1. Introduction
Realgar is a traditional Chinese medicine. In the clinic, it has been used for the treatment of acute promyelocytic leukemia (APL) and other cancers [
1]. Realgar is frequently used in combination with herbs to achieve better efficacy and reduce toxicity. In the 2015 edition of the Pharmacopeia of China (Volume 1), approximately 2.48% of the recipes contain realgar among the listed total of 1439 traditional recipes [
2]. The major component of realgar is disulfide arsenic (As
2S
2) [
3]. Although the toxicity of realgar is far less than that of arsenic, increasing evidence supports that the arsenic in realgar can be absorbed into the blood and accumulated in various organs causing systemic damage, predominantly liver, kidney and nerve damage [
4]. The liver is the main site of arsenic metabolism as well as the major target of realgar toxicity. More and more studies have reported that the mice exposed to realgar exhibited obvious liver damage [
5,
6,
7]. However, the mechanism of realgar-induced liver injury and the therapeutic target has not been completely elucidated.
The nucleotide-binding oligomerization domain (NOD)-like pyrin domain-containing protein 3 (NLRP3) inflammasome, which consists of NLRP3, adaptor apoptosis-associated speck-like protein (ASC), and pro-caspase-1 is deemed as a vital driving factor of various liver diseases and chemical/toxin-induced liver injury [
8]. NLRP3 recruits and cleaves pro-caspase-1 into its active forms, which cleaves pro-interleukin-1β (pro-IL-1β) and interleukin-18 (pro-IL-18) into mature interleukin-1β (IL-1β) and interleukin-18 (IL-18), and finally triggers inflammation occurrence [
9]. Furthermore, the active-caspase-1 cleaves gasdermin D (GSDMD) into an N-terminal fragment (N-GSDMD), leading to the formation of membrane pores to mediate pyroptosis. Our recent study has demonstrated that sub-chronic realgar exposure upregulated the NLRP3 inflammasome and promoted the expression and release of IL-1β, leading to inflammatory damage in mouse livers [
7].
Autophagy is a process that eliminates damaged organelles and proteins through cytoplasmic degradation [
10]. Increasing evidence has revealed that there is a close relationship between autophagy and NLRP3 inflammasome activation [
11]. A number of studies have shown that autophagy has a protective effect on exogenous or drugs/toxicant-induced liver injury [
12]. However, till now, little is known about the role of autophagy in realgar-induced liver injury and the relationship between autophagy and NLRP3 inflammasome activation in liver tissues of realgar-exposed mice.
Therefore, in this study, 3-methyladenine (3-MA, an inhibitor of autophagy) or rapamycin (RAPA, an inducer of autophagy) was used to regulate autophagy in realgar- induced liver injury mice model to explore the function of autophagy in realgar-induced liver injury and elucidate the relationship between autophagy and NLRP3 inflammasome activation in liver tissues. We aimed to provide insightful viewpoints into the mechanism and therapeutic target of realgar-induced liver injury.
2. Material and Methods
2.1. Chemical and Reagents
Water-processed realgar was purchased from Sanmenxia Yuhuangshan Pharmaceutical Corporation (Sanmenxia, Henan, China, Lot: W1-180321). N-acetylcysteine (NAC, purity ≥ 98%, #PHR1098) was purchased from Sigma (St. Louis, MO, USA). 3-MA (purity ≥ 99%, #T1879) and Rapa (RAPA, purity ≥ 98%, #T1537) were purchased from TargetMol (Shanghai, China). Alanine aminotransferase (ALT, #c009-2-1), aspartate aminotransferase (AST, #c010-2-1), alkaline phosphatase (ALP, #A059-1-1), total bilirubin (TBIL, #c019-1), lactate lactic dehydrogenase (LDH, #A020-2-2), reactive oxygen species (ROS, #E004-1-1) kits were purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing, China). Assay kits for urea (UN, #R21723-100T) and albumin (ALB, #CEB028Rb) were obtained from Shenyang BoKe Biological Technology Co. Itd and Shenyang Bolande Trading Co., Ltd. (Shenyang, China). Mouse γ- glutamyl transferase (GGT, #JM-11634M2) ELISA Kits were provided by JIANGSU JINGMEI Biotechnology Co., Ltd. (Yancheng, Jiangsu, China). Mouse serum creatinine (Cr, #RX-28589) ELISA kits were purchased from Mode Biological Technology Co. Itd (Shenyang, China). IL-1β (#EK0394) ELISA Array Kits were purchased from Boster Biological Technology Co. Itd (Wuhan, China). Monodansylcadaverine (MDC, #G0170) was purchased from solarbio (Beijing, China). PrimeScript™ RT reagent Kit with gDNA Eraser (#RR820A), TB Green® Premix Ex Taq™ II (RR047A) were purchased from TaKaRa Biotechnology Company (Dalian, China). Antibodies against NLRP3 (#19771-1-AP), Caspase-1 (#22915-1-AP), thioredoxin (TRX, #14999-1-AP), p62 (#18420-1-AP), LC3 (#14600-1-AP), vacuolar protein sorting 34 (VPS34, #12452-1-AP), the mammalian target of rapamycin (mTOR, #66888-1-Ig), nuclear receptor-κB p65 (NF-κB p65, #AF5006) and Lamin B1 (#12987-1-AP) were purchased from Proteintech (Wuhan, China). Antibodies against ASC (#67824) and thioredoxin-interacting protein (TXNIP, #14715) were obtained from Cell Signaling Technology (Beverly, MA, USA). Antibodies against IL-1β (#ab254360), IL-18 (#ab207323) and GSDMD (#ab209845) were obtained from Abcam (Cambridge, UK). Pure Proteome Protein A/G Mix Magnetic Beads (#LSKMAGAG02) were purchased from Merck Millipore (Burlington, MA, USA).
2.2. Animals and Treatments
Six-week-old healthy male Kunming (KM) mice (weighing 20~22 g) were obtained from the Laboratory Animal Center of China Medical University (Permission No SCXK2015-0001; Shenyang, Liaoning, China). The experimental mice were housed under SPF conditions under a 12 h light-dark cycle (8:00am–8:00pm) in an air-conditioned room with a room temperature of (25 ± 2) °C and (55 ± 10)% humidity and providing free access to food and water. The experimental protocol was processed according to the National Institute of Health Guide for the Care and Use of laboratory Animals (NIH Publication No.80-23, 1996) and approved by the Animal Ethics Committee of China Medical University. All efforts were exerted to minimize the number of mice used and their suffering.
