
TRUE Gene Silencing


Research Institute for Healthy Living, Niigata University of Pharmacy and Applied Life Sciences, Niigata 956-8603, Japan
Int. J. Mol. Sci. 2022, 23(10), 5387; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105387
Submission received: 11 April 2022
/
Revised: 9 May 2022
/
Accepted: 10 May 2022
/
Published: 11 May 2022
(This article belongs to the Collection Regulation by Non-coding RNAs)
Abstract
:TRUE gene silencing is an RNA-mediated gene expression control technology and is termed after tRNase ZL-utilizing efficacious gene silencing. In this review, I overview the potentiality of small guide RNA (sgRNA) for TRUE gene silencing as novel therapeutics. First, I describe the physiology of tRNase ZL and cellular small RNA, and then sgRNA and TRUE gene silencing. An endoribonuclease, tRNase ZL, which can efficiently remove a 3′ trailer from pre-tRNA, is thought to play the role in tRNA maturation in the nucleus and mitochondria. There exist various small RNAs including miRNA and fragments from tRNA and rRNA, which can function as sgRNA, in living cells, and human cells appear to be harnessing cytosolic tRNase ZL for gene regulation together with these small RNAs. By utilizing the property of tRNase ZL to recognize and cleave micro-pre-tRNA, a pre-tRNA-like or micro-pre-tRNA-like complex, as well as pre-tRNA, tRNase ZL can be made to cleave any target RNA at any desired site under the direction of an artificial sgRNA that binds a target RNA and forms the pre-tRNA-like or micro-pre-tRNA-like complex. This general RNA cleavage method underlies TRUE gene silencing. Various examples of the application of TRUE gene silencing are reviewed including the application to several human cancer cells in order to induce apoptosis. Lastly, I discuss the potentiality of sgRNA as novel therapeutics for multiple myeloma.
1. Introduction
After a long-term struggle for their development, six antisense oligonucleotide therapeutics and two siRNA therapeutics for several diseases have been approved by US Food and Drug Administration [1,2,3], and this encourages us to further develop novel oligonucleotide therapeutics for various diseases. In this review, I overview the potentiality of small guide RNA (sgRNA) for RNA-mediated gene expression control technology, TRUE gene silencing (after tRNase ZL-utilizing efficacious gene silencing), as novel therapeutics. sgRNA guides target mRNA cleavage by tRNase ZL as antisense oligonucleotide and siRNA guide target mRNA cleavage by RNase H1 and Argonaute 2, respectively.
Since TRUE gene silencing has been studied by only us and a few collaborators of us, this review focuses primarily on our studies. First, I describe the physiology of tRNase ZL and cellular small RNA in Section 2, and then sgRNA and TRUE gene silencing in Section 3. In Section 3, I review four types of sgRNA, how to choose target sites for sgRNA, and examples of application of TRUE gene silencing, and discuss the potentiality of sgRNA as novel therapeutics for multiple myeloma in Section 3.3.
Although the development of sgRNA is still in its infancy, I expect that this review will stimulate readers’ interest in TRUE gene silencing to bring breakthroughs in sgRNA therapeutics for various diseases. Lastly, I would like to note that “sgRNA” in this review, which has been used since 2000, is completely different from sgRNA for the CRISPR/Cas9 system, to avoid confusion.
2. Gene Regulation by tRNase ZL and Cellular Small RNA
2.1. Human tRNase Z
Human tRNase Z (tRNA 3′ processing endoribonuclease, 3′ tRNase, or RNase Z) (EC 3.1.26.11) is encoded on chromosomes 17 and 18. The long form (tRNase ZL) that consists of 826 amino acids is on the former and the short form (tRNase ZS) that consists of 363 amino acids is on the latter [4]. tRNase ZL, which can efficiently remove a 3′ trailer from pre-tRNA, is thought to play a role in tRNA maturation in the nucleus and mitochondria [4], while tRNase ZS, which can efficiently remove a phosphate from the CCA-less tRNA ending with the 2′,3′-cyclic phosphate, is thought to play a role in tRNA recycling in cytosol [5].
2.2. Targeted RNA Cleavage by tRNase ZL under the Direction of sgRNA
tRNase ZL can recognize and cleave micro-pre-tRNA, a pre-tRNA-like or micro-pre-tRNA-like complex, as well as pre-tRNA [6]. By utilizing this property, tRNase ZL can be made to cleave any target RNA at any desired site under the direction of an artificial sgRNA that binds a target RNA and forms the pre-tRNA-like or micro-pre-tRNA-like complex (Figure 1). There exist various small RNAs including miRNA and fragments from tRNA and rRNA, which can function as sgRNA, in living cells, and human cells appear to be harnessing cytosolic tRNase ZL for gene regulation together with these small RNAs [7,8].
It has been shown that a 5′-half-tRNAGlu and a 28S rRNA fragment in HEK293 cells, which co-immunoprecipitate with tRNase ZL, can function as sgRNA for tRNase ZL and that these RNA fragments downregulate the PPM1F and DYNC1H1 mRNAs, respectively, in HEK293 cells [7]. Furthermore, the transcriptome analysis for HEK293 cells that overexpress tRNase ZL has suggested that tRNase ZL is likely to be involved in the p53 signaling pathway and apoptosis.
