
Extracellular Vesicles Released from Stem Cells as a New Therapeutic Strategy for Primary and Secondary Glomerulonephritis


Abstract
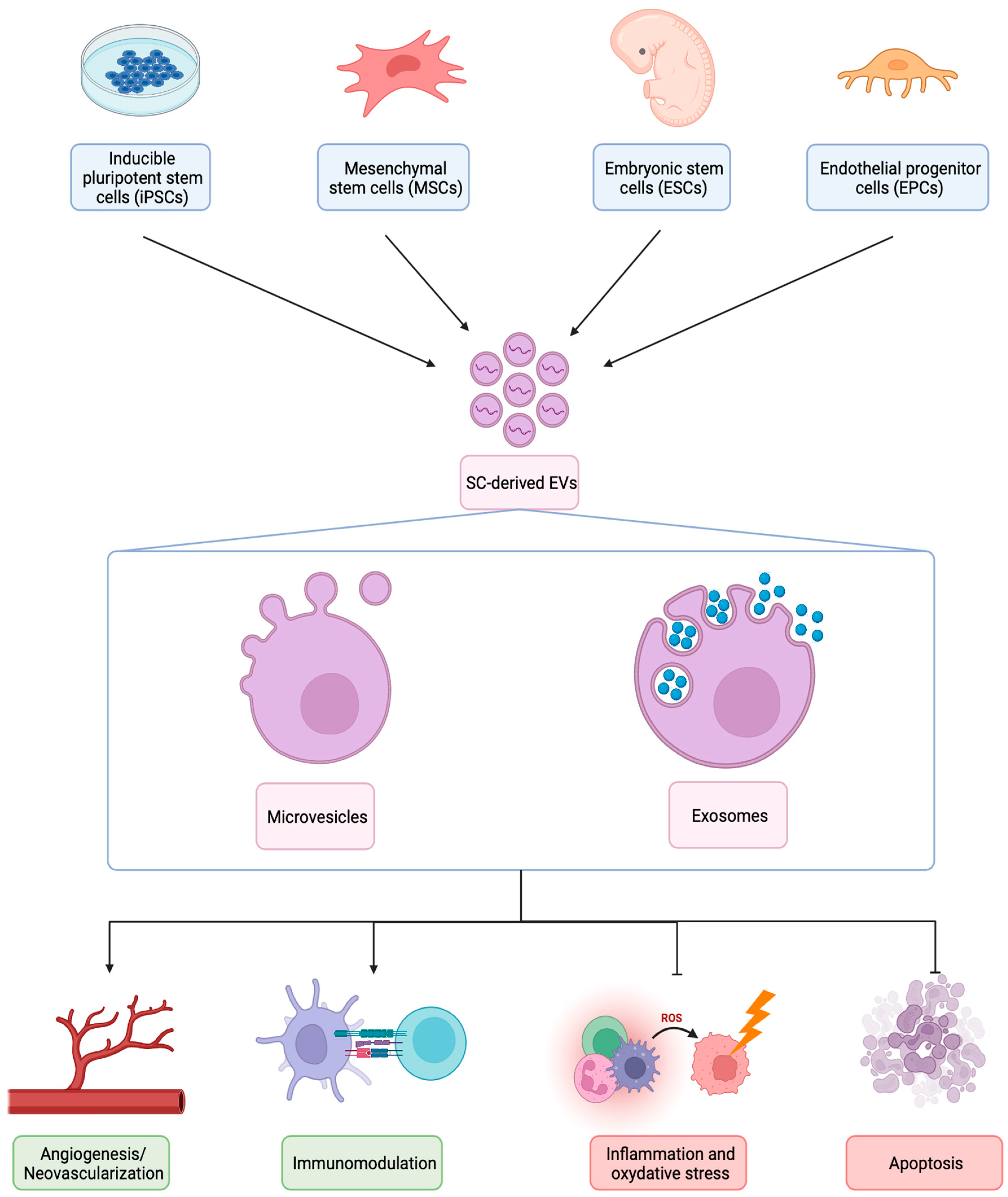
:1. Introduction
2. General Features of Stem Cells and Extracellular Vesicles Released from Stem Cells
3. Therapeutic Actions of Stem Cells and Extracellular Vesicles in Renal Disease
3.1. Cytoprotective Effects
3.2. Regenerative Effects
3.3. Immunomodulatory Effects
4. Stem Cells and Extracellular Vesicles as Therapeutic Tools for Glomerulonephritis
4.1. Primary Glomerulonephritis
4.1.1. Mesangial Proliferative Glomerulonephritis and IgA Nephropathy
4.1.2. Focal Segmental Glomerulosclerosis
4.1.3. Idiopathic Membranous Nephropathy
4.2. Secondary Glomerulonephritis
4.2.1. Lupus Nephritis
Mesenchymal Stromal Cell-Based Therapy for Lupus Nephritis
Therapy with Mesenchymal Stromal Cell-Derived Extracellular Vesicles for Lupus Nephritis
4.2.2. Anti-Glomerular Basement Membrane and ANCA-Associated Crescentic Glomerulonephritis
4.2.3. Diabetic Nephropathy
Mesenchymal Stromal Cells-Based Therapy for Diabetic Nephropathy
Endothelial Progenitor Cell-Based Therapy for Diabetic Nephropathy
Induced Pluripotent Stem Cell-Based Therapy for Diabetic Nephropathy
Urine-Derived Stem Cell-Based Therapy for Diabetic Nephropathy
5. Limitations and Perspectives
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AAV | ANCA-associated vasculitis |
| AD-MSC | Adipose tissue-derived Mesenchymal Stromal Cell |
| ANA | anti-nuclear antibodies |
| αSMA | α Smooth Muscle Actin |
| BM | Bone Marrow |
| BM-MSC | Bone Marrow-derived Mesenchymal Stromal Cell |
| BMP | Bone Morphogenetic Protein |
| CKD | chronic kidney disease |
| DC | Dendritic cells |
| DN | diabetic nephropathy |
| DM | diabetes mellitus |
| EGF | Epidermal Growth Factor |
| EMT | epithelial-to-mesenchymal transition |
| EPC | Endothelial Progenitor Cell |
| ESRD | end-stage renal disease |
| EV | Extracellular Vesicle |
| FSGS | focal segmental glomerulosclerosis |
| F/U | Follow-up |
| GBM | glomerular basement membrane |
| GFR | Glomerular Filtration Rate |
| GN | Glomerulonephritis |
| HGF | Hepatocyte Growth Factor |
| HSC | Hematopoietic Stem Cell |
| IgAN | IgAN nephropathy |
| IMN | Idiopathic Membranous Nephropathy |
| iPSC | induced pluripotent stem cells |
| LN | Lupus Nephritis |
| LPS | Lipopolysaccharide |
| MCP-1 | Monocyte Chemotactic Protein-1 |
| miRNA | microRNA |
| MSC | Mesenchymal Stromal Cell |
| NF-kB | Nuclear Factor kappa B |
| PBMC | Peripheral Blood Mononuclear Cell |
| pMSC | placental-derived Mesenchymal Stem Cell |
| RCT | Randomized Controlled Trial |
| RPC | Renal Progenitor Cell |
| RTEC | renal tubular epithelial cell |
| SC | Stem Cell |
| SLE | Systemic Lupus Erythematosus |
| SLEDAI | Systemic Lupus Erythematosus Disease Activity Index |
| STZ | Streptozocin |
| TNF | Tumor Necrosis Factor |
| Th | T-helper cells |
| Treg | T-regulatory cells |
| UC | Umbilical Cord |
| UC-MSCs | Umbilical Cord-derived Mesenchymal Stromal Cell |
| UCB-MSC | umbilical cord blood-derived Mesenchymal Stromal Cell |
| uEV | urinary Extracellular Vesicle |
| USC | Urine-derived Stem Cell |
References
- Kazi, A.M.; Hashmi, M.F. Glomerulonephritis. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Sethi, S.; Fervenza, F.C. Standardized Classification and Reporting of Glomerulonephritis. Nephrol. Dial. Transplant. 2019, 34, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Thomas, B. The Global Burden of Diabetic Kidney Disease: Time Trends and Gender Gaps. Curr. Diab. Rep. 2019, 19, 18. [Google Scholar] [CrossRef] [PubMed]
- Li, P.K.-T.; Ma, T.K.-W. Global Impact of Nephropathies. Nephrology 2017, 22 (Suppl. S4), 9–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Floege, J.; Amann, K. Primary Glomerulonephritides. Lancet 2016, 387, 2036–2048. [Google Scholar] [CrossRef]
- Couser, W.G. Pathogenesis and Treatment of Glomerulonephritis-an Update. J. Bras. Nefrol. 2016, 38, 107–122. [Google Scholar] [CrossRef]
- Sung, S.-S.J.; Fu, S.M. Interactions among Glomerulus Infiltrating Macrophages and Intrinsic Cells via Cytokines in Chronic Lupus Glomerulonephritis. J. Autoimmun. 2020, 106, 102331. [Google Scholar] [CrossRef]
- Holdsworth, S.R.; Gan, P.-Y. Cytokines: Names and Numbers You Should Care About. Clin. J. Am. Soc. Nephrol. 2015, 10, 2243–2254. [Google Scholar] [CrossRef]
- Anton-Pampols, P.; Diaz-Requena, C.; Martinez-Valenzuela, L.; Gomez-Preciado, F.; Fulladosa, X.; Vidal-Alabro, A.; Torras, J.; Lloberas, N.; Draibe, J. The Role of Inflammasomes in Glomerulonephritis. Int. J. Mol. Sci. 2022, 23, 4208. [Google Scholar] [CrossRef]
- Chen, P.; Chen, Y.; Jiang, M.; Mo, Y.; Ying, H.; Tang, X.; Zhang, J. Usefulness of the Cytokines Expression of Th1/Th2/Th17 and Urinary CD80 Excretion in Adult-Onset Minimal Change Disease. PeerJ 2020, 8, e9854. [Google Scholar] [CrossRef]
- Ahn, W.; Bomback, A.S. Approach to Diagnosis and Management of Primary Glomerular Diseases Due to Podocytopathies in Adults: Core Curriculum 2020. Am. J. Kidney Dis. 2020, 75, 955–964. [Google Scholar] [CrossRef]
- Kaartinen, K.; Safa, A.; Kotha, S.; Ratti, G.; Meri, S. Complement Dysregulation in Glomerulonephritis. Semin Immunol. 2019, 45, 101331. [Google Scholar] [CrossRef] [PubMed]
- Rajasekaran, A.; Julian, B.A.; Rizk, D.V. IgA Nephropathy: An Interesting Autoimmune Kidney Disease. Am. J. Med. Sci. 2021, 361, 176–194. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Gao, C.; Dai, H.; Zheng, Y.; Dong, Z.; Gao, Y.; Liu, F.; Zhang, Z.; Liu, Z.; Liu, W.; et al. Immunological Pathogenesis of Membranous Nephropathy: Focus on PLA2R1 and Its Role. Front Immunol. 2019, 10, 1809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathomthongtaweechai, N.; Chutipongtanate, S. AGE/RAGE Signaling-Mediated Endoplasmic Reticulum Stress and Future Prospects in Non-Coding RNA Therapeutics for Diabetic Nephropathy. Biomed. Pharmacother. 2020, 131, 110655. [Google Scholar] [CrossRef]
- Kida, Y. Peritubular Capillary Rarefaction: An Underappreciated Regulator of CKD Progression. Int. J. Mol. Sci. 2020, 21, E8255. [Google Scholar] [CrossRef]
- Tang, C.; Livingston, M.J.; Liu, Z.; Dong, Z. Autophagy in Kidney Homeostasis and Disease. Nat. Rev. Nephrol. 2020, 16, 489–508. [Google Scholar] [CrossRef]
- Turkmen, K. Inflammation, Oxidative Stress, Apoptosis, and Autophagy in Diabetes Mellitus and Diabetic Kidney Disease: The Four Horsemen of the Apocalypse. Int. Urol. Nephrol. 2017, 49, 837–844. [Google Scholar] [CrossRef]
- Zhou, B.; Wan, Y.; Chen, R.; Zhang, C.; Li, X.; Meng, F.; Glaser, S.; Wu, N.; Zhou, T.; Li, S.; et al. The Emerging Role of Cellular Senescence in Renal Diseases. J. Cell. Mol. Med. 2020, 24, 2087–2097. [Google Scholar] [CrossRef]
- Djudjaj, S.; Boor, P. Cellular and Molecular Mechanisms of Kidney Fibrosis. Mol. Aspects Med. 2019, 65, 16–36. [Google Scholar] [CrossRef]
- Tang, S.C.W.; Yiu, W.H. Innate Immunity in Diabetic Kidney Disease. Nat. Rev. Nephrol. 2020, 16, 206–222. [Google Scholar] [CrossRef]
