
The Regulation of Collagen Processing by miRNAs in Disease and Possible Implications for Bone Turnover

 , , ,
, , ,
Abstract
:1. Introduction
2. Collagen Formation
2.1. Collagen Expression
2.1.1. COL1A1 Expression Regulation by miRNAs in Skeletal Tissues
2.1.2. Regulation of COL1A1 Expression by miRNAs in Non-Skeletal Tissues
2.1.3. COL1A1 and miRNAs in the Development of Neoplasms
2.2. Intracellular Stages of Collagen Formation
2.2.1. Hydroxylation of Proline and Lysine Residues, Glycosylation of Some Hydroxylysine Residues
2.2.2. Procollagen Glycosylation
2.2.3. Formation of Intra-Chain and Inter-Chain Disulphide Bonds in Terminal Peptides
2.2.4. Procollagen Triple Helix Formation
2.3. Extracellular Stages of Collagen Formation
2.3.1. Cleavage of Amino- and Carboxy Propeptides
2.3.2. Aggregation of Collagen Fibres
2.3.3. Crosslinking
3. Collagen Interactions with Non-Collagenous Proteins and Their Impact on Bone Quality
3.1. Osteopontin
3.2. Thrombospondin-2
3.3. Biglycan and Decorin
3.4. Osteonectin
4. Interactions between Collagen and the Mineral Phase in Bone
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Buckwalter, J.A.; Glimcher, M.J.; Cooper, R.R.; Recker, R. Bone biology. I: Structure, blood supply, cells, matrix, and mineralization. Instr. Course Lect. 1996, 45, 371–386. [Google Scholar] [PubMed]
- Saito, M.; Marumo, K. Effects of Collagen Crosslinking on Bone Material Properties in Health and Disease. Calcif. Tissue Int. 2015, 97, 242–261. [Google Scholar] [CrossRef] [PubMed]
- Cava, C.; Bertoli, G.; Castiglioni, I. Portrait of Tissue-Specific Coexpression Networks of Noncoding RNAs (miRNA and lncRNA) and mRNAs in Normal Tissues. Comput. Math. Methods Med. 2019, 2019, 9029351. [Google Scholar] [CrossRef]
- Bartel, D.P. Metazoan MicroRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Jia, L.; Zheng, Y.; Li, W. Bone remodeling induced by mechanical forces is regulated by miRNAs. Biosci. Rep. 2018, 38, 38. [Google Scholar] [CrossRef]
- Gao, Y.; Patil, S.; Qian, A. The Role of MicroRNAs in Bone Metabolism and Disease. Int. J. Mol. Sci 2020, 21, 6081. [Google Scholar] [CrossRef]
- Bellavia, D.; De Luca, A.; Carina, V.; Costa, V.; Raimondi, L.; Salamanna, F.; Alessandro, R.; Fini, M.; Giavaresi, G. Deregulated miRNAs in bone health: Epigenetic roles in osteoporosis. Bone 2019, 122, 52–75. [Google Scholar] [CrossRef]
- Karsenty, G.; Park, R.W. Regulation of type I collagen genes expression. Int. Rev. Immunol. 1995, 12, 177–185. [Google Scholar] [CrossRef]
- Moradifard, S.; Hoseinbeyki, M.; Emam, M.M.; Parchiniparchin, F.; Ebrahimi-Rad, M. Association of the Sp1 binding site and -1997 promoter variations in COL1A1 with osteoporosis risk: The application of meta-analysis and bioinformatics approaches offers a new perspective for future research. Mutat. Res. Rev. Mutat. Res. 2020, 786, 108339. [Google Scholar] [CrossRef]
- Tadić, T.; Erceg, I.; Stover, M.L.; Rowe, D.W.; Lichtler, A.C. Dlx5 induces expression of COL1A1 promoter contained in a retrovirus vector. Croat. Med. J. 2001, 42, 436–439. [Google Scholar] [PubMed]
- Komori, T. Regulation of bone development and extracellular matrix protein genes by RUNX2. Cell Tissue Res. 2010, 339, 189–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, L.; Xiao, Y.; Wu, Q.; Liu, L.; Zhang, C.; Pan, X. TLR4/NF-ÎşB axis signaling pathway-dependent up-regulation of miR-625–5p contributes to human intervertebral disc degeneration by targeting COL1A1. Am. J. Transl Res. 2018, 11, 1374–1388. [Google Scholar]
- Kashiyama, K.; Mitsutake, N.; Matsuse, M.; Ogi, T.; Saenko, V.A.; Ujifuku, K.; Utani, A.; Hirano, A.; Yamashita, S. miR-196a downregulation increases the expression of type I and III collagens in keloid fibroblasts. J. Investig. Dermatol. 2012, 132, 1597–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gauglitz, G.G.; Korting, H.C.; Pavicic, T.; Ruzicka, T.; Jeschke, M.G. Hypertrophic scarring and keloids: Pathomechanisms and current and emerging treatment strategies. Mol. Med. 2011, 17, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Bi, S.; Chai, L.; Yuan, X.; Cao, C.; Li, S. MicroRNA-98 inhibits the cell proliferation of human hypertrophic scar fibroblasts via targeting Col1A1. Biol. Res. 2017, 50, 22. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Feng, C.; Song, K.; Qi, Z.; Huang, W.; Wang, Y. lncRNA-H19/miR-29a axis affected the viability and apoptosis of keloid fibroblasts through acting upon COL1A1 signaling. J. Cell Biochem. 2020, 121, 4364–4376. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Itami, S.; Kuroda, M.; Yoshizato, K.; Kawada, N.; Murakami, Y. MiR-29a Assists in Preventing the Activation of Human Stellate Cells and Promotes Recovery from Liver Fibrosis in Mice. Mol. Ther. 2016, 24, 1848–1859. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Chen, Y.; Wang, Y.; Li, Y.; Cheng, Q. MicroRNA-513b-5p targets COL1A1 and COL1A2 associated with the formation and rupture of intracranial aneurysm. Sci. Rep. 2021, 11, 14897. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, S.; Zhang, W.; Ni, L.; Hu, Z.; Sheng, Z.; Yin, B. MiR-128 inhibits the osteogenic differentiation in osteoporosis by down-regulating SIRT6 expression. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, D.; An, S.; Choi, B.G.; Kim, K.; Choi, Y.M.; Ahn, K.J.; An, I.S.; Cha, H.J. MicroRNA-378b regulates α-1-type 1 collagen expression via sirtuin 6 interference. Mol. Med. Rep. 2017, 16, 8520–8524. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; An, S.; Choi, B.G.; Joo, D.; Choi, Y.M.; Ahn, K.J.; An, I.S.; Cha, H.J. Arctiin regulates collagen type 1α chain 1 mRNA expression in human dermal fibroblasts via the miR-378b-SIRT6 axis. Mol. Med. Rep. 2017, 16, 9120–9124. [Google Scholar] [CrossRef] [PubMed]
- Probert, C.; Dottorini, T.; Speakman, A.; Hunt, S.; Nafee, T.; Fazeli, A.; Wood, S.; Brown, J.E.; James, V. Communication of prostate cancer cells with bone cells via extracellular vesicle RNA; a potential mechanism of metastasis. Oncogene 2019, 38, 1751–1763. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Sui, B.; Fan, W.; Lei, L.; Zhou, L.; Yang, L.; Diao, Y.; Zhang, Y.; Li, Z.; Liu, J.; et al. Exosomes derived from osteogenic tumor activate osteoclast differentiation and concurrently inhibit osteogenesis by transferring COL1A1-targeting miRNA-92a-1–5p. J. Extracell Vesicles 2021, 10, e12056. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yu, J. MiR-129-5p suppresses gastric cancer cell invasion and proliferation by inhibiting COL1A1. Biochem. Cell Biol. 2018, 96, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Lu, G.; Mao, H.; Zhou, S.; Tong, X.; Wu, J.; Sun, Q.; Xu, H.; Fang, F. miR-133b Suppresses Invasion and Migration of Gastric Cancer Cells via the COL1A1/TGF-Î Axis. OncoTargets Ther. 2020, 13, 7985–7995. [Google Scholar] [CrossRef]
- He, B.; Lin, X.; Tian, F.; Yu, W.; Qiao, B. MiR-133a-3p Inhibits Oral Squamous Cell Carcinoma (OSCC) Proliferation and Invasion by Suppressing COL1A1. J. Cell Biochem. 2018, 119, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Du, L.; Li, X.; Zhang, X.; Gao, Y. miR-133a-3p suppresses cell proliferation, migration, and invasion and promotes apoptosis in esophageal squamous cell carcinoma. J. Cell Physiol. 2019, 234, 12757–12770. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Rao, X.; Yao, W.; Zou, X. Downregulation of MiR-196b-5p impedes cell proliferation and metastasis in breast cancer through regulating COL1A1. Am. J. Transl. Res. 2018, 10, 3122–3132. [Google Scholar]
- Lu, Q.; Guo, Z.; Xie, W.; Jin, W.; Zhu, D.; Chen, S.; Ren, T. The lncRNA H19 Mediates Pulmonary Fibrosis by Regulating the miR-196a/COL1A1 Axis. Inflammation 2018, 41, 896–903. [Google Scholar] [CrossRef]
- Ito, S.; Nagata, K. Mutants of collagen-specific molecular chaperone Hsp47 causing osteogenesis imperfecta are structurally unstable with weak binding affinity to collagen. Biochem. Biophys. Res. Commun. 2016, 469, 437–442. [Google Scholar] [CrossRef]
- Yamauchi, M.; Noyes, C.; Kuboki, Y.; Mechanic, G.L. Collagen structural microheterogeneity and a possible role for glycosylated hydroxylysine in type I collagen. Proc. Natl. Acad. Sci. USA 1982, 79, 7684–7688. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Yu, F.; Di, M.; Li, M.; Chen, Y.; Zhang, Y.; Liu, X.; Huang, X.; Zhang, M. MicroRNA-124-3p inhibits collagen synthesis in atherosclerotic plaques by targeting prolyl 4-hydroxylase subunit alpha-1 (P4HA1) in vascular smooth muscle cells. Atherosclerosis 2018, 277, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.; Lees, J.F.; Bulleid, N.J. Protein disulfide isomerase acts as a molecular chaperone during the assembly of procollagen. J. Biol. Chem. 1998, 273, 9637–9643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.; Sun, S.; Zhang, X.Q.; Zhang, P.D.; Ho, A.S.; Kiang, K.M.; Fung, C.F.; Lui, W.M.; Leung, G.K. MicroRNA-210 and Endoplasmic Reticulum Chaperones in the Regulation of Chemoresistance in Glioblastoma. J. Cancer 2015, 6, 227–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Xiang, H.; Cong, W.; Yang, H.; Zhang, G.; Wang, Y.; Guo, Z.; Shen, Y.; Chen, B. PLOD1, a target of miR-34c, contributes to cell growth and metastasis via repressing LATS1 phosphorylation and inactivating Hippo pathway in osteosarcoma. Biochem. Biophys. Res. Commun. 2020, 527, 29–36. [Google Scholar] [CrossRef]
- Huang, J.; Zhou, X.H.; Huang, S.; Li, S.; Wu, T.T. Involvement of microRNA-124 in biological behaviors of laryngeal carcinoma via PLOD2 signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 4855–4862. [Google Scholar]