Various 3′-truncated tRNAs of ~67 nt have been found to exist as relatively stable complexes with tRNase ZL in mouse FM3A cell extracts [9,10,11,12,13]. These RNA/enzyme complexes (generally termed RNase 65) have been shown to function as four-base-recognizing RNA cutters, in which the 4-nt 5′-terminal sequences of the 3′-truncated tRNAs are used to recognize target RNAs via 4 base-pairings (Figure 1). Although little is known about the physiological role and substrate of RNase 65, it has been shown that the level of a 3′-truncated tRNAArg increases in human BJAB cells after adding 5-fluorouracil [7].
It has been reported that various 5′-half-tRNAs and 3′-truncated tRNAGly and tRNAVal exist in mouse epididymis and that the 5′-half-tRNAGly suppresses genes associated with the endogenous retroelement MERVL [14]. Although these 5′-half-tRNAs and 3′-truncated tRNAs should work as sgRNAs for tRNase ZL, it remains to be seen what their cellular target RNAs are and whether they can guide the target RNA cleavages by tRNase ZL in the cells.
Furthermore, it has been found that various potential sgRNAs exist in human plasma [15]. Among them, a 31/32-nt RNA fragment derived from 94-nt Y4-RNA is particularly noteworthy [16]. Although its genuine target RNAs are not known, the observations that it is overwhelmingly abundant in plasma and that it can function as sgRNA for tRNase ZL have suggested that the 31/32-nt Y4-RNA fragment may work as an intercellular signaling molecule [17].
In addition, the observation that the white blood cell count and the C-reactive protein level show moderate positive correlations with the Y4-RNA fragment level in plasma has suggested that it could be used as a novel inflammatory marker [18]. It would be also notable that the Y4-RNA fragment levels of all four multiple myeloma patients examined were beyond the normal level even though their white blood cell counts were normal.
3. TRUE Gene Silencing
3.1. Four Types of sgRNA
In parallel with the studies on physiological aspects of tRNase ZL and intracellular and extracellular sgRNAs mentioned above, the types of sgRNA and the interactions between tRNase ZL and its RNA substrates have been intensively investigated and this has led to the development of the general RNA cleavage method [6,19,20,21,22,23,24,25]. With this method, any target RNA can be cleaved at any desired site by tRNase ZL with the aid of appropriate sgRNA in a test tube (Figure 1). Incidentally, a similar method has been developed using RNase P and external guide RNA [25].
tRNase ZL primarily recognizes the L-shaped folding structure formed from the cloverleaf-like secondary structure of pre-tRNA and cleaves it after the discriminator nucleotide to remove its 3′ trailer [4]. In the same fashion, a pre-tRNA-like RNA complex formed between a target RNA and a 5′-half-tRNA or 3′-truncated tRNA can be recognized by tRNase ZL and the target RNA can be cleaved [13,19].
tRNase ZL can also recognize and cleave micro-pre-tRNA that lacks the D, anticodon, and extra arms after a nucleotide corresponding to the discriminator as efficiently as pre-tRNA [24]. In similar ways, tRNase ZL can recognize a micro-pre-tRNA-like RNA complex formed between a target RNA and a hook-shaped RNA, ~14-nt linear RNA, or heptamer RNA and cleave the target RNA [6,20,22,24].
On the basis of the above property of tRNase ZL, we can cleave a target RNA with various specificities in sequence and length by selecting appropriate guide RNAs. tRNase ZL can work as an ~4-nt sequence specific RNA cutter with 3′-truncated tRNAs or hook-shaped RNAs, and as 12-nt and ~14-nt sequence specific RNA cutters with 5′-half-tRNAs and ~14-nt linear RNAs, respectively. Since an ~5-base-pair T-arm-like structure is needed for recognition of a target RNA by tRNase ZL when using heptamer RNA, roughly speaking, the target RNA can be cleaved with an ~12-nt (not merely 7-nt) sequence specificity [20]. The last four guide RNAs are termed small guide RNA, or sgRNA for tRNase ZL (Figure 1).
With respect to heptamer-type sgRNA, how chemical modifications on it affect its guiding efficiency has been investigated. The guiding efficiencies (Vmax/Km/Kd) of the sgRNA with full 2′-O-methyl modifications, the sgRNA with full phosphorothioate modifications, sgDNA, and sgDNA with full phosphorothioate modifications have been shown to be 89, 53, 21, and 6%, respectively, of that of the natural sgRNA [26]. The efficiencies of heptamer-guided target RNA cleavage by tRNase ZL have been shown to decrease as the number of LNA-modified riboses increases [27].
Furthermore, from the investigation on the interaction between tRNase ZL and pre-tRNA, several pieces of information important for selecting sgRNA have been obtained. On the whole, the efficiency of pre-tRNA cleavage by tRNase ZL decreases as its 3′ trailer becomes longer [28]. The cleavage efficiency for pre-tRNA with its 3′ trailer starting with G or A is the highest, while that for pre-tRNA with its 3′ trailer starting with C is the lowest [28]. The cleavage of pre-tRNA with its 5′ leader of more than 8 nt is severely inhibited, and that of pre-tRNA with its 5′ leader that forms more than 2 base-pairs with its 3′ trailer resulting in an extended acceptor stem is also suppressed [29].
Although tRNase ZL usually cleaves pre-tRNA after the discriminator nucleotide, in some cases, it cleaves a pre-tRNA-like or micro-pre-tRNA-like complex at multiple sites near a nucleotide corresponding to the discriminator [19,20,22]. This would be due to the presence of multiple conformers of the RNA complex and/or the presence of multiple modes of the RNA complex/tRNase ZL interaction. The experiments for pre-tRNA variants have shown that, in general, pre-tRNA variants containing a total of >11 base pairs in the acceptor stem and the T stem are cleaved only after the discriminator and that pre-tRNA variants with a total of N base pairs (N is <12) are cleaved 12 minus N and 13 minus N nt downstream of the discriminator [30].
