi, A.; Simon, P.; Koslowski, M.; Huber, C.; Türeci, O.; Sahin, U. Modification of antigen-encoding RNA increases stability, translational efficacy, and T-cell stimulatory capacity of dendritic cells. Blood 2006, 108, 4009–4017. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, C.-C.; Tsou, H.-K.; Chang, H.-H.; Chan, L.Y.; Zhuo, G.-Y.; Maeda, T.; Lin, C.-Y. Runx1 Messenger RNA Delivered by Polyplex Nanomicelles Alleviate Spinal Disc Hydration Loss in a Rat Disc Degeneration Model. Int. J. Mol. Sci. 2022, 23, 565. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010565
Chang C-C, Tsou H-K, Chang H-H, Chan LY, Zhuo G-Y, Maeda T, Lin C-Y. Runx1 Messenger RNA Delivered by Polyplex Nanomicelles Alleviate Spinal Disc Hydration Loss in a Rat Disc Degeneration Model. International Journal of Molecular Sciences. 2022; 23(1):565. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010565
Chicago/Turabian StyleChang, Cheng-Chung, Hsi-Kai Tsou, Hsu-Hsin Chang, Long Yi Chan, Guan-Yu Zhuo, Tomoji Maeda, and Chin-Yu Lin. 2022. "Runx1 Messenger RNA Delivered by Polyplex Nanomicelles Alleviate Spinal Disc Hydration Loss in a Rat Disc Degeneration Model" International Journal of Molecular Sciences 23, no. 1: 565. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010565