- Guo, J.; Zheng, H.J.; Zhang, W.; Lou, W.; Xia, C.; Han, X.T.; Huang, W.J.; Zhang, F.; Wang, Y.; Liu, W.J. Accelerated Kidney Aging in Diabetes Mellitus. Oxid. Med. Cell. Longev. 2020, 2020, 1234059. [Google Scholar] [CrossRef] [PubMed]
- Shikata, K.; Makino, H. Microinflammation in the Pathogenesis of Diabetic Nephropathy. J. Diabetes Investig. 2013, 4, 142–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobeika, L.; Ng, L.; Lee, I.J. Moving Forward with Biologics in Lupus Nephritis. Adv. Chronic Kidney Dis. 2019, 26, 338–350. [Google Scholar] [CrossRef]
- García-Carro, C.; Vergara, A.; Agraz, I.; Jacobs-Cachá, C.; Espinel, E.; Seron, D.; Soler, M.J. The New Era for Reno-Cardiovascular Treatment in Type 2 Diabetes. J. Clin. Med. 2019, 8, E864. [Google Scholar] [CrossRef] [Green Version]
- Amir, R.; Suhl, S.; Alexander, C.M. Renal Evaluation and Protection. Clin. Geriatr. Med. 2020, 36, 431–445. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Xie, Y.; Li, Q.; Chen, X. Stem Cell-Based Cell Therapy for Glomerulonephritis. Biomed. Res. Int. 2014, 2014, 124730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perico, N.; Casiraghi, F.; Remuzzi, G. Clinical Translation of Mesenchymal Stromal Cell Therapies in Nephrology. J. Am. Soc. Nephrol. 2018, 29, 362–375. [Google Scholar] [CrossRef]
- Bochon, B.; Kozubska, M.; Surygała, G.; Witkowska, A.; Kuźniewicz, R.; Grzeszczak, W.; Wystrychowski, G. Mesenchymal Stem Cells-Potential Applications in Kidney Diseases. Int. J. Mol. Sci. 2019, 20, 2462. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, K.; Kitamura, S.; Wada, J. Immunomodulatory and Regenerative Effects of Mesenchymal Stem Cell-Derived Extracellular Vesicles in Renal Diseases. Int. J. Mol. Sci. 2020, 21, 756. [Google Scholar] [CrossRef] [Green Version]
- Cable, J.; Fuchs, E.; Weissman, I.; Jasper, H.; Glass, D.; Rando, T.A.; Blau, H.; Debnath, S.; Oliva, A.; Park, S.; et al. Adult Stem Cells and Regenerative Medicine-a Symposium Report. Ann. N. Y. Acad. Sci. 2020, 1462, 27–36. [Google Scholar] [CrossRef]
- Mahla, R.S. Stem Cells Applications in Regenerative Medicine and Disease Therapeutics. Int. J. Cell Biol. 2016, 2016, 6940283. [Google Scholar] [CrossRef] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Andrzejewska, A.; Lukomska, B.; Janowski, M. Concise Review: Mesenchymal Stem Cells: From Roots to Boost. Stem Cells 2019, 37, 855–864. [Google Scholar] [CrossRef] [Green Version]
- Alessio, N.; Stellavato, A.; Aprile, D.; Cimini, D.; Vassallo, V.; Di Bernardo, G.; Galderisi, U.; Schiraldi, C. Timely Supplementation of Hydrogels Containing Sulfated or Unsulfated Chondroitin and Hyaluronic Acid Affects Mesenchymal Stromal Cells Commitment toward Chondrogenic Differentiation. Front Cell Dev. Biol. 2021, 9, 641529. [Google Scholar] [CrossRef]
- Yuan, L.; Liu, H.-Q.; Wu, M.-J. Human Embryonic Mesenchymal Stem Cells Participate in Differentiation of Renal Tubular Cells in Newborn Mice. Exp. Ther. Med. 2016, 12, 641–648. [Google Scholar] [CrossRef] [Green Version]
- Naderi-Meshkin, H.; Bahrami, A.R.; Bidkhori, H.R.; Mirahmadi, M.; Ahmadiankia, N. Strategies to Improve Homing of Mesenchymal Stem Cells for Greater Efficacy in Stem Cell Therapy. Cell Biol. Int. 2015, 39, 23–34. [Google Scholar] [CrossRef]
- Deng, M.; Mei, T.; Hou, T.; Luo, K.; Luo, F.; Yang, A.; Yu, B.; Pang, H.; Dong, S.; Xu, J. TGFβ3 Recruits Endogenous Mesenchymal Stem Cells to Initiate Bone Regeneration. Stem Cell Res. Ther. 2017, 8, 258. [Google Scholar] [CrossRef] [Green Version]
- Ullah, I.; Subbarao, R.B.; Rho, G.J. Human Mesenchymal Stem Cells—Current Trends and Future Prospective. Biosci. Rep. 2015, 35, e00191. [Google Scholar] [CrossRef]
- Jiang, W.; Xu, J. Immune Modulation by Mesenchymal Stem Cells. Cell Prolif. 2020, 53, e12712. [Google Scholar] [CrossRef]
- Biancone, L.; Cantaluppi, V.; Duò, D.; Deregibus, M.C.; Torre, C.; Camussi, G. Role of L-Selectin in the Vascular Homing of Peripheral Blood-Derived Endothelial Progenitor Cells. J. Immunol. 2004, 173, 5268–5274. [Google Scholar] [CrossRef] [Green Version]
- Borgovan, T.; Crawford, L.; Nwizu, C.; Quesenberry, P. Stem Cells and Extracellular Vesicles: Biological Regulators of Physiology and Disease. Am. J. Physiol. Cell Physiol. 2019, 317, C155–C166. [Google Scholar] [CrossRef]
- Tetta, C.; Deregibus, M.C.; Camussi, G. Stem Cells and Stem Cell-Derived Extracellular Vesicles in Acute and Chronic Kidney Diseases: Mechanisms of Repair. Ann. Transl. Med. 2020, 8, 570. [Google Scholar] [CrossRef]
- Imafuku, A.; Sjoqvist, S. Extracellular Vesicle Therapeutics in Regenerative Medicine. Adv. Exp. Med. Biol. 2021, 1312, 131–138. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding Light on the Cell Biology of Extracellular Vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Cantaluppi, V.; Gatti, S.; Medica, D.; Figliolini, F.; Bruno, S.; Deregibus, M.C.; Sordi, A.; Biancone, L.; Tetta, C.; Camussi, G. Microvesicles Derived from Endothelial Progenitor Cells Protect the Kidney from Ischemia-Reperfusion Injury by MicroRNA-Dependent Reprogramming of Resident Renal Cells. Kidney Int. 2012, 82, 412–427. [Google Scholar] [CrossRef] [Green Version]
- Bruno, S.; Grange, C.; Deregibus, M.C.; Calogero, R.A.; Saviozzi, S.; Collino, F.; Morando, L.; Busca, A.; Falda, M.; Bussolati, B.; et al. Mesenchymal Stem Cell-Derived Microvesicles Protect against Acute Tubular Injury. J. Am. Soc. Nephrol. 2009, 20, 1053–1067. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Jia, H.; Zhang, B.; Wang, J.; Ji, C.; Zhu, X.; Yan, Y.; Yin, L.; Yu, J.; Qian, H.; et al. Pre-Incubation with HucMSC-Exosomes Prevents Cisplatin-Induced Nephrotoxicity by Activating Autophagy. Stem Cell Res. Ther. 2017, 8, 75. [Google Scholar] [CrossRef] [Green Version]
- Bruno, S.; Tapparo, M.; Collino, F.; Chiabotto, G.; Deregibus, M.C.; Soares Lindoso, R.; Neri, F.; Kholia, S.; Giunti, S.; Wen, S.; et al. Renal Regenerative Potential of Different Extracellular Vesicle Populations Derived from Bone Marrow Mesenchymal Stromal Cells. Tissue Eng. Part A 2017, 23, 1262–1273. [Google Scholar] [CrossRef]
- Zhou, Y.; Xu, H.; Xu, W.; Wang, B.; Wu, H.; Tao, Y.; Zhang, B.; Wang, M.; Mao, F.; Yan, Y.; et al. Exosomes Released by Human Umbilical Cord Mesenchymal Stem Cells Protect against Cisplatin-Induced Renal Oxidative Stress and Apoptosis In Vivo and In Vitro. Stem Cell Res. Ther. 2013, 4, 34. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Zou, X.; Miao, S.; Chen, J.; Du, T.; Zhong, L.; Ju, G.; Liu, G.; Zhu, Y. The Anti-Oxidative Role of Micro-Vesicles Derived from Human Wharton-Jelly Mesenchymal Stromal Cells through NOX2/Gp91(Phox) Suppression in Alleviating Renal Ischemia-Reperfusion Injury in Rats. PLoS ONE 2014, 9, e92129. [Google Scholar] [CrossRef]
- Zhang, G.; Zou, X.; Huang, Y.; Wang, F.; Miao, S.; Liu, G.; Chen, M.; Zhu, Y. Mesenchymal Stromal Cell-Derived Extracellular Vesicles Protect against Acute Kidney Injury through Anti-Oxidation by Enhancing Nrf2/ARE Activation in Rats. Kidney Blood Press. Res. 2016, 41, 119–128. [Google Scholar] [CrossRef]
