
Plant Defense Responses to a Novel Plant Elicitor Candidate LY5-24-2


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Protection of A. thaliana against H. arabidopsidis by LY5-24-2
2.2. Enhancement of Accumulation of Cell Wall Components and Facilitation of Stomatal Closure by LY5-24-2
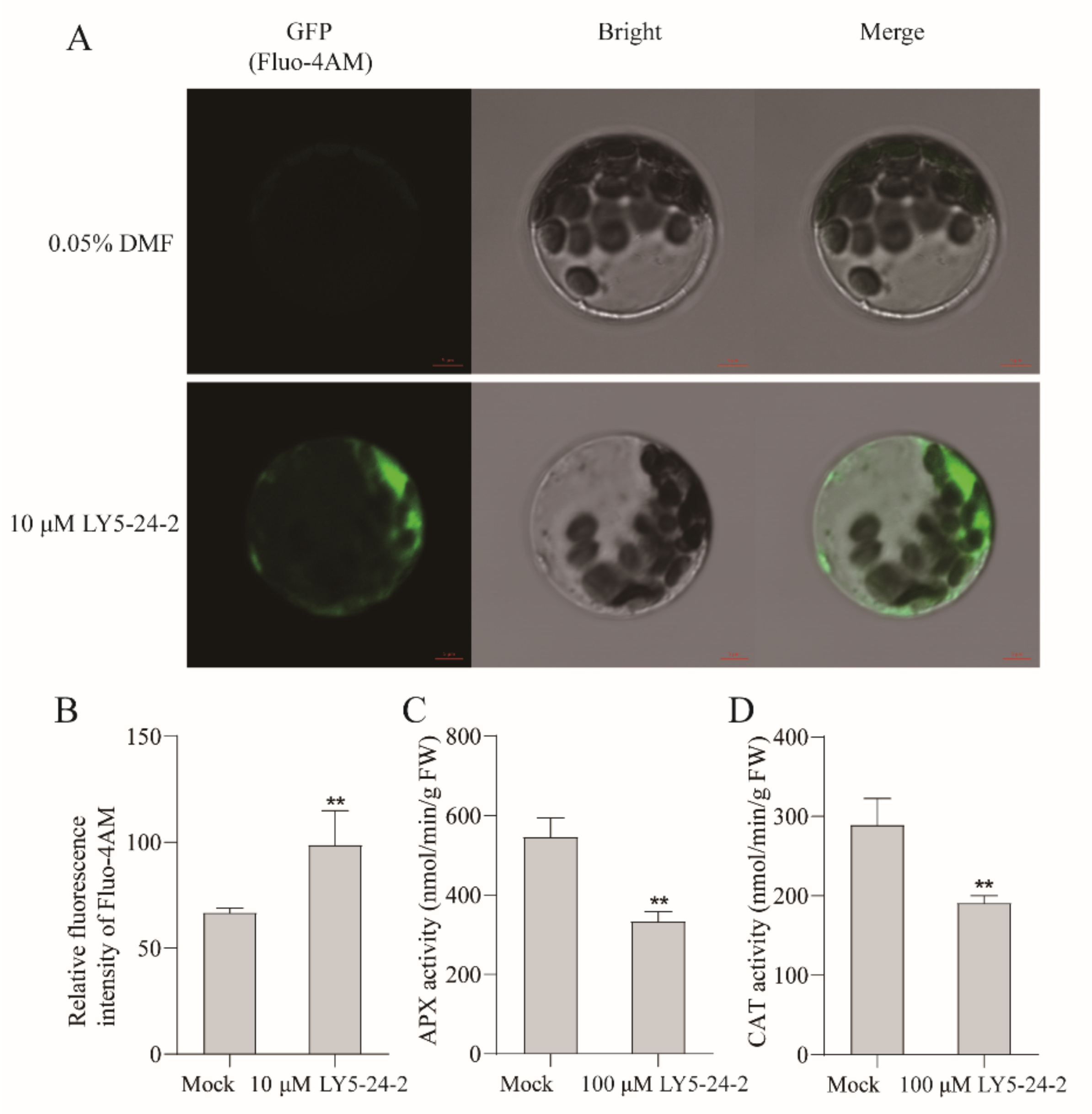
2.3. Enhancement of Transduction of Intracellular Defense Signals by LY5-24-2
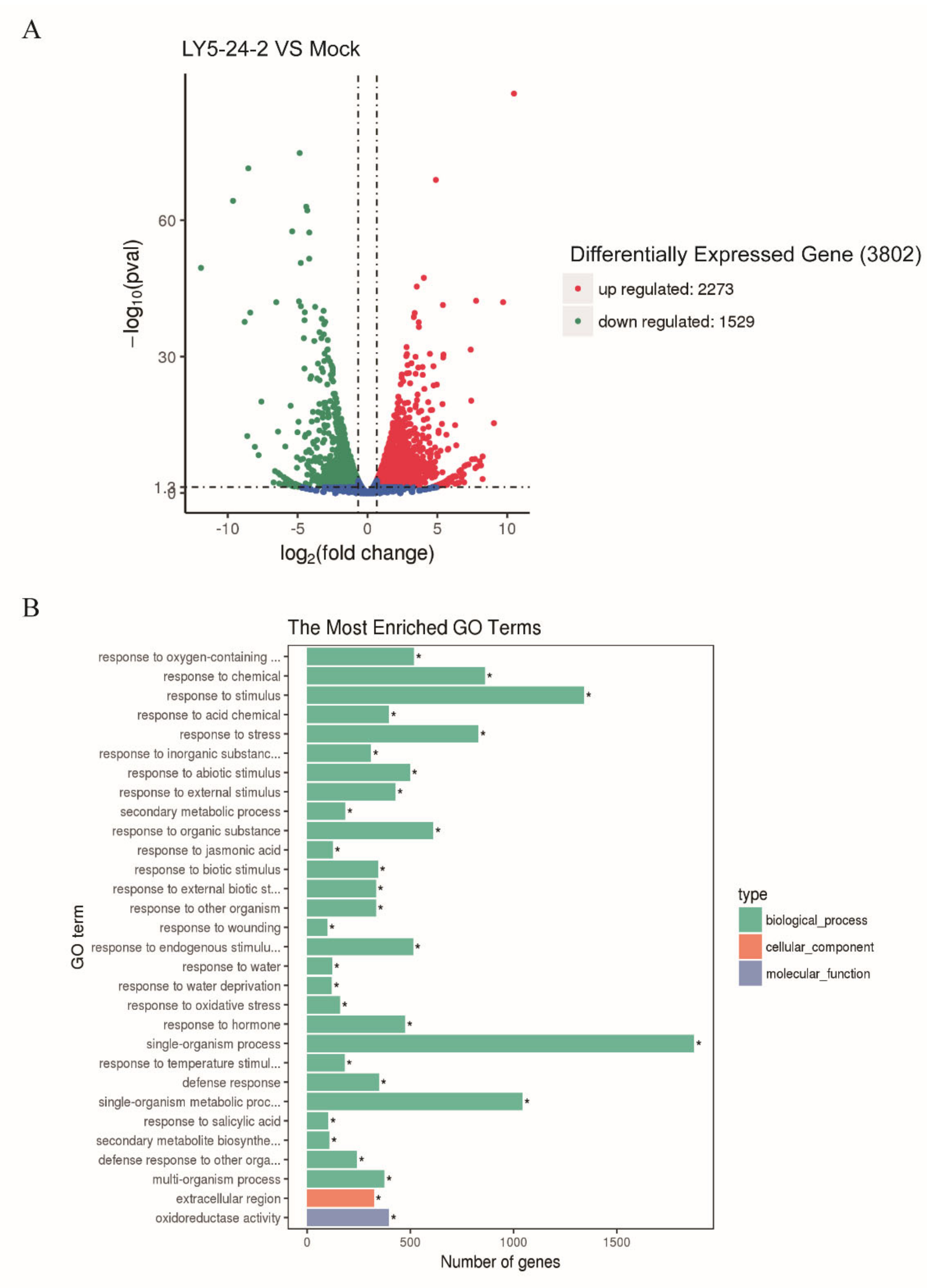
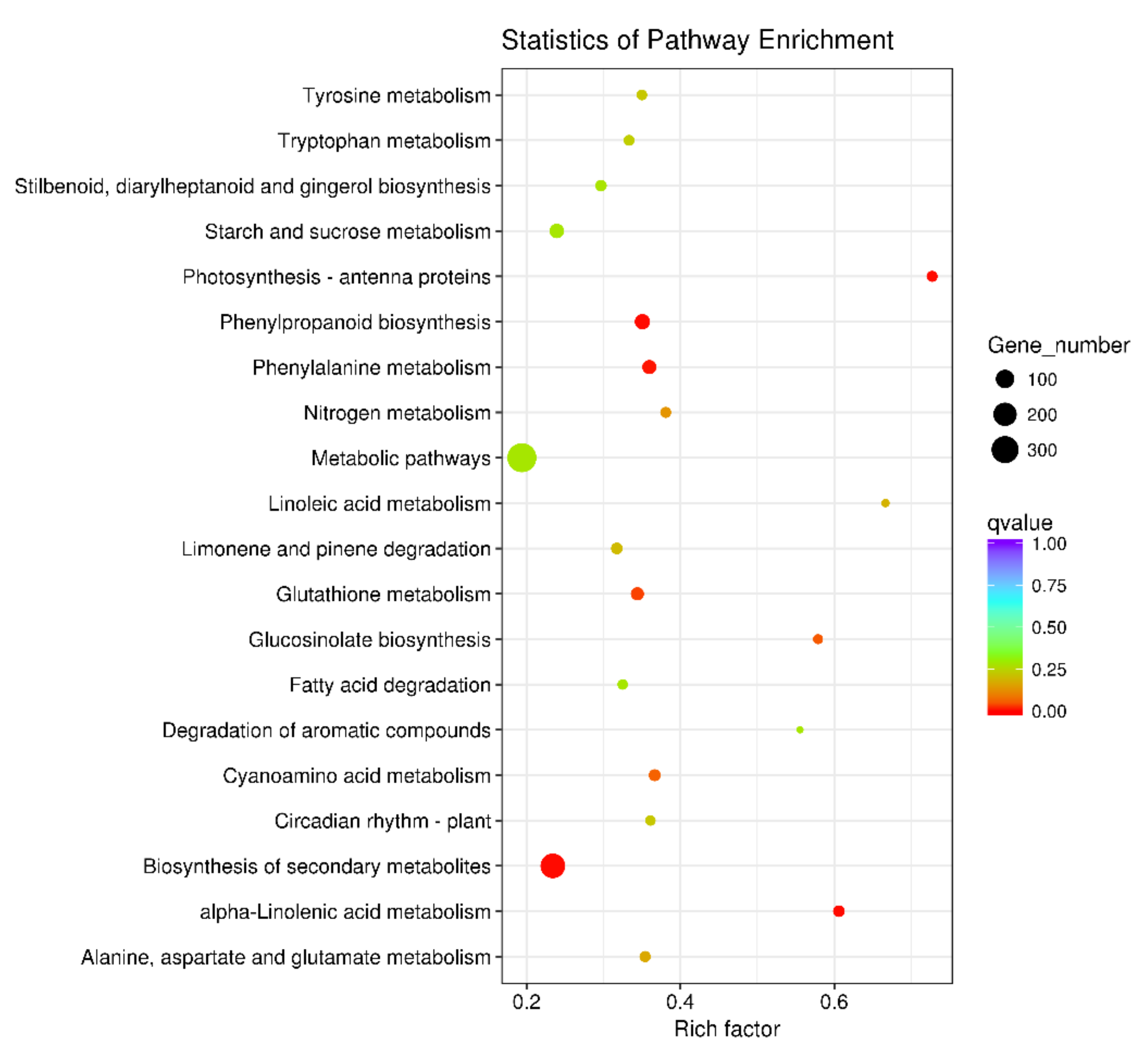
2.4. Analyses of Differentially Expressed Genes (DEGs) in A. thaliana after a LY5-24-2 Treatment
2.5. Mode of Action of LY5-24-2 on Downstream SA Signal Transduction and Dependence on AtNPR1
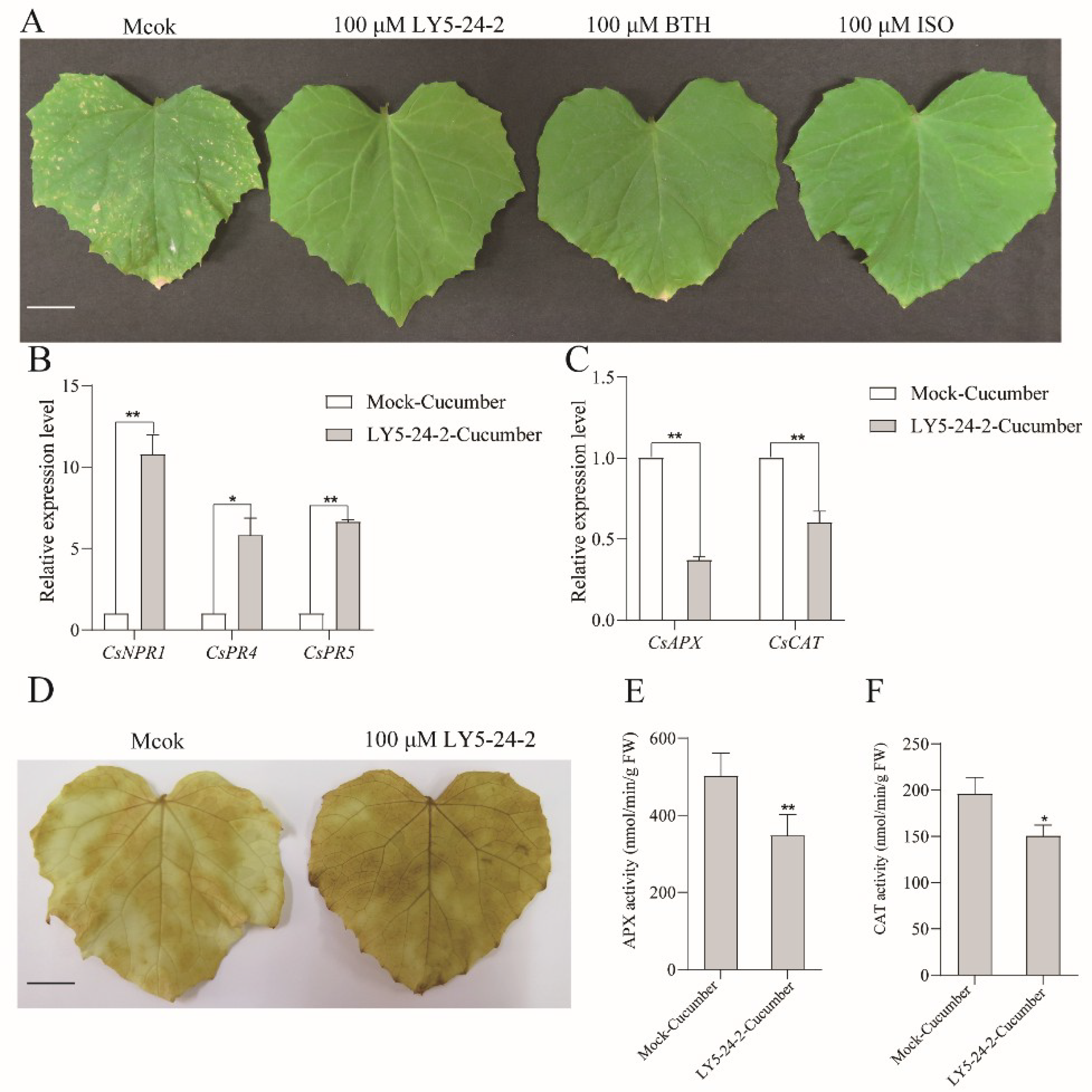
2.6. Defense Enhancement of Cucumber against P. cubensis by LY5-24-2
3. Discussion
4. Materials and Methods
4.1. Synthesis of LY5-24-2
4.2. Plant Materials, Growth Conditions, Treatments with Compound Solutions and Pathogen Inoculation
4.3. Lactophenol Trypan Blue Staining
4.4. Cell Wall Component Measurements
4.5. Stomatal Morphology of A. thaliana Leaves
4.6. Analysis of Ca2+ Content
4.7. Transcriptome Analysis
4.8. RNA Extraction and Quantitative Real-Time PCR
4.9. Measurement of AXP and CAT Activity
4.10. Measurement of ROS Accumulation
4.11. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhou, J.M.; Zhang, Y. Plant immunity: Danger perception and signaling. Cell 2020, 181, 978–989. [Google Scholar] [CrossRef] [PubMed]