As explained in detail above, tRNase ZL can cleave target RNA with varying sequence/length specificities under the direction of appropriate sgRNA. Target RNA can be as small as miRNA and as long as mRNA, and any site can be targeted including a 3′-terminal nucleotide with the exception of tightly folded regions and 5′-terminal 4 sites.
Target sites for each type of sgRNA can be chosen as follows. With respect to 5′-half-tRNA-type and heptamer-type sgRNA, first, hairpin structures resembling the T-arm of tRNA in a target RNA are selected with the aid of an appropriate computer program and/or visually. Then, candidate target sites are chosen from among them by excluding the sites in potentially tightly folded RNA regions, which can be predicted by a computer program for RNA secondary structure prediction. With respect to hook-type and 14-nt linear-type sgRNA, the first step is not needed. Although RNA primary structures of target sites seem to hardly affect RNA cleavage efficiency by tRNase ZL, some sequence constraints could be found by further analyses.
3.2. Modulation of Gene Expression by TRUE Gene Silencing
The efficacy of the general RNA cleavage method using tRNase ZL and sgRNA in living cells has been examined by measuring expression levels of reporter genes and endogenous genes in cultured cells or in mice. As below, its efficacy has been shown with various degrees, and this technology is called TRUE gene silencing after tRNase ZL-utilizing efficacious gene silencing (Figure 2). Some of the examples are the application of TRUE gene silencing to several human cancer cells in order to induce apoptosis in them, and are listed in Table 1.
It has been demonstrated that the expression of chloramphenicol acetyltransferase can be downregulated by a 5′-half-tRNA-type sgRNA targeting its mRNA which is introduced into Madin–Darby canine kidney epithelial cells as an expression plasmid or a synthetic 2′-O-methyl RNA [26]. It has also been shown that 2′-O-methyl heptamer-type sgRNAs targeting luciferase mRNA variants can downregulate their mRNA levels in HEK293 cells and that a 2′-O-methyl heptamer-type sgRNA targeting the BCL2 mRNA reduces viability of Sarcoma 180 mouse cells [26].
It has been shown that HIV-1 gene expression is suppressed in COS cells by transiently expressing 5′-half-tRNA-type sgRNAs targeting the gag mRNA and that the expression of HIV-1 gag p24 is almost completely suppressed in Jurkat cells stably expressing a 5′-half-tRNA-type sgRNA targeting the gag mRNA for at least 18 days after challenge with HIV-1 [38].
The involvement of tRNase ZL in downregulation of cellular mRNAs has been shown by the observation that the efficacy of luciferase downregulation by a 5′-half-tRNA-type or 14-nt linear-type 2′-O-methyl sgRNA targeting the luciferase mRNA increases with an increasing cellular tRNase ZL level in HEK293 cells [39]. It has been observed that BCL2 and GSK3B expression can be downregulated in HEK293 cells that stably express a 5′-half-tRNA-type or 14-nt linear-type sgRNA targeting the BCL2 and GSK3B mRNAs, respectively [39]. Furthermore, the expression of chloramphenicol acetyltransferase and luciferase has been shown to be suppressed in the livers of postnatal mice that were tail-vein-injected with a 5′-half-tRNA-type 2′-O-methyl sgRNA targeting the chloramphenicol acetyltransferase mRNA and a heptamer-type 2′-O-methyl sgRNA targeting the luciferase mRNA, respectively [39].
To discover anti-angiogenic sgRNAs, TRUE gene silencing has been applied to the VEGF gene by using four 5′-half-tRNA-type sgRNAs and four 14-nt linear-type sgRNAs targeting the human VEGF mRNA [40]. It has been found that many of them downregulate the exogenous VEGF mRNA efficiently in HEK293 and HeLa cells and that two of them downregulate the endogenous VEGF gene expression in HeLa cells much more efficiently than an siRNA.
It has been shown that five naked heptamer-type sgRNAs with 2–7 LNA modifications targeting the BCL2 mRNA reduce its mRNA levels in Jurkat cells as efficiently as a heptamer-type sgRNA with no LNA modification [27].
It has been demonstrated that a naked 14-nt linear-type sgRNA targeting human miR-16 significantly reduces the miR-16 level in HEK293 and HL60 cells [41]. Three other naked 14-nt linear-type sgRNAs targeting miR-142-3p, miR-206, and miR-19a/b have also been shown to downregulate the respective miRNA levels in various mammalian cell lines [41]. It appears that, in general, a specific cellular miRNA can be eliminated at least by ~50% by using a naked linear-type sgRNA.
A naked heptamer-type sgRNA targeting the human BCL2 mRNA has been shown to moderately downregulate its mRNA level in the human HL60 leukemia cells and to induce apoptosis in them (Table 1) [32]. In xenograft experiments using nude mice implanted with the HL60 cells, it has been shown that the median survival of the mice treated with this sgRNA is longer than that of the control mice [32].
It has been demonstrated that naked heptamer-type sgRNAs targeting the WT1 mRNA can reduce its mRNA levels and WT1 protein amounts in the WT1-expressing leukemia cells and that these sgRNAs efficiently induce apoptosis in these cells but not in WT1-nonexpressing cells (Table 1) [33].
