
Proteomics and Extracellular Vesicles as Novel Biomarker Sources in Peritoneal Dialysis in Children

 and
and
Abstract
:1. Introduction
2. Proteomics in Pediatric Peritoneal Dialysis
3. Proteomics Applied to Extracellular Vesicles and Exosomes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviation
References
- Vasudevan, A.; Phadke, K.; Yap, H.K. Peritoneal dialysis for the management of pediatric patients with acute kidney injury. Pediatr. Nephrol. 2017, 32, 1145–1156. [Google Scholar] [CrossRef] [PubMed]
- Hickman, R.O. Chronic peritoneal dialysis in childhood. Contrib. Nephrol. 1979, 17, 30–32. [Google Scholar] [PubMed]
- Perl, J.; Bargman, J. Peritoneal dialysis: From bench to bedside and bedside to bench. Am. J. Physiol. Ren. Physiol. 2016, 311, F999–F1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanna, R. Solute and Water Transport in Peritoneal Dialysis: A Case-Based Primer. Am. J. Kidney Dis. 2017, 69, 461–472. [Google Scholar] [CrossRef]
- García-López, E.; Lindholm, B.; Davies, S. An update on peritoneal dialysis solutions. Nat. Rev. Nephrol. 2012, 8, 224–233. [Google Scholar] [CrossRef]
- Jung, H.-Y.; Jeon, Y.; Park, Y.; Kim, Y.S.; Kang, S.-W.; Yang, C.W.; Kim, N.-H.; Choi, J.-Y.; Cho, J.-H.; Park, S.-H.; et al. Better Quality of Life of Peritoneal Dialysis compared to Hemodialysis over a Two-year Period after Dialysis Initiation. Sci. Rep. 2019, 9, 10266. [Google Scholar] [CrossRef]
- Sukul, N.; Zhao, J.; Fuller, D.S.; Karaboyas, A.; Bieber, B.; Sloand, J.A.; Subramanian, L.; Johnson, D.W.; Oliver, M.J.; Tungsanga, K.; et al. Patient-reported advantages and disadvantages of peritoneal dialysis: Results from the PDOPPS. BMC Nephrol. 2019, 20, 116. [Google Scholar] [CrossRef] [Green Version]
- Madden, I.; Blaauw, M.; Baugh, N.; Rees, L.; Shroff, R. Chronic peritoneal dialysis in children. Paediatr. Child Health 2020, 30, 319–327. [Google Scholar] [CrossRef]
- McCormick, B.B.; Bargman, J.M. Noninfectious complications of peritoneal dialysis: Implications for patient and technique survival. J. Am. Soc. Nephrol. 2007, 18, 3023–3025. [Google Scholar] [CrossRef] [Green Version]
- Cueto-Manzano, A.M.; González-Espinoza, L.; Del Campo, F.M.; Fortes, P.C.; Pecoits-Filho, R. Inflammation in peritoneal Dialysis: A Latin-American perspective. Perit. Dial. Int. 2007, 27, 347–352. [Google Scholar] [CrossRef]
- Baroni, G.; Schuinski, A.; de Moraes, T.P.; Meyer, F.; Pecoits-Filho, R. Inflammation and the peritoneal membrane: Causes and impact on structure and function during peritoneal dialysis. Mediat. Inflamm. 2012, 2012, 912595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, K.; Hamada, C. Impact of uremia, diabetes, and peritoneal dialysis itself on the pathogenesis of peritoneal sclerosis: A quantitative study of peritoneal membrane morphology. Clin. J. Am. Soc. Nephrol. 2008, 3, 720–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodrow, G.; Fan, S.L.; Reid, C.; Denning, J.; Pyrah, A.N. Renal Association Clinical Practice Guideline on peritoneal dialysis in adults and children. BMC Nephrol. 2017, 18, 333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, J.D.; Craig, K.J.; Topley, N.; Von Ruhland, C.; Fallon, M.; Newman, G.R.; MacKenzie, R.K.; Williams, G.T. Morphologic changes in the peritoneal membrane of patients with renal disease. J. Am. Soc. Nephrol. 2002, 13, 470–479. [Google Scholar] [CrossRef] [PubMed]