- Kourelis, J.; Van, D. Defended to the nines: 25 years of resistance gene cloning identifies nine mechanisms for R protein function. Plant Cell 2018, 30, 285–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Weyer, A.L.; Monteiro, F.; Furzer, O.J.; Nishimura, M.T.; Cevik, V.; Witek, K.; Jones, J.D.G.; Dangl, J.L.; Weigel, D.; Bemm, F. A species wide inventory of NLR genes and alleles in Arabidopsis thaliana. Cell 2019, 178, 1260–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Z.Q.; Dong, X. Systemic acquired resistance: Turning local infection into global defense. Annu. Rev. Plant Biol. 2013, 64, 839–886. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Sun, T.; Ao, K.; Peng, Y.; Zhang, Y.; Li, X.; Zhang, Y. Opposite roles of salicylic acid receptors NPR1 and NPR3/NPR4 in transcriptional regulation of Plant immunity. Cell 2018, 173, 1454–1467. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, D.; Chu, J.Y.; Boyle, P.; Wang, Y.; Brindle, I.D.; De Luca, V.; Després, C. The Arabidopsis NPR1 protein is a receptor for the plant defense hormone salicylic acid. Cell Rep. 2012, 1, 639–647. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, X. Salicylic acid: Biosynthesis, perception, and contributions to plant immunity. Curr. Opin. Plant Biol. 2019, 50, 29–36. [Google Scholar] [CrossRef]
- Damalas, C.A.; Eleftherohorinos, I.G. Pesticide exposure, safety issues, and risk assessment indicators. Int. J. Environ. Res. Public Health 2010, 8, 1402–14197. [Google Scholar] [CrossRef]