To search for potential therapeutic sgRNAs for leukemia, a library composed of 156 heptamer-type sgRNAs has been screened, and 39 sgRNAs have been found to be able to efficiently induce apoptosis in human HL60 leukemia cells (Table 1) [34,35,42]. Furthermore, it has been demonstrated that 4 of the 39 sgRNAs can reduce growth rates of HL60 cells in mouse xenograft model experiments [34].
To find potential sgRNA therapeutics for head and neck squamous cell carcinoma, six heptamer-type, two linear-type, and two 5′-half-tRNA-type sgRNAs targeting the human CCND1 mRNA have been examined (Table 1) [31]. It has been demonstrated that all of these sgRNAs downregulate the mRNA level in HSC-2 or HSC-3 cells and that a subset of the sgRNAs reduce its protein level and induce apoptosis in HSC-3 cells [31].
In the course of an experiment to investigate the ability of a heptamer-type sgRNA targeting the OCT4 mRNA to differentiate human amnion stem cells, it has been observed unexpectedly that the amnion cells exhibit a morphology resembling initialized cells [43]. Further examination of its effect on human HL60 leukemia cells has shown that this OCT4-mRNA-targeting sgRNA can upregulate the OCT4 expression and the expression of alkaline phosphatase in the cells [43]. Although nothing is known about the molecular mechanism of the OCT4 upregulation, the sgRNA might happen to suppress genes that suppress the OCT4 expression.
3.3. Potential sgRNA Therapeutics for Multiple Myeloma
To explore the possibility of sgRNA as therapeutic agents, multiple myeloma was chosen as a target disease, and an intensive investigation was conducted. Multiple myeloma is a refractory plasma cell cancer [44]. In spite of various treatment modalities [45,46], achieving complete remission appears to be very hard because drug-resistant mutations emerge almost inevitably in the course of myeloma cell evolution resulting in relapse of myeloma in most patients.
To search for potential sgRNA therapeutics for multiple myeloma, the library of 156 heptamer-type sgRNAs has been screened, and 7 and 5 sgRNAs have been shown to be able to efficiently induce apoptosis in human KMM-1 and RPMI-8226 myeloma cells, respectively (Table 1) [34,35,42].
It has been demonstrated that two consecutively aligned heptamer-type sgRNAs targeting the human BCL2 mRNA can guide target RNA cleavage by tRNase ZL as efficiently as a corresponding 14-nt linear-type sgRNA and that the double heptamer-type sgRNA, but not the 14-nt linear-type sgRNA, downregulates the BCL2 mRNA in HL60 cells [27]. It has been shown that the double heptamer-type sgRNA and each single heptamer-type sgRNA reduce viability of RPMI-8226 and KMM-1 cells and also suppress the growth of KMM-1 cells in a mouse xenograft experiment (Table 1) [36]. The absence of stable downregulation of the BCL2 mRNA has suggested that the double heptamer-type sgRNA targets other cellular RNAs to reduce the myeloma cell viability. Furthermore, two sets of double heptamer-type sgRNA targeting the human CCND1 mRNA have been shown to synergistically reduce RPMI-8226 cell viability [36].
The interplay between the bone marrow microenvironment and myeloma cells is essentially important for the pathology. To search for heptamer-type sgRNAs that can shift tumor-associated macrophages toward the M1 state, a heptamer-type sgRNA library has been screened for the ability to upregulate IL-12b gene expression in human macrophage-like cell lines, and three such sgRNAs have been found [37]. It has been demonstrated that one of the sgRNAs also shows such ability in fresh human macrophages and mouse macrophage-like cell lines and efficiently suppresses human KMM-1 myeloma cell growth in immunocompromised mice that is deficient in acquired immunity (Table 1) [37]. The observation that this sgRNA cannot induce apoptosis in KMM-1 myeloma cells suggests that it affects mouse macrophages in the tumor microenvironment and induces them to eliminate the implanted myeloma cells.
With respect to the above two sgRNAs that worked efficiently in mouse experiments [36,37], in the next stage, cellular target RNAs of these two sgRNAs need to be identified by transcriptome analyses to elucidate molecular mechanisms for how these sgRNAs work. In parallel, the efficacy of these two sgRNAs in mouse xenograft experiments needs to be examined by their systemic administration rather than local administration.
Although the development of sgRNA therapeutics for multiple myeloma is still in its infancy, there are several advantages worth mentioning: sgRNA can be taken up by cells without any transfection reagent [32,33]; since tRNase ZL exists ubiquitously in a human cell, in theory, any cellular RNA in cytosol, nucleus, and mitochondria can be targeted [7]; since discontinuous capillaries are formed in bone marrow, myeloma is one of the best targets of sgRNA therapeutics from the viewpoint of drug delivery. Concerning the first advantage, the cellular uptake of naked sgRNA is thought to occur by endocytosis, although the molecular mechanism is not fully understood [47].
Since the above two sgRNAs were obtained from screenings of libraries composed of only less than 160 heptamer-type sgRNAs, much better heptamer-type sgRNAs would be expected to be discovered from a screening of the full library composed of 16,384 heptamer-type sgRNAs. In parallel, by examining 5′-half-tRNA-type and 14-nt linear-type sgRNAs designed to target appropriate mRNAs such as the BCL2, CCND1, and IRF4 mRNAs for their ability to induce apoptosis in myeloma cells, potential sgRNA therapeutics would be expected to be found. Further nucleotide modifications of sgRNAs in addition to the 2′-O-methyl modification would also improve their efficacy in inducing apoptosis in myeloma cells. Lastly, since heptamer-type, 5′-half-tRNA-type, and 14-nt linear-type sgRNAs consist of 16,384, 16,777,216, and 268,435,456 different sequence sgRNAs, respectively, sgRNA therapeutics may help promote personalized medicine by selecting the most appropriate sgRNA depending on a patient’s own mRNA sequence and changing patient conditions.
