
ALCAM/CD166 Is Involved in the Binding and Uptake of Cancer-Derived Extracellular Vesicles

 , , , , , and
, , , , , and 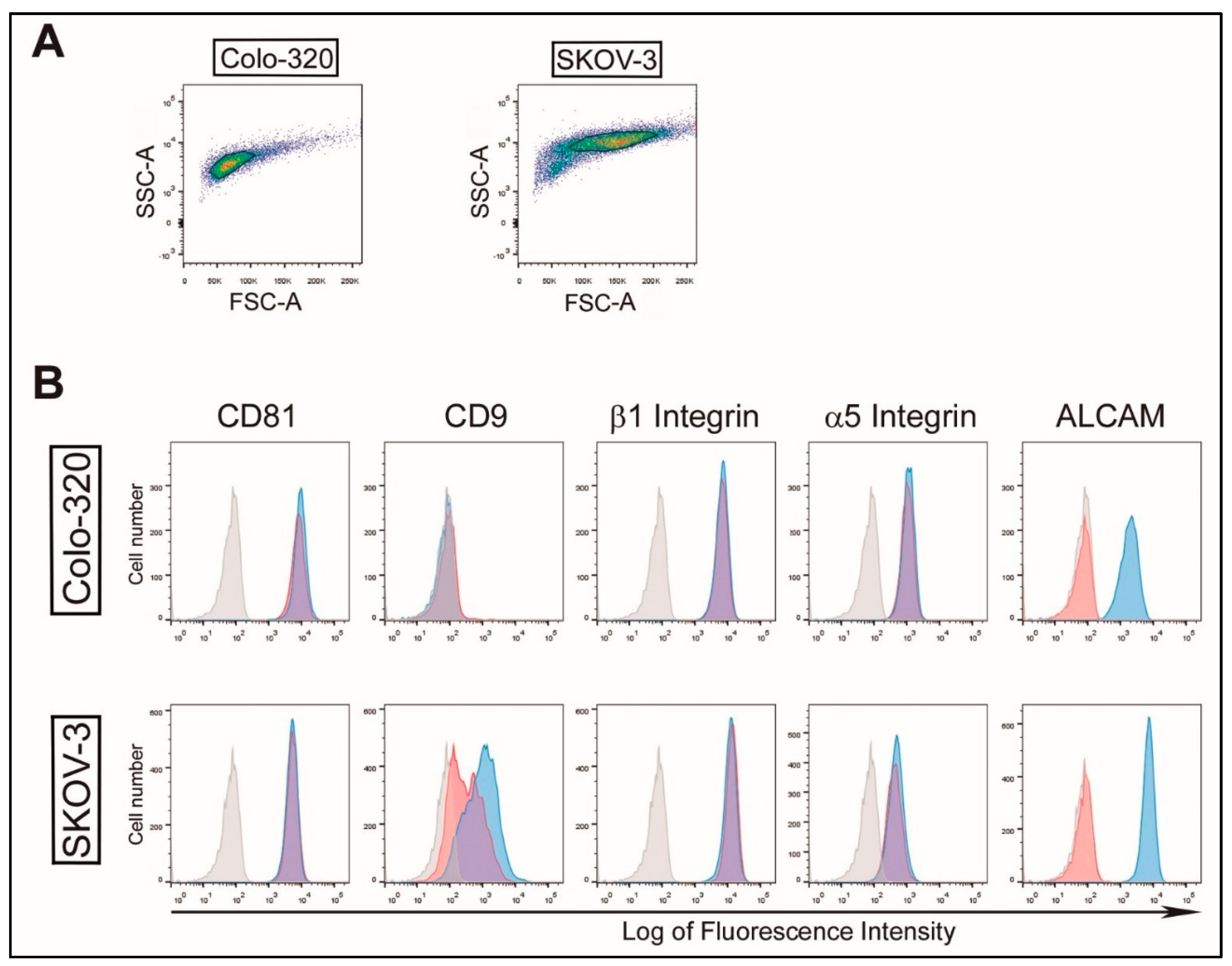{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
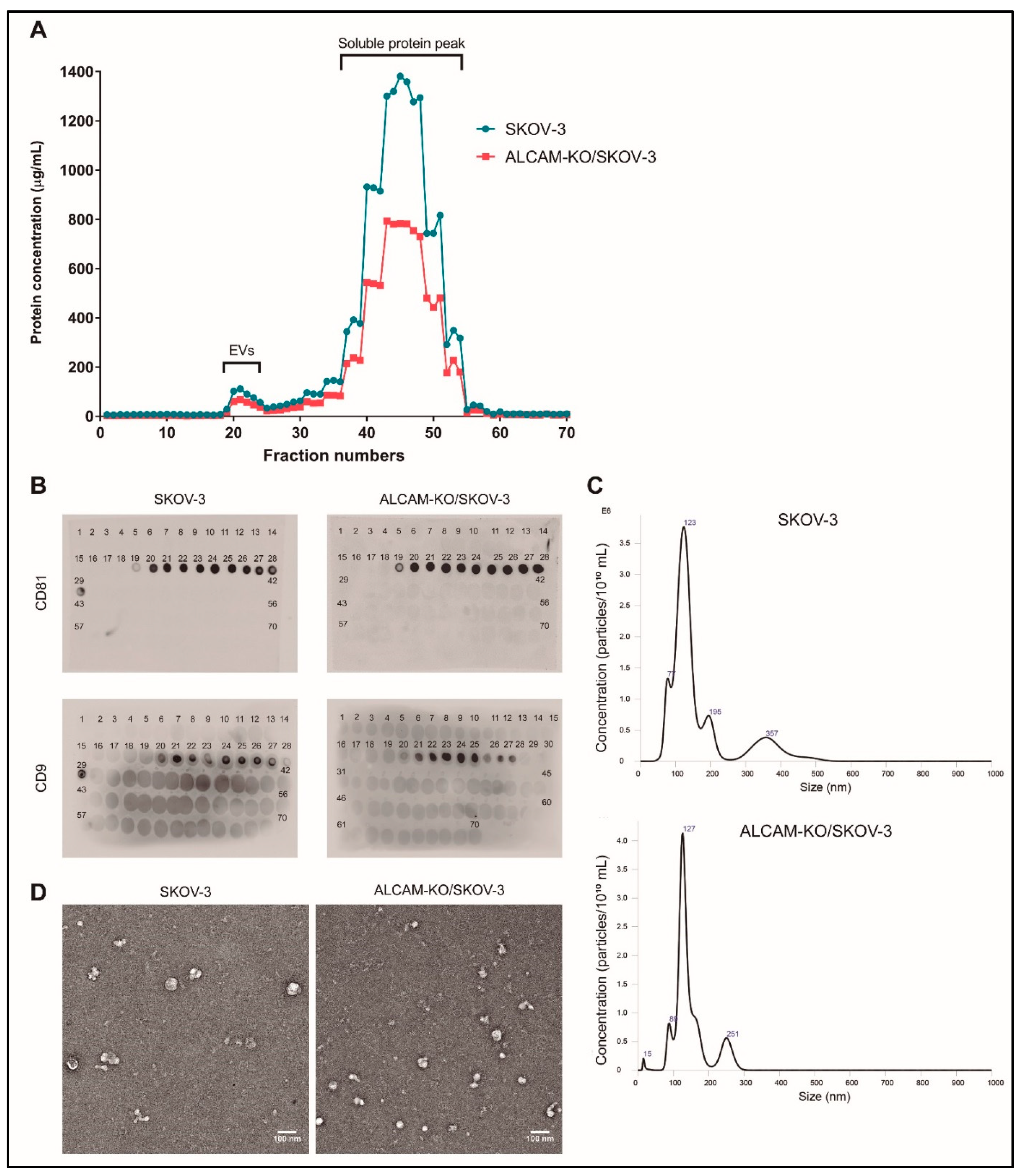
2.1. Characterization of EVs Derived from CRC Colo-320 and OvC SKOV-3 Cells
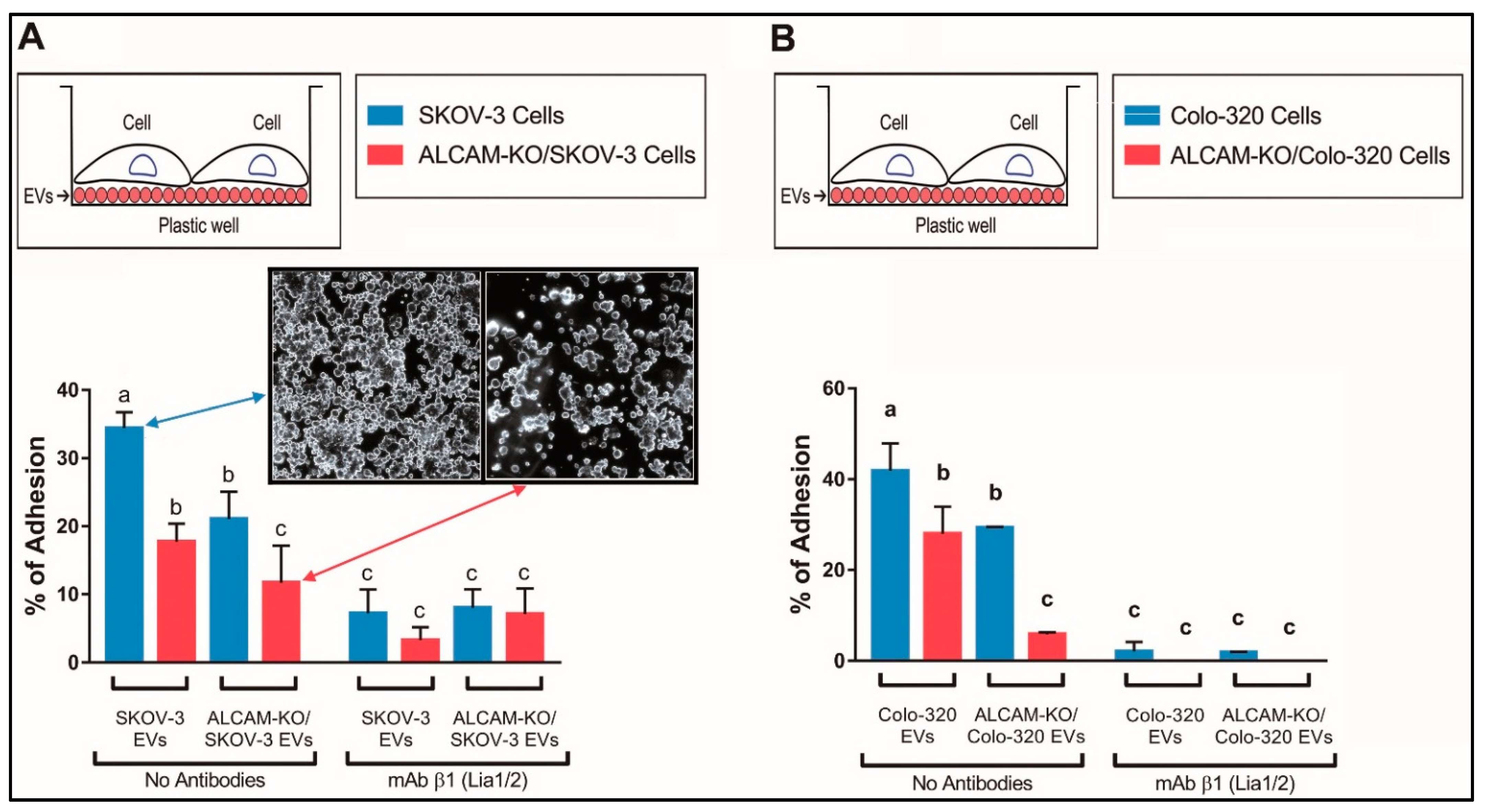
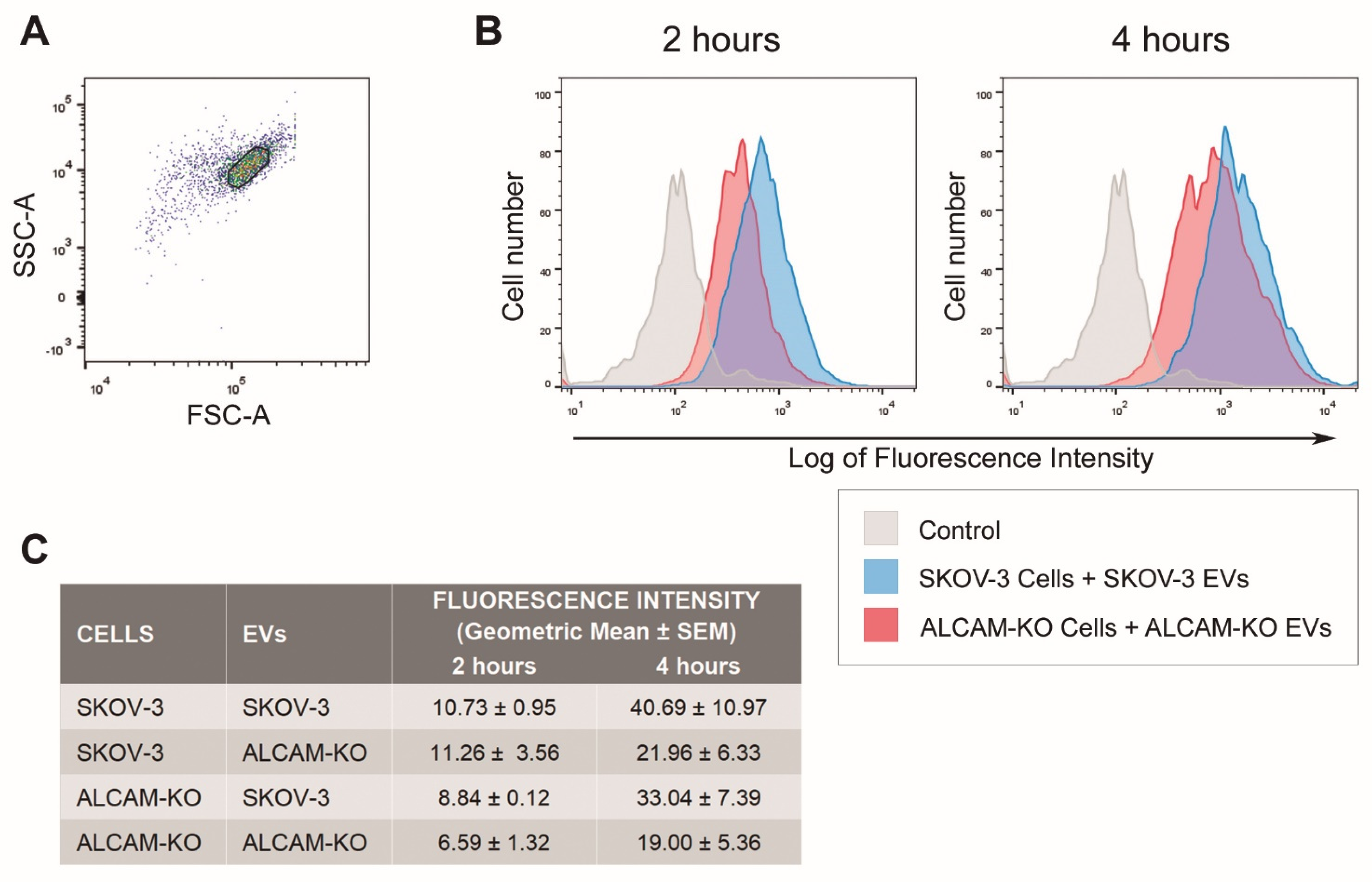
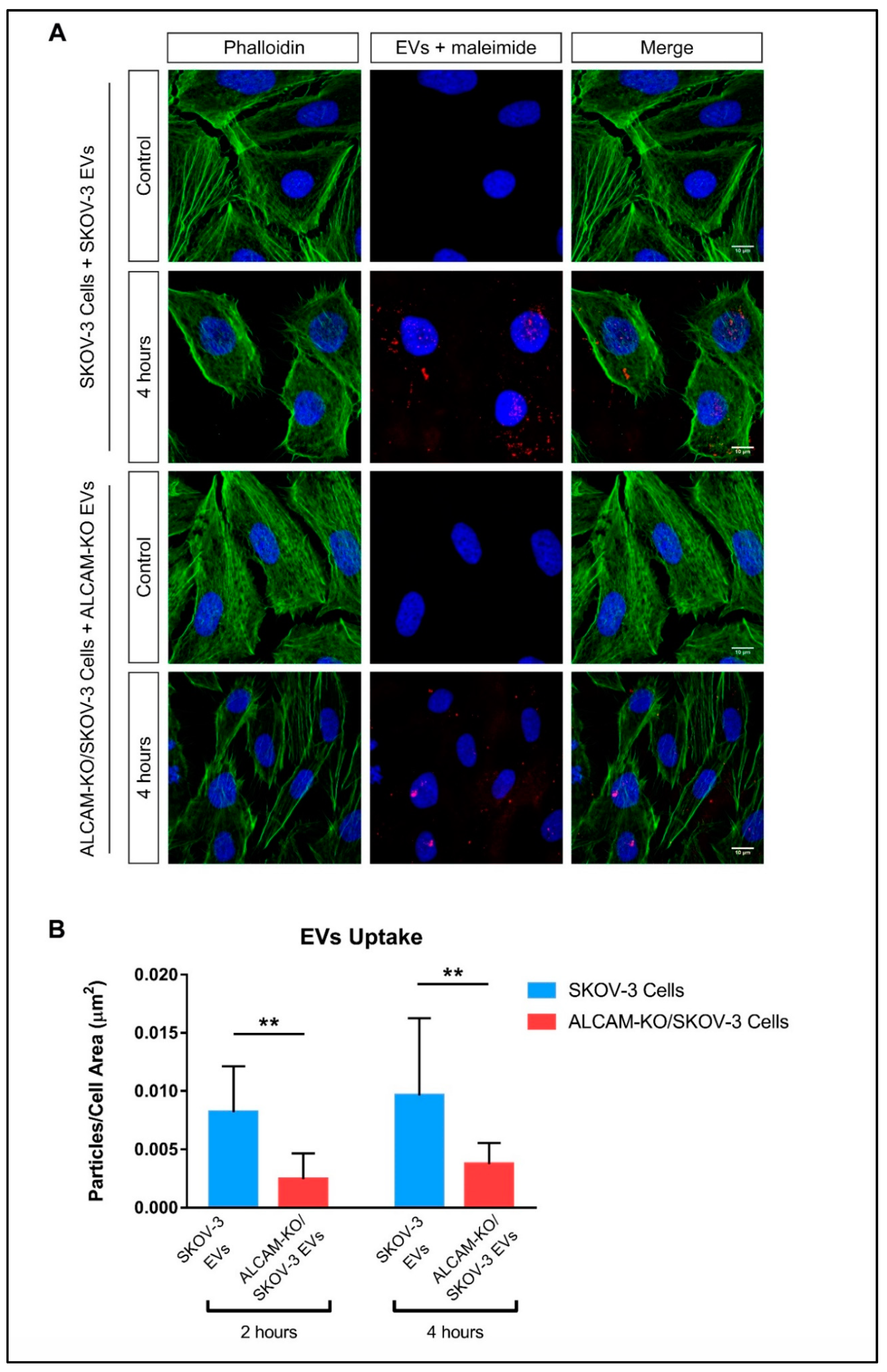
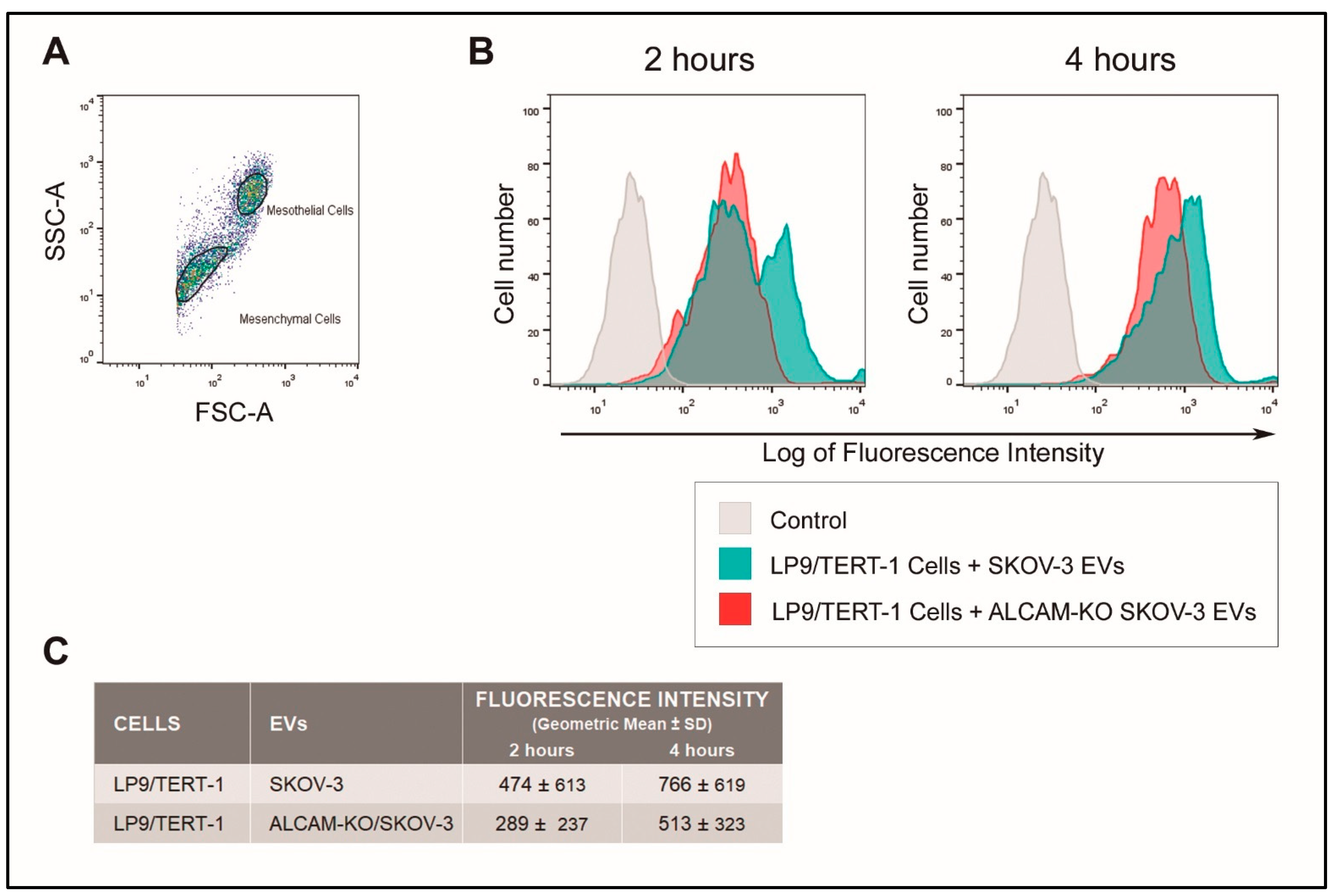
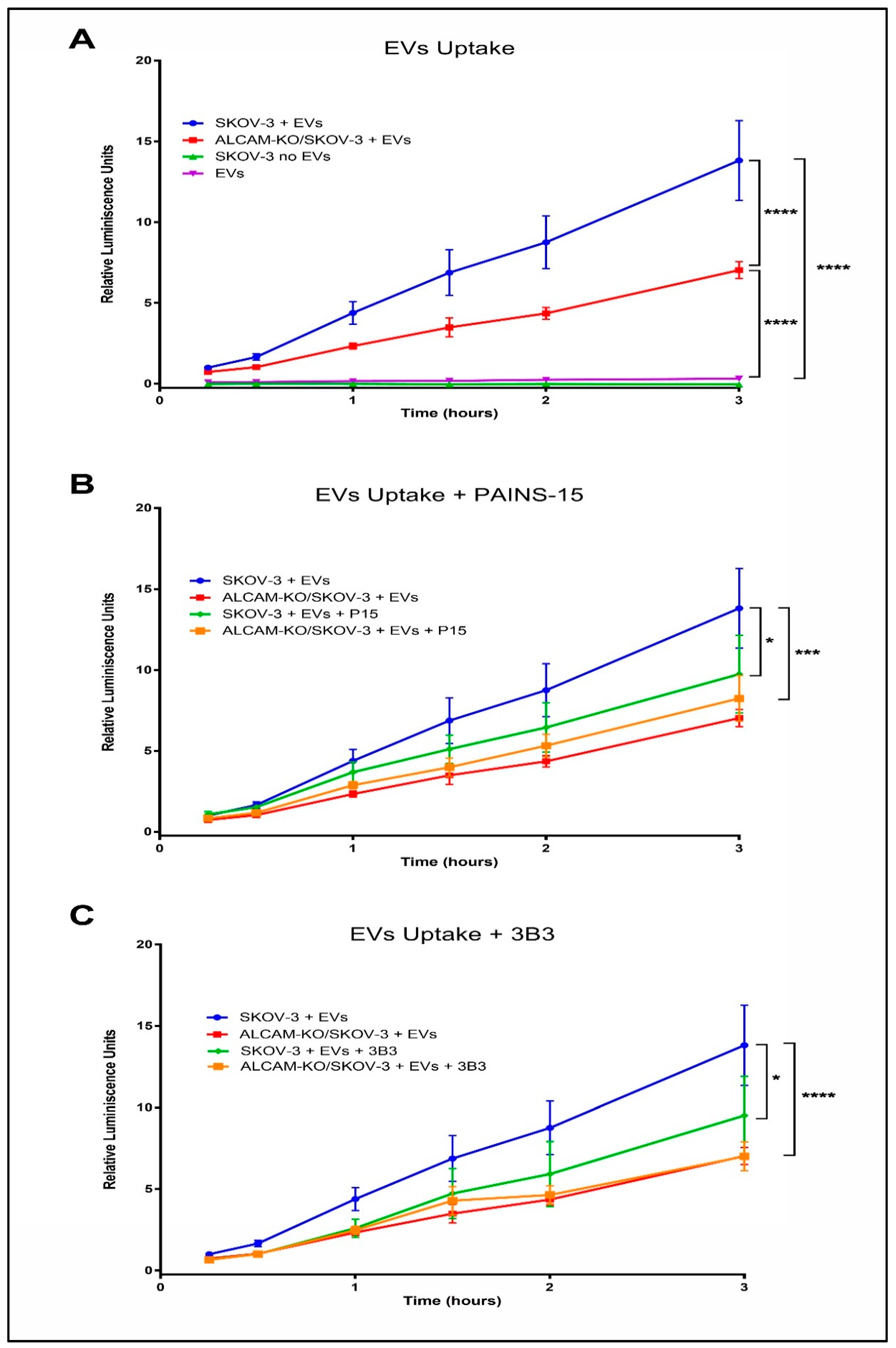
2.2. ALCAM/CD166 Mediates Interactions of Tumor-Derived EVs with Recipient Cancer Cells
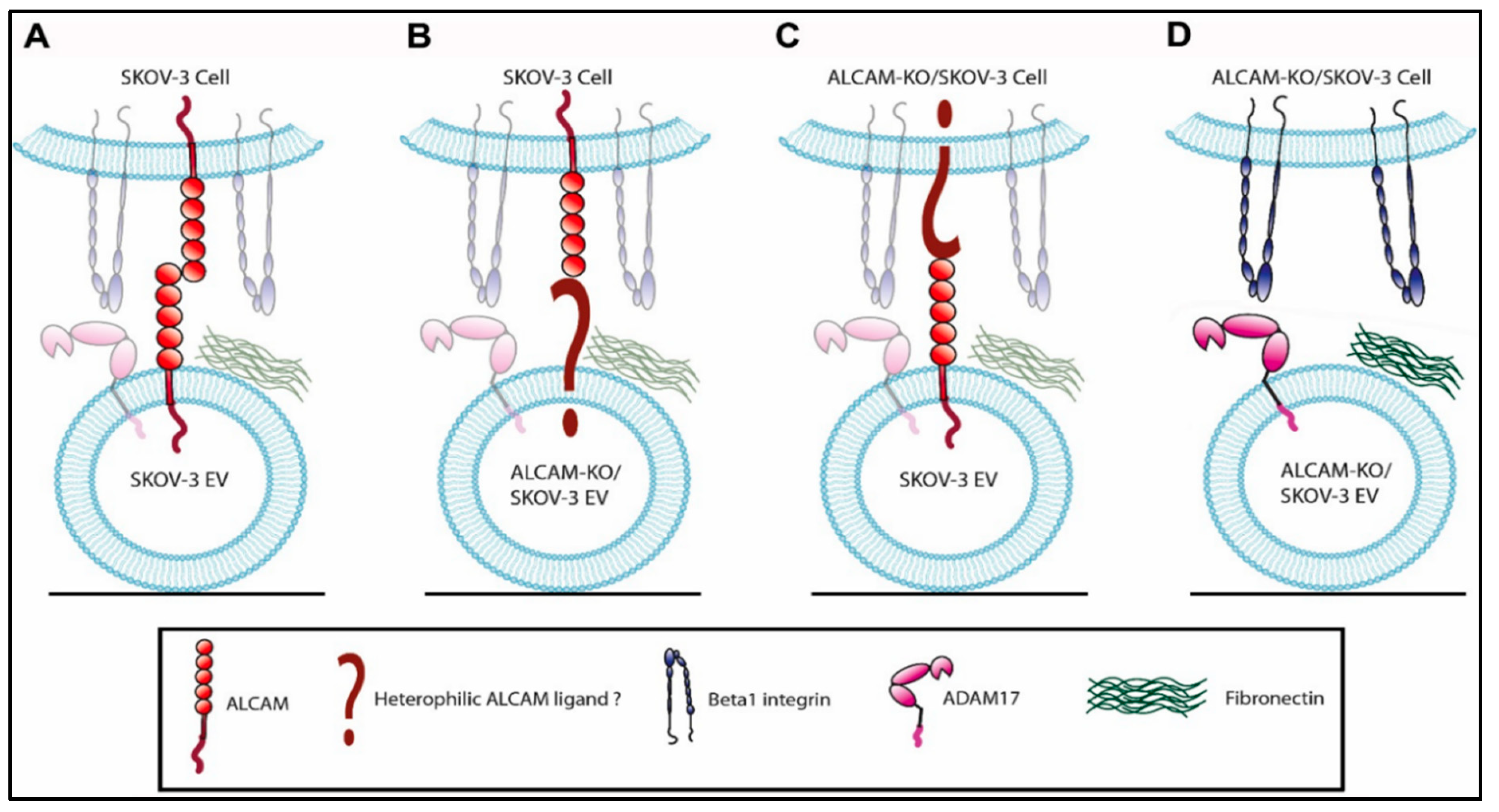
3. Discussion
4. Materials and Methods
4.1. Cells and Antibodies
4.2. Flow Cytometry
4.3. EVs Purification
4.4. Dot Blot
4.5. Nano Tracking Analysis (NTA)
4.6. Transmission Electron Microscopy
4.7. Cell Adhesion Assays
4.8. Measurement of Cellular EV Uptake
4.9. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Bedina Zavec, A.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological Properties of Extracellular Vesicles and Their Physiological Functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [Green Version]
- Kowal, J.; Tkach, M.; Théry, C. Biogenesis and Secretion of Exosomes. Curr. Opin. Cell Biol. 2014, 29, 116–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Thery, C. Specificities of Secretion and Uptake of Exosomes and Other Extracellular Vesicles for Cell-to-Cell Communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Becker, A.; Thakur, B.K.; Weiss, J.M.; Kim, H.S.; Peinado, H.; Lyden, D. Extracellular Vesicles in Cancer: Cell-to-Cell Mediators of Metastasis. Cancer Cell 2016, 30, 836–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Chengalvala, V.; Hu, H.; Sun, D. Tumor-Derived Exosomes: Nanovesicles Made by Cancer Cells to Promote Cancer Metastasis. Acta Pharm. Sin. B 2021, 11, 2136–2149. [Google Scholar] [CrossRef]