- De Lima, S.M.A.; Otoni, A. Inflammation, neoangiogenesis and fibrosis in peritoneal dialysis. Clin. Chim. Acta 2013, 421, 46–50. [Google Scholar] [CrossRef]
- Krediet, R.T. Ultrafiltration Failure is a Reflection of Peritoneal Alterations in Patients Treated with Peritoneal Dialysis. Front. Physiol. 2018, 9, 1815. [Google Scholar] [CrossRef] [Green Version]
- Shroff, R.; Stefanidis, C.J.; Askiti, V.; Edefonti, A.; Testa, S.; Ekim, M.; Kavaz, A.; Ariceta, G.; Bakkaloglu, S.; Fischbach, M.; et al. Encapsulating peritoneal sclerosis in children on chronic PD: A survey from the European Paediatric Dialysis Working Group. Nephrol. Dial. Transplant. 2013, 28, 1908–1914. [Google Scholar] [CrossRef] [Green Version]
- Moinuddin, Z.; Summers, A. Encapsulating peritoneal sclerosis-a rare but devastating peritoneal disease. Front. Physiol. 2015, 6, 1–12. [Google Scholar]
- Jagirdar, R.M.; Bozikas, A.; Zarogiannis, S.G.; Bartosova, M.; Schmitt, C.P.; Liakopoulos, V. Encapsulating peritoneal sclerosis: Pathophysiology and current treatment options. Int. J. Mol. Sci. 2019, 20, 5765. [Google Scholar] [CrossRef] [Green Version]
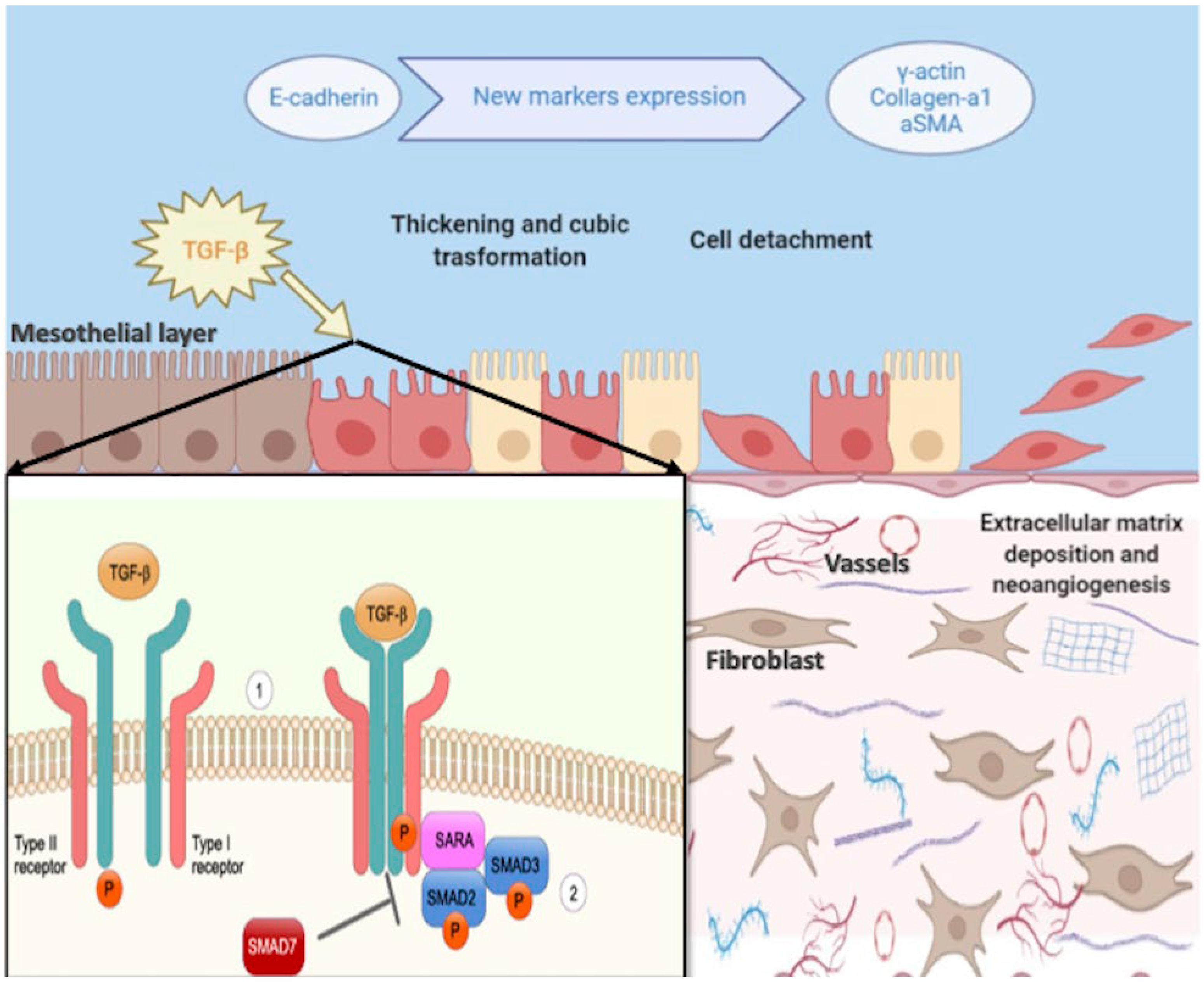
- Strippoli, R.; Moreno-Vicente, R.; Battistelli, C.; Cicchini, C.; Noce, V.; Amicone, L.; Marchetti, A.; del Pozo, M.A.; Tripodi, M. Molecular Mechanisms Underlying Peritoneal EMT and Fibrosis. Stem Cells Int. 2016, 2016, 3543678. [Google Scholar] [CrossRef] [Green Version]
- Terri, M.; Trionfetti, F.; Montaldo, C.; Cordani, M.; Tripodi, M.; Lopez-Cabrera, M.; Strippoli, R. Mechanisms of Peritoneal Fibrosis: Focus on Immune Cells–Peritoneal Stroma Interactions. Front. Immunol. 2021, 12, 607204. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Chen, W.P. Persistent transforming growth factor-beta 1 expression may predict peritoneal fibrosis in CAPD patients with frequent peritonitis occurrence. Am. J. Nephrol. 1998, 18, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Mó, M.; Lara-Pezzi, E. Peritoneal Dialysis and Epithelial-to-Mesenchymal Transition of Mesothelial Cells. N. Engl. J. Med. 2003, 348, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Jörres, A. Novel peritoneal dialysis solutions-What are the clinical implications? Blood Purif. 2012, 33, 153–159. [Google Scholar] [CrossRef]
- Piccapane, F.; Bonomini, M. A novel formulation of glucose-sparing peritoneal dialysis solutions with l-carnitine improves biocompatibility on human mesothelial cells. Int. J. Mol. Sci. 2021, 22, 123. [Google Scholar] [CrossRef]
- Libetta, C.; Esposito, P. Effects of different peritoneal dialysis fluids on the TH1/TH2 balance. Eur. Cytokine Netw. 2011, 22, 24–31. [Google Scholar] [CrossRef]
- Kim, C.D.; Kwon, H.M.; Park, S.-H.; Oh, E.-J.; Kim, M.-H.; Choi, S.-Y.; Choi, M.-J.; Kim, I.-S.; Park, M.-S.; Kim, Y.-J.; et al. Effects of low glucose degradation products peritoneal dialysis fluid on the peritoneal fibrosis and vascularization in a chronic rat model. Ther. Apher. Dial. 2007, 11, 56–64. [Google Scholar] [CrossRef]
- Provenzano, M.; Rotundo, S.; Chiodini, P.; Gagliardi, I.; Michael, A.; Angotti, E.; Borrelli, S.; Serra, R.; Foti, D.; De Sarro, G.; et al. Contribution of predictive and prognostic biomarkers to clinical research on chronic kidney disease. Int. J. Mol. Sci. 2020, 21, 5846. [Google Scholar] [CrossRef]