Funding
This work was supported in part by the Science Research Promotion Fund from the Promotion and Mutual Aid Corporation for Private Schools of Japan.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
I would like to thank the present lab members Masayuki Takahashi and Mineaki Seki, former lab members and collaborators for their cooperation in the development of TRUE gene silencing and give special thanks to Masato Tamura at Hokkaido University for his support and cooperation for more than a quarter century.
Conflicts of Interest
The author is an advisor of Veritas In Silico Inc., and owns stock of the company.
References
- Herkt, M.; Thum, T. Pharmacokinetics and Proceedings in Clinical Application of Nucleic Acid Therapeutics. Mol. Ther. 2021, 29, 521–539. [Google Scholar] [CrossRef]
- Scharner, J.; Aznarez, I. Clinical Applications of Single-Stranded Oligonucleotides: Current Landscape of Approved and In-Development Therapeutics. Mol. Ther. 2021, 29, 540–554. [Google Scholar] [CrossRef] [PubMed]
- Rossi, J.J.; Rossi, D.J. siRNA Drugs: Here to Stay. Mol. Ther. 2021, 29, 431–432. [Google Scholar] [CrossRef] [PubMed]
- Takaku, H.; Minagawa, A.; Takagi, M.; Nashimoto, M. A candidate prostate cancer susceptibility gene encodes tRNA 3′ processing endoribonuclease. Nucleic Acids Res. 2003, 31, 2272–2278. [Google Scholar] [CrossRef] [PubMed]
- Yip, M.C.J.; Savickas, S.; Gygi, S.P.; Shao, S. ELAC1 Repairs tRNAs Cleaved during Ribosome-Associated Quality Control. Cell Rep. 2020, 30, 2106–2114.e5. [Google Scholar] [CrossRef] [Green Version]
- Shibata, H.S.; Takaku, H.; Takagi, M.; Nashimoto, M. The T loop structure is dispensable for substrate recognition by tRNase ZL. J. Biol. Chem. 2005, 280, 22326–22334. [Google Scholar] [CrossRef] [Green Version]
- Elbarbary, R.A.; Takaku, H.; Uchiumi, N.; Tamiya, H.; Abe, M.; Takahashi, M.; Nishida, H.; Nashimoto, M. Modulation of gene expression by human cytosolic tRNase ZL through 5′-half-tRNA. PLoS ONE 2009, 4, e5908. [Google Scholar] [CrossRef]
- Elbarbary, R.A.; Takaku, H.; Uchiumi, N.; Tamiya, H.; Abe, M.; Nishida, H.; Nashimoto, M. Human cytosolic tRNase ZL can downregulate gene expression through miRNA. FEBS Lett. 2009, 583, 3241–3246. [Google Scholar] [CrossRef] [Green Version]
- Nashimoto, M.; Kominami, R.; Nishi, S.; Mishima, Y. A novel spermidine-dependent endoribonuclease activity caused by RNA-protein complex in mouse FM3A cell extracts. Biochem. Biophys. Res. Commun. 1991, 176, 1163–1169. [Google Scholar] [CrossRef]
- Nashimoto, M.; Sakai, M.; Nishi, S. Transfer RNA lacking its 3′ terminus is required for spermidine-dependent ribonuclease 65 activity in mouse FM3A cell extracts. Biochem. Biophys. Res. Commun. 1991, 178, 1247–1252. [Google Scholar] [CrossRef]
- Nashimoto, M. Characterization of the spermidine-dependent, sequence-specific endoribonuclease that requires transfer RNA for its activity. Nucleic Acids Res. 1992, 20, 3737–3742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nashimoto, M. 3′ truncated tRNAArg is essential for in vitro specific cleavage of partially synthesized mouse 18S rRNA. Nucleic Acids Res. 1993, 21, 4696–4702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nashimoto, M. Conversion of mammalian tRNA 3′ processing endoribonuclease to four-base-recognizing RNA cutters. Nucleic Acids Res. 1995, 23, 3642–3647. [Google Scholar] [CrossRef] [PubMed]
- Sharma, U.; Conine, C.C.; Shea, J.M.; Boskovic, A.; Derr, A.G.; Bing, X.Y.; Belleannee, C.; Kucukural, A.; Serra, R.W.; Sun, F.; et al. Biogenesis and function of tRNA fragments during sperm maturation and fertilization in mammals. Science 2016, 351, 391–396. [Google Scholar] [CrossRef] [Green Version]
- Ninomiya, S.; Kawano, M.; Abe, T.; Ishikawa, T.; Takahashi, M.; Tamura, M.; Takahashi, Y.; Nashimoto, M. Potential small guide RNAs for tRNase ZL from human plasma, peripheral blood mononuclear cells, and cultured cell lines. PLoS ONE 2015, 10, e0118631. [Google Scholar] [CrossRef]