- Bektas, Y.; Eulgem, T. Synthetic plant defense elicitors. Front. Plant Sci. 2014, 5, 804. [Google Scholar] [CrossRef]
- Kassanis, B.; White, R.F. Polyacrylic acid -induced resistance to tobacco mosaic virus in Tobacco cv. Xanthi. Ann. Appl. Biol. 1975, 79, 215–220. [Google Scholar] [CrossRef]
- Langcake, P.; Wickins, S.G.A. Studies on the action of the dichlorocyclopropanes on the host-parasite relationship in the rice blast disease. Physiol. Plant Pathol. 1975, 7, 113–126. [Google Scholar] [CrossRef]
- Oostendorp, M.; Kunz, W.; Dietrich, B.; Staub, T. Induced disease resistance in plants by chemicals. Eur. J. Plant Pathol. 2001, 107, 19–28. [Google Scholar] [CrossRef]
- White, R.F. Acetylsalicylic acid (aspirin) induces resistance to tobacco mosaic virus in Tobacco. Virology 1979, 99, 410–412. [Google Scholar] [CrossRef]
- Kunz, W.; Staub, T.; Metraux, J.P.; Hoegerle, K.; Nyfeler, R.; Ahl, P. A Method for Protecting Plants against Diseases. European Patent 0268775A1, 6 January 1988. [Google Scholar]
- Tsubata, K.; Kuroda, K.; Yamamoto, Y.; Yasokawa, N. Development of a novel plant activator for rice diseases, tiadinil. J. Pestic. Sci. 2006, 31, 161–162. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Ogino, A.; Yamada, K.; Sonoda, R. Induction of disease resistance in tea (Camellia sinensis L.) by plant activators. JARQ 2010, 44, 391–398. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, M. Regulation mechanisms of systemic acquired resistance induced by plant activators. J. Pestic. Sci. 2007, 32, 281–282. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Itou, A.; Yamamoto, R.; Tobino, T.; Murakawa, H.; Kei Toda, K. Determination of isotianil in brown rice and soil using supercritical fluid extraction and gas chromatography/mass spectrometry. Anal. Sci. 2013, 29, 919–922. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, H.; Konishi, K.; Nakagawa, T.; Sekido, S.; Yamaguchi, I. Characteristics of n-phenylsulfonyl-2-chloroisonicotinamide as an anti-rice blast agent. J. Pestic. Sci. 1990, 15, 199–203. [Google Scholar] [CrossRef] [Green Version]
- Nishioka, M.; Nakashita, H.; Suzuki, H.; Akiyama, S.; Yoshida, S.; Yamaguchi, I. Induction of resistance against rice blast disease by a novel class of plant activators, pyrazolecarboxylic acid derivatives. J. Pestic. Sci. 2003, 28, 416–421. [Google Scholar] [CrossRef] [Green Version]
- Nishioka, M. Studies on regulation of plant physiology by pesticides. J. Pestic. Sci. 2021, 46, 393–398. [Google Scholar] [CrossRef]
- Fan, Z.J.; Shi, Z.G.; Zhang, H.K.; Liu, X.F.; Bao, L.L.; Ma, L.; Zuo, X.; Zheng, Q.; Mi, N. Synthesis and biological activity evaluation of 1,2,3-thiadiazole derivatives as potential elicitors with highly systemic acquired resistance. J. Agric. Food Chem. 2009, 57, 4279–4286. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Guo, X.F.; Fan, Z.J.; Zhang, N.L.; Zhu, Y.J.; Zhang, Z.M.; Inna Khazhieva, I.; Yurievich, M.Y.; Belskaya, N.P.; Bakulev, V.A. Synthesis and fungicidal activity of 3,4-dichloroisothiazole based strobilurins as potent fungicide candidates. RSC Adv. 2017, 7, 3145–3151. [Google Scholar] [CrossRef] [Green Version]
- Qiu, D.W.; Zeng, H.M. Research and development of small chemical molecular plant immunity inducers. In Technology of Plant Immunity Induction; Science Press: Beijing, China, 2021; pp. 177–197. ISBN 978-7-03-068345-8. [Google Scholar]
- Qi, X.; Chen, L.; Zhang, Y.; Gao, W.; Chen, L.; Wang, D.; Tang, L.; Wang, Z.; Wang, N.; Fan, Z. The methoxyacrylate fungicide candidate CL-15C also functions as a plant elicitor in Arabidopsis thaliana and Oryza sativa L. J. Agric. Food Chem. 2022, 70, 3142–3150. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, X.; Fan, Z.; Liang, X.; Li, Y.; Mao, W.; Li, J.; Wang, D.; Wang, S.; Zhou, L.; et al. A Kind of 3,4-Dichloroisothiazole Derivatives and Its Preparation Methods and Application. CN102942565A, 27 February 2013. [Google Scholar]
- Herlihy, J.; Ludwig, N.R.; Gvd, A.; McDowell, J.M. Oomycetes used in Arabidopsis research. Arab. Book 2019, 17, e0188. [Google Scholar]
- Qiu, D.; Dong, Y.; Zhang, Y.; Li, S.; Shi, F. Plant immunity inducer development and application. Mol. Plant Microbe Interact. 2017, 30, 355. [Google Scholar] [CrossRef] [Green Version]