- Fang, X.; Lu, M.; Xia, Z.; Gao, C.; Cao, Y.; Wang, R.; Wang, M.; Wu, H. Use of liquid chromatography-tandem mass spectrometry to perform urinary proteomic analysis of children with IgA nephropathy and Henoch-Schönlein purpura nephritis. J. Proteom. 2021, 230, 103979. [Google Scholar] [CrossRef]
- Rinschen, M.M.; Limbutara, K. From molecules to mechanisms: Functional proteomics and its application to renal tubule physiology. Physiol. Rev. 2018, 98, 2571–2606. [Google Scholar] [CrossRef] [Green Version]
- Aufricht, C.; Beelen, R. Biomarker research to improve clinical outcomes of peritoneal dialysis: Consensus of the European Training and Research in Peritoneal Dialysis (EuTRiPD) network. Kidney Int. 2017, 92, 824–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreto, D.L.; Krediet, R.T. Current status and practical use of effluent biomarkers in peritoneal dialysis patients. Am. J. Kidney Dis. 2013, 62, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Krediet, R.T.; Struijk, D.G. Peritoneal changes in patients on long-term peritoneal dialysis. Nat. Rev. Nephrol. 2013, 9, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Bruschi, M.; Candiano, G.; Santucci, L.; Petretto, A.; Mangraviti, S.; Canepa, A.; Perri, K.; Ghiggeri, G.M.; Verrina, E. Proteome profile of peritoneal effluents in children on glucose- or icodextrin-based peritoneal dialysis. Nephrol. Dial. Transplant. 2011, 26, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Pearson, L.J.; Klaharn, I.Y.; Thongsawang, B.; Manuprasert, W.; Saejew, T.; Somparn, P.; Chuengsaman, P.; Kanjanabuch, T.; Pisitkun, T. Multiple extracellular vesicle types in peritoneal dialysis effluent are prominent and contain known biomarkers. PLoS ONE 2017, 12, e0178601. [Google Scholar] [CrossRef] [Green Version]
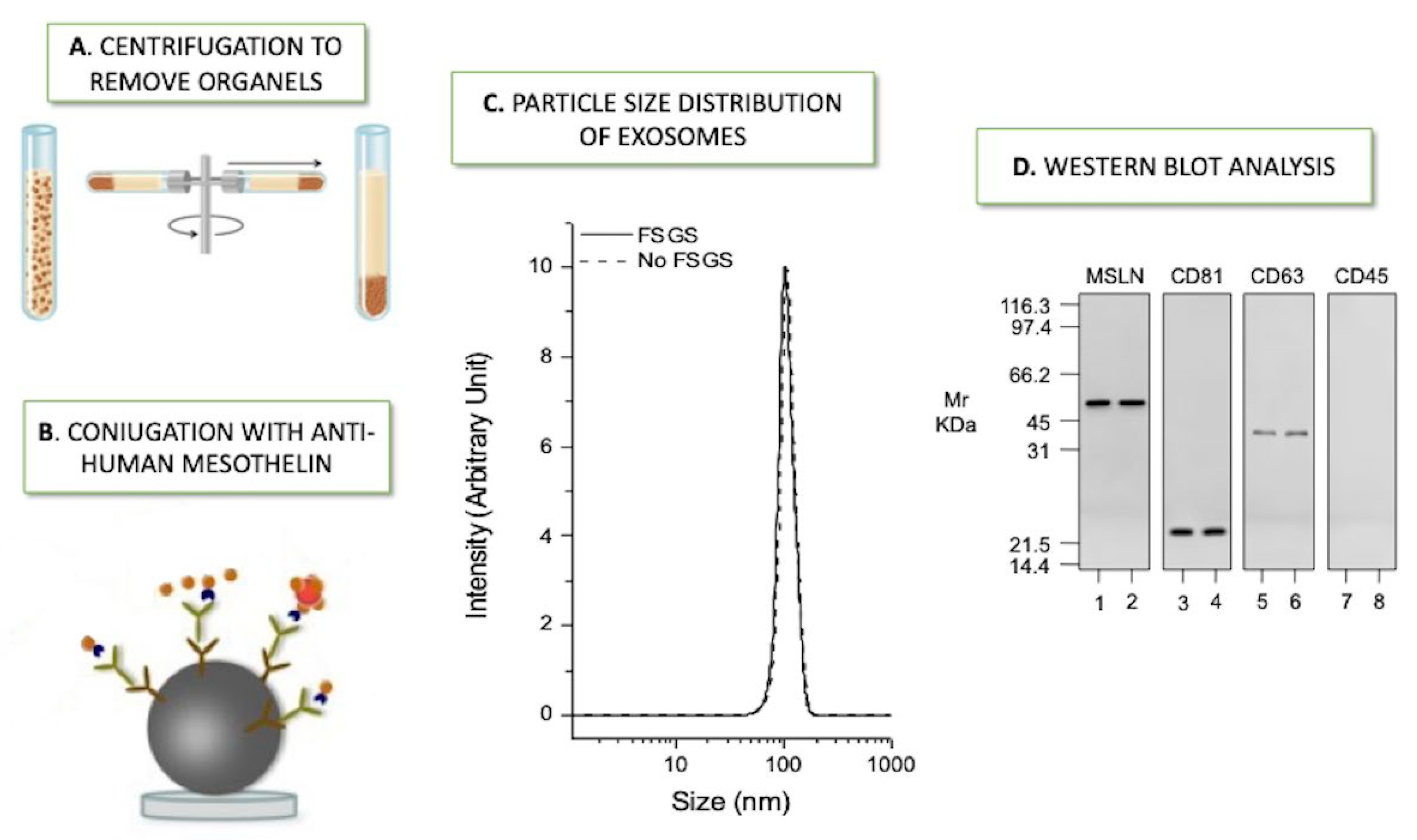
- Bruschi, M.; La Porta, E.; Panfoli, I.; Candiano, G.; Petretto, A.; Vidal, E.; Kajana, X.; Bartolucci, M.; Granata, S.; Ghiggeri, G.M.; et al. Proteomic profile of mesothelial exosomes isolated from peritoneal dialysis effluent of children with focal segmental glomerulosclerosis. Sci. Rep. 2021, 11, 20807. [Google Scholar] [CrossRef]
- Bartosova, M.; Zhang, C.; Schaefer, B.; Herzog, R.; Ridinger, D.; Damgov, I.; Levai, E.; Marinovic, I.; Eckert, C.; Romero, P.; et al. Glucose Derivative Induced Vasculopathy in Children on Chronic Peritoneal Dialysis. Circ. Res. 2021, 129, E102–E118. [Google Scholar] [CrossRef]
- Bartosova, M.; Schaefer, B.; Bermejo, J.L.; Tarantino, S.; Lasitschka, F.; Macher-Goeppinger, S.; Sinn, H.-P.; Warady, B.A.; Zaloszyc, A.; Parapatics, K.; et al. Complement activation in peritoneal dialysis—Induced arteriolopathy. J. Am. Soc. Nephrol. 2018, 29, 268–282. [Google Scholar] [CrossRef]
- Bruschi, M.; Candiano, G. Combinatorial peptide ligand library and two-dimensional electrophoresis: New frontiers in the study of peritoneal dialysis effluent in pediatric patients. J. Proteom. 2015, 116, 68–80. [Google Scholar] [CrossRef]
- Raaijmakers, R.; Pluk, W. Proteomic profiling and identification in peritoneal fluid of children treated by peritoneal dialysis. Nephrol. Dial. Transplant. 2008, 23, 2402–2405. [Google Scholar] [CrossRef] [Green Version]