- Ishikawa, T.; Haino, A.; Seki, M.; Terada, H.; Nashimoto, M. The Y4-RNA fragment, a potential diagnostic marker, exists in saliva. Noncoding RNA Res. 2017, 2, 122–128. [Google Scholar] [CrossRef]
- Ninomiya, S.; Ishikawa, T.; Takahashi, M.; Seki, M.; Nashimoto, M. Potential physiological roles of the 31/32-nucleotide Y4-RNA fragment in human plasma. Noncoding RNA Res. 2019, 4, 135–140. [Google Scholar] [CrossRef]
- Ishikawa, T.; Haino, A.; Seki, M.; Kurihara, T.; Hirose, T.; Imai, Y.; Ishiguro, T.; Chou, T.; Toshima, M.; Terada, H.; et al. The 31-nucleotide Y4-RNA fragment in plasma is a potential novel biomarker. Noncoding RNA Res. 2019, 5, 37–40. [Google Scholar] [CrossRef]
- Nashimoto, M. Specific cleavage of target RNAs from HIV-1 with 5′ half tRNA by mammalian tRNA 3′ processing endoribonuclease. RNA 1996, 2, 523–534. [Google Scholar]
- Nashimoto, M.; Geary, S.; Tamura, M.; Kaspar, R. RNA heptamers that direct RNA cleavage by mammalian tRNA 3′ processing endoribonuclease. Nucleic Acids Res. 1998, 26, 2565–2572. [Google Scholar] [CrossRef] [Green Version]
- Nashimoto, M.; Tamura, M.; Kaspar, R.L. Minimum requirements for substrates of mammalian tRNA 3′ processing endoribonuclease. Biochemistry 1999, 38, 12089–12096. [Google Scholar] [CrossRef] [PubMed]
- Nashimoto, M. Anomalous RNA substrates for mammalian tRNA 3′ processing endoribonuclease. FEBS Lett. 2000, 472, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Takaku, H.; Minagawa, A.; Takagi, M.; Nashimoto, M. The N-terminal half-domain of the long form of tRNase Z is required for the RNase 65 activity. Nucleic Acids Res. 2004, 32, 4429–4438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takaku, H.; Minagawa, A.; Takagi, M.; Nashimoto, M. A novel 4-base-recognizing RNA cutter that can remove the single 3′ terminal nucleotides from RNA molecules. Nucleic Acids Res. 2004, 32, e91. [Google Scholar] [CrossRef] [Green Version]
- Scherer, L.; Rossi, J.J. Therapeutic potential of RNA-mediated control of gene expression: Options and designs. In RNA and the Regulation of Gene Expression: A Hidden Layer of Complexity; Morris, K.V., Ed.; Caister Academic Press: Norfolk, UK, 2008; pp. 201–226. [Google Scholar]
- Tamura, M.; Nashimoto, C.; Miyake, N.; Daikuhara, Y.; Ochi, K.; Nashimoto, M. Intracellular mRNA cleavage by 3′ tRNase under the direction of 2′-O-methyl RNA heptamers. Nucleic Acids Res. 2003, 31, 4354–4360. [Google Scholar] [CrossRef] [Green Version]
- Sano, T.; Takahashi, M.; Nozaki, T.; Takahashi, Y.; Tamura, M.; Nashimoto, M. Expanding the utility of heptamer-type sgRNA for TRUE gene silencing. Biochem. Biophys. Res. Commun. 2011, 416, 427–432. [Google Scholar] [CrossRef]
- Nashimoto, M. Distribution of both lengths and 5′ terminal nucleotides of mammalian pre-tRNA 3′ trailers reflects properties of 3′ processing endoribonuclease. Nucleic Acids Res. 1997, 25, 1148–1154. [Google Scholar] [CrossRef] [Green Version]
- Nashimoto, M.; Wesemann, D.R.; Geary, S.; Tamura, M.; Kaspar, R.L. Long 5′ leaders inhibit removal of a 3′ trailer from a precursor tRNA by mammalian tRNA 3′ processing endoribonuclease. Nucleic Acids Res. 1999, 27, 2770–2776. [Google Scholar] [CrossRef] [Green Version]
- Nashimoto, M.; Tamura, M.; Kaspar, R.L. Selection of cleavage site by mammalian tRNA 3′ processing endoribonuclease. J. Mol. Biol. 1999, 287, 727–740. [Google Scholar] [CrossRef]
- Iizuka, S.; Oridate, N.; Nashimoto, M.; Fukuda, S.; Tamura, M. Growth inhibition of head and neck squamous cell carcinoma cells by sgRNA targeting the cyclin D1 mRNA based on TRUE gene silencing. PLoS ONE 2014, 9, e114121. [Google Scholar] [CrossRef]