- Willms, E.; Cabañas, C.; Mäger, I.; Wood, M.J.A.; Vader, P. Extracellular Vesicle Heterogeneity: Subpopulations, Isolation Techniques, and Diverse Functions in Cancer Progression. Front. Immunol. 2018, 9, 738. [Google Scholar] [CrossRef] [Green Version]
- Kranenburg, O.; van der Speeten, K.; de Hingh, I. Peritoneal Metastases From Colorectal Cancer: Defining and Addressing the Challenges. Front. Oncol. 2021, 11, 650098. [Google Scholar] [CrossRef]
- Lengyel, E. Ovarian Cancer Development and Metastasis. Am. J. Pathol. 2010, 177, 1053–1064. [Google Scholar] [CrossRef]
- Pretzsch, E.; Bosch, F.; Neumann, J.; Ganschow, P.; Bazhin, A.; Guba, M.; Werner, J.; Angele, M. Mechanisms of Metastasis in Colorectal Cancer and Metastatic Organotropism: Hematogenous versus Peritoneal Spread. J. Oncol. 2019, 2019, 7407190. [Google Scholar] [CrossRef]
- Kipps, E.; Tan, D.S.P.; Kaye, S.B. Meeting the Challenge of Ascites in Ovarian Cancer: New Avenues for Therapy and Research. Nat. Rev. Cancer 2013, 13, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Sangisetty, S.L.; Miner, T.J. Malignant Ascites: A Review of Prognostic Factors, Pathophysiology and Therapeutic Measures. World J. Gastrointest. Surg. 2012, 4, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Dean, D.C.; Hornicek, F.J.; Shi, H.; Duan, Z. Exosomes Promote Pre-Metastatic Niche Formation in Ovarian Cancer. Mol. Cancer 2019, 18, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, K.; Sawada, K.; Kobayashi, M.; Miyamoto, M.; Shimizu, A.; Yamamoto, M.; Kinose, Y.; Kimura, T. Role of the Exosome in Ovarian Cancer Progression and Its Potential as a Therapeutic Target. Cancers 2019, 11, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.; Zhu, X.; Fei, J.; Shi, P.; Yu, S.; Zhou, J. Advances of Exosome in the Development of Ovarian Cancer and Its Diagnostic and Therapeutic Prospect. OncoTargets Ther. 2018, 11, 2831–2841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascual-Antón, L.; Cardeñes, B.; Sainz de la Cuesta, R.; González-Cortijo, L.; López-Cabrera, M.; Cabañas, C.; Sandoval, P. Mesothelial-to-Mesenchymal Transition and Exosomes in Peritoneal Metastasis of Ovarian Cancer. Int. J. Mol. Sci. 2021, 22, 11496. [Google Scholar] [CrossRef]