- Kurozumi, A.; Kato, M.; Goto, Y.; Matsushita, R.; Nishikawa, R.; Okato, A.; Fukumoto, I.; Ichikawa, T.; Seki, N. Regulation of the collagen cross-linking enzymes LOXL2 and PLOD2 by tumor-suppressive microRNA-26a/b in renal cell carcinoma. Int. J. Oncol. 2016, 48, 1837–1846. [Google Scholar] [CrossRef] [Green Version]
- Amodio, G.; Sasso, E.; D’Ambrosio, C.; Scaloni, A.; Moltedo, O.; Franceschelli, S.; Zambrano, N.; Remondelli, P. Identification of a microRNA (miR-663a) induced by ER stress and its target gene PLOD3 by a combined microRNome and proteome approach. Cell Biol. Toxicol. 2016, 32, 285–303. [Google Scholar] [CrossRef]
- Schegg, B.; Hülsmeier, A.J.; Rutschmann, C.; Maag, C.; Hennet, T. Core glycosylation of collagen is initiated by two beta(1-O)galactosyltransferases. Mol. Cell Biol. 2009, 29, 943–952. [Google Scholar] [CrossRef] [Green Version]
- Hennet, T. Collagen glycosylation. Curr. Opin. Struct. Biol. 2019, 56, 131–138. [Google Scholar] [CrossRef]
- Dall’Olio, F.; Trinchera, M. Epigenetic Bases of Aberrant Glycosylation in Cancer. Int. J. Mol. Sci. 2017, 18, 998. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, N.; Kinoshita, T.; Nohata, N.; Yoshino, H.; Itesako, T.; Fujimura, L.; Mitsuhashi, A.; Usui, H.; Enokida, H.; Nakagawa, M.; et al. Tumor-suppressive microRNA-29a inhibits cancer cell migration and invasion via targeting HSP47 in cervical squamous cell carcinoma. Int. J. Oncol. 2013, 43, 1855–1863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Ghazwani, M.; Li, J.; Sun, M.; Stolz, D.B.; He, F.; Fan, J.; Xie, W.; Li, S. MiR-29b inhibits collagen maturation in hepatic stellate cells through down-regulating the expression of HSP47 and lysyl oxidase. Biochem. Biophys. Res. Commun. 2014, 446, 940–944. [Google Scholar] [CrossRef] [Green Version]
- Yamada, Y.; Sugawara, S.; Arai, T.; Kojima, S.; Kato, M.; Okato, A.; Yamazaki, K.; Naya, Y.; Ichikawa, T.; Seki, N. Molecular pathogenesis of renal cell carcinoma: Impact of the anti-tumor miR-29 family on gene regulation. Int. J. Urol. 2018, 25, 953–965. [Google Scholar] [CrossRef] [Green Version]
- Luna, C.; Li, G.; Qiu, J.; Epstein, D.L.; Gonzalez, P. Role of miR-29b on the regulation of the extracellular matrix in human trabecular meshwork cells under chronic oxidative stress. Mol. Vis. 2009, 15, 2488–2497. [Google Scholar]
- Wu, X.; Liu, T.; Fang, O.; Leach, L.J.; Hu, X.; Luo, Z. miR-194 suppresses metastasis of non-small cell lung cancer through regulating expression of BMP1 and p27(kip1). Oncogene 2014, 33, 1506–1514. [Google Scholar] [CrossRef]
- Tong, Y.; Wang, M.; Dai, Y.; Bao, D.; Zhang, J.; Pan, H. LncRNA HOXA-AS3 Sponges miR-29c to Facilitate Cell Proliferation, Metastasis, and EMT Process and Activate the MEK/ERK Signaling Pathway in Hepatocellular Carcinoma. Hum. Gene Ther. Clin. Dev. 2019, 30, 129–141. [Google Scholar] [CrossRef]
- Ushiki, T. Collagen fibers, reticular fibers and elastic fibers. A comprehensive understanding from a morphological viewpoint. Arch. Histol. Cytol. 2002, 65, 109–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.Z.; Xu, X.; Ning, L.F.; Jiang, W.Y.; Xing, C.; Tang, Q.Q.; Huang, H.Y. miR-27 impairs the adipogenic lineage commitment via targeting lysyl oxidase. Obesity 2015, 23, 2445–2453. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.L.; Tsai, M.C.; Chang, Y.H.; Wang, C.C.; Chu, P.Y.; Lin, H.Y.; Huang, Y.H. MIR29A Impedes Metastatic Behaviors in Hepatocellular Carcinoma via Targeting LOX, LOXL2, and VEGFA. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Muther, C.; Jobeili, L.; Garion, M.; Heraud, S.; Thepot, A.; Damour, O.; Lamartine, J. An expression screen for aged-dependent microRNAs identifies miR-30a as a key regulator of aging features in human epidermis. Aging 2017, 9, 2376–2396. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Shi, L.; Li, X.; Wang, X.; Wang, H.; Liu, Y. RNF144A-AS1, a TGF-Î1- and hypoxia-inducible gene that promotes tumor metastasis and proliferation via targeting the miR-30c-2-3p/LOX axis in gastric cancer. Cell Biosci. 2021, 11, 177. [Google Scholar] [CrossRef]
- Saatci, O.; Kaymak, A.; Raza, U.; Ersan, P.G.; Akbulut, O.; Banister, C.E.; Sikirzhytski, V.; Tokat, U.M.; Aykut, G.; Ansari, S.A.; et al. Targeting lysyl oxidase (LOX) overcomes chemotherapy resistance in triple negative breast cancer. Nat. Commun. 2020, 11, 2416. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.; Li, L.; Li, Y. Involvement of miR-30b in kynurenine-mediated lysyl oxidase expression. J. Physiol Biochem 2019, 75, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Shi, E.; Gu, T.; Tang, R.; Gao, S.; Wang, Y.; Liu, H. Overexpression of microRNA-30a contributes to the development of aortic dissection by targeting lysyl oxidase. J. Thorac. Cardiovasc. Surg. 2017, 154, 1862–1869. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Shen, J.X.; Wu, H.T.; Li, X.L.; Wen, X.F.; Du, C.W.; Zhang, G.J. Collagen 1A1 (COL1A1) promotes metastasis of breast cancer and is a potential therapeutic target. Discov. Med. 2018, 25, 211–223. [Google Scholar] [PubMed]