After one week of adaptive feeding, all mice were randomly divided into the following eight groups: (1) Control group; (2) Realgar group; (3) NAC group; (4) NAC+Realgar group; (5) 3-MA group; (6) 3-MA+Realgar group; (7) RAPA group, and (8) RAPA+Realgar group. Mice in the Control, NAC, 3-MA and RAPA groups received 0.5% CMC-Na; mice in the Realgar, NAC+Realgar, 3-MA+Realgar and RAPA+Realgar groups received 1.35 g/kg realgar by gavage once daily for 8 consecutive weeks. For the intervention studies, NAC, 3-MA, and RAPA were dissolved in 0.5% dimethylsulfoxide (DMSO). Mice in the NAC and NAC+Realgar groups were injected intraperitoneally with NAC at a dose of 20 mg/kg; mice in the RAPA and RAPA+Realgar groups were injected intraperitoneally with RAPA at a dose of 2 mg/kg; mice in the 3-MA and 3-MA+Realgar groups were injected intraperitoneally with 3-MA at a dose of 15 mg/kg. Mice in the Control and Realgar groups were injected intraperitoneally with 0.5% DMSO. All injections were given every two days. NAC was given for eight weeks, while 3-MA and RAPA were given for three weeks before isoflurane anaesthesia.
2.3. Measurement of Liver Index and ALT, AST, ALP, γ-GGT, TBIL, ALB, UN, Cr, LDH, IL-1β in Plasma
The liver was removed carefully and rinsed with phosphate buffer. Then, the liver weights were measured after the liver tissues were dried with filter paper to remove the excess solution. The liver index was calculated as follows: liver index% = liver mass (g)/body mass (g) × 100%. The plasma ALT, AST, ALP, γ-GGT, TBIL, ALB, UN, Cr, LDH and IL-1β levels were detected with commercial kits on the basis of the manufacturer’s instructions.
2.4. Histopathological Examination
The heart perfusion-fix technique was used to prepare the specimen for histopathological examination. The PBS was perfused initially to clear the blood from the liver followed by 4% paraformaldehyde and 2.5% (w/v) glutaraldehyde buffer. After perfusion for about 20 min, the liver was excised. Cross-sections (5 µm) of the tissue were cut and stained with hematoxylin and eosin (H&E) and then assessed under a microscope (OLYMPUS, Tokyo, Japan). For transmission electron microscopy (TEM) observation, the liver specimen was cut into 1 mm3 in size and fixed in 1% osmium tetroxide. Next, the samples were dehydrated through a graded series of ethanol and acetone and then embedded in the epoxy resin. After being sliced into 1-mm-thick sections and stained with lead citrate, the samples were detected by Hitachi (Ibaraki, Japan) transmission electron microscope.
2.5. Evaluation of ROS in Mouse Livers
The intracellular ROS was determined by detecting the fluorescent intensity of 2,7-dichlorodihydrofluorescein diacetate (DCFH-DA). Each liver tissue sample was homogenized in ice-cooled Tris-HCl buffer (40 mM, pH = 7.4, 4 °C). The homogenate 190 μL was mixed with 10 μL DCFH-DA working solution (1 mmol/L), and then incubated in the dark at 37 °C for 30 min. Finally, the fluorescence intensity of DCF was assessed using a FLUOstar Omega® multi-functional microplate reader (Biotek, Winooski, VT, USA) with an excitation wavelength of 502 nm and emission wavelength of 530 nm.
2.6. MDC Measurement
The intensity of autophagic vacuoles (AVOs) was measured with MDC. The fresh liver tissue was minced in PBS and digested in 1 mL of trypsin solution at 37 °C for 30 min. Afterwards, 5 mL of DEME was added to terminate digestion. After filtration with sieves, the samples were washed two times using cold PBS and resuspended in binding buffer. The number of viable cells was counted and adjusted to 1 × 106 cells/mL. The single-cell suspension was incubated with MDC (50 μM) for 45 min at room temperature avoiding light, followed by measuring with the help of a microplate reader (Biotek, Winooski, VT, USA) at the excitation wavelength of 335 nm, and the emission wavelength of 518 nm.
2.7. RT-qPCR Assay
For RNA extraction, total RNAs from mouse liver tissue were extracted by Trizol Reagent (TaKaRa, Maebashi, Japan) according to the manufacturer’s instructions and the concentration of RNA was calculated. Approximately 1 μg of total RNA from each sample was reverse transcribed to cDNA by the PrimeScript RT reagent kit (TaKaRa, Maebashi, Japan). The levels of mRNA expression were quantified using SYBR Green PCR Master Mix and an ABI prim 7500 Sequence Detection System (Applied Biosystems, Waltham, MA, USA). The PCR program included 95 °C for 5 s and then 95 °C for 30 s and 60 °C for 30 s for 40 cycles. Relative expression of target genes was normalized to
Gapdh, analyzed by the 2
−ΔΔCt method and presented as a ratio compared with the control. The primers of target genes were designed by Sangon Biotech (Shanghai, China) and the sequences are shown in
Table 1.
2.8. Western Blot
After treatment, RIPA lysis buffer supplemented with protease and phosphatase inhibitors was used to extract the liver protein samples of mice according to the standard procedure. A BCA protein detection kit (Dingguo Changsheng, Beijing, China) was used for the quantification of protein content. After SDS-PAGE electrophoresis, the protein was transferred onto the PVDF membrane. The membranes were blocked using 5% skim milk for 1 h at room temperature. Then, the membrane was hatched by a specific primary antibody at 4 °C overnight. After washing three times with TBST, the membranes were incubated with horseradish peroxidase-conjugated secondary antibodies (Dingguo Changsheng, Beijing, China) for 1 h. The following primary antibodies were used: NLRP3 (1:1000), ASC (1:1000), Caspase-1 (1:1000), IL-1β (1:1000), GSDMD (1:1000), NF-κB (1:1000), TRX (1:1000), TXNIP (1:1000), IL-18 (1:1000), P62 (1:1000), LC3B (1:1500), VPS34 (1:1000), mTOR (1:5000), GAPDH (1:3000), and Lambin B1 (1:2500). GAPDH was used as an invariant control for the target proteins. The expressions of nuclear NF-κB proteins were normalized to Lambin B1 as a reference.