- Tian, W.; Lei, N.; Zhou, J.; Chen, M.; Guo, R.; Qin, B.; Li, Y.; Chang, L. Extracellular Vesicles in Ovarian Cancer Chemoresistance, Metastasis, and Immune Evasion. Cell Death Dis. 2022, 13, 64. [Google Scholar] [CrossRef]
- Alharbi, M.; Zuñiga, F.; Elfeky, O.; Guanzon, D.; Lai, A.; Rice, G.E.; Perrin, L.; Hooper, J.; Salomon, C. The Potential Role of MiRNAs and Exosomes in Chemotherapy in Ovarian Cancer. Endocr.-Relat. Cancer 2018, 25, R663–R685. [Google Scholar] [CrossRef] [Green Version]
- Schillaci, O.; Fontana, S.; Monteleone, F.; Taverna, S.; Di Bella, M.A.; Di Vizio, D.; Alessandro, R. Exosomes from Metastatic Cancer Cells Transfer Amoeboid Phenotype to Non-Metastatic Cells and Increase Endothelial Permeability: Their Emerging Role in Tumor Heterogeneity. Sci. Rep. 2017, 7, 4711. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Wang, C.; Zhu, H.; Wang, Y.; Wang, X.; Cheng, X.; Ge, W.; Lu, W. Exosome-Mediated Transfer of CD44 from High-Metastatic Ovarian Cancer Cells Promotes Migration and Invasion of Low-Metastatic Ovarian Cancer Cells. J. Ovarian Res. 2021, 14, 38. [Google Scholar] [CrossRef]
- Consuegra-Fernandez, M.; Lin, F.; Fox, D.A.; Lozano, F. Clinical and Experimental Evidence for Targeting CD6 in Immune-Based Disorders. Autoimmun. Rev. 2018, 17, 493–503. [Google Scholar] [CrossRef]
- Hassan, N.J.; Barclay, A.N.; Brown, M.H. Frontline: Optimal T Cell Activation Requires the Engagement of CD6 and CD166. Eur. J. Immunol. 2004, 34, 930–940. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, A.W.; Joosten, B.; Torensma, R.; Parnes, J.R.; van Leeuwen, F.N.; Figdor, C.G. Long-Term Engagement of CD6 and ALCAM Is Essential for T-Cell Proliferation Induced by Dendritic Cells. Blood 2006, 107, 3212–3220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weidle, U.H.; Eggle, D.; Klostermann, S.; Swart, G.W. ALCAM/CD166: Cancer-Related Issues. Cancer Genom. Proteom. 2010, 7, 231–243. [Google Scholar]