- Hou, L.; Lin, T.; Wang, Y.; Liu, B.; Wang, M. Collagen type 1 alpha 1 chain is a novel predictive biomarker of poor progression-free survival and chemoresistance in metastatic lung cancer. J. Cancer 2021, 12, 5723–5731. [Google Scholar] [CrossRef]
- Lv, H.; Sun, Y.; Zhang, Y. MiR-133 is Involved in Estrogen Deficiency-Induced Osteoporosis through Modulating Osteogenic Differentiation of Mesenchymal Stem Cells. Med. Sci. Monit. 2015, 21, 1527–1534. [Google Scholar] [PubMed] [Green Version]
- Viguet-Carrin, S.; Garnero, P.; Delmas, P.D. The role of collagen in bone strength. Osteoporos Int. 2006, 17, 319–336. [Google Scholar] [CrossRef]
- Shi, R.; Gao, S.; Zhang, J.; Xu, J.; Graham, L.M.; Yang, X.; Li, C. Collagen prolyl 4-hydroxylases modify tumor progression. Acta Biochim. Biophys. Sin. 2021, 53, 805–814. [Google Scholar] [CrossRef]
- Li, J.; Ghazwani, M.; Zhang, Y.; Lu, J.; Li, J.; Fan, J.; Gandhi, C.R.; Li, S. miR-122 regulates collagen production via targeting hepatic stellate cells and suppressing P4HA1 expression. J. Hepatol. 2013, 58, 522–528. [Google Scholar] [CrossRef] [Green Version]
- Seeliger, C.; Karpinski, K.; Haug, A.T.; Vester, H.; Schmitt, A.; Bauer, J.S.; Van Griensven, M. Five freely circulating miRNAs and bone tissue miRNAs are associated with osteoporotic fractures. J. Bone Miner. Res. 2014, 29, 1718–1728. [Google Scholar] [CrossRef]
- Feng, G.X.; Li, J.; Yang, Z.; Zhang, S.Q.; Liu, Y.X.; Zhang, W.Y.; Ye, L.H.; Zhang, X.D. Hepatitis B virus X protein promotes the development of liver fibrosis and hepatoma through downregulation of miR-30e targeting P4HA2 mRNA. Oncogene 2017, 36, 6895–6905. [Google Scholar] [CrossRef] [Green Version]
- Aghajanian, P.; Hall, S.; Wongworawat, M.D.; Mohan, S. The Roles and Mechanisms of Actions of Vitamin C in Bone: New Developments. J. Bone Miner. Res. 2015, 30, 1945–1955. [Google Scholar] [CrossRef] [Green Version]
- Nagata, K.; Saga, S.; Yamada, K.M. A major collagen-binding protein of chick embryo fibroblasts is a novel heat shock protein. J. Cell Biol. 1986, 103, 223–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trackman, P.C. Enzymatic and non-enzymatic functions of the lysyl oxidase family in bone. Matrix Biol 2015, 52, 7–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Y.; Xu, R. Roles of PLODs in Collagen Synthesis and Cancer Progression. Front. Cell Dev. Biol. 2018, 6, 66. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Shi, Y.; Zheng, L.; Zhou, B.; Inose, H.; Wang, J.; Guo, X.E.; Grosschedl, R.; Karsenty, G. miR-34s inhibit osteoblast proliferation and differentiation in the mouse by targeting SATB2. J. Cell Biol. 2012, 197, 509–521. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Holmstrøm, K.; Qiu, W.; Ditzel, N.; Shi, K.; Hokland, L.; Kassem, M. MicroRNA-34a inhibits osteoblast differentiation and in vivo bone formation of human stromal stem cells. Stem. Cells 2014, 32, 902–912. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Zhao, C.; Zhang, P.; Liu, Y.; Jiang, Y.; Wu, E.; Xue, H.; Liu, C.; Li, Z. miR-26b modulates OA induced BMSC osteogenesis through regulating GSK3beta-catenin pathway. Exp. Mol. Pathol. 2019, 107, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Gelse, K.; Pöschl, E.; Aigner, T. Collagens-structure, function, and biosynthesis. Adv. Drug. Deliv. Rev. 2003, 55, 1531–1546. [Google Scholar] [CrossRef] [Green Version]
- Sornay-Rendu, E.; Boutroy, S.; Munoz, F.; Delmas, P.D. Alterations of cortical and trabecular architecture are associated with fractures in postmenopausal women, partially independent of decreased BMD measured by DXA: The OFELY study. J. Bone Miner. Res. 2007, 22, 425–433. [Google Scholar] [CrossRef]
- Sroga, G.E.; Vashishth, D. Effects of bone matrix proteins on fracture and fragility in osteoporosis. Curr. Osteoporos Rep. 2012, 10, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Bottomley, M.J.; Batten, M.R.; Lumb, R.A.; Bulleid, N.J. Quality control in the endoplasmic reticulum: PDI mediates the ER retention of unassembled procollagen C-propeptides. Curr. Biol. 2001, 11, 1114–1118. [Google Scholar] [CrossRef]
- Sharma, U.; Carrique, L.; Vadon-Le Goff, S.; Mariano, N.; Georges, R.N.; Delolme, F.; Koivunen, P.; Myllyharju, J.; Moali, C.; Aghajari, N.; et al. Structural basis of homo- and heterotrimerization of collagen I. Nat. Commun. 2017, 8, 14671. [Google Scholar] [CrossRef]
- Koski, M.K.; Anantharajan, J.; Kursula, P.; Dhavala, P.; Murthy, A.V.; Bergmann, U.; Myllyharju, J.; Wierenga, R.K. Assembly of the elongated collagen prolyl 4-hydroxylase α2β2 heterotetramer around a central α2 dimer. Biochem. J. 2017, 474, 751–769. [Google Scholar] [CrossRef] [Green Version]