2.9. Co-Immunoprecipitation (Co-IP)
In order to detect the protein-protein interactions, a Co-IP assay was performed. RIPA lysis of 1 mL was added to 0.05 g of the liver sample, and then homogenized and centrifuged at 12,000 rpm at 4 °C for 10 min. The supernatant was collected and incubated with mouse monoclonal TXNIP antibody or IgG control antibody for 4 h at 4 °C. Then 20 μL of protein A/G beads (Santa Cruz Biotechnology, Dallas, TX, USA) were added to the protein-antibody complex and incubated for 2 h at 4 °C. Then, the protein-bead mixture was centrifuged and the supernatant was removed. The beads were washed three times with PBST buffer followed by denaturation with 1 × SDS-containing loading buffer at 100 °C for 5 min. Then, the expressions of captured proteins were assessed by Western blot. TXNIP-binding proteins from protein extract were immunoprecipitated with TXNIP antibodies. NLRP3 and TRX levels in pulled-down proteins were measured using Western blot analysis with NLRP3 and TRX antibody.
2.10. Statistical Analysis
Data are presented as the mean ± standard error (SEM). IBM SPSS 24.0 software (IBM, Armonk, NY, USA) was used for data analysis. The graphs were made with GraphPad Prism 5.0 software. The Student’s t-test was used for statistical analysis of two groups. p < 0.05 was considered statistically significant.
4. Discussion
Arsenic is a highly toxic substance, which has been classified as a class I carcinogen by the International Agency for Research on Cancer (IARC). As the main organ of arsenic metabolism and accumulation, the liver is easily affected by arsenic toxicity. Epidemiology studies have indicated a close link between chronic arsenic exposure and abnormal liver function, hepatomegaly, hepatoportal sclerosis, liver fibrosis, cirrhosis and liver cancer in humans [
16]. Medicinal exposure is one of the sources of arsenic exposure in humans. Arsenic trioxide (ATO) is a valuable drug in traditional Chinese medicine (TCM), which is often used to treat APL and other cancers [
17]. There is a report that for long-term use (>1 month), ATO induced liver damage in about 7.9% of cases [
18].
Many herbal medicines also contain arsenic, such as Salviae Miltiorrhizae, Safflower, Gentiana, Flower of Silktree Albizzia, etc. [
19]. Realgar is the most famous arsenic-containing drug, which is the essential component of some popular medicinal preparations in Asian and Western countries for the treatment of APL [
20]. Realgar has been widely used in diverse formulae in traditional Chinese medicine (TCM), such as
Niuhuang Jiedu tablet,
Angong Niuhuang Wan, etc. These realgar-containing TCMs have shown beneficial effects on detoxication, insecticidal, dampness elimination and dispelling phlegm [
1]. Studies have shown that realgar has multi-system toxicity [
4]. Excessive or long-term exposure to realgar has detrimental influences on the liver [
4]. Autophagy is a process of “self-digestion” in which proteins, organelles and other cellular components damaged are transported to lysosomes for degradation. There is evidence that autophagy has a protective effect on drug-induced liver injury [
21]. To date, the direct effect of autophagy on realgar-induced liver injury has not been studied.
In previous studies, we found that mouse exposure to realgar at 1.35 g/kg for 8 weeks exhibits obvious liver injury [
5,
7,
22,
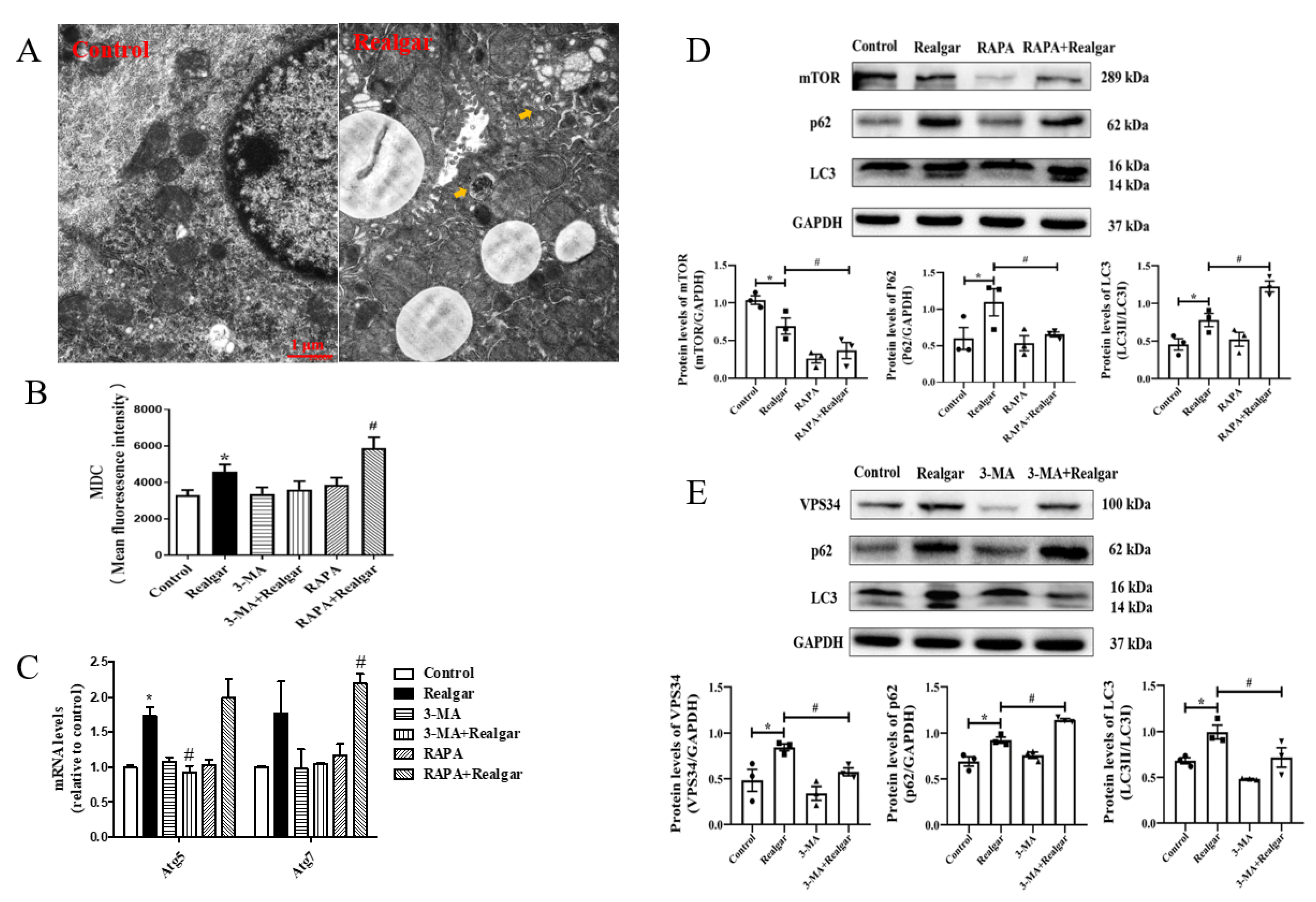
23]. Herein, mice were given 1.35 g/kg realgar for 8 weeks to induce liver injury. We first examined the effect of realgar exposure on autophagy in mouse livers. TEM is the most classical method for observing autophagic vacuoles. Under TEM, autophagosomes have a double membrane containing morphologically intact cytoplasm, including ribosomes and rough ER, and the limiting membrane that is partially visible as two bilayers separated by a narrow electron-lucent cleft [
24]. Autolysosomes typically have only one limiting membrane, which usually contains electron-dense cytoplasmic material and/or organelles at various stages of degradation [
25]. In the experiment, an increase in the numbers of autophagic vacuoles in mouse livers after realgar exposure was observed (shown in
Figure 1A). MDC, which can accumulate as a selective fluorescent marker for autophagic vacuoles under in vivo conditions by interacting with membrane lipids that are highly concentrated in the autophagic compartments [