- Gu, D.; Zou, X.; Ju, G.; Zhang, G.; Bao, E.; Zhu, Y. Mesenchymal Stromal Cells Derived Extracellular Vesicles Ameliorate Acute Renal Ischemia Reperfusion Injury by Inhibition of Mitochondrial Fission through MiR-30. Stem Cells Int. 2016, 2016, 2093940. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Liu, W.; Zhang, B.; Wang, J.; Wu, P.; Tandra, N.; Liang, Z.; Ji, C.; Yin, L.; Hu, X.; et al. HucMSC Exosomes-Delivered 14-3-3ζ Enhanced Autophagy via Modulation of ATG16L in Preventing Cisplatin-Induced Acute Kidney Injury. Am. J. Transl. Res. 2018, 10, 101–113. [Google Scholar]
- Xiang, J.; Jiang, T.; Zhang, W.; Xie, W.; Tang, X.; Zhang, J. Human Umbilical Cord-Derived Mesenchymal Stem Cells Enhanced HK-2 Cell Autophagy through MicroRNA-145 by Inhibiting the PI3K/AKT/MTOR Signaling Pathway. Exp. Cell Res. 2019, 378, 198–205. [Google Scholar] [CrossRef]
- Zou, X.; Zhang, G.; Cheng, Z.; Yin, D.; Du, T.; Ju, G.; Miao, S.; Liu, G.; Lu, M.; Zhu, Y. Microvesicles Derived from Human Wharton’s Jelly Mesenchymal Stromal Cells Ameliorate Renal Ischemia-Reperfusion Injury in Rats by Suppressing CX3CL1. Stem Cell Res. Ther. 2014, 5, 40. [Google Scholar] [CrossRef] [Green Version]
- Eirin, A.; Zhu, X.-Y.; Puranik, A.S.; Tang, H.; McGurren, K.A.; van Wijnen, A.J.; Lerman, A.; Lerman, L.O. Mesenchymal Stem Cell-Derived Extracellular Vesicles Attenuate Kidney Inflammation. Kidney Int. 2017, 92, 114–124. [Google Scholar] [CrossRef]
- Choi, H.Y.; Lee, H.G.; Kim, B.S.; Ahn, S.H.; Jung, A.; Lee, M.; Lee, J.E.; Kim, H.J.; Ha, S.K.; Park, H.C. Mesenchymal Stem Cell-Derived Microparticles Ameliorate Peritubular Capillary Rarefaction via Inhibition of Endothelial-Mesenchymal Transition and Decrease Tubulointerstitial Fibrosis in Unilateral Ureteral Obstruction. Stem Cell Res. Ther. 2015, 6, 18. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Yao, K.; Huuskes, B.M.; Shen, H.-H.; Zhuang, J.; Godson, C.; Brennan, E.P.; Wilkinson-Berka, J.L.; Wise, A.F.; Ricardo, S.D. Mesenchymal Stem Cells Deliver Exogenous MicroRNA-Let7c via Exosomes to Attenuate Renal Fibrosis. Mol. Ther. 2016, 24, 1290–1301. [Google Scholar] [CrossRef] [Green Version]
- Grange, C.; Tritta, S.; Tapparo, M.; Cedrino, M.; Tetta, C.; Camussi, G.; Brizzi, M.F. Stem Cell-Derived Extracellular Vesicles Inhibit and Revert Fibrosis Progression in a Mouse Model of Diabetic Nephropathy. Sci. Rep. 2019, 9, 4468. [Google Scholar] [CrossRef] [Green Version]
- Zhong, L.; Liao, G.; Wang, X.; Li, L.; Zhang, J.; Chen, Y.; Liu, J.; Liu, S.; Wei, L.; Zhang, W.; et al. Mesenchymal Stem Cells-Microvesicle-MiR-451a Ameliorate Early Diabetic Kidney Injury by Negative Regulation of P15 and P19. Exp. Biol. Med. 2018, 243, 1233–1242. [Google Scholar] [CrossRef]
- He, J.; Wang, Y.; Sun, S.; Yu, M.; Wang, C.; Pei, X.; Zhu, B.; Wu, J.; Zhao, W. Bone Marrow Stem Cells-Derived Microvesicles Protect against Renal Injury in the Mouse Remnant Kidney Model. Nephrology 2012, 17, 493–500. [Google Scholar] [CrossRef]
- Wang, Y.; Fu, B.; Sun, X.; Li, D.; Huang, Q.; Zhao, W.; Chen, X. Differentially Expressed MicroRNAs in Bone Marrow Mesenchymal Stem Cell-Derived Microvesicles in Young and Older Rats and Their Effect on Tumor Growth Factor-Β1-Mediated Epithelial-Mesenchymal Transition in HK2 Cells. Stem Cell Res. Ther. 2015, 6, 185. [Google Scholar] [CrossRef] [Green Version]
- Ju, G.; Cheng, J.; Zhong, L.; Wu, S.; Zou, X.; Zhang, G.; Gu, D.; Miao, S.; Zhu, Y.; Sun, J.; et al. Microvesicles Derived from Human Umbilical Cord Mesenchymal Stem Cells Facilitate Tubular Epithelial Cell Dedifferentiation and Growth via Hepatocyte Growth Factor Induction. PLoS ONE 2015, 10, e0121534. [Google Scholar] [CrossRef] [Green Version]
- Gatti, S.; Bruno, S.; Deregibus, M.C.; Sordi, A.; Cantaluppi, V.; Tetta, C.; Camussi, G. Microvesicles Derived from Human Adult Mesenchymal Stem Cells Protect against Ischaemia-Reperfusion-Induced Acute and Chronic Kidney Injury. Nephrol. Dial. Transplant. 2011, 26, 1474–1483. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.; Gu, D.; Xing, X.; Cheng, Z.; Gong, D.; Zhang, G.; Zhu, Y. Human Mesenchymal Stromal Cell-Derived Extracellular Vesicles Alleviate Renal Ischemic Reperfusion Injury and Enhance Angiogenesis in Rats. Am. J. Transl. Res. 2016, 8, 4289–4299. [Google Scholar]
- Zhang, C.; Ma, P.; Zhao, Z.; Jiang, N.; Lian, D.; Huo, P.; Yang, H. MiRNA-mRNA Regulatory Network Analysis of Mesenchymal Stem Cell Treatment in Cisplatin-induced Acute Kidney Injury Identifies Roles for MiR-210/Serpine1 and MiR-378/Fos in Regulating Inflammation. Mol. Med. Rep. 2019, 20, 1509–1522. [Google Scholar] [CrossRef] [Green Version]
- Ti, D.; Hao, H.; Tong, C.; Liu, J.; Dong, L.; Zheng, J.; Zhao, Y.; Liu, H.; Fu, X.; Han, W. LPS-Preconditioned Mesenchymal Stromal Cells Modify Macrophage Polarization for Resolution of Chronic Inflammation via Exosome-Shuttled Let-7b. J. Transl. Med. 2015, 13, 308. [Google Scholar] [CrossRef] [Green Version]
- Lopatina, T.; Bruno, S.; Tetta, C.; Kalinina, N.; Porta, M.; Camussi, G. Platelet-Derived Growth Factor Regulates the Secretion of Extracellular Vesicles by Adipose Mesenchymal Stem Cells and Enhances Their Angiogenic Potential. Cell Commun. Signal. 2014, 12, 26. [Google Scholar] [CrossRef] [Green Version]
- Crain, S.K.; Robinson, S.R.; Thane, K.E.; Davis, A.M.; Meola, D.M.; Barton, B.A.; Yang, V.K.; Hoffman, A.M. Extracellular Vesicles from Wharton’s Jelly Mesenchymal Stem Cells Suppress CD4 Expressing T Cells Through Transforming Growth Factor Beta and Adenosine Signaling in a Canine Model. Stem Cells Dev. 2019, 28, 212–226. [Google Scholar] [CrossRef]
- Shen, G.; Krienke, S.; Schiller, P.; Nießen, A.; Neu, S.; Eckstein, V.; Schiller, M.; Lorenz, H.-M.; Tykocinski, L.-O. Microvesicles Released by Apoptotic Human Neutrophils Suppress Proliferation and IL-2/IL-2 Receptor Expression of Resting T Helper Cells. Eur. J. Immunol. 2017, 47, 900–910. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Huang, Y.; Han, J.; Yu, L.; Li, Y.; Lu, Z.; Li, H.; Liu, Z.; Shi, C.; Duan, F.; et al. Immunomodulatory Effects of Mesenchymal Stromal Cells-Derived Exosome. Immunol. Res. 2016, 64, 831–840. [Google Scholar] [CrossRef]
- Qiu, P.; Zhou, J.; Zhang, J.; Dong, Y.; Liu, Y. Exosome: The Regulator of the Immune System in Sepsis. Front Pharmacol. 2021, 12, 671164. [Google Scholar] [CrossRef]
- Perez-Hernandez, J.; Redon, J.; Cortes, R. Extracellular Vesicles as Therapeutic Agents in Systemic Lupus Erythematosus. Int. J. Mol. Sci. 2017, 18, E717. [Google Scholar] [CrossRef] [Green Version]
- Elahi, F.M.; Farwell, D.G.; Nolta, J.A.; Anderson, J.D. Preclinical Translation of Exosomes Derived from Mesenchymal Stem/Stromal Cells. Stem Cells 2020, 38, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Chen, Y.; Yin, G.; Xie, Q. Therapeutic Prospects of MicroRNAs Carried by Mesenchymal Stem Cells-Derived Extracellular Vesicles in Autoimmune Diseases. Life Sci. 2021, 277, 119458. [Google Scholar] [CrossRef]
- Grange, C.; Papadimitriou, E.; Dimuccio, V.; Pastorino, C.; Molina, J.; O’Kelly, R.; Niedernhofer, L.J.; Robbins, P.D.; Camussi, G.; Bussolati, B. Urinary Extracellular Vesicles Carrying Klotho Improve the Recovery of Renal Function in an Acute Tubular Injury Model. Mol. Ther. 2020, 28, 490–502. [Google Scholar] [CrossRef] [Green Version]