- Cayrol, R.; Wosik, K.; Berard, J.L.; Dodelet-Devillers, A.; Ifergan, I.; Kebir, H.; Haqqani, A.S.; Kreymborg, K.; Krug, S.; Moumdjian, R.; et al. Activated Leukocyte Cell Adhesion Molecule Promotes Leukocyte Trafficking into the Central Nervous System. Nat. Immunol. 2008, 9, 137–145. [Google Scholar] [CrossRef]
- Masedunskas, A.; King, J.A.; Tan, F.; Cochran, R.; Stevens, T.; Sviridov, D.; Ofori-Acquah, S.F. Activated Leukocyte Cell Adhesion Molecule Is a Component of the Endothelial Junction Involved in Transendothelial Monocyte Migration. FEBS Lett. 2006, 580, 2637–2645. [Google Scholar] [CrossRef] [Green Version]
- Swart, G.W.; Lunter, P.C.; Kilsdonk, J.W.; Kempen, L.C. Activated Leukocyte Cell Adhesion Molecule (ALCAM/CD166): Signaling at the Divide of Melanoma Cell Clustering and Cell Migration? Cancer Metastasis Rev. 2005, 24, 223–236. [Google Scholar] [CrossRef]
- Darvishi, B.; Boroumandieh, S.; Majidzadeh, A.K.; Salehi, M.; Jafari, F.; Farahmand, L. The Role of Activated Leukocyte Cell Adhesion Molecule (ALCAM) in Cancer Progression, Invasion, Metastasis and Recurrence: A Novel Cancer Stem Cell Marker and Tumor-Specific Prognostic Marker. Exp. Mol. Pathol. 2020, 115, 104443. [Google Scholar] [CrossRef]
- Ferragut, F.; Vachetta, V.S.; Troncoso, M.F.; Rabinovich, G.A.; Elola, M.T. ALCAM/CD166: A Pleiotropic Mediator of Cell Adhesion, Stemness and Cancer Progression. Cytokine Growth Factor Rev. 2021, 61, 27–37. [Google Scholar] [CrossRef]
- Ofori-Acquah, S.F.; King, J.A. Activated Leukocyte Cell Adhesion Molecule: A New Paradox in Cancer. Transl. Res. 2008, 151, 122–128. [Google Scholar] [CrossRef]
- von Lersner, A.; Droesen, L.; Zijlstra, A. Modulation of Cell Adhesion and Migration through Regulation of the Immunoglobulin Superfamily Member ALCAM/CD166. Clin. Exp. Metastasis 2019, 36, 87–95. [Google Scholar] [CrossRef]