- Minami, S.; Hora, K. Relationship between effluent levels of β2-microglobulin and peritoneal injury markers in 7.5% icodextrin-based peritoneal dialysis solution. Ther. Apher. Dial. 2007, 11, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Bantscheff, M.; Schirle, M.; Sweetman, G.; Rick, J.; Kuster, B. Quantitative mass spectrometry in proteomics: A critical review. Anal. Bioanal. Chem. 2007, 389, 1017–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sritippayawan, S.; Chiangjong, W. Proteomic Analysis of Peritoneal Dialysate Fluid in Patients with Different Types of Peritoneal Membranes. J. Proteome. Res. 2007, 6, 4356–4362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, W.T.; Tsai, C.C.; Chen, C.-Y.; Lee, W.-J.; Su, C.-C.; Wu, Y.-J. Proteomic analysis of peritoneal dialysate fluid in patients with dialysis-related peritonitis. Ren. Fail. 2008, 30, 772–777. [Google Scholar] [CrossRef] [Green Version]
- Carozzi, S.; Nasini, G. Peritoneal macrophage beta-2 microglobulin production and bacterial paeritonitis in CAPD patients. ASAIO Trans. 1990, 36, M369-71. [Google Scholar]
- Carozzi, S.; Nasini, G. Bacterial peritonitis and beta-2 microglobulin production by peritoneal macrophages in CAPD patients. Adv. Perit Dial. 1990, 6, 106–109. [Google Scholar]
- Zavvos, V.; Buxton, A.T. A prospective, proteomics study identified potential biomarkers of encapsulating peritoneal sclerosis in peritoneal effluent. Kidney Int. 2017, 92, 988–1002. [Google Scholar] [CrossRef] [Green Version]
- Strippoli, R.; Sandoval, P.; Moreno-Vicente, R.; Rossi, L.; Battistelli, C.; Terri, M.; Pascual-Antón, L.; Loureiro, M.; Matteini, F.; Calvo, E.; et al. Caveolin1 and YAP drive mechanically induced mesothelial to mesenchymal transition and fibrosis. Cell Death Dis. 2020, 11, 647. [Google Scholar] [CrossRef]
- Strippoli, R.; Loureiro, J. Caveolin-1 deficiency induces a MEK-ERK 1/2-Snail-1-dependent epithelial–mesenchymal transition and fibrosis during peritoneal dialysis. EMBO Mol. Med. 2015, 7, 357. [Google Scholar] [CrossRef]
- Ferrantelli, E.; Farhat, K. Effluent and serum protein N-glycosylation is associated with inflammation and peritoneal membrane transport characteristics in peritoneal dialysis patients. Sci. Rep. 2018, 8, 979. [Google Scholar] [CrossRef] [Green Version]
- Bonomini, M.; Borras, F.E.; Troya-Saborido, M.; Carreras-Planella, L.; Di Liberato, L.; Arduini, A. Proteomic research in peritoneal dialysis. Int. J. Mol. Sci. 2020, 21, 5489. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; El Andaloussi, S.; Wood, M.J. Exosomes and microvesicles: Extracellular vesicles for genetic information transfer and gene therapy. Hum. Mol. Genet. 2012, 21, R125–R134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campanella, C.; Bavisotto, C.C.; Logozzi, M.; Gammazza, A.M.; Mizzoni, D.; Cappello, F.; Fais, S. On the choice of the extracellular vesicles for therapeutic purposes. Int. J. Mol. Sci. 2019, 20, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Théry, C.; Ostrowski, M. Membrane vesicles as conveyors of immune responses. Nat. Rev. Immunol. 2009, 9, 581–593. [Google Scholar] [CrossRef]
- Hristov, M.; Erl, W. Apoptotic bodies from endothelial cells enhance the number and initiate the differentiation of human endothelial progenitor cells in vitro. Blood 2004, 104, 2761–2766. [Google Scholar] [CrossRef]
- Margetts, P.J.; Churchill, D.N. Acquired ultrafiltration dysfunction in peritoneal dialysis patients. J. Am. Soc. Nephrol. 2002, 13, 2787–2794. [Google Scholar] [CrossRef]