- Ito, S.; Nagata, K. Biology of Hsp47 (Serpin H1), a collagen-specific molecular chaperone. Semin. Cell Dev. Biol. 2017, 62, 142–151. [Google Scholar] [CrossRef]
- Ito, S.; Nagata, K. Roles of the endoplasmic reticulum-resident, collagen-specific molecular chaperone Hsp47 in vertebrate cells and human disease. J. Biol. Chem. 2019, 294, 2133–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Xiong, G.; Fu, H.; Evers, B.M.; Zhou, B.P.; Xu, R. Chaperone Hsp47 Drives Malignant Growth and Invasion by Modulating an ECM Gene Network. Cancer Res. 2015, 75, 1580–1591. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Li, Z.; Wang, Y.; Li, L.; Wang, D.; Zhang, W.; Liu, L.; Jiang, H.; Yang, J.; Cheng, J. Overexpression of miR-29b reduces collagen biosynthesis by inhibiting heat shock protein 47 during skin wound healing. Transl. Res. 2016, 178, 38–53. [Google Scholar] [CrossRef] [PubMed]
- Gorur, A.; Yuan, L.; Kenny, S.J.; Baba, S.; Xu, K.; Schekman, R. COPII-coated membranes function as transport carriers of intracellular procollagen I. J. Cell Biol. 2017, 216, 1745–1759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malhotra, V.; Erlmann, P. The pathway of collagen secretion. Annu. Rev. Cell Dev. Biol. 2015, 31, 109–124. [Google Scholar] [CrossRef]
- Hulmes, D.J.S. Roles of the procollagen C-propeptides in health and disease. Essays Biochem. 2019, 63, 313–323. [Google Scholar]
- Rosell-Garcia, T.; Paradela, A.; Bravo, G.; Dupont, L.; Bekhouche, M.; Colige, A.; Rodriguez-Pascual, F. Differential cleavage of lysyl oxidase by the metalloproteinases BMP1 and ADAMTS2/14 regulates collagen binding through a tyrosine sulfate domain. J. Biol. Chem. 2019, 294, 11087–11100. [Google Scholar] [CrossRef]
- Li, Z.; Hassan, M.Q.; Jafferji, M.; Aqeilan, R.I.; Garzon, R.; Croce, C.M.; Van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; Lian, J.B. Biological functions of miR-29b contribute to positive regulation of osteoblast differentiation. J. Biol. Chem. 2009, 284, 15676–15684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNerny, E.M.B.; Gardinier, J.D.; Kohn, D.H. Exercise increases pyridinoline cross-linking and counters the mechanical effects of concurrent lathyrogenic treatment. Bone 2015, 81, 327–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, T.; Stoop, R.; Sakai, T.; Hess, A.; Deutzmann, R.; Schlotzer-Schrehardt, U.; Chu, M.L.; Von der Mark, K. Loss of fibulin-4 results in abnormal collagen fibril assembly in bone, caused by impaired lysyl oxidase processing and collagen cross-linking. Matrix Biol. 2016, 50, 53–66. [Google Scholar] [CrossRef]
- Bird, T.A.; Levene, C.I. Lysyl oxidase: Evidence that pyridoxal phosphate is a cofactor. Biochem. Biophys. Res. Commun. 1982, 108, 1172–1180. [Google Scholar] [CrossRef]
- Wang, S.X.; Mure, M.; Medzihradszky, K.F.; Burlingame, A.L.; Brown, D.E.; Dooley, D.M.; Smith, A.J.; Kagan, H.M.; Klinman, J.P. A crosslinked cofactor in lysyl oxidase: Redox function for amino acid side chains. Science 1996, 273, 1078–1084. [Google Scholar] [CrossRef]
- Nagaoka, H.; Mochida, Y.; Atsawasuwan, P.; Kaku, M.; Kondoh, T.; Yamauchi, M. 1,25(OH)2D3 regulates collagen quality in an osteoblastic cell culture system. Biochem. Biophys. Res. Commun. 2008, 377, 674–678. [Google Scholar] [CrossRef]
- Depalle, B.; Duarte, A.G.; Fiedler, I.A.K.; Pujo-Menjouet, L.; Buehler, M.J.; Berteau, J.P. The different distribution of enzymatic collagen cross-links found in adult and children bone result in different mechanical behavior of collagen. Bone 2018, 110, 107–114. [Google Scholar] [CrossRef] [Green Version]
- You, L.; Pan, L.; Chen, L.; Gu, W.; Chen, J. MiR-27a is Essential for the Shift from Osteogenic Differentiation to Adipogenic Differentiation of Mesenchymal Stem Cells in Postmenopausal Osteoporosis. Cell Physiol. Biochem. 2016, 39, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Zhu, S.; Li, X.; Weng, J.; Chen, S. miR-27b-3p Suppressed Osteogenic Differentiation of Maxillary Sinus Membrane Stem Cells by Targeting Sp7. Implant. Dent. 2017, 26, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.; Peng, Y.T.; Guo, P. MiR-29a promotes osteogenic differentiation of mesenchymal stem cells via targeting HDAC4. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3318–3326. [Google Scholar] [PubMed]
- Gauthier, R.; Follet, H.; Langer, M.; Gineyts, E.; Rongieras, F.; Peyrin, F.; Mitton, D. Relationships between human cortical bone toughness and collagen cross-links on paired anatomical locations. Bone 2018, 112, 202–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collier, T.A.; Nash, A.; Birch, H.L.; De Leeuw, N.H. Intra-molecular lysine-arginine derived advanced glycation end-product cross-linking in Type I collagen: A molecular dynamics simulation study. Biophys. Chem 2016, 218, 42–46. [Google Scholar] [CrossRef] [Green Version]