- Abe-Yoshio, Y.; Abe, K.; Miyazaki, M.; Furusu, A.; Nishino, T.; Harada, T.; Koji, T.; Kohno, S. Involvement of Bone Marrow-Derived Endothelial Progenitor Cells in Glomerular Capillary Repair in Habu Snake Venom-Induced Glomerulonephritis. Virchows Arch. 2008, 453, 97–106. [Google Scholar] [CrossRef]
- Cantaluppi, V.; Medica, D.; Mannari, C.; Stiaccini, G.; Figliolini, F.; Dellepiane, S.; Quercia, A.D.; Migliori, M.; Panichi, V.; Giovannini, L.; et al. Endothelial Progenitor Cell-Derived Extracellular Vesicles Protect from Complement-Mediated Mesangial Injury in Experimental Anti-Thy1.1 Glomerulonephritis. Nephrol. Dial. Transplant. 2015, 30, 410–422. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Dai, S.-C.; Luan, X.; Chen, J.; Cannavicci, A. Dysfunction and Therapeutic Potential of Endothelial Progenitor Cells in Diabetes Mellitus. J. Clin. Med. Res. 2018, 10, 752–757. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.-Y.; Cheong, S.-K.; Mok, P.-L.; Leong, C.-F. Differentiation of Human Mesenchymal Stem Cells into Mesangial Cells in Post-Glomerular Injury Murine Model. Pathology 2008, 40, 52–57. [Google Scholar] [CrossRef]
- Ronconi, E.; Sagrinati, C.; Angelotti, M.L.; Lazzeri, E.; Mazzinghi, B.; Ballerini, L.; Parente, E.; Becherucci, F.; Gacci, M.; Carini, M.; et al. Regeneration of Glomerular Podocytes by Human Renal Progenitors. J. Am. Soc. Nephrol. 2009, 20, 322–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radmanesh, F.; Mahmoudi, M.; Yazdanpanah, E.; Keyvani, V.; Kia, N.; Nikpoor, A.R.; Zafari, P.; Esmaeili, S.-A. The Immunomodulatory Effects of Mesenchymal Stromal Cell-Based Therapy in Human and Animal Models of Systemic Lupus Erythematosus. IUBMB Life 2020, 72, 2366–2381. [Google Scholar] [CrossRef] [PubMed]
- El-Jawhari, J.J.; El-Sherbiny, Y.; McGonagle, D.; Jones, E. Multipotent Mesenchymal Stromal Cells in Rheumatoid Arthritis and Systemic Lupus Erythematosus; From a Leading Role in Pathogenesis to Potential Therapeutic Saviors? Front Immunol. 2021, 12, 643170. [Google Scholar] [CrossRef]
- Zhang, B.; Yeo, R.W.Y.; Lai, R.C.; Sim, E.W.K.; Chin, K.C.; Lim, S.K. Mesenchymal Stromal Cell Exosome-Enhanced Regulatory T-Cell Production through an Antigen-Presenting Cell-Mediated Pathway. Cytotherapy 2018, 20, 687–696. [Google Scholar] [CrossRef]
- Zhang, B.; Yin, Y.; Lai, R.C.; Tan, S.S.; Choo, A.B.H.; Lim, S.K. Mesenchymal Stem Cells Secrete Immunologically Active Exosomes. Stem Cells Dev. 2014, 23, 1233–1244. [Google Scholar] [CrossRef]
- Lo Sicco, C.; Reverberi, D.; Balbi, C.; Ulivi, V.; Principi, E.; Pascucci, L.; Becherini, P.; Bosco, M.C.; Varesio, L.; Franzin, C.; et al. Mesenchymal Stem Cell-Derived Extracellular Vesicles as Mediators of Anti-Inflammatory Effects: Endorsement of Macrophage Polarization. Stem Cells Transl. Med. 2017, 6, 1018–1028. [Google Scholar] [CrossRef]
- Kordelas, L.; Rebmann, V.; Ludwig, A.-K.; Radtke, S.; Ruesing, J.; Doeppner, T.R.; Epple, M.; Horn, P.A.; Beelen, D.W.; Giebel, B. MSC-Derived Exosomes: A Novel Tool to Treat Therapy-Refractory Graft-versus-Host Disease. Leukemia 2014, 28, 970–973. [Google Scholar] [CrossRef]
- Li, M.; Soder, R.; Abhyankar, S.; Abdelhakim, H.; Braun, M.W.; Trinidad, C.V.; Pathak, H.B.; Pessetto, Z.; Deighan, C.; Ganguly, S.; et al. WJMSC-Derived Small Extracellular Vesicle Enhance T Cell Suppression through PD-L1. J. Extracell. Vesicles 2021, 10, e12067. [Google Scholar] [CrossRef]
- Ma, H.; Liu, C.; Shi, B.; Zhang, Z.; Feng, R.; Guo, M.; Lu, L.; Shi, S.; Gao, X.; Chen, W.; et al. Mesenchymal Stem Cells Control Complement C5 Activation by Factor H in Lupus Nephritis. EBioMedicine 2018, 32, 21–30. [Google Scholar] [CrossRef]
- Yuan, X.; Qin, X.; Wang, D.; Zhang, Z.; Tang, X.; Gao, X.; Chen, W.; Sun, L. Mesenchymal Stem Cell Therapy Induces FLT3L and CD1c+ Dendritic Cells in Systemic Lupus Erythematosus Patients. Nat. Commun. 2019, 10, 2498. [Google Scholar] [CrossRef]
- Uchimura, H.; Marumo, T.; Takase, O.; Kawachi, H.; Shimizu, F.; Hayashi, M.; Saruta, T.; Hishikawa, K.; Fujita, T. Intrarenal Injection of Bone Marrow-Derived Angiogenic Cells Reduces Endothelial Injury and Mesangial Cell Activation in Experimental Glomerulonephritis. J. Am. Soc. Nephrol. 2005, 16, 997–1004. [Google Scholar] [CrossRef]
- Chang, H.-H.; Hsu, S.-P.; Chien, C.-T. Intrarenal Transplantation of Hypoxic Preconditioned Mesenchymal Stem Cells Improves Glomerulonephritis through Anti-Oxidation, Anti-ER Stress, Anti-Inflammation, Anti-Apoptosis, and Anti-Autophagy. Antioxidants 2019, 9, E2. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.W.; Qiu, H.; Mei, Y.M.; Fu, H.; Zheng, H.G. Repair Effects of Umbilical Cord Mesenchymal Stem Cells on Podocyte Damage Of IgA Nephropathy. J. Biol. Regul. Homeost. Agents 2015, 29, 609–617. [Google Scholar]
- Yang, R.-C.; Zhu, X.-L.; Wang, J.; Wan, F.; Zhang, H.-Q.; Lin, Y.; Tang, X.-L.; Zhu, B. Bone Marrow Mesenchymal Stem Cells Attenuate the Progression of Focal Segmental Glomerulosclerosis in Rat Models. BMC Nephrol. 2018, 19, 335. [Google Scholar] [CrossRef]
- de Carvalho Ribeiro, P.; Oliveira, L.F.; Filho, M.A.; Caldas, H.C. Differentiating Induced Pluripotent Stem Cells into Renal Cells: A New Approach to Treat Kidney Diseases. Stem Cells Int. 2020, 2020, 8894590. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, Y.; Cai, R.; Jin, J.; He, Q. Differential Expression of Urinary Exosomal Small RNAs in Idiopathic Membranous Nephropathy. Biomed. Res. Int. 2020, 2020, 3170927. [Google Scholar] [CrossRef]
- Ma, H.; Xu, Y.; Zhang, R.; Guo, B.; Zhang, S.; Zhang, X. Differential Expression Study of Circular RNAs in Exosomes from Serum and Urine in Patients with Idiopathic Membranous Nephropathy. Arch. Med. Sci. 2019, 15, 738–753. [Google Scholar] [CrossRef]
- Müller-Deile, J.; Sopel, N.; Ohs, A.; Rose, V.; Gröner, M.; Wrede, C.; Hegermann, J.; Daniel, C.; Amann, K.; Zahner, G.; et al. Glomerular Endothelial Cell-Derived MicroRNA-192 Regulates Nephronectin Expression in Idiopathic Membranous Glomerulonephritis. J. Am. Soc. Nephrol. 2021, 32, 2777–2794. [Google Scholar] [CrossRef]
- Liu, D.; Liu, F.; Wang, X.; Qiao, Y.; Pan, S.; Yang, Y.; Hu, Y.; Zhang, Y.; Tian, F.; Liu, Z. MiR-130a-5p Prevents Angiotensin II-Induced Podocyte Apoptosis by Modulating M-Type Phospholipase A2 Receptor. Cell Cycle 2018, 17, 2484–2495. [Google Scholar] [CrossRef] [Green Version]
- Nalewajska, M.; Gurazda, K.; Styczyńska-Kowalska, E.; Marchelek-Myśliwiec, M.; Pawlik, A.; Dziedziejko, V. The Role of MicroRNAs in Selected Forms of Glomerulonephritis. Int. J. Mol. Sci. 2019, 20, E5050. [Google Scholar] [CrossRef] [Green Version]
- Lateef, A.; Petri, M. Unmet Medical Needs in Systemic Lupus Erythematosus. Arthritis Res. Ther. 2012, 14 (Suppl. S4), S4. [Google Scholar] [CrossRef] [Green Version]
- Parikh, S.V.; Almaani, S.; Brodsky, S.; Rovin, B.H. Update on Lupus Nephritis: Core Curriculum 2020. Am. J. Kidney Dis. 2020, 76, 265–281. [Google Scholar] [CrossRef]