26], is widely used to detect autophagic vacuoles. In our experiment, the intensity of MDC fluorescence was remarkably elevated in the realgar group as compared with controls (
Figure 1B), which further confirmed that realgar led to increased autophagic vacuoles accumulation in mouse livers. In addition, we further detected autophagy-related markers to determine the effect of realgar on autophagy. Vacuolar protein sorting 34 (VPS34) plays an essential role in phagophores nucleation and elongation [
27]. In the nucleation process, VPS34 forms a multiprotein complex with Beclin-1. It was found that realgar markedly upregulated the protein expression of VPS34 in mouse livers (
Figure 1E). Atg5 and Atg7 are involved in the expansion of the phagophore. The Atg5 conjugated by Atg12 is present on the outer side of phagophores and essential for elongation [
28]. Under stress conditions, Atg proteins are activated, resulting in the formation and maturation of autophagosomes from phagophores. In our study, we found that realgar upregulated Atg5 and Atg7 mRNA levels (
Figure 1C), suggesting that realgar promotes phagophores elongation. LC3, a marker of the autophagosome, is conversed from LC3I to LC3II when autophagy is induced. Therefore, the ratio of LC3II and LC3I is used as a sign of autophagy induction [
29]. In the realgar-exposed mouse livers, the ratio of LC3II to LC3I is remarkably elevated in comparison to that in the control mice (
Figure 1D,E), suggesting an induction of autophagy by realgar treatment. P62 serves as an autophagy receptor protein, which can selectively recognize autophagic cargo and mediate its engulfment into autophagosomes to initiate the degradation [
30]. P62 accumulates when autophagy is inhibited, and the level of P62 decreases when autophagy is induced. In this study, a significantly upregulated expression of P62 was found in realgar exposed mouse livers (
Figure 1D,E), indicating that the autophagy-lysosome pathway was inhibited and autophagy dysfunction occurred in the liver of mice exposed to realgar. This is consistent with the increased numbers of autophagic vacuoles in realgar exposed mouse livers observed by electron microscopy. Similar results were found in the cerebral cortex of realgar exposed rats, which suggests that realgar exposure promotes the initiation of autophagy and the bilayer membrane structure of autophagosome, but blocks autophagy degradation functions [
31]. It is well known that autophagy can be induced under stressful conditions, such as oxidative stress, and plays an important role in the modulation of cell death and survival. It has been reported in our previous study that the level of GSH and the activities of GSH-Px, SOD were significantly decreased and the level of ROS markedly increased in the liver tissues of mice treated with the same realgar dose and exposure time as this study [
7,
23]. Therefore, it is conceivable that autophagy enhancement in realgar-exposed mouse livers is likely a stress-protective response under extensive oxidative stress induced by realgar [
5].
RAPA and 3-MA were used to regulate the level of autophagy in the livers of realgar-exposed mice. 3-MA is the most commonly used autophagy inhibitor, and it functions by inhibiting VPS34. There is a report that pointed out that the generation of phosphatidyl-inositol-3-phosphate (PI3P) by VPS34 and the recruitment of PI3P-binding proteins is a critical step for phagophores nucleation and elongation [
27]. Therefore, 3-MA inhibited autophagy in an early stage. In this experiment, we found that 3-MA significantly inhibited the levels of Atg5 mRNA and VPS34, LC3II/I proteins, while upregulating the expression of P62 in realgar-exposed mouse livers (
Figure 1C–E), suggesting that 3-MA inhibits autophagy in realgar exposed mouse livers. RAPA is an inhibitor of the mammalian target of rapamycin (mTOR), which can induce autophagy by inhibiting the mTOR signaling pathway. After RAPA treatment, the intensity of MDC fluorescence, Atg7 mRNA and LC3II/I ratio were further increased in realgar exposed mouse livers, while the expression of P62 was decreased (
Figure 1C–E). The results showed that RAPA could significantly enhance autophagy in the liver tissues of mice from the Realgar group.