- Takahashi, M.; Elbarbary, R.A.; Nakashima, A.; Abe, M.; Watanabe, N.; Narita, M.; Takahashi, M.; Tamura, M.; Yoshida, T.; Nashimoto, M. A naked RNA heptamer targeting the human Bcl-2 mRNA induces apoptosis of HL60 leukemia cells. Cancer Lett. 2013, 328, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, N.; Narita, M.; Yamahira, A.; Taniguchi, T.; Furukawa, T.; Yoshida, T.; Miyazawa, T.; Nashimoto, M.; Takahashi, M. Induction of apoptosis of leukemic cells by TRUE gene silencing using small guide RNAs targeting the WT1 mRNA. Leuk. Res. 2013, 37, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Elbarbary, R.A.; Watanabe, N.; Goto, A.; Kamiya, D.; Watabe, Y.; Uchiyama, T.; Narita, M.; Takahashi, M.; Takahashi, Y.; et al. Screening of a heptamer-type sgRNA library for potential therapeutic agents against hematological malignancies. Leuk. Res. 2014, 38, 808–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haino, A.; Ishikawa, T.; Seki, M.; Nashimoto, M. TRUE Gene Silencing: Screening of a Heptamer-type Small Guide RNA Library for Potential Cancer Therapeutic Agents. J. Vis. Exp. 2016, 112, 53879. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, T.; Haino, A.; Ichiyanagi, T.; Seki, M.; Nashimoto, M. Evaluation of double heptamer-type sgRNA as a potential therapeutic agent against multiple myeloma. Blood Cells Mol. Dis. 2019, 79, 102341. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, T.; Haino, A.; Ichiyanagi, T.; Takahashi, M.; Seki, M.; Nashimoto, M. Heptamer-type small guide RNA that can shift macrophages toward the M1 state. Blood Cells Mol. Dis. 2021, 86, 102503. [Google Scholar] [CrossRef] [PubMed]
- Habu, Y.; Miyano-Kurosaki, N.; Kitano, M.; Endo, Y.; Yukita, M.; Ohira, S.; Takaku, H.; Nashimoto, M.; Takaku, H. Inhibition of HIV-1 gene expression by retroviral vector-mediated small-guide RNAs that direct specific RNA cleavage by tRNase ZL. Nucleic Acids Res. 2005, 33, 235–243. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, A.; Takaku, H.; Shibata, H.S.; Negishi, Y.; Takagi, M.; Tamura, M.; Nashimoto, M. Gene silencing by the tRNA maturase tRNase ZL under the direction of small-guide RNA. Gene Ther. 2007, 14, 78–85. [Google Scholar] [CrossRef] [Green Version]
- Elbarbary, R.A.; Takaku, H.; Tamura, M.; Nashimoto, M. Inhibition of vascular endothelial growth factor expression by TRUE gene silencing. Biochem. Biophys. Res. Commun. 2009, 379, 924–927. [Google Scholar] [CrossRef]
- Takahashi, M.; Elbarbary, R.A.; Abe, M.; Sato, M.; Yoshida, T.; Yamada, Y.; Tamura, M.; Nashimoto, M. Elimination of specific miRNAs by naked 14-nt sgRNAs. PLoS ONE 2012, 7, e38496. [Google Scholar] [CrossRef] [Green Version]
- Shivarov, V. TRUE gene silencing for hematologic malignancies. Leuk. Res. 2014, 38, 729. [Google Scholar] [CrossRef] [PubMed]
- Nozaki, T.; Takahashi, M.; Ishikawa, T.; Haino, A.; Seki, M.; Kikuchi, H.; Yuan, B.; Nashimoto, M. The heptamer sgRNA targeting the human OCT4 mRNA can upregulate the OCT4 expression. Biochem. Biophys. Rep. 2021, 26, 100918. [Google Scholar] [CrossRef] [PubMed]
- Key Statistics about Multiple Myeloma. Available online: https://www.cancer.org/cancer/multiple-myeloma/about/key-statistics.html (accessed on 8 May 2022).
- Chim, C.S.; Kumar, S.K.; Orlowski, R.Z.; Cook, G.; Richardson, P.G.; Gertz, M.A.; Giralt, S.; Mateos, M.V.; Leleu, X.; Anderson, K.C. Management of relapsed and refractory multiple myeloma: Novel agents, antibodies, immunotherapies and beyond. Leukemia 2018, 32, 252–262. [Google Scholar] [CrossRef] [PubMed]
- García-Guerrero, E.; Sierro-Martínez, B.; Pérez-Simón, J.A. Overcoming Chimeric Antigen Receptor (CAR) Modified T-Cell Therapy Limitations in Multiple Myeloma. Front. Immunol. 2020, 11, 1128. [Google Scholar] [CrossRef] [PubMed]
- Deprey, K.; Batistatou, N.; Kritzer, J.A. A critical analysis of methods used to investigate the cellular uptake and subcellular localization of RNA therapeutics. Nucleic Acids Res. 2020, 48, 7623–7639. [Google Scholar] [CrossRef]
Figure 1.
Substrates of tRNase ZL. The RNA complexes of the four types of sgRNA and their target RNAs are shown in a blue box. Arrows denote cleavage sites by tRNase ZL.
Figure 1.
Substrates of tRNase ZL. The RNA complexes of the four types of sgRNA and their target RNAs are shown in a blue box. Arrows denote cleavage sites by tRNase ZL.