- Wendehenne, D.; Durner, J.; Chen, Z.; Klessig, D.F. Benzothiadiazole, an inducer of plant defenses, inhibits catalase and ascorbate peroxidase. Phytochemistry 1998, 47, 651–657. [Google Scholar] [CrossRef]
- Knoth, C.; Salus, M.S.; Girke, T.; Eulgem, T. The synthetic elicitor 3,5-dichloroanthranilic acid induces NPR1-dependent and NPR1-independent mechanisms of disease resistance in Arabidopsis. Plant Physiol. 2009, 150, 333–347. [Google Scholar] [CrossRef] [Green Version]
- Ogava, M.; Kadowaki, A.; Yamada, T.; Kadooka, O. Applied Development of a Novel Fungicide Isotianil (Stout); R&D Report, No. I; Health &; Crop Sciences Research Laboratory: Takarazuka, Japan; Sumitomo Chemical Co., Ltd: Tokyo, Japan, 2011; pp. 1–16. [Google Scholar]
- Feys, B.J.; Wiermer, M.; Bhat, R.A.; Moisan, L.J.; Medina-Escobar, N.; Neu, C.; Cabral, A.; Parker, J.E. Arabidopsis SENESCENCE-ASSOCIATED GENE101 stabilizes and signals within an ENHANCED DISEASE SUSCEPTIBILITY1 complex in plant innate immunity. Plant Cell 2005, 17, 2601–2613. [Google Scholar] [CrossRef] [Green Version]
- Shi, Q.; Ji, Y.; Shi, Y.; Zhao, Z.; Zhu, W.; Xu, Y.; Li, B.; Qian, X. Floro-pyrazolo[3,4-d]pyrimidine derivative as a novel plant activator induces two-pathway immune system. Phytochemistry 2021, 184, 112657. [Google Scholar] [CrossRef]