- Bech-Serra, J.J.; Santiago-Josefat, B.; Esselens, C.; Saftig, P.; Baselga, J.; Arribas, J.; Canals, F. Proteomic Identification of Desmoglein 2 and Activated Leukocyte Cell Adhesion Molecule as Substrates of ADAM17 and ADAM10 by Difference Gel Electrophoresis. Mol. Cell. Biol. 2006, 26, 5086–5095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilsanz, A.; Sánchez-Martín, L.; Gutiérrez-López, M.D.; Ovalle, S.; Machado-Pineda, Y.; Reyes, R.; Swart, G.W.; Figdor, C.G.; Lafuente, E.M.; Cabañas, C. ALCAM/CD166 Adhesive Function Is Regulated by the Tetraspanin CD9. Cell. Mol. Life Sci. 2013, 70, 475–493. [Google Scholar] [CrossRef] [PubMed]
- Rosso, O.; Piazza, T.; Bongarzone, I.; Rossello, A.; Mezzanzanica, D.; Canevari, S.; Orengo, A.M.; Puppo, A.; Ferrini, S.; Fabbi, M. The ALCAM Shedding by the Metalloprotease ADAM17/TACE Is Involved in Motility of Ovarian Carcinoma Cells. Mol. Cancer Res. MCR 2007, 5, 1246–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, A.G.; Freeman, T.J.; Arnold, S.A.; Starchenko, A.; Jones-Paris, C.R.; Gilger, M.A.; Washington, M.K.; Fan, K.-H.; Shyr, Y.; Beauchamp, R.D.; et al. Elevated ALCAM Shedding in Colorectal Cancer Correlates with Poor Patient Outcome. Cancer Res. 2013, 73, 2955–2964. [Google Scholar] [CrossRef] [Green Version]
- Carbotti, G.; Orengo, A.M.; Mezzanzanica, D.; Bagnoli, M.; Brizzolara, A.; Emionite, L.; Puppo, A.; Centurioni, M.G.; Bruzzone, M.; Marroni, P.; et al. Activated Leukocyte Cell Adhesion Molecule Soluble Form: A Potential Biomarker of Epithelial Ovarian Cancer Is Increased in Type II Tumors. Int. J. Cancer 2013, 132, 2597–2605. [Google Scholar] [CrossRef]
- Cardenes, B.; Clares, I.; Toribio, V.; Pascual, L.; Lopez-Martin, S.; Torres-Gomez, A.; Sainz de la Cuesta, R.; Lafuente, E.M.; Lopez-Cabrera, M.; Yanez-Mo, M.; et al. Cellular Integrin Alpha5beta1 and Exosomal ADAM17 Mediate the Binding and Uptake of Exosomes Produced by Colorectal Carcinoma Cells. Int. J. Mol. Sci. 2021, 22, 9938. [Google Scholar] [CrossRef]
- Machado-Pineda, Y.; Cardeñes, B.; Reyes, R.; López-Martín, S.; Toribio, V.; Sánchez-Organero, P.; Suarez, H.; Grötzinger, J.; Lorenzen, I.; Yáñez-Mó, M.; et al. CD9 Controls Integrin A5β1-Mediated Cell Adhesion by Modulating Its Association With the Metalloproteinase ADAM17. Front. Immunol. 2018, 9, 2474. [Google Scholar] [CrossRef] [Green Version]