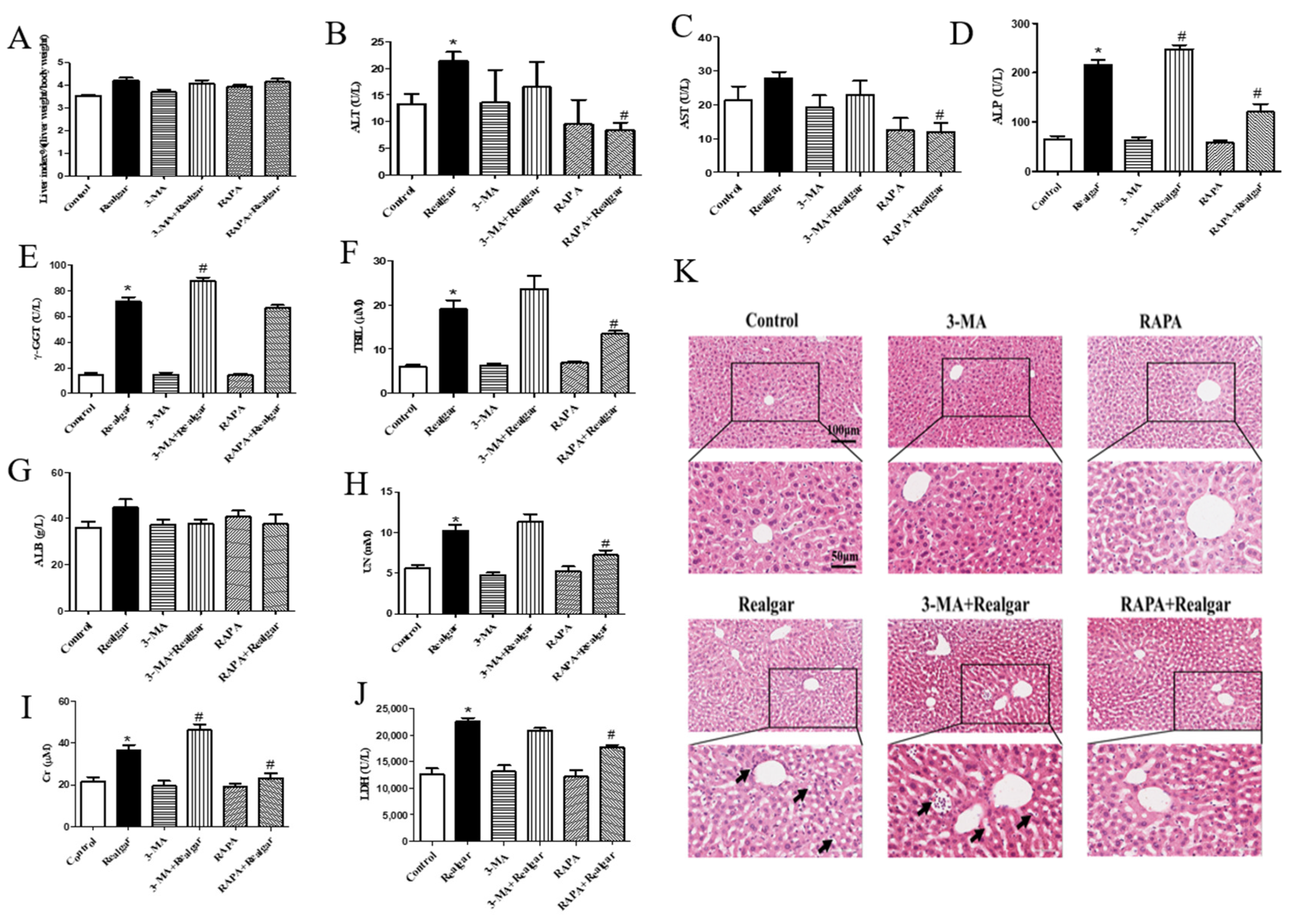
Next, we investigated the effect of autophagy on realgar-induced liver injury. It was found that after 8-week realgar exposure, the plasma ALT, ALP, γ-GGT activities and TBIL, UN, Cr, LDH levels are all apparently increased. In particular, the ALP, γ-GGT activities and TBIL levels are far higher than those in the controls, which may indicate that realgar may induce cholestatic liver injury in mouse livers [
32]. The results are consistent with our previous study that reported a disturbance in bile acid synthesis and metabolism in liver tissues of mice treated with the same realgar dose and exposure time as this study [
7]. RAPA and 3-MA intervention significantly altered the liver functions of realgar-exposed mice. It was found that RAPA markedly restored the elevated levels of ALT, AST, ALP, TBIL, UN, Cr and LDH in mouse plasma induced by realgar, while 3-MA aggravates realgar-induced liver function disorder by further increasing the levels of ALP, γ-GGT, TBIL and Cr in plasma of realgar exposed mice. H&E staining results also