- Macejova, Z.; Madarasova Geckova, A.; Husarova, D.; Zarikova, M.; Kotradyova, Z. Living with Systematic Lupus Erythematosus: A Profile of Young Female Patients. Int. J. Environ. Res. Public Health 2020, 17, E1315. [Google Scholar] [CrossRef] [Green Version]
- Kronbichler, A.; Brezina, B.; Gauckler, P.; Quintana, L.F.; Jayne, D.R.W. Refractory Lupus Nephritis: When, Why and How to Treat. Autoimmun. Rev. 2019, 18, 510–518. [Google Scholar] [CrossRef]
- Morales, E.; Galindo, M.; Trujillo, H.; Praga, M. Update on Lupus Nephritis: Looking for a New Vision. Nephron 2021, 145, 1–13. [Google Scholar] [CrossRef]
- Sattwika, P.D.; Mustafa, R.; Paramaiswari, A.; Herningtyas, E.H. Stem Cells for Lupus Nephritis: A Concise Review of Current Knowledge. Lupus 2018, 27, 1881–1897. [Google Scholar] [CrossRef]
- Dorraji, S.E.; Hovd, A.-M.K.; Kanapathippillai, P.; Bakland, G.; Eilertsen, G.Ø.; Figenschau, S.L.; Fenton, K.A. Mesenchymal Stem Cells and T Cells in the Formation of Tertiary Lymphoid Structures in Lupus Nephritis. Sci. Rep. 2018, 8, 7861. [Google Scholar] [CrossRef]
- Fathollahi, A.; Gabalou, N.B.; Aslani, S. Mesenchymal Stem Cell Transplantation in Systemic Lupus Erythematous, a Mesenchymal Stem Cell Disorder. Lupus 2018, 27, 1053–1064. [Google Scholar] [CrossRef]
- Jang, E.; Jeong, M.; Kim, S.; Jang, K.; Kang, B.-K.; Lee, D.Y.; Bae, S.-C.; Kim, K.S.; Youn, J. Infusion of Human Bone Marrow-Derived Mesenchymal Stem Cells Alleviates Autoimmune Nephritis in a Lupus Model by Suppressing Follicular Helper T-Cell Development. Cell Transplant. 2016, 25, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Tani, C.; Vagnani, S.; Carli, L.; Querci, F.; Kühl, A.A.; Spieckermann, S.; Cieluch, C.P.; Pacini, S.; Fazzi, R.; Mosca, M. Treatment with Allogenic Mesenchymal Stromal Cells in a Murine Model of Systemic Lupus Erythematosus. Int. J. Stem Cells 2017, 10, 160–168. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, W.; Liu, C.; Yan, M.; Raman, I.; Du, Y.; Fang, X.; Zhou, X.J.; Mohan, C.; Li, Q.-Z. Delivering Oxidation Resistance-1 (OXR1) to Mouse Kidney by Genetic Modified Mesenchymal Stem Cells Exhibited Enhanced Protection against Nephrotoxic Serum Induced Renal Injury and Lupus Nephritis. J. Stem Cell Res. Ther. 2014, 4, 231. [Google Scholar] [CrossRef]
- Yang, X.; Yang, J.; Li, X.; Ma, W.; Zou, H. Bone Marrow-Derived Mesenchymal Stem Cells Inhibit T Follicular Helper Cell in Lupus-Prone Mice. Lupus 2018, 27, 49–59. [Google Scholar] [CrossRef]
- Zhang, W.; Feng, Y.-L.; Pang, C.-Y.; Lu, F.-A.; Wang, Y.-F. Transplantation of Adipose Tissue-Derived Stem Cells Ameliorates Autoimmune Pathogenesis in MRL/Lpr Mice: Modulation of the Balance between Th17 and Treg. Z. Rheumatol. 2019, 78, 82–88. [Google Scholar] [CrossRef]
- Zhang, Z.; Niu, L.; Tang, X.; Feng, R.; Yao, G.; Chen, W.; Li, W.; Feng, X.; Chen, H.; Sun, L. Mesenchymal Stem Cells Prevent Podocyte Injury in Lupus-Prone B6.MRL-Faslpr Mice via Polarizing Macrophage into an Anti-Inflammatory Phenotype. Nephrol. Dial. Transplant. 2019, 34, 597–605. [Google Scholar] [CrossRef]
- Liu, J.; Lu, X.; Lou, Y.; Cai, Y.; Cui, W.; Wang, J.; Nie, P.; Chen, L.; Li, B.; Luo, P. Xenogeneic Transplantation of Human Placenta-Derived Mesenchymal Stem Cells Alleviates Renal Injury and Reduces Inflammation in a Mouse Model of Lupus Nephritis. Biomed. Res. Int. 2019, 2019, 9370919. [Google Scholar] [CrossRef]
- Liang, J.; Zhang, H.; Hua, B.; Wang, H.; Lu, L.; Shi, S.; Hou, Y.; Zeng, X.; Gilkeson, G.S.; Sun, L. Allogenic Mesenchymal Stem Cells Transplantation in Refractory Systemic Lupus Erythematosus: A Pilot Clinical Study. Ann. Rheum. Dis. 2010, 69, 1423–1429. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Wang, D.; Liang, J.; Zhang, H.; Feng, X.; Wang, H.; Hua, B.; Liu, B.; Ye, S.; Hu, X.; et al. Umbilical Cord Mesenchymal Stem Cell Transplantation in Severe and Refractory Systemic Lupus Erythematosus. Arthritis Rheum. 2010, 62, 2467–2475. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, H.; Liang, J.; Li, X.; Feng, X.; Wang, H.; Hua, B.; Liu, B.; Lu, L.; Gilkeson, G.S.; et al. Allogeneic Mesenchymal Stem Cell Transplantation in Severe and Refractory Systemic Lupus Erythematosus: 4 Years of Experience. Cell Transplant. 2013, 22, 2267–2277. [Google Scholar] [CrossRef]
- Wang, D.; Li, J.; Zhang, Y.; Zhang, M.; Chen, J.; Li, X.; Hu, X.; Jiang, S.; Shi, S.; Sun, L. Umbilical Cord Mesenchymal Stem Cell Transplantation in Active and Refractory Systemic Lupus Erythematosus: A Multicenter Clinical Study. Arthritis Res. Ther. 2014, 16, R79. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Wang, S.; Huang, S.; Zhang, Z.; Yuan, X.; Feng, X.; Lu, L.; Sun, L. Serum IFN-γ Predicts the Therapeutic Effect of Mesenchymal Stem Cells Transplantation in Systemic Lupus Erythematosus Patients. Stem Cells Transl. Med. 2017, 6, 1777–1785. [Google Scholar] [CrossRef]
- Gu, F.; Wang, D.; Zhang, H.; Feng, X.; Gilkeson, G.S.; Shi, S.; Sun, L. Allogeneic Mesenchymal Stem Cell Transplantation for Lupus Nephritis Patients Refractory to Conventional Therapy. Clin. Rheumatol. 2014, 33, 1611–1619. [Google Scholar] [CrossRef]
- Barbado, J.; Tabera, S.; Sánchez, A.; García-Sancho, J. Therapeutic Potential of Allogeneic Mesenchymal Stromal Cells Transplantation for Lupus Nephritis. Lupus 2018, 27, 2161–2165. [Google Scholar] [CrossRef]
- Deng, D.; Zhang, P.; Guo, Y.; Lim, T.O. A Randomised Double-Blind, Placebo-Controlled Trial of Allogeneic Umbilical Cord-Derived Mesenchymal Stem Cell for Lupus Nephritis. Ann. Rheum. Dis. 2017, 76, 1436–1439. [Google Scholar] [CrossRef]
- Zhou, T.; Liao, C.; Li, H.-Y.; Lin, W.; Lin, S.; Zhong, H. Efficacy of Mesenchymal Stem Cells in Animal Models of Lupus Nephritis: A Meta-Analysis. Stem Cell Res. Ther. 2020, 11, 48. [Google Scholar] [CrossRef]
- Zhao, Y.; Wei, W.; Liu, M.-L. Extracellular Vesicles and Lupus Nephritis—New Insights into Pathophysiology and Clinical Implications. J. Autoimmun. 2020, 115, 102540. [Google Scholar] [CrossRef]
- Ye, H.; Su, B.; Ni, H.; Li, L.; Chen, X.; You, X.; Zhang, H. MicroRNA-199a May Be Involved in the Pathogenesis of Lupus Nephritis via Modulating the Activation of NF-ΚB by Targeting Klotho. Mol. Immunol. 2018, 103, 235–242. [Google Scholar] [CrossRef]
- Furuhashi, K.; Tsuboi, N.; Shimizu, A.; Katsuno, T.; Kim, H.; Saka, Y.; Ozaki, T.; Sado, Y.; Imai, E.; Matsuo, S.; et al. Serum-Starved Adipose-Derived Stromal Cells Ameliorate Crescentic GN by Promoting Immunoregulatory Macrophages. J. Am. Soc. Nephrol. 2013, 24, 587–603. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Yan, M.; Yang, J.; Raman, I.; Du, Y.; Min, S.; Fang, X.; Mohan, C.; Li, Q.-Z. Glutathione S-Transferase Mu 2-Transduced Mesenchymal Stem Cells Ameliorated Anti-Glomerular Basement Membrane Antibody-Induced Glomerulonephritis by Inhibiting Oxidation and Inflammation. Stem Cell Res. Ther. 2014, 5, 19. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Iyoda, M.; Shibata, T.; Ohtaki, H.; Matsumoto, K.; Shindo-Hirai, Y.; Kuno, Y.; Wada, Y.; Yamamoto, Y.; Kawaguchi, M.; et al. Therapeutic Effects of Human Mesenchymal Stem Cells in Wistar-Kyoto Rats with Anti-Glomerular Basement Membrane Glomerulonephritis. PLoS ONE 2013, 8, e67475. [Google Scholar] [CrossRef] [Green Version]