- Ovalle, S.; Gutierrez-Lopez, M.D.; Olmo, N.; Turnay, J.; Lizarbe, M.A.; Majano, P.; Molina-Jimenez, F.; Lopez-Cabrera, M.; Yanez-Mo, M.; Sanchez-Madrid, F.; et al. The Tetraspanin CD9 Inhibits the Proliferation and Tumorigenicity of Human Colon Carcinoma Cells. Int. J. Cancer 2007, 121, 2140–2152. [Google Scholar] [CrossRef]
- Andreu, Z.; Yanez-Mo, M. Tetraspanins in Extracellular Vesicle Formation and Function. Front. Immunol. 2014, 5, 442. [Google Scholar] [CrossRef] [Green Version]
- Kowal, J.; Arras, G.; Colombo, M.; Jouve, M.; Morath, J.P.; Primdal-Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Théry, C. Proteomic Comparison Defines Novel Markers to Characterize Heterogeneous Populations of Extracellular Vesicle Subtypes. Proc. Natl. Acad. Sci. USA 2016, 113, E968–E977. [Google Scholar] [CrossRef] [Green Version]
- Mulcahy, L.A.; Pink, R.C.; Carter, D.R. Routes and Mechanisms of Extracellular Vesicle Uptake. J. Extracell. Vesicles 2014, 3, 24641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- French, K.C.; Antonyak, M.A.; Cerione, R.A. Extracellular Vesicle Docking at the Cellular Port: Extracellular Vesicle Binding and Uptake. Semin. Cell Dev. Biol. 2017, 67, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Gurung, S.; Perocheau, D.; Touramanidou, L.; Baruteau, J. The Exosome Journey: From Biogenesis to Uptake and Intracellular Signalling. Cell Commun. Signal. CCS 2021, 19, 47. [Google Scholar] [CrossRef] [PubMed]
- Dickson, M.A.; Hahn, W.C.; Ino, Y.; Ronfard, V.; Wu, J.Y.; Weinberg, R.A.; Louis, D.N.; Li, F.P.; Rheinwald, J.G. Human Keratinocytes That Express HTERT and Also Bypass a P16(INK4a)-Enforced Mechanism That Limits Life Span Become Immortal yet Retain Normal Growth and Differentiation Characteristics. Mol. Cell. Biol. 2000, 20, 1436–1447. [Google Scholar] [CrossRef] [Green Version]