- Poundarik, A.A.; Wu, P.C.; Evis, Z.; Sroga, G.E.; Ural, A.; Rubin, M.; Vashishth, D. A direct role of collagen glycation in bone fracture. J. Mech. Behav. Biomed. Mater. 2015, 52, 120–130. [Google Scholar] [CrossRef] [Green Version]
- Depalle, B.; Qin, Z.; Shefelbine, S.J.; Buehler, M.J. Influence of cross-link structure, density and mechanical properties in the mesoscale deformation mechanisms of collagen fibrils. J. Mech. Behav. Biomed. Mater. 2015, 52, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, F.N.; Zimmermann, E.A.; Campbell, G.M.; Sroga, G.E.; Puschel, K.; Amling, M.; Tang, S.Y.; Vashishth, D.; Busse, B. Assessment of collagen quality associated with non-enzymatic cross-links in human bone using Fourier-transform infrared imaging. Bone 2017, 97, 243–251. [Google Scholar] [CrossRef] [Green Version]
- Stock, S.R. The Mineral-Collagen Interface in Bone. Calcif. Tissue Int. 2015, 97, 262–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, S.; Poundarik, A.A.; Vashishth, D. Do Non-collagenous Proteins Affect Skeletal Mechanical Properties? Calcif. Tissue Int. 2015, 97, 281–291. [Google Scholar] [CrossRef]
- Icer, M.A.; Gezmen-Karadag, M. The multiple functions and mechanisms of osteopontin. Clin. Biochem. 2018, 59, 17–24. [Google Scholar] [CrossRef]
- Bailey, S.; Karsenty, G.; Gundberg, C.; Vashishth, D. Osteocalcin and osteopontin influence bone morphology and mechanical properties. Ann. N. Y. Acad. Sci. 2017, 1409, 79–84. [Google Scholar] [CrossRef]
- Rodriguez, D.E.; Thula-Mata, T.; Toro, E.J.; Yeh, Y.W.; Holt, C.; Holliday, L.S.; Gower, L.B. Multifunctional role of osteopontin in directing intrafibrillar mineralization of collagen and activation of osteoclasts. Acta Biomater. 2014, 10, 494–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Gill, G.; Kaur, H.; Amhmed, M.; Jakhu, H. Role of osteopontin in bone remodeling and orthodontic tooth movement: A review. Prog. Orthod. 2018, 19, 18. [Google Scholar] [CrossRef]
- Tu, M.; Li, Y.; Zeng, C.; Deng, Z.; Gao, S.; Xiao, W.; Luo, W.; Jiang, W.; Li, L.; Lei, G. MicroRNA-127-5p regulates osteopontin expression and osteopontin-mediated proliferation of human chondrocytes. Sci. Rep. 2016, 6, 25032. [Google Scholar] [CrossRef] [Green Version]
- Song, K.; Liu, N.; Yang, Y.; Qiu, X. Regulation of osteosarcoma cell invasion through osteopontin modification by miR-4262. Tumour. Biol. 2016, 37, 6493–6499. [Google Scholar] [CrossRef] [PubMed]
- Calabro, N.E.; Kristofik, N.J.; Kyriakides, T.R. Thrombospondin-2 and extracellular matrix assembly. Biochim. Biophys. Acta 2014, 1840, 2396–2402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alford, A.I.; Golicz, A.Z.; Cathey, A.L.; Reddy, A.B. Thrombospondin-2 facilitates assembly of a type-I collagen-rich matrix in marrow stromal cells undergoing osteoblastic differentiation. Connect. Tissue Res. 2013, 54, 275–282. [Google Scholar] [CrossRef] [Green Version]
- Manley, E., Jr.; Perosky, J.E.; Khoury, B.M.; Reddy, A.B.; Kozloff, K.M.; Alford, A.I. Thrombospondin-2 deficiency in growing mice alters bone collagen ultrastructure and leads to a brittle bone phenotype. J. Appl. Physiol. 2015, 119, 872–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Li, H.; Ma, Y.; Zhu, X.; Zhang, S.; Li, J. MiR-221-3p is down-regulated in preeclampsia and affects trophoblast growth, invasion and migration partly via targeting thrombospondin 2. Biomed. Pharmacother. 2019, 109, 127–134. [Google Scholar] [CrossRef]
- Sun, X.Y.; Han, X.M.; Zhao, X.L.; Cheng, X.M.; Zhang, Y. MiR-93-5p promotes cervical cancer progression by targeting THBS2/MMPS signal pathway. Eur Rev. Med. Pharmacol. Sci. 2019, 23, 5113–5121. [Google Scholar]
- Qian, Z.; Gong, L.; Mou, Y.; Han, Y.; Zheng, S. MicroRNA-203a-3p is a candidate tumor suppressor that targets thrombospondin 2 in colorectal carcinoma. Oncol. Rep. 2019, 42, 1825–1832. [Google Scholar] [CrossRef]
- Zhang, Y.; Wei, Q.S.; Ding, W.B.; Zhang, L.L.; Wang, H.C.; Zhu, Y.J.; He, W.; Chai, Y.N.; Liu, Y.W. Increased microRNA-93-5p inhibits osteogenic differentiation by targeting bone morphogenetic protein-2. PLoS ONE. 2017, 12, e0182678. [Google Scholar] [CrossRef] [Green Version]
- Fan, F.Y.; Deng, R.; Qiu, L.; Wen, Q.; Zeng, Y.; Gao, L.; Zhang, C.; Kong, P.; Zhong, J.; Zeng, N.; et al. miR-203a-3p.1 is involved in the regulation of osteogenic differentiation by directly targeting Smad9 in MM-MSCs. Oncol Lett. 2019, 18, 6339–6346. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Gao, Y.; Cai, L.; Li, F.; Lou, Y.; Xu, N.; Kang, Y.; Yang, H. MicroRNA-221 is involved in the regulation of osteoporosis through regulates RUNX2 protein expression and osteoblast differentiation. Am. J. Transl. Res. 2017, 9, 126–135. [Google Scholar]
- Balooch, G.; Balooch, M.; Nalla, R.K.; Schilling, S.; Filvaroff, E.H.; Marshall, G.W.; Marshall, S.J.; Ritchie, R.O.; Derynck, R.; Alliston, T. TGF-beta regulates the mechanical properties and composition of bone matrix. Proc. Natl. Acad. Sci. USA 2005, 102, 18813–18818. [Google Scholar] [CrossRef] [Green Version]