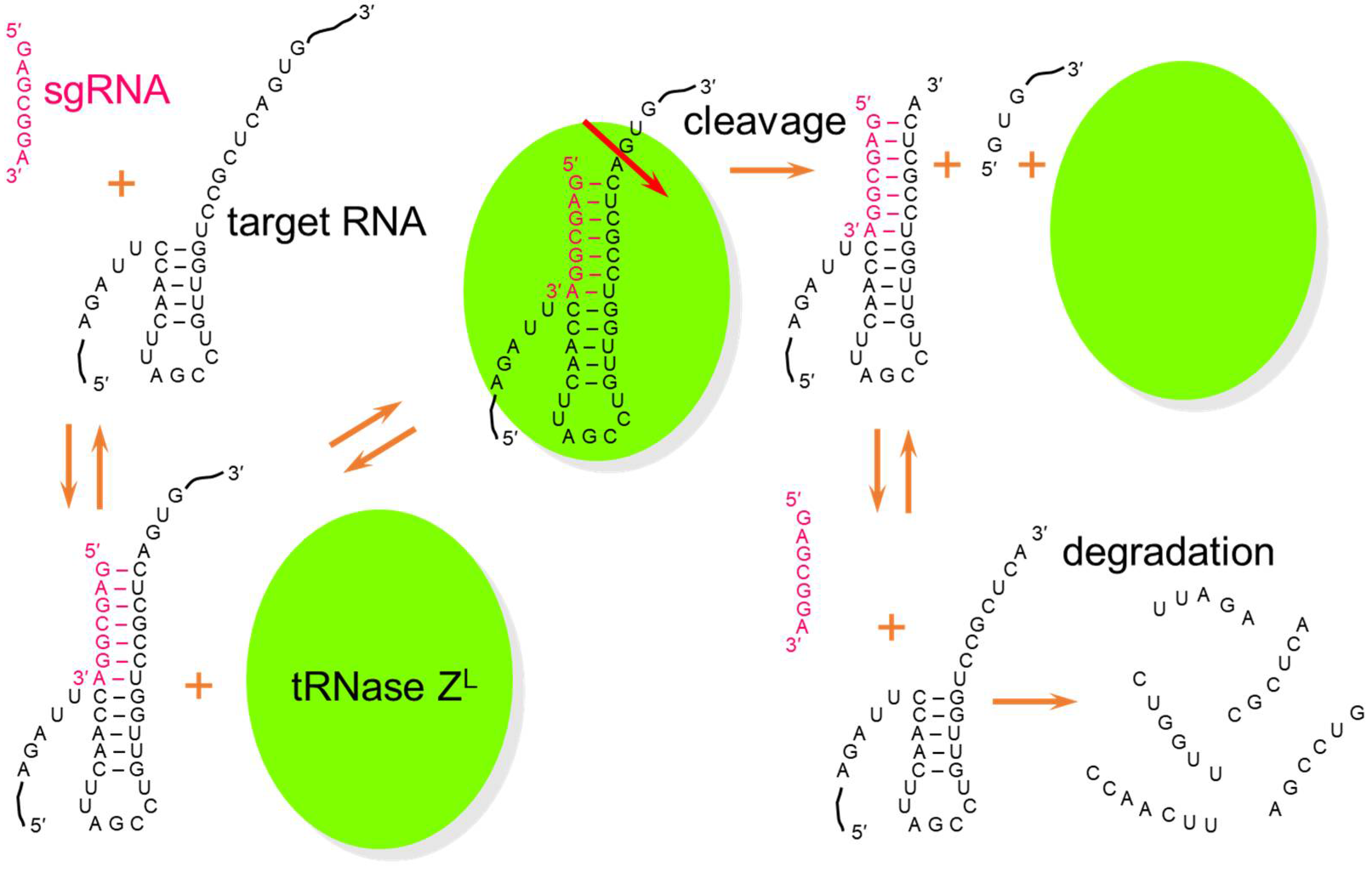
Figure 2.
Molecular mechanism of TRUE gene silencing. A heptamer-type sgRNA binds a target RNA immediately after a 5-base-pair T-arm-like structure to form an sgRNA/target RNA complex. tRNase ZL recognizes this RNA complex and cleaves target RNA after a nucleotide corresponding the discriminator. The cleaved RNAs are further degraded by other nucleases, and the sgRNA and tRNase ZL can be reused to cleave another target RNA.
Figure 2.
Molecular mechanism of TRUE gene silencing. A heptamer-type sgRNA binds a target RNA immediately after a 5-base-pair T-arm-like structure to form an sgRNA/target RNA complex. tRNase ZL recognizes this RNA complex and cleaves target RNA after a nucleotide corresponding the discriminator. The cleaved RNAs are further degraded by other nucleases, and the sgRNA and tRNase ZL can be reused to cleave another target RNA.


{kind=link}
{kind=link}
Table 1.
Application of TRUE gene silencing to human cancer cells in order to induce apoptosis.1.
| Cancer | Target Cell | Target mRNA 2 | sgRNA Type | Experimental Type 3 | Reference |
|---|---|---|---|---|---|
| head and neck squamous cell carcinoma | HSC-2 HSC-3 | CCND1 | Heptamer 14-nt linear 5′-half-tRNA | in vitro | [31] |
| leukemia | HL60 C2F8 | BCL2 WT1 screening | heptamer | in vitro in vivo | [32,33,34,35] |
| multiple myeloma | KMM-1 RPMI-8226 | BCL2 CCND1 screening | heptamer | in vitro in vivo | [34,35,36] |
| THP-1 U937 fresh monocyte J774.1 (mouse) RAW264.7 (mouse) | screening | heptamer | in vitro in vivo | [37] |
1 The last example is the application to macrophage-like cells in order to shift them toward the M1 state. 2 “screening” denotes that target RNAs of the sgRNAs, which were obtained by screening of sgRNA libraries, are unknown. 3 “in vitro” and “in vivo” denote cultured cell experiments and mouse xenograft experiments, respectively.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nashimoto, M. TRUE Gene Silencing. Int. J. Mol. Sci. 2022, 23, 5387. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105387
AMA Style
Nashimoto M. TRUE Gene Silencing. International Journal of Molecular Sciences. 2022; 23(10):5387. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105387
Chicago/Turabian StyleNashimoto, Masayuki. 2022. "TRUE Gene Silencing" International Journal of Molecular Sciences 23, no. 10: 5387. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105387
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.