- Zheng, X.; Xing, J.; Zhang, K.; Pang, X.; Zhao, Y.; Wang, G.; Zang, J.; Huang, R.; Dong, J. Ethylene response factor ERF11 activates BT4 transcription to regulate immunity to Pseudomonas syringae. Plant Physiol. 2019, 180, 1132–1151. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Salus, M.; Bektas, Y.; Schroeder, M.; Knoth, C.; Eulgem, T. The synthetic elicitor 2-(5-bromo-2-hydroxy-phenyl)-thiazolidine-4-carboxylic acid links plant immunity to hormesis. Plant Physiol. 2016, 170, 444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDowell, J.M.; Hoff, T.; Anderson, R.G.; Deegan, D. Propagation, storage, and assays with Hyaloperonospora arabidopsidis: A model oomycete pathogen of Arabidopsis. Methods Mol. Biol. 2011, 712, 137–151. [Google Scholar] [CrossRef]
- Yoo, S.D.; Cho, H.Y.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, X.; Li, K.; Chen, L.; Zhang, Y.; Zhang, N.; Gao, W.; Li, Y.; Liu, X.; Fan, Z. Plant Defense Responses to a Novel Plant Elicitor Candidate LY5-24-2. Int. J. Mol. Sci. 2022, 23, 5348. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105348
Qi X, Li K, Chen L, Zhang Y, Zhang N, Gao W, Li Y, Liu X, Fan Z. Plant Defense Responses to a Novel Plant Elicitor Candidate LY5-24-2. International Journal of Molecular Sciences. 2022; 23(10):5348. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105348
Chicago/Turabian StyleQi, Xin, Kun Li, Lei Chen, Yue Zhang, Nailou Zhang, Wei Gao, Yuedong Li, Xingzhong Liu, and Zhijin Fan. 2022. "Plant Defense Responses to a Novel Plant Elicitor Candidate LY5-24-2" International Journal of Molecular Sciences 23, no. 10: 5348. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105348