- Garcia, J.; Delany, A.M. MicroRNAs regulating TGFÎ and BMP signaling in the osteoblast lineage. Bone 2021, 143, 115791. [Google Scholar] [CrossRef] [PubMed]
- Corsi, A.; Xu, T.; Chen, X.D.; Boyde, A.; Liang, J.; Mankani, M.; Sommer, B.; Iozzo, R.V.; Eichstetter, I.; Robey, P.G.; et al. Phenotypic effects of biglycan deficiency are linked to collagen fibril abnormalities, are synergized by decorin deficiency, and mimic Ehlers-Danlos-like changes in bone and other connective tissues. J. Bone Miner. Res. 2002, 17, 1180–1189. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.L.; Bai, Y.M.; Zhao, B.Y.; Wang, Q.H.; Zhang, H.S. Silencing of miR-330-5p stimulates osteogenesis in bone marrow mesenchymal stem cells and inhibits bone loss in osteoporosis by activating Bgn-mediated BMP/Smad pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 4095–4102. [Google Scholar] [PubMed]
- Bhushan, R.; Grünhagen, J.; Becker, J.; Robinson, P.N.; Ott, C.E.; Knaus, P. miR-181a promotes osteoblastic differentiation through repression of TGF-Î signaling molecules. Int. J. Biochem Cell Biol. 2013, 45, 696–705. [Google Scholar] [CrossRef] [PubMed]
- Kwan, P.; Ding, J.; Tredget, E.E. MicroRNA 181b regulates decorin production by dermal fibroblasts and may be a potential therapy for hypertrophic scar. PLoS ONE 2015, 10, e0123054. [Google Scholar] [CrossRef] [Green Version]
- Hensley, A.P.; McAlinden, A. The role of microRNAs in bone development. Bone 2021, 143, 115760. [Google Scholar] [CrossRef]
- Machado do Reis, L.; Kessler, C.B.; Adams, D.J.; Lorenzo, J.; Jorgetti, V.; Delany, A.M. Accentuated osteoclastic response to parathyroid hormone undermines bone mass acquisition in osteonectin-null mice. Bone 2008, 43, 264–273. [Google Scholar] [CrossRef] [Green Version]
- Thurner, P.J.; Chen, C.G.; Ionova-Martin, S.; Sun, L.; Harman, A.; Porter, A.; Ager, J.W. , III; Ritchie, R.O.; Alliston, T. Osteopontin deficiency increases bone fragility but preserves bone mass. Bone 2010, 46, 1564–1573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapinas, K.; Kessler, C.B.; Delany, A.M. miR-29 suppression of osteonectin in osteoblasts: Regulation during differentiation and by canonical Wnt signaling. J. Cell Biochem. 2009, 108, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Dole, N.S.; Kapinas, K.; Kessler, C.B.; Yee, S.P.; Adams, D.J.; Pereira, R.C.; Delany, A.M. A single nucleotide polymorphism in osteonectin 3′ untranslated region regulates bone volume and is targeted by miR-433. J. Bone Miner. Res. 2015, 30, 723–732. [Google Scholar] [CrossRef]
- Wei, J.; Chen, H.; Fu, Y.; Zhang, B.; Zhang, L.; Tao, S.; Lin, F. Experimental study of expression profile and specific role of human microRNAs in regulating atrophic bone nonunion. Medicine 2020, 99, e21653. [Google Scholar] [CrossRef]
- Moura, S.R.; Bras, J.P.; Freitas, J.; Osório, H.; Barbosa, M.A.; Santos, S.G.; Almeida, M.I. miR-99a in bone homeostasis: Regulating osteogenic lineage commitment and osteoclast differentiation. Bone 2020, 134, 115303. [Google Scholar] [CrossRef]
- Zheng, C.; Bai, C.; Sun, Q.; Zhang, F.; Yu, Q.; Zhao, X.; Kang, S.; Li, J.; Jia, Y. Long noncoding RNA XIST regulates osteogenic differentiation of human bone marrow mesenchymal stem cells by targeting miR-9-5p. Mech. Dev. 2020, 162, 103612. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Bian, M.; Zhou, Z.; Wu, X.; Ge, X.; Xiao, T.; Yu, J. Circular RNA SIPA1L1 regulates osteoblastic differentiation of stem cells from apical papilla via miR-204-5p/ALPL pathway. Stem. Cell Res. Ther. 2020, 11, 461. [Google Scholar] [CrossRef]
- Bini, F.; Pica, A.; Marinozzi, A.; Marinozzi, F. A 3D Model of the Effect of Tortuosity and Constrictivity on the Diffusion in Mineralized Collagen Fibril. Sci. Rep. 2019, 9, 2658. [Google Scholar] [CrossRef]
- Depalle, B.; Qin, Z.; Shefelbine, S.J.; Buehler, M.J. Large Deformation Mechanisms, Plasticity, and Failure of an Individual Collagen Fibril with Different Mineral Content. J. Bone Miner. Res. 2016, 31, 380–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jager, I.; Fratzl, P. Mineralized collagen fibrils: A mechanical model with a staggered arrangement of mineral particles. Biophys. J. 2000, 79, 1737–1746. [Google Scholar] [CrossRef] [Green Version]
- Landis, W.J.; Song, M.J.; Leith, A.; McEwen, L.; McEwen, B.F. Mineral and organic matrix interaction in normally calcifying tendon visualized in three dimensions by high-voltage electron microscopic tomography and graphic image reconstruction. J. Struct. Biol. 1993, 110, 39–54. [Google Scholar] [CrossRef] [PubMed]