- Akbari, S.; Abou-Arkoub, R. Microparticle formation in peritoneal dialysis: A proof of concept study. Can. J. Kidney Health Dis. 2017, 4, 1–8. [Google Scholar] [CrossRef]
- Raeven, P.; Zipperle, J. Extracellular vesicles as markers and mediators in sepsis. Theranostics 2018, 8, 3348–3365. [Google Scholar] [CrossRef]
- Carreras-Planella, L.; Soler-Majoral, J. Proteomic profiling of peritoneal dialysis effluent-derived extracellular vesicles: A longitudinal study. J. Nephrol. 2019, 32, 1021–1031. [Google Scholar] [CrossRef]
- Peter, M.R.; Jerkic, M.; Sotov, V.; Douda, D.N.; Ardelean, D.S.; Ghamami, N.; Lakschevitz, F.; Khan, M.A.; Robertson, S.J.; Glogauer, M.; et al. Impaired resolution of inflammation in the Endoglin heterozygous mouse model of chronic colitis. Mediat. Inflamm. 2014, 2014, 767185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, J.; Tong, Y.; Ji, O.; Wei, S.; Chen, Z.; Song, A.; Li, P.; Zhang, Y.; Zhang, H.; Ruan, H.; et al. Glycoprotein 96 in Peritoneal Dialysis Effluent-Derived Extracellular Vesicles: A Tool for Evaluating Peritoneal Transport Properties and Inflammatory Status. Front. Immunol. 2022, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Fais, S.; O’Driscoll, L. Evidence-Based Clinical Use of Nanoscale Extracellular Vesicles in Nanomedicine. ACS Nano 2016, 10, 3886–3899. [Google Scholar] [CrossRef]
- Wen, J.; Ma, Z.; Livingston, M.J.; Zhang, W.; Yuan, Y.; Guo, C.; Liu, Y.; Fu, P.; Dong, Z. Decreased secretion and profibrotic activity of tubular exosomes in diabetic kidney disease. Am. J. Physiol. Ren. Physiol. 2020, 319, F664–F673. [Google Scholar] [CrossRef] [PubMed]
- Erdbrügger, U.; Blijdorp, C.J. Urinary extracellular vesicles: A position paper by the Urine Task Force of the International Society for Extracellular Vesicles. J. Extracell. Vesicles 2021, 10, e12093. [Google Scholar] [CrossRef] [PubMed]
- Miranda, K.C.; Bond, D.T.; McKee, M.; Skog, J.; Paunescu, T.; Da Silva, N.; Brown, D.; Russo, L.M. Potential Biomarkers for Renal Disease. Kidney Int. 2015, 78, 191–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corciulo, S.; Nicoletti, M.C.; Mastrofrancesco, L.; Milano, S.; Mastrodonato, M.; Carmosino, M.; Gerbino, A.; Corciulo, R.; Russo, R.; Svelto, M.; et al. AQP1-Containing Exosomes in Peritoneal Dialysis Effluent as Biomarker of Dialysis Efficiency. Cells 2019, 8, 330. [Google Scholar] [CrossRef] [Green Version]
- Gou, R.; Zhu, L. Annexin A8 can serve as potential prognostic biomarker and therapeutic target for ovarian cancer: Based on the comprehensive analysis of Annexins. J. Transl. Med. 2019, 17, 275. [Google Scholar] [CrossRef] [Green Version]
- Xue, G.L.; Zhang, C.; Zheng, G.; Zhang, L.; Bi, J. Annexin A13 predicts poor prognosis for lung adenocarcinoma patients and accelerates the proliferation and migration of lung adenocarcinoma cells by modulating epithelial—Mesenchymal transition. Fundam. Clin. Pharmacol. 2020, 34, 687–696. [Google Scholar] [CrossRef]
- Gupta, M.P.; Tandalam, S.; Ostrager, S.; Lever, A.S.; Fung, A.R.; Hurley, D.D.; Alegre, G.B.; Espinal, J.E.; Remmel, H.L.; Mukherjee, S.; et al. Non-reversible tissue fixation retains extracellular vesicles for in situ imaging. Nat. Methods 2019, 16, 1269–1273. [Google Scholar] [CrossRef]
- Hurwitz, S.N.; Olcese, J.M.; Meckes, D.G., Jr. Extraction of Extracellular Vesicles from Whole Tissue. J. Vis. Exp. 2019, 144, e59143. [Google Scholar] [CrossRef] [PubMed]