- Iseri, K.; Iyoda, M.; Ohtaki, H.; Matsumoto, K.; Wada, Y.; Suzuki, T.; Yamamoto, Y.; Saito, T.; Hihara, K.; Tachibana, S.; et al. Therapeutic Effects and Mechanism of Conditioned Media from Human Mesenchymal Stem Cells on Anti-GBM Glomerulonephritis in WKY Rats. Am. J. Physiol. Renal Physiol. 2016, 310, F1182–F1191. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Tang, S.C.W. Recent Progress in Stem Cell Therapy for Diabetic Nephropathy. Kidney Dis. 2016, 2, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Ezquer, M.E.; Ezquer, F.E.; Arango-Rodríguez, M.L.; Conget, P.A. MSC Transplantation: A Promising Therapeutic Strategy to Manage the Onset and Progression of Diabetic Nephropathy. Biol. Res. 2012, 45, 289–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, R.H.; Seo, M.J.; Reger, R.L.; Spees, J.L.; Pulin, A.A.; Olson, S.D.; Prockop, D.J. Multipotent Stromal Cells from Human Marrow Home to and Promote Repair of Pancreatic Islets and Renal Glomeruli in Diabetic NOD/Scid Mice. Proc. Natl. Acad. Sci. USA 2006, 103, 17438–17443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Zhao, F.; Zhang, W.; Lv, J.; Lv, J.; Yin, A. BMSCs and MiR-124a Ameliorated Diabetic Nephropathy via Inhibiting Notch Signalling Pathway. J. Cell. Mol. Med. 2018, 22, 4840–4855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, H.; Dai, C. Effects of Bone Marrow Mesenchymal Stem Cells on Plasminogen Activator Inhibitor-1 and Renal Fibrosis in Rats with Diabetic Nephropathy. Arch. Med. Res. 2016, 47, 71–77. [Google Scholar] [CrossRef]
- Bai, Y.; Wang, J.; He, Z.; Yang, M.; Li, L.; Jiang, H. Mesenchymal Stem Cells Reverse Diabetic Nephropathy Disease via Lipoxin A4 by Targeting Transforming Growth Factor β (TGF-β)/Smad Pathway and Pro-Inflammatory Cytokines. Med. Sci. Monit. 2019, 25, 3069–3076. [Google Scholar] [CrossRef]
- Yuan, Y.; Yuan, L.; Li, L.; Liu, F.; Liu, J.; Chen, Y.; Cheng, J.; Lu, Y. Mitochondrial Transfer from Mesenchymal Stem Cells to Macrophages Restricts Inflammation and Alleviates Kidney Injury in Diabetic Nephropathy Mice via PGC-1α Activation. Stem Cells 2021, 39, 913–928. [Google Scholar] [CrossRef]
- Li, Y.; Liu, J.; Liao, G.; Zhang, J.; Chen, Y.; Li, L.; Li, L.; Liu, F.; Chen, B.; Guo, G.; et al. Early Intervention with Mesenchymal Stem Cells Prevents Nephropathy in Diabetic Rats by Ameliorating the Inflammatory Microenvironment. Int. J. Mol. Med. 2018, 41, 2629–2639. [Google Scholar] [CrossRef] [Green Version]
- Lv, S.-S.; Liu, G.; Wang, J.-P.; Wang, W.-W.; Cheng, J.; Sun, A.-L.; Liu, H.-Y.; Nie, H.-B.; Su, M.-R.; Guan, G.-J. Mesenchymal Stem Cells Transplantation Ameliorates Glomerular Injury in Streptozotocin-Induced Diabetic Nephropathy in Rats via Inhibiting Macrophage Infiltration. Int. Immunopharmacol. 2013, 17, 275–282. [Google Scholar] [CrossRef]
- Nagaishi, K.; Mizue, Y.; Chikenji, T.; Otani, M.; Nakano, M.; Konari, N.; Fujimiya, M. Mesenchymal Stem Cell Therapy Ameliorates Diabetic Nephropathy via the Paracrine Effect of Renal Trophic Factors Including Exosomes. Sci. Rep. 2016, 6, 34842. [Google Scholar] [CrossRef]
- Ebrahim, N.; Ahmed, I.A.; Hussien, N.I.; Dessouky, A.A.; Farid, A.S.; Elshazly, A.M.; Mostafa, O.; Gazzar, W.B.E.; Sorour, S.M.; Seleem, Y.; et al. Mesenchymal Stem Cell-Derived Exosomes Ameliorated Diabetic Nephropathy by Autophagy Induction through the MTOR Signaling Pathway. Cells 2018, 7, E226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, R.; Shen, J.; Hu, X. BMSCs-Derived Exosomal MicroRNA-Let-7a Plays a Protective Role in Diabetic Nephropathy via Inhibition of USP22 Expression. Life Sci. 2021, 268, 118937. [Google Scholar] [CrossRef] [PubMed]
- Ni, W.; Fang, Y.; Xie, L.; Liu, X.; Shan, W.; Zeng, R.; Liu, J.; Liu, X. Adipose-Derived Mesenchymal Stem Cells Transplantation Alleviates Renal Injury in Streptozotocin-Induced Diabetic Nephropathy. J. Histochem. Cytochem. 2015, 63, 842–853. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Tian, X.; Bai, S.; Fan, J.; Hou, W.; Tong, H.; Li, D. Autologous Transplantation of Adipose-Derived Mesenchymal Stem Cells Ameliorates Streptozotocin-Induced Diabetic Nephropathy in Rats by Inhibiting Oxidative Stress, pro-Inflammatory Cytokines and the P38 MAPK Signaling Pathway. Int. J. Mol. Med. 2012, 30, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, K.; Liu, X.; Li, D.; Luo, C.; Fu, B.; Cui, S.; Zhu, F.; Zhao, R.C.; Chen, X. Repeated Systemic Administration of Human Adipose-Derived Stem Cells Attenuates Overt Diabetic Nephropathy in Rats. Stem Cells Dev. 2013, 22, 3074–3086. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wang, N.; Zhang, L.; Hanyu, Z.; Xueyuan, B.; Fu, B.; Shaoyuan, C.; Zhang, W.; Xuefeng, S.; Li, R.; et al. Mesenchymal Stem Cells Protect Podocytes from Apoptosis Induced by High Glucose via Secretion of Epithelial Growth Factor. Stem Cell Res. Ther. 2013, 4, 103. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.; Luo, Q.; Wang, Y.; Ma, Y.; Chen, F.; Zhu, X.; Shi, J. Adipose Mesenchymal Stem Cell-Derived Extracellular Vesicles Containing MicroRNA-26a-5p Target TLR4 and Protect against Diabetic Nephropathy. J. Biol. Chem. 2020, 295, 12868–12884. [Google Scholar] [CrossRef]
- An, X.; Liao, G.; Chen, Y.; Luo, A.; Liu, J.; Yuan, Y.; Li, L.; Yang, L.; Wang, H.; Liu, F.; et al. Intervention for Early Diabetic Nephropathy by Mesenchymal Stem Cells in a Preclinical Nonhuman Primate Model. Stem Cell Res. Ther. 2019, 10, 363. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.; Zhang, Q.; Wang, W.; Liu, Q.; Liu, S.; Song, Y.; Wang, X.; Zhang, Y.; Li, S.; et al. Human Umbilical Cord Mesenchymal Stem Cells Attenuate Podocyte Injury under High Glucose via TLR2 and TLR4 Signaling. Diabetes Res. Clin. Pract. 2021, 173, 108702. [Google Scholar] [CrossRef]
- Park, J.H.; Hwang, I.; Hwang, S.H.; Han, H.; Ha, H. Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells Prevent Diabetic Renal Injury through Paracrine Action. Diabetes Res. Clin. Pract. 2012, 98, 465–473. [Google Scholar] [CrossRef]
- Li, H.; Rong, P.; Ma, X.; Nie, W.; Chen, Y.; Zhang, J.; Dong, Q.; Yang, M.; Wang, W. Mouse Umbilical Cord Mesenchymal Stem Cell Paracrine Alleviates Renal Fibrosis in Diabetic Nephropathy by Reducing Myofibroblast Transdifferentiation and Cell Proliferation and Upregulating MMPs in Mesangial Cells. J. Diabetes Res. 2020, 2020, 3847171. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Ma, J.; Li, S.; Liu, W. Applicability of Adipose-Derived Mesenchymal Stem Cells in Treatment of Patients with Type 2 Diabetes. Stem Cell Res. Ther. 2019, 10, 274. [Google Scholar] [CrossRef]