- Murshed, M. Mechanism of Bone Mineralization. Cold Spring Harb. Perspect. Med. 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Anh, D.J.; Dimai, H.P.; Hall, S.L.; Farley, J.R. Skeletal alkaline phosphatase activity is primarily released from human osteoblasts in an insoluble form, and the net release is inhibited by calcium and skeletal growth factors. Calcif. Tissue Int. 1998, 62, 332–340. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhang, L.; Tong, X.; Zhang, M.; Zhao, Y.; Guo, J.; Lei, L.; Chen, X.; Tickner, J.; Xu, J.; et al. Mechanical Stress Regulates Bone Metabolism Through MicroRNAs. J. Cell Physiol. 2017, 232, 1239–1245. [Google Scholar] [CrossRef]
- Robling, A.G.; Turner, C.H. Mechanical signaling for bone modeling and remodeling. Crit. Rev. Eukaryot Gene Expr. 2009, 19, 319–338. [Google Scholar] [CrossRef] [Green Version]
- Qi, Z.; Liu, W.; Lu, J. The mechanisms underlying the beneficial effects of exercise on bone remodeling: Roles of bone-derived cytokines and microRNAs. Prog. Biophys. Mol. Biol. 2016, 122, 131–139. [Google Scholar] [CrossRef]



{kind=link}
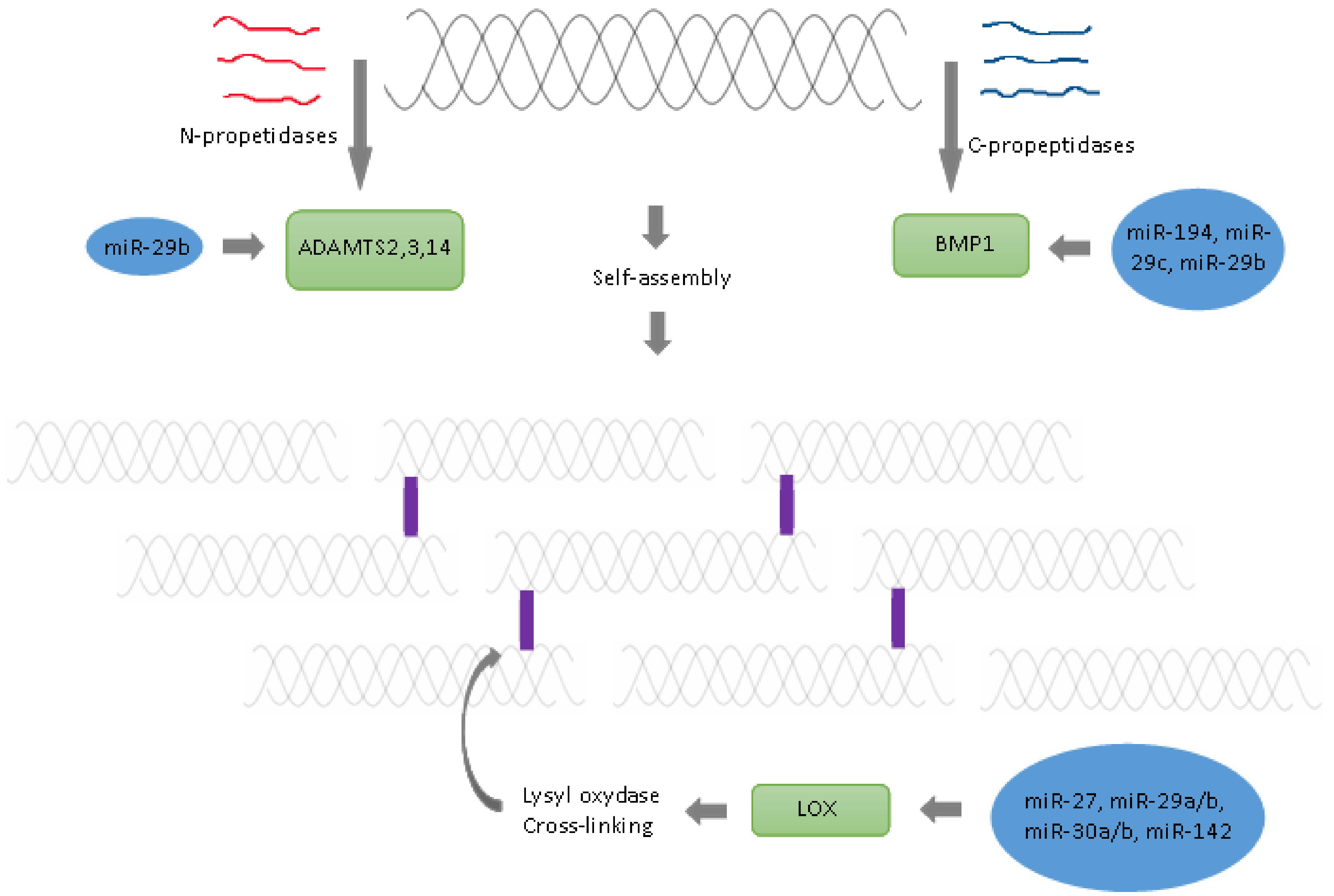{kind=link}
| Intracellular Stage of Collagen Formation | Enzyme | Gene | miRNA | References | |
|---|---|---|---|---|---|
| 1. | COL1A1 gene expression | − | COL1A1 | miR-625-5p, miR-92a-1-5p, miR-129-5p, miR-133b, miR-133a-3p, miR-196b-5p, miR-196a, miR-98, miR-29a, miR-513b-5p | [12,13,15,16,17,18,23,24,25,26,27,28,29] |
| 2. | Hydroxylation of proline residues | Prolyl,4- hydroxylase | P4HA1, P4HA2, P4HA3, P4HB | miR-124-3p, miR-122, miR-30e, − miR-210 | [30,31,32] [33,34] |
| 3. | Hydroxylation of lysine residues | Lysyl hydroxylase | PLOD1, PLOD2, PLOD3 | miR-34c, miR-124, miR-26, miR-663a | [35,36,37,38] |
| 4. | Glycosylation of some hydroxylysine residues | Glycosyltransferase | GLT25D1 GLT25D2 | − | [39,40,41] |
| 5. | Formation of intra-chain and inter-chain disulphide bonds in terminal peptides | Protein disulphide isomerase | P4HB | miR-210 | [33,34] |
| 6. | Procollagen triple helix formation | Hsp-47 protein | SERPINH1 | miR-29b, miR-29a | [42,43,44] |
| Extracellular stage of collagen formation | Enzyme | Gene | miRNA | Sources | |
| 7. | Cleavage of amino propeptides | Specific N-propeptidases | ADAM2,3 and 14 | miR-29b | [45] |
| 8. | Cleavage of carboxy propeptides | Specific C- propeptidase | BMP1 | miR-194, miR-29c, miR-29b | [45,46,47] |
| 9. | Aggregation of collagen fibres | Self-assembly | − | − | [48] |
| 10. | Crosslinking Oxidative deamination of epsilon-amino groups of lysyl and hydroxylysine residues located in telopeptide domains to aldehydes | Lysyl oxidase | LOX | miR-27, miR29b, miR29a, miR-30a, miR-30b, miR-30b-2-3p, miR-142-3p | [43,49,50,51,52,53,54,55] |
| Protein | Function | Gene | miRNA | References |
|---|---|---|---|---|
| TGF-β | Regulates the elastic modulus and the hardness of the bone | TGFB1 | See review | [118] |
| Decorin | Models the effects of TGF-β, participates in fibrogenesis of collagen, prevents the mineralisation of collagen [30] | DCN | miR-181b | [122] |
| Biglycan | Inhibits the effects of TGF-β | BGN | miR-330-5p | [120] |
| Osteonectin | Responsible for bone density | SPARC | miRs-29a and -29c | [127] |
| Osteopontin | Crucial for the deformability and resistance of bones to fractures, provides strong adhesion for hydroxyapatite and bone sialoprotein I (BSP-1 or BNSP) | OPN | miRNA-127-5p miR-4262 | [106, 107] |
| Thrombospondin-2 | Incorporates collagen into the insoluble cross-linked bone matrix | THBS2 | miR-221-3p miR-93-5p miR-203a-3p | [111,112,113] |
| Alkaline phosphatase | Alters the Pi/PPi ratio in the bone microenvironment to favour bone mineralisation | ALPL | hsa-miR-149, miR-99a-5p MiR-9-5p miR-204-5p | [128,129,130,131] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lehmann, T.P.; Guderska, U.; Kałek, K.; Marzec, M.; Urbanek, A.; Czernikiewicz, A.; Sąsiadek, M.; Karpiński, P.; Pławski, A.; Głowacki, M.; et al. The Regulation of Collagen Processing by miRNAs in Disease and Possible Implications for Bone Turnover. Int. J. Mol. Sci. 2022, 23, 91. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010091
Lehmann TP, Guderska U, Kałek K, Marzec M, Urbanek A, Czernikiewicz A, Sąsiadek M, Karpiński P, Pławski A, Głowacki M, et al. The Regulation of Collagen Processing by miRNAs in Disease and Possible Implications for Bone Turnover. International Journal of Molecular Sciences. 2022; 23(1):91. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010091
Chicago/Turabian StyleLehmann, Tomasz P., Urszula Guderska, Klaudia Kałek, Maria Marzec, Agnieszka Urbanek, Alicja Czernikiewicz, Maria Sąsiadek, Paweł Karpiński, Andrzej Pławski, Maciej Głowacki, and et al. 2022. "The Regulation of Collagen Processing by miRNAs in Disease and Possible Implications for Bone Turnover" International Journal of Molecular Sciences 23, no. 1: 91. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010091