suggested that RAPA mitigated realgar-induced liver injury, whereas 3-MA exacerbated. Inflammation is regarded as one of the major mechanisms of realgar-induced liver injury. NLRP3 inflammasome plays an important role in initiating and sustaining inflammation [
33]. A previous study has reported that realgar-induced liver injury in mice is related to the activation of the NLRP3 inflammasome pathway [
7]. Autophagy is closely related to the activation of NLRP3 inflammasome [
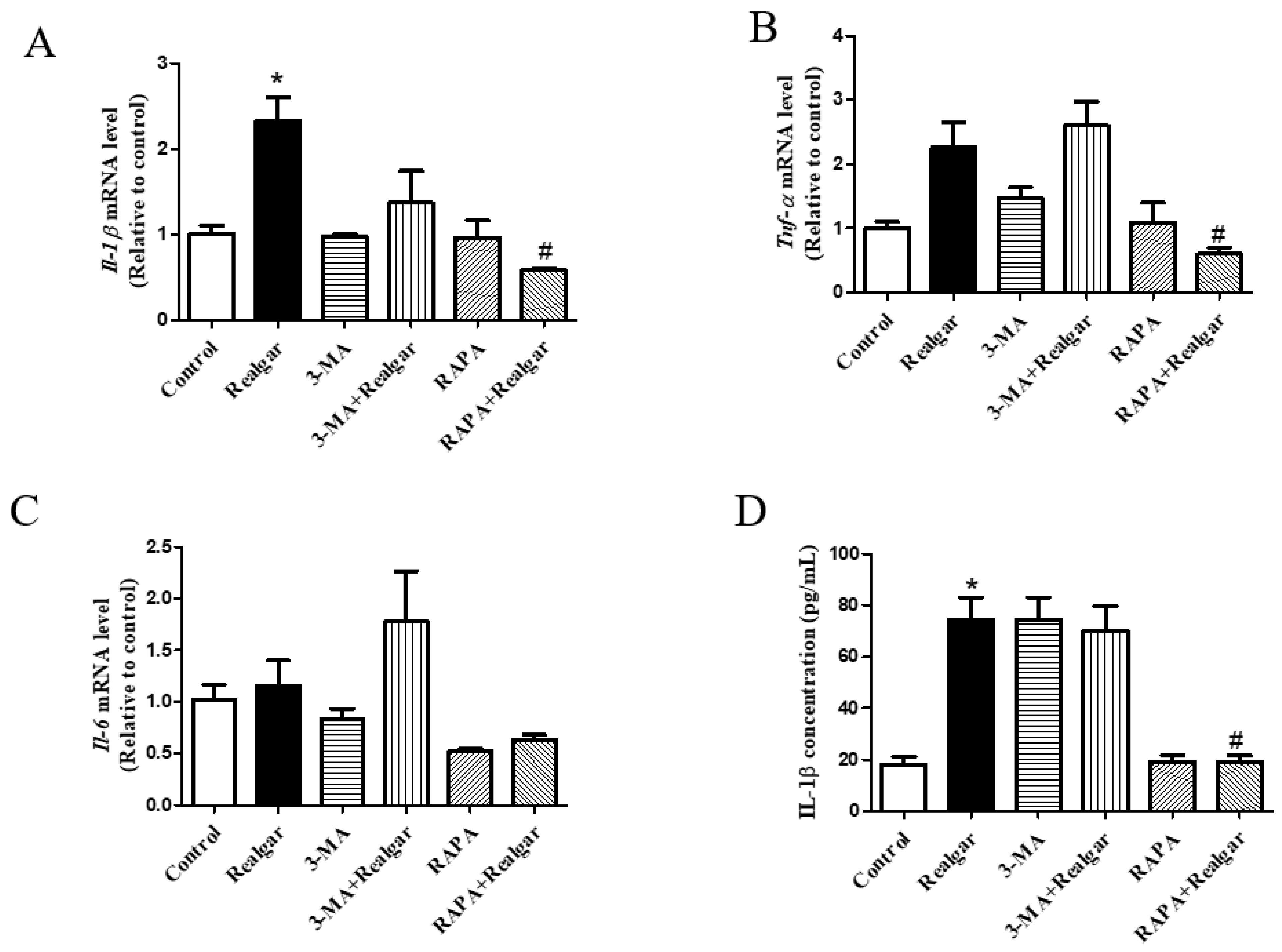
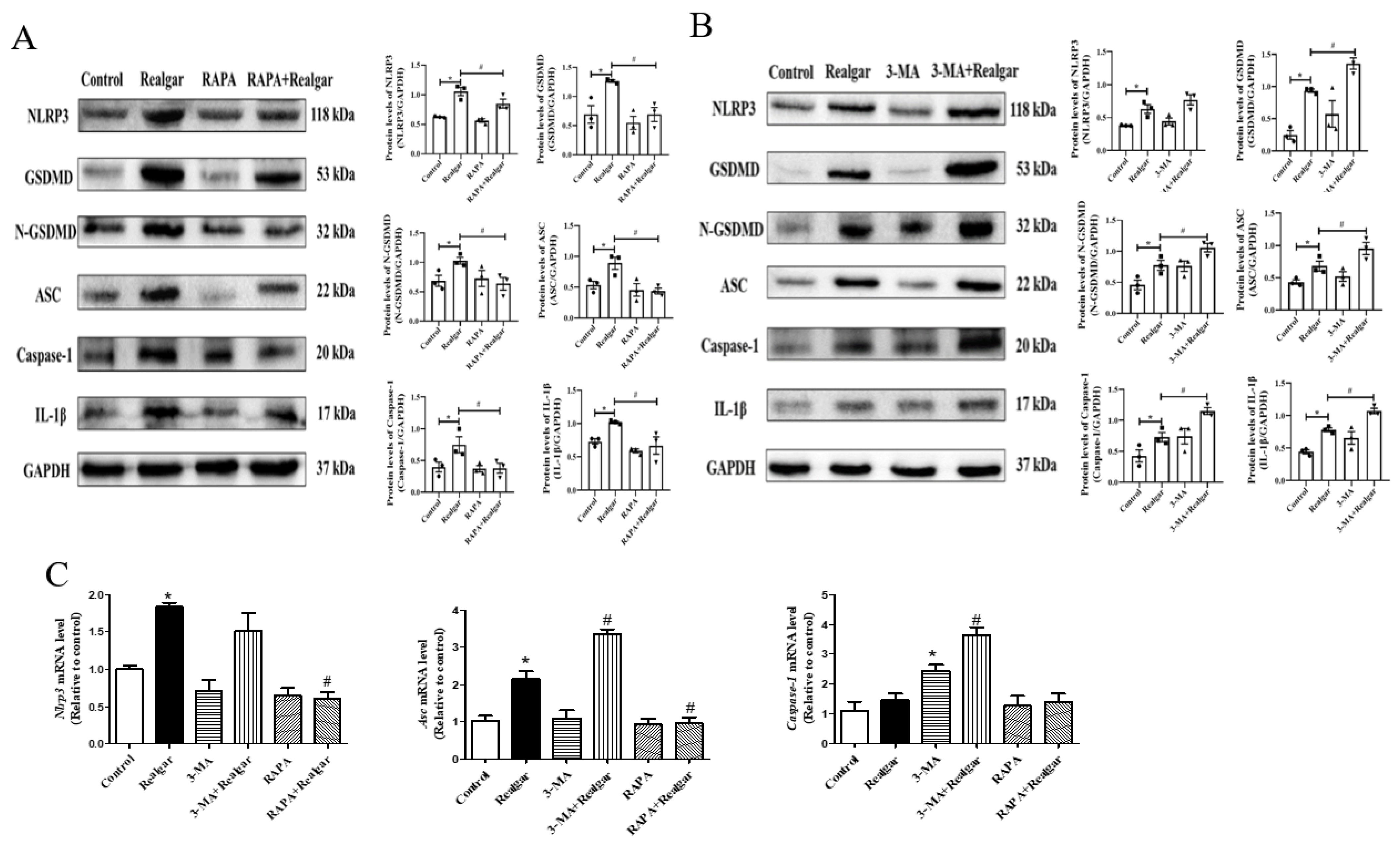
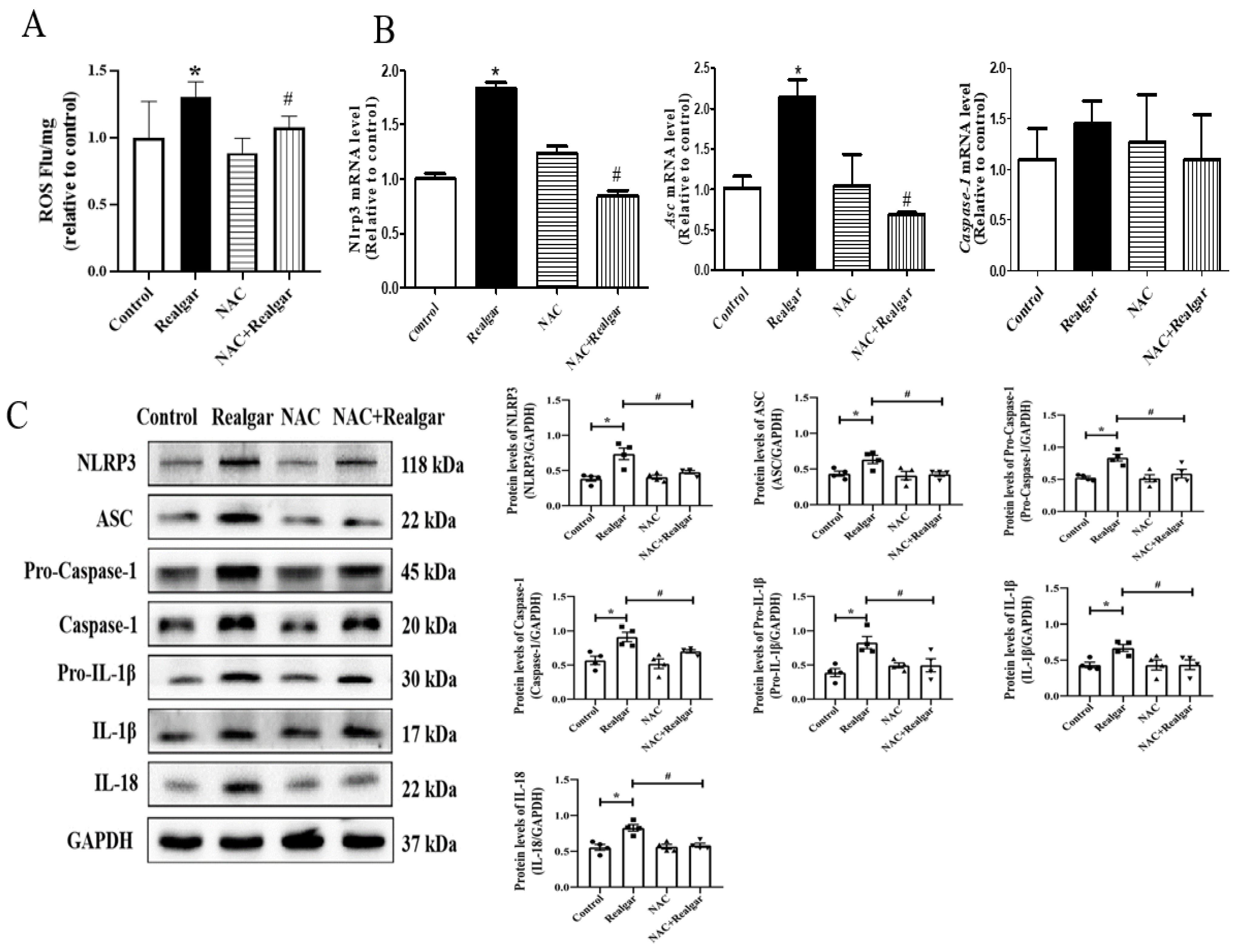
11]. The effect of autophagy on NLRP3 inflammasome activation induced by realgar has not been investigated. In this study, we found that autophagy enhancement by RAPA treatment mitigated realgar-induced NLRP3 inflammasome activation in mouse livers and reduced the expression and release of inflammatory cytokines. Oppositely, autophagy inhibition by 3-MA treatment aggravated realgar-induced NLRP3 inflammasome activation in mouse livers and promoted the expression and release of inflammatory