- Ezquer, F.E.; Ezquer, M.E.; Parrau, D.B.; Carpio, D.; Yañez, A.J.; Conget, P.A. Systemic Administration of Multipotent Mesenchymal Stromal Cells Reverts Hyperglycemia and Prevents Nephropathy in Type 1 Diabetic Mice. Biol. Blood Marrow Transplant. 2008, 14, 631–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Li, Y.; Zhao, J.; Zhang, J.; Huang, Y. Mesenchymal Stem Cells Ameliorate Podocyte Injury and Proteinuria in a Type 1 Diabetic Nephropathy Rat Model. Biol. Blood Marrow Transplant. 2013, 19, 538–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Tian, H.; Long, Y.; Zhang, X.; Zhong, L.; Deng, L.; Chen, X.; Li, X. Mesenchymal Stem Cells Transplantation Mildly Ameliorates Experimental Diabetic Nephropathy in Rats. Chin. Med. J. 2009, 122, 2573–2579. [Google Scholar]
- Lin, L.; Lin, H.; Wang, D.; Bao, Z.; Cai, H.; Zhang, X. Bone Marrow Mesenchymal Stem Cells Ameliorated Kidney Fibrosis by Attenuating TLR4/NF-ΚB in Diabetic Rats. Life Sci. 2020, 262, 118385. [Google Scholar] [CrossRef]
- Pan, X.-H.; Yang, X.-Y.; Yao, X.; Sun, X.-M.; Zhu, L.; Wang, J.-X.; Pang, R.-Q.; Cai, X.-M.; Dai, J.-J.; Ruan, G.-P. Bone-Marrow Mesenchymal Stem Cell Transplantation to Treat Diabetic Nephropathy in Tree Shrews. Cell Biochem. Funct. 2014, 32, 453–463. [Google Scholar] [CrossRef]
- Liu, D.; Zheng, W.; Pan, S.; Liu, Z. Concise Review: Current Trends on Applications of Stem Cells in Diabetic Nephropathy. Cell Death Dis. 2020, 11, 1000. [Google Scholar] [CrossRef]
- Ende, N.; Chen, R.; Reddi, A.S. Transplantation of Human Umbilical Cord Blood Cells Improves Glycemia and Glomerular Hypertrophy in Type 2 Diabetic Mice. Biochem. Biophys. Res. Commun. 2004, 321, 168–171. [Google Scholar] [CrossRef]
- Masoad, R.E.; Ewais, M.M.S.; Tawfik, M.K.; Abd El-All, H.S. Effect of Mononuclear Cells versus Pioglitazone on Streptozotocin-Induced Diabetic Nephropathy in Rats. Pharmacol. Rep. 2012, 64, 1223–1233. [Google Scholar] [CrossRef]
- Park, J.H.; Park, J.; Hwang, S.H.; Han, H.; Ha, H. Delayed Treatment with Human Umbilical Cord Blood-Derived Stem Cells Attenuates Diabetic Renal Injury. Transplant. Proc. 2012, 44, 1123–1126. [Google Scholar] [CrossRef] [PubMed]
- Mohler, E.R.; Shi, Y.; Moore, J.; Bantly, A.; Hamamdzic, D.; Yoder, M.; Rader, D.J.; Putt, M.; Zhang, L.; Parmacek, M.; et al. Diabetes Reduces Bone Marrow and Circulating Porcine Endothelial Progenitor Cells, an Effect Ameliorated by Atorvastatin and Independent of Cholesterol. Cytometry A 2009, 75, 75–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leicht, S.F.; Schwarz, T.M.; Hermann, P.C.; Seissler, J.; Aicher, A.; Heeschen, C. Adiponectin Pretreatment Counteracts the Detrimental Effect of a Diabetic Environment on Endothelial Progenitors. Diabetes 2011, 60, 652–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pala, C.; Altun, I.; Koker, Y.; Kurnaz, F.; Sivgin, S.; Koçyiğit, I.; Tanrıverdi, F.; Kaynar, L.; Elmali, F.; Cetin, M.; et al. The Effect of Diabetes Mellitus and End-Stage Renal Disease on the Number of CD34+ Cells in the Blood. Ann. Hematol. 2013, 92, 1189–1194. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, S.; Severini, G.M.; Zauli, G.; Secchiero, P. Cell-Based Therapies for Diabetic Complications. Exp. Diabetes Res. 2012, 2012, 872504. [Google Scholar] [CrossRef]
- Bahlmann, F.H.; DeGroot, K.; Duckert, T.; Niemczyk, E.; Bahlmann, E.; Boehm, S.M.; Haller, H.; Fliser, D. Endothelial Progenitor Cell Proliferation and Differentiation Is Regulated by Erythropoietin. Kidney Int. 2003, 64, 1648–1652. [Google Scholar] [CrossRef] [Green Version]
- Mohler, E.R.; Zhang, L.; Medenilla, E.; Rogers, W.; French, B.; Bantly, A.; Moore, J.S.; Huan, Y.; Murashima, M.; Berns, J.S. Effect of Darbepoetin Alfa on Endothelial Progenitor Cells and Vascular Reactivity in Chronic Kidney Disease. Vasc. Med. 2011, 16, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Song, B.; Niclis, J.C.; Alikhan, M.A.; Sakkal, S.; Sylvain, A.; Kerr, P.G.; Laslett, A.L.; Bernard, C.A.; Ricardo, S.D. Generation of Induced Pluripotent Stem Cells from Human Kidney Mesangial Cells. J. Am. Soc. Nephrol. 2011, 22, 1213–1220. [Google Scholar] [CrossRef]
- Zhou, T.; Benda, C.; Duzinger, S.; Huang, Y.; Li, X.; Li, Y.; Guo, X.; Cao, G.; Chen, S.; Hao, L.; et al. Generation of Induced Pluripotent Stem Cells from Urine. J. Am. Soc. Nephrol. 2011, 22, 1221–1228. [Google Scholar] [CrossRef]
- Rogers, I. Induced Pluripotent Stem Cells from Human Kidney. J. Am. Soc. Nephrol. 2011, 22, 1179–1180. [Google Scholar] [CrossRef]
- Song, B.; Smink, A.M.; Jones, C.V.; Callaghan, J.M.; Firth, S.D.; Bernard, C.A.; Laslett, A.L.; Kerr, P.G.; Ricardo, S.D. The Directed Differentiation of Human IPS Cells into Kidney Podocytes. PLoS ONE 2012, 7, e46453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tajiri, S.; Yamanaka, S.; Fujimoto, T.; Matsumoto, K.; Taguchi, A.; Nishinakamura, R.; Okano, H.J.; Yokoo, T. Regenerative Potential of Induced Pluripotent Stem Cells Derived from Patients Undergoing Haemodialysis in Kidney Regeneration. Sci. Rep. 2018, 8, 14919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, G.; Tao, L.; Ma, W.-J.; Gong, M.-J.; Zhao, L.; Shen, L.-J.; Long, C.-L.; Zhang, D.-Y.; Zhang, Y.-Y.; Wei, G.-H. Urine-Derived Stem Cells for the Therapy of Diabetic Nephropathy Mouse Model. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 1316–1324. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Liu, Y.; Niu, X.; Yin, J.; Hu, B.; Guo, S.; Fan, Y.; Wang, Y.; Wang, N. Exosomes Secreted by Human Urine-Derived Stem Cells Could Prevent Kidney Complications from Type I Diabetes in Rats. Stem Cell Res. Ther. 2016, 7, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Y.-R.; Chen, B.-P.; Chen, F.; Yang, S.-X.; Zhu, C.-Y.; Ma, Y.-L.; Li, Y.; Shi, J. Exosomal MicroRNA-16-5p from Human Urine-Derived Stem Cells Ameliorates Diabetic Nephropathy through Protection of Podocyte. J. Cell. Mol. Med. 2021, 25, 10798–10813. [Google Scholar] [CrossRef] [Green Version]
- Kunter, U.; Rong, S.; Boor, P.; Eitner, F.; Müller-Newen, G.; Djuric, Z.; van Roeyen, C.R.; Konieczny, A.; Ostendorf, T.; Villa, L.; et al. Mesenchymal Stem Cells Prevent Progressive Experimental Renal Failure but Maldifferentiate into Glomerular Adipocytes. J. Am. Soc. Nephrol. 2007, 18, 1754–1764. [Google Scholar] [CrossRef] [Green Version]
- Smeets, B.; Angelotti, M.L.; Rizzo, P.; Dijkman, H.; Lazzeri, E.; Mooren, F.; Ballerini, L.; Parente, E.; Sagrinati, C.; Mazzinghi, B.; et al. Renal Progenitor Cells Contribute to Hyperplastic Lesions of Podocytopathies and Crescentic Glomerulonephritis. J. Am. Soc. Nephrol. 2009, 20, 2593–2603. [Google Scholar] [CrossRef] [Green Version]
- Lasagni, L.; Romagnani, P. Glomerular Epithelial Stem Cells: The Good, the Bad, and the Ugly. J. Am. Soc. Nephrol. 2010, 21, 1612–1619. [Google Scholar] [CrossRef] [Green Version]
- Peired, A.; Lazzeri, E.; Lasagni, L.; Romagnani, P. Glomerular Regeneration: When Can the Kidney Regenerate from Injury and What Turns Failure into Success? Nephron Exp. Nephrol. 2014, 126, 70. [Google Scholar] [CrossRef]
- Rota, C.; Morigi, M.; Imberti, B. Stem Cell Therapies in Kidney Diseases: Progress and Challenges. Int. J. Mol. Sci. 2019, 20, E2790. [Google Scholar] [CrossRef] [Green Version]