- Carreras-Planella, L.; Soler, J.; Rubio-Esteve, C.; Lozano-Ramos, S.I.; Franquesa, M.; Bonet, J.; Troya-Saborido, M.I.; Borràs, F.E. Characterization and proteomic profile of extracellular vesicles from peritoneal dialysis efflux. PLoS ONE 2017, 12, e0176987. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Biomarkers | Main Findings | Reference |
|---|---|---|
| ANXA13 | Most significant potential biomarker in detecting peritoneal dialysis effluent exosomes of FSGS from No FSGS patients | [36] |
| TIMP1 | Down-regulated protein in FSGS | [36] |
| PTP4A1 | PD vintage and decreased PM function | [36] |
| CENP-E, FCN2 | Up-regulated proteins in FSGS | [36] |
| Caspase-3, IL-6, ZO-1 | Lumen narrowing of parietal peritoneal arterioles of patients exposed to high-GDP | [37] |
| C1q and C3d | Abundance in PD-associated glucose exposure; correlation with the degree of arteriolopathy and high level of p-SMAD2/3 | [38] |
| p-SMAD2/3 | Microvasculature damage mechanisms of the peritoneum vessels | [38] |
| Intelectin-1 | Inflammation and fibrosis | [39] |
| Defensive role against intestinal bacterial permeation and against a parasite | [40] | |
| Cystatin C and B2M | Peritoneal dialysis efficiency | [41] |
| Paraoxonase | Protection against toxic oxidative modification; possible correlation with early atherosclerotic changes in peritoneal dialysis | [40] |
| Gelsolin | Protective role in mesothelial cell damage against infection | [40] |
| Study | Population/Patients and Study Design | Main Topics/Research Aim | Main Biomarkers | Proteomic Analysis | AI Methods and Approaches | Ref. |
|---|---|---|---|---|---|---|
| Fang J. et al. 2022 | ADULTS Sixty patients undergoing peritoneal dialysis (PD) divided into two groups: high/high average transport group and low/low average transport group | Proteomic analysis on PDE-EVs to identify potential biomarkers related to different Peritoneal membrane phenotypes | Glycoprotein 96 (GP96) | Liquid chromatography-tandem mass spectrometry. Mass spectrometer (Q Exactive HFX) coupled with nanoLC1200 system | Spectronaut 12.0 Pulsar (Biognosys). Data-dependent acquisition) technology. Blast2Go software was applied to associate Gene Ontology (GO) terms with the differentially expressed proteins | [62] |
| Bruschi M. et al. 2021 | PEDIATRIC 6 patients with FSGS vs. 6 pediatric patients affected by other primary renal diseases (no FSGS) | Comparative proteomic analysis of mesothelial exosomes from PDE | Annexin A13 (ANXA13), Centromere-associated protein E (CENP-E) Ficolin-2 (FNC2), inhibitor matrix metalloproteinase 1 (TIMP1) Protein Tyrosine Phosphatase 4A1 (PTP4A1) | Orbitrap Fusion Tribrid mass spectrometer (ThermoScientific) | Andromeda engine, incorporated into MaxQuant software, was used to search spectra against Uniprot human database. Weight gene co-expression network analysis (WGCNA) package in R. T-test, machine learning methods such as nonlinear support vector machine (SVM) learning, and partial least squares discriminant analysis (PLS-DA) | [36] |