cytokines. These results suggest that the ameliorative effect of autophagy on realgar-induced liver injury may be attributed to the inhibition of NLRP3 inflammasome activation.
To elucidate the mechanism by which autophagy inhibits the activation of NLRP3 inflammasome in realgar-induced liver injury mice, we focus on the role of ROS. As we know, ROS is one of the major endogenous danger-associated molecular patterns (DAMPs) that leads to the activation of NLRP3 [
34]. In order to clarify whether ROS production is an upstream mechanism for NLRP3 activation induced by realgar, we used NAC, an ROS scavenger, to intervene. The results illustrated that NAC intervention obviously inhibited NLRP3 inflammasome activation in realgar-exposed mouse liver, which suggests that realgar-induced NLRP3 inflammasome activation in mice liver is mediated by ROS.
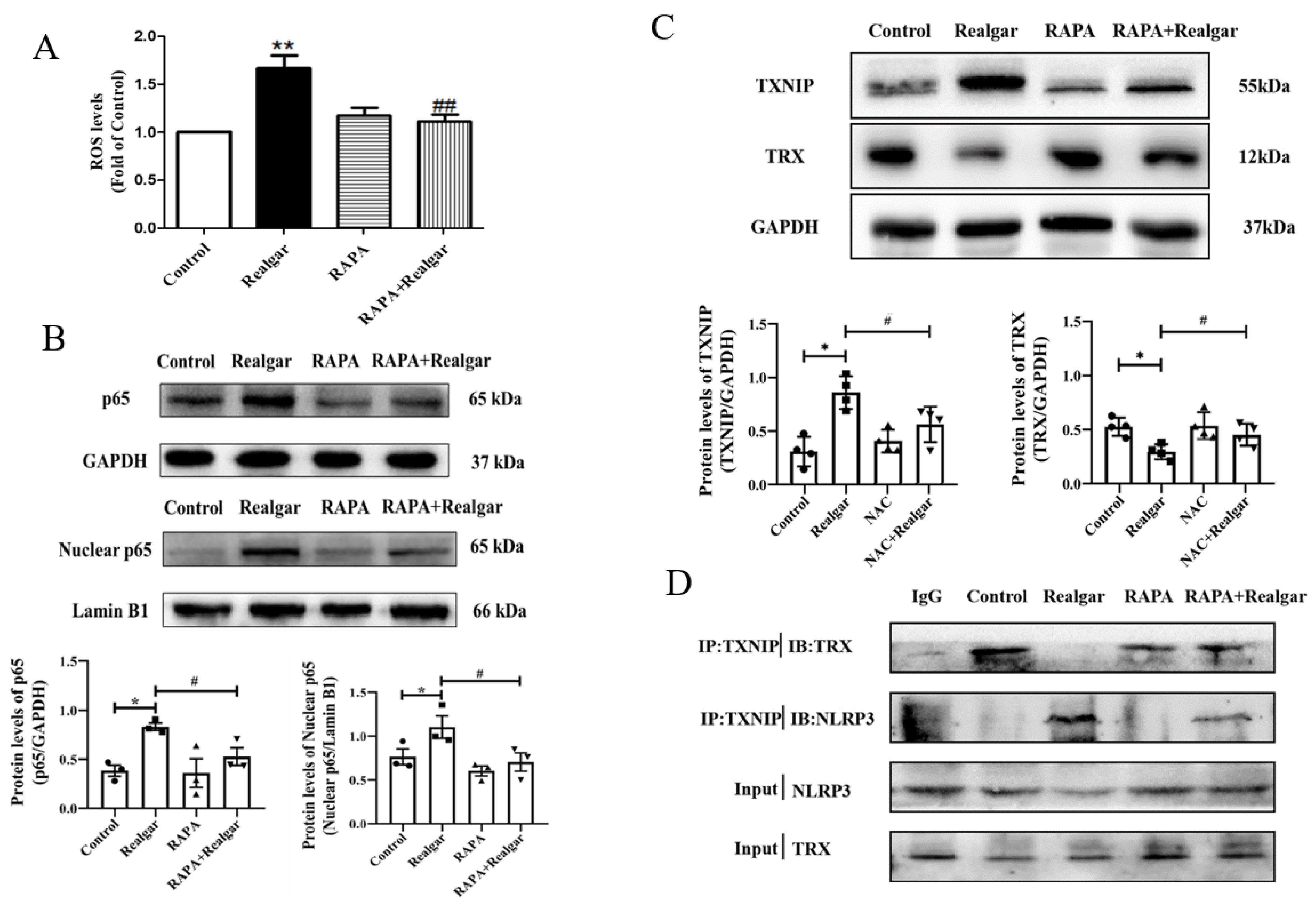
Autophagy may inhibit NLRP3 inflammasome activation by removing excess ROS. In this experiment, RAPA treatment apparently restored the increased ROS levels induced by realgar in mouse livers.