- Shukla, A.; MacPherson, M.B.; Hillegass, J.; Ramos-Nino, M.E.; Alexeeva, V.; Vacek, P.M.; Bond, J.P.; Pass, H.I.; Steele, C.; Mossman, B.T. Alterations in Gene Expression in Human Mesothelial Cells Correlate with Mineral Pathogenicity. Am. J. Respir. Cell Mol. Biol. 2009, 41, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Connell, N.D.; Rheinwald, J.G. Regulation of the Cytoskeleton in Mesothelial Cells: Reversible Loss of Keratin and Increase in Vimentin during Rapid Growth in Culture. Cell 1983, 34, 245–253. [Google Scholar] [CrossRef]
- Toribio, V.; Morales, S.; López-Martín, S.; Cardeñes, B.; Cabañas, C.; Yáñez-Mó, M. Development of a Quantitative Method to Measure EV Uptake. Sci. Rep. 2019, 9, 10522. [Google Scholar] [CrossRef]
- Nazarenko, I.; Rana, S.; Baumann, A.; McAlear, J.; Hellwig, A.; Trendelenburg, M.; Lochnit, G.; Preissner, K.T.; Zöller, M. Cell Surface Tetraspanin Tspan8 Contributes to Molecular Pathways of Exosome-Induced Endothelial Cell Activation. Cancer Res. 2010, 70, 1668–1678. [Google Scholar] [CrossRef] [Green Version]
- Reyes, R.; Cardenes, B.; Machado-Pineda, Y.; Cabanas, C. Tetraspanin CD9: A Key Regulator of Cell Adhesion in the Immune System. Front. Immunol. 2018, 9, 863. [Google Scholar] [CrossRef]
- Zech, D.; Rana, S.; Büchler, M.W.; Zöller, M. Tumor-Exosomes and Leukocyte Activation: An Ambivalent Crosstalk. Cell Commun. Signal. 2012, 10, 37. [Google Scholar] [CrossRef] [Green Version]
- Suárez, H.; Andreu, Z.; Mazzeo, C.; Toribio, V.; Pérez-Rivera, A.E.; López-Martín, S.; García-Silva, S.; Hurtado, B.; Morato, E.; Peláez, L.; et al. CD9 Inhibition Reveals a Functional Connection of Extracellular Vesicle Secretion with Mitophagy in Melanoma Cells. J. Extracell. Vesicles 2021, 10, e12082. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Lopez, M.D.; Ovalle, S.; Yanez-Mo, M.; Sanchez-Sanchez, N.; Rubinstein, E.; Olmo, N.; Lizarbe, M.A.; Sanchez-Madrid, F.; Cabanas, C. A Functionally Relevant Conformational Epitope on the CD9 Tetraspanin Depends on the Association with Activated Beta1 Integrin. J. Biol. Chem. 2003, 278, 208–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vences-Catalán, F.; Rajapaksa, R.; Kuo, C.