| Bartosova M. et al. 2021 | PEDIATRIC 107 CKD5 patients and 90 patients on PD with PD fluids containing very low or high concentrations of GDP. | Impact of GDP on vasculopathy of children in chronic peritoneal dialysis. Microdissected arterioles were isolated for transcriptome and proteome analysis (n = 8 in CKD5, n = 6 for high-GDP, and n = 5 for low-GDP group) | Caspase-3, TGF-β-induced-pSMAD2/3 interleukin-6, zonula occludens-1 (ZO-1) | Liquid chromatography–mass spectrometry (QExactive. Thermo Fisher) | Data were submitted to PRIDE (Proteomics Identification Database). Ingenuity Pathway Analysis software (Qiagen, Hilden, Germany). Similarity data (edges representing shared genes) were generated using R and visualized using Cytoscape 3.8.0 | [37] |
| Carreras Planella L. et al. 2019 | ADULTS 11 patients. Follow-up 24 months, collecting samples every six months. | To outline the peritoneal dialysis-efflux—extravesicles (PDE-EV) proteome capacity of showing alterations much earlier than PET) | Endoglin, Thy-1 membrane glycoprotein (THY-1 or CD90) and biglycan (BGN), kininogen-1 (KNG1) | Liquid chromatography–mass spectrometry (LC-MS) (VelosOrbitrap-Thermo Fisher Scientific, Carlsbad, CA, USA) | Data were analyzed using Progenesis QI for proteomics software v3.0 (Nonlinear dynamics, Newcastle upon Tyne, UK). Peak lists generated s were analyzed with the Mascot search engine (v2.2, Matrix Science, London, UK). Protein identification was performed using the SwissProt-human database. Protein enrichment analysis was performed using Gene set enrichment analysis software (GSEA v3.0, Broad Institute, Cambridge, MA, USA) and GSEA Molecular Signatures Database (MSigDB v6.2, Broad Institute, Cambridge, MA, USA) | [60] |
| Bartosova M. et al. 2018 | PEDIATRIC Peritoneal arterioles were obtained from patients with CKD5 (n = 15), established PD (n = 15), and healthy control (n = 5) | Multi-omic analysis to understand the mechanisms of CKD-associated arteriopathy. They showed activation of the arteriolar complement system and correlation with severity of arteriolar vasculopathy in PD | C1q and C3d (terminal complement complex), pSMAD2/3 | LC-MS. Q Exactive mass spectrometer (Thermo Fisher Scientific, Carlsbad, CA, USA) | Data were processed and searched against the human SwissProt database with Andromeda search engine using MaxQuant. Gene enrichment analyses were conducted using the PANTHER online database. | [38] |
| Pearson LJ. et al. 2017 | ADULTS 8 patients on stable PD | To demonstrate EVs in PDE and to characterize the related proteome | Mesothelin and cancer cell antigen 125 (MUC16). vWF, CD109, CD14 and its coactivator lipopolysaccharide binding protein. EMT-related proteins: E-cadherin and extracellular matrix proteins collagen I and III, respectively. αSMA, TGF-β related proteins. | LC-MS. mass spectrometer (Q Exactive Plus Hybrid Quadrupole-Orbitrap, Thermo Fisher Scientific, Carlsbad, CA, USA) through an EASY-Spray nanoelectrospray ion source (Thermo Fisher Scientific). | Raw data were searched by X! Tandem (CYCLONE, 2013.2.01) against human databases (ENSEMBL v.76 Homo sapiens GRCh38). Gene ontology was performed using David bioinformatics resource 6.8. Protein lists generated were compared using gene ontology enrichment analysis and visualization tool, GOrilla | [35] |
| Carreras-Planella L. et al. 2017 | ADULTS 9 patients in PD divided in two groups: Newly-enrolled Patients and Longer-treated Patients | Identifying, isolating, and characterizing peritoneal dialysis efflux-extravesicles of patients on PD | CD9 CD63- CD81 Galectin 3-binding protein (LGALS3BP) Ezrin (EZR) | LC-MS/MS on a LTQ Orbitrap Velos (Thermo Fisher, Carlsbad, CA, USA). | Data were analyzed with Max Quant software against Uniprot human database. Further analyses were made using the Intensity-Based Absolute Quantification (iBAQ) values obtained from MaxQuant, and analyzed using Perseus software (version 1.5.6.0), InteractiVenn, and the EVs specific databases EVpedia, Exocarta, and Vesiclepedia. Subsequent analysis was performed Non-supervised hierarchical clustering approach | [72] |