NF-κB plays an important role in the priming stage of NLRP3 inflammasome activation. The NF-κB enters into the nuclear and upregulates the expression of inflammasome component NLRP3 and pro-IL-1β. It has been reported that autophagy can directly regulate NF-κB to regulate inflammatory responses [
35]. Our previous study has shown that sub-chronic realgar exposure activated the NF-κB signaling pathway [
7]. In this study, we investigated the effect of autophagy on the NF-κB signaling pathway in realgar-induced liver injury mice. The results showed that RAPA treatment inhibits NF-κBp65 nuclear translocation and subsequently reduced the transcription of
Nlrp3,
Asc and
Il-1β. Therefore, we hypothesized that autophagy may down-regulate the realgar-induced NLRP3 inflammasome activation pathway by inhibiting NF-κB nuclear translocation, thereby alleviating realgar-induced inflammatory injury in mouse livers.
Growing evidence has shown that TXNIP is an active regulator of NLRP3 [
36]. It can directly interact with NLRP3 leading to the activation of NLRP3 inflammasome [
37]. Heo et al. demonstrated that hepatocytes treated with ethanol resulted in TXNIP overexpression, activating NLRP3 inflammasome [
38]. In a previous study, it was found that the gene and protein expression of TXNIP was increased in the liver tissues of mice exposed to 1.35 g/kg realgar for 8 weeks, and the interaction between NLRP3 and TXNIP was enhanced [
7]. The TXNIP/NLRP3 axis is regulated by ROS levels. Under high ROS concentrations, TXNIP dissociates from TRX, binds to NLRP3 protein, and activates the NLRP3 inflammasome. We found that activation of autophagy reduced TXNIP protein levels in the liver tissues of realgar-induced liver injury mice and weakened the interaction between TXNIP and NLRP3.
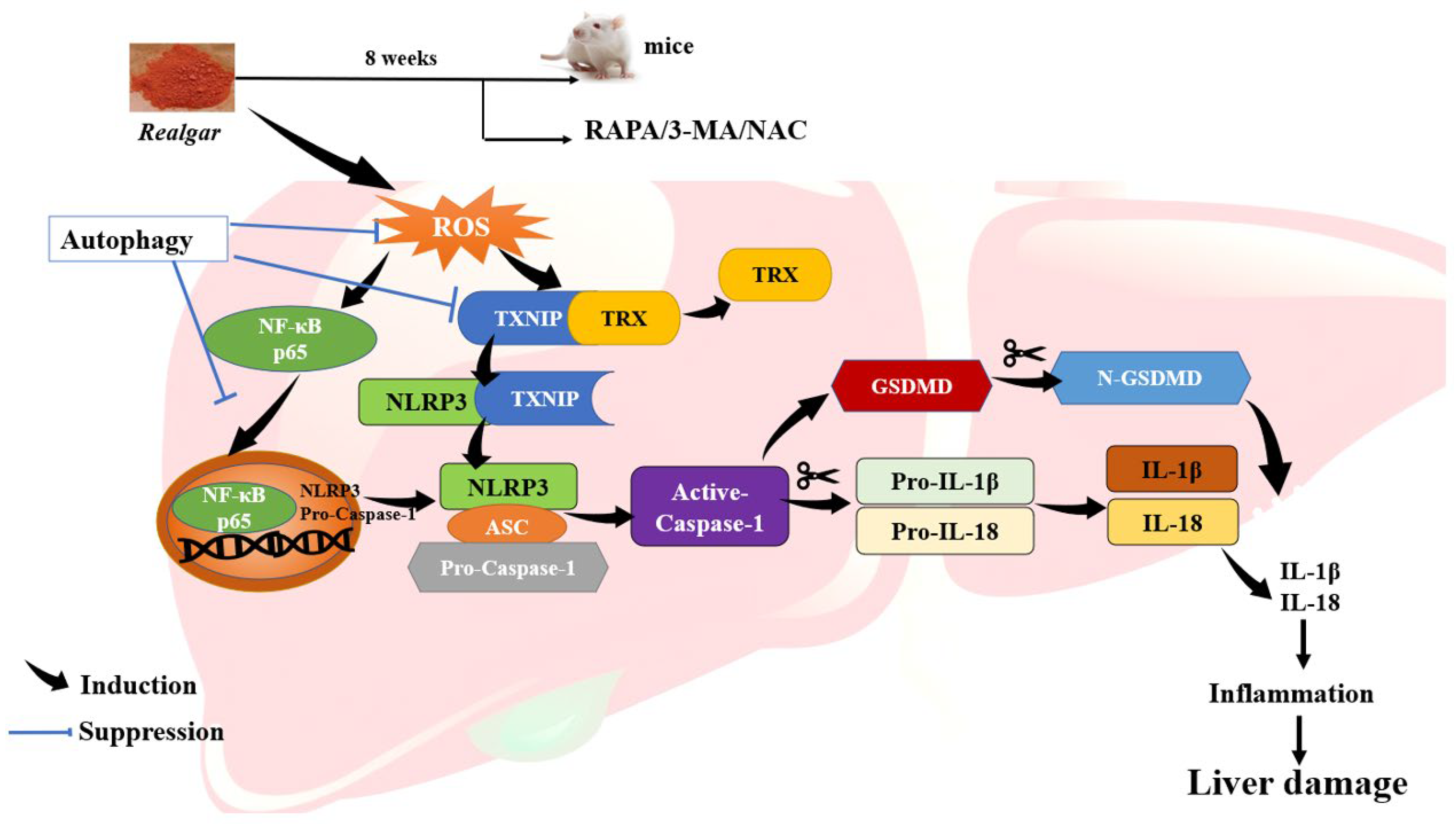
Taken together, this study suggests that autophagy may exert a protective effect on realgar-induced liver injury by eliminating excessive ROS, inhibiting NF-κB nuclear translocation and down-regulating TXNIP/NLRP3 axis, consequently inhibiting NLRP3 inflammasome activation in mouse livers.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}