-C.; Miller, C.L.; Lee, A.; Ramani, V.C.; Jeffrey, S.S.; Levy, R.; Levy, S. Targeting the Tetraspanin CD81 Reduces Cancer Invasion and Metastasis. Proc. Natl. Acad. Sci. USA 2021, 118, e2018961118. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, A.G.; Sanchez-Mateos, P.; Campanero, M.R.; Martin-Padura, I.; Dejana, E.; Sanchez-Madrid, F. Regulation of the VLA Integrin-Ligand Interactions through the Beta 1 Subunit. J. Cell Biol. 1992, 117, 659–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campanero, M.R.; Arroyo, A.G.; Pulido, R.; Ursa, A.; de Matías, M.S.; Sánchez-Mateos, P.; Kassner, P.D.; Chan, B.M.; Hemler, M.E.; Corbí, A.L.; et al. Functional Role of Alpha 2/Beta 1 and Alpha 4/Beta 1 Integrins in Leukocyte Intercellular Adhesion Induced through the Common Beta 1 Subunit. Eur. J. Immunol. 1992, 22, 3111–3119. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardeñes, B.; Clares, I.; Bezos, T.; Toribio, V.; López-Martín, S.; Rocha, A.; Peinado, H.; Yáñez-Mó, M.; Cabañas, C. ALCAM/CD166 Is Involved in the Binding and Uptake of Cancer-Derived Extracellular Vesicles. Int. J. Mol. Sci. 2022, 23, 5753. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105753
Cardeñes B, Clares I, Bezos T, Toribio V, López-Martín S, Rocha A, Peinado H, Yáñez-Mó M, Cabañas C. ALCAM/CD166 Is Involved in the Binding and Uptake of Cancer-Derived Extracellular Vesicles. International Journal of Molecular Sciences. 2022; 23(10):5753. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105753
Chicago/Turabian StyleCardeñes, Beatriz, Irene Clares, Tamara Bezos, Víctor Toribio, Soraya López-Martín, Almudena Rocha, Héctor Peinado, María Yáñez-Mó, and Carlos Cabañas. 2022. "ALCAM/CD166 Is Involved in the Binding and Uptake of Cancer-Derived Extracellular Vesicles" International Journal of Molecular Sciences 23, no. 10: 5753. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105753