| Bruschi M. et al. 2015 | PEDIATRIC 19 patients with different kidney disease (primarily kidney dysplasia in 6 cases, nephronophthisis in 5 cases). | Proteomic characterization of PDE samples collected in patients with different APD treatment by the combined use of Combinatorial Peptide Ligand Library (CPLL) technology and two-dimensional electrophoresis | Gelsolin, intelectin-1 | Matrix-assisted laser-desorption ionization (MALDI)—Mass Spectrometry analy- sis. | PDQuest Advance software for 2-DE experiments and QuantyOne software (Bio-Rad, Hercules, CA, USA) for western blot experiments. | [39] |
| Bruschi M. et al. 2011 | PEDIATRIC 16 patients with different kidney diseases | Proteome profile of PDE obtained with icodextrin or glucose-based solutions | β2-microglobulin cystatin C leptin. | LTQ linear ion trap mass spectrometer (Thermo Electron, San Jose, CA, USA) coupled to an HPLC Surveyor (Thermo Electron) | Protein identification was performed using SEQUEST software (Thermo Electron, San Jose, CA, USA) All digitalized images were analyzed with PDQuest Advance or QuantyOne software (Bio-Rad, Hercules, CA, USA).. | [34] |
| Raaijmakers R. et al. 2008 | PEDIATRIC 9 patients in PD | To obtain the first representative overview of the proteome of PDE. Identified proteins in PDE reflect local peritoneal processes | gelsolin intelectin paraoxonase | cyclotron resonance mass spectrometer (LIT FT-ICR MS) | Data were searched against the NCBI database using the Mascot search program. Protein identifications were validated and clustered using the PROVALT algorithm. Gene ontology classifications were made with Protein Center (www.proxeon.com, accessed on 1 April 2022). To provide an estimation of protein concentration exponentially modified protein abundance index (emPAI) was used | [40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trincianti, C.; Meleca, V.; La Porta, E.; Bruschi, M.; Candiano, G.; Garbarino, A.; Kajana, X.; Preda, A.; Lugani, F.; Ghiggeri, G.M.; et al. Proteomics and Extracellular Vesicles as Novel Biomarker Sources in Peritoneal Dialysis in Children. Int. J. Mol. Sci. 2022, 23, 5655. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105655
Trincianti C, Meleca V, La Porta E, Bruschi M, Candiano G, Garbarino A, Kajana X, Preda A, Lugani F, Ghiggeri GM, et al. Proteomics and Extracellular Vesicles as Novel Biomarker Sources in Peritoneal Dialysis in Children. International Journal of Molecular Sciences. 2022; 23(10):5655. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105655
Chicago/Turabian StyleTrincianti, Chiara, Vincenzo Meleca, Edoardo La Porta, Maurizio Bruschi, Giovanni Candiano, Andrea Garbarino, Xhuliana Kajana, Alberto Preda, Francesca Lugani, Gian Marco Ghiggeri, and et al. 2022. "Proteomics and Extracellular Vesicles as Novel Biomarker Sources in Peritoneal Dialysis in Children" International Journal of Molecular Sciences 23, no. 10: 5655. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105655