- Habib, H.A.; Heeba, G.H.; Khalifa, M.M.A. Effect of Combined Therapy of Mesenchymal Stem Cells with GLP-1 Receptor Agonist, Exenatide, on Early-Onset Nephropathy Induced in Diabetic Rats. Eur. J. Pharmacol. 2021, 892, 173721. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Lv, S.; Liu, J.; Liu, S.; Wang, Y.; Liu, G. Mesenchymal Stem Cells Modified with Angiotensin-Converting Enzyme 2 Are Superior for Amelioration of Glomerular Fibrosis in Diabetic Nephropathy. Diabetes Res. Clin. Pract. 2020, 162, 108093. [Google Scholar] [CrossRef] [PubMed]
- Takemura, S.; Shimizu, T.; Oka, M.; Sekiya, S.; Babazono, T. Transplantation of Adipose-Derived Mesenchymal Stem Cell Sheets Directly into the Kidney Suppresses the Progression of Renal Injury in a Diabetic Nephropathy Rat Model. J. Diabetes Investig. 2020, 11, 545–553. [Google Scholar] [CrossRef] [Green Version]
- Bukulmez, H.; Horkayne-Szakaly, I.; Bilgin, A.; Baker, T.P.; Caplan, A.I.; Jones, O.Y. Intrarenal Injection of Mesenchymal Stem Cell for Treatment of Lupus Nephritis in Mice—A Pilot Study. Lupus 2021, 30, 52–60. [Google Scholar] [CrossRef]
- Becherucci, F.; Mazzinghi, B.; Allinovi, M.; Angelotti, M.L.; Romagnani, P. Regenerating the Kidney Using Human Pluripotent Stem Cells and Renal Progenitors. Expert Opin. Biol. Ther. 2018, 18, 795–806. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Sun, F.; Liu, J.; Ding, T.; She, J.; Mao, F.; Xu, W.; Qian, H.; Yan, Y. Emerging Role of Mesenchymal Stem Cell-Derived Exosomes in Regenerative Medicine. Curr. Stem Cell Res. Ther. 2019, 14, 482–494. [Google Scholar] [CrossRef]
- Bastos-Amador, P.; Royo, F.; Gonzalez, E.; Conde-Vancells, J.; Palomo-Diez, L.; Borras, F.E.; Falcon-Perez, J.M. Proteomic Analysis of Microvesicles from Plasma of Healthy Donors Reveals High Individual Variability. J. Proteom. 2012, 75, 3574–3584. [Google Scholar] [CrossRef]
- Börger, V.; Bremer, M.; Ferrer-Tur, R.; Gockeln, L.; Stambouli, O.; Becic, A.; Giebel, B. Mesenchymal Stem/Stromal Cell-Derived Extracellular Vesicles and Their Potential as Novel Immunomodulatory Therapeutic Agents. Int. J. Mol. Sci. 2017, 18, E1450. [Google Scholar] [CrossRef] [Green Version]
- Nargesi, A.A.; Lerman, L.O.; Eirin, A. Mesenchymal Stem Cell-Derived Extracellular Vesicles for Renal Repair. Curr. Gene Ther. 2017, 17, 29–42. [Google Scholar] [CrossRef]
- Wang, H.; Wang, B.; Zhang, A.; Hassounah, F.; Seow, Y.; Wood, M.; Ma, F.; Klein, J.D.; Price, S.R.; Wang, X.H. Exosome-Mediated MiR-29 Transfer Reduces Muscle Atrophy and Kidney Fibrosis in Mice. Mol. Ther. 2019, 27, 571–583. [Google Scholar] [CrossRef] [Green Version]
- Aghajani Nargesi, A.; Lerman, L.O.; Eirin, A. Mesenchymal Stem Cell-Derived Extracellular Vesicles for Kidney Repair: Current Status and Looming Challenges. Stem Cell Res. Ther. 2017, 8, 273. [Google Scholar] [CrossRef] [PubMed]
- Salih, M.; Zietse, R.; Hoorn, E.J. Urinary Extracellular Vesicles and the Kidney: Biomarkers and Beyond. Am. J. Physiol. Renal Physiol. 2014, 306, F1251–F1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimuccio, V.; Peruzzi, L.; Brizzi, M.F.; Cocchi, E.; Fop, F.; Boido, A.; Gili, M.; Gallo, S.; Biancone, L.; Camussi, G.; et al. Acute and Chronic Glomerular Damage Is Associated with Reduced CD133 Expression in Urinary Extracellular Vesicles. Am. J. Physiol. Renal Physiol. 2020, 318, F486–F495. [Google Scholar] [CrossRef] [PubMed]
- Lazzeri, E.; Romagnani, P.; Lasagni, L. Stem Cell Therapy for Kidney Disease. Expert Opin. Biol. Ther. 2015, 15, 1455–1468. [Google Scholar] [CrossRef]
- Packham, D.K.; Fraser, I.R.; Kerr, P.G.; Segal, K.R. Allogeneic Mesenchymal Precursor Cells (MPC) in Diabetic Nephropathy: A Randomized, Placebo-Controlled, Dose Escalation Study. EBioMedicine 2016, 12, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Maezawa, Y.; Takemoto, M.; Yokote, K. Cell Biology of Diabetic Nephropathy: Roles of Endothelial Cells, Tubulointerstitial Cells and Podocytes. J. Diabetes Investig. 2015, 6, 3–15. [Google Scholar] [CrossRef]
- Griffin, T.P.; Martin, W.P.; Islam, N.; O’Brien, T.; Griffin, M.D. The Promise of Mesenchymal Stem Cell Therapy for Diabetic Kidney Disease. Curr. Diab. Rep. 2016, 16, 42. [Google Scholar] [CrossRef]
- Peired, A.J.; Sisti, A.; Romagnani, P. Mesenchymal Stem Cell-Based Therapy for Kidney Disease: A Review of Clinical Evidence. Stem Cells Int. 2016, 2016, 4798639. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Chen, W.; Sun, L. An Update for Mesenchymal Stem Cell Therapy in Lupus Nephritis. Kidney Dis. 2021, 7, 79–89. [Google Scholar] [CrossRef]
- Al-Adhoubi, N.K.; Bystrom, J. Systemic Lupus Erythematosus and Diffuse Alveolar Hemorrhage, Etiology and Novel Treatment Strategies. Lupus 2020, 29, 355–363. [Google Scholar] [CrossRef]
- Shi, D.; Wang, D.; Li, X.; Zhang, H.; Che, N.; Lu, Z.; Sun, L. Allogeneic Transplantation of Umbilical Cord-Derived Mesenchymal Stem Cells for Diffuse Alveolar Hemorrhage in Systemic Lupus Erythematosus. Clin. Rheumatol. 2012, 31, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Gu, F.; Wang, H.; Hua, B.; Hou, Y.; Shi, S.; Lu, L.; Sun, L. Mesenchymal Stem Cell Transplantation for Diffuse Alveolar Hemorrhage in SLE. Nat. Rev. Rheumatol. 2010, 6, 486–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, G.; Isenberg, D.A. New Therapies for Systemic Lupus Erythematosus—Past Imperfect, Future Tense. Nat. Rev. Rheumatol. 2019, 15, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Gregorini, M.; Maccario, R.; Avanzini, M.A.; Corradetti, V.; Moretta, A.; Libetta, C.; Esposito, P.; Bosio, F.; Dal Canton, A.; Rampino, T. Antineutrophil Cytoplasmic Antibody-Associated Renal Vasculitis Treated with Autologous Mesenchymal Stromal Cells: Evaluation of the Contribution of Immune-Mediated Mechanisms. Mayo. Clin. Proc. 2013, 88, 1174–1179. [Google Scholar] [CrossRef] [Green Version]
- Appel, G.B.; Lau, W.L. Treatment of Rapidly Progressive Glomerulonephritis in the Elderly. Blood Purif. 2018, 45, 208–212. [Google Scholar] [CrossRef]
- Zhang, J.; Tao, X.; Sun, M.; Ying, R.; Su, W.; Wei, W.; Meng, X. A Rat Model of Radiation Vasculitis for the Study of Mesenchymal Stem Cell-Based Therapy. Biomed. Res. Int. 2019, 2019, 3727635. [Google Scholar] [CrossRef]
- Yin, W.; Ghebrehiwet, B.; Peerschke, E.I.B. Expression of Complement Components and Inhibitors on Platelet Microparticles. Platelets 2008, 19, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Trivioli, G.; Vaglio, A. The Rise of Complement in ANCA-Associated Vasculitis: From Marginal Player to Target of Modern Therapy. Clin. Exp. Immunol. 2020, 202, 403–406. [Google Scholar] [CrossRef]
- Grange, C.; Skovronova, R.; Marabese, F.; Bussolati, B. Stem Cell-Derived Extracellular Vesicles and Kidney Regeneration. Cells 2019, 8, E1240. [Google Scholar] [CrossRef] [Green Version]
- Al Mushafi, A.; Ooi, J.D.; Odobasic, D. Crescentic Glomerulonephritis: Pathogenesis and Therapeutic Potential of Human Amniotic Stem Cells. Front Physiol. 2021, 12, 724186. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Property | Effect | Study Type | Injury Type | EV-Carried RNA | References |
|---|---|---|---|---|---|
| Cytoprotection | Anti-apoptotic | In vivo (mice, rat) In vitro | I/R and drug-induced AKI; Unilateral ureteral obstruction-induced CKD | miR-21/-30/-125b/-130a/ 199a/-210/-223/-242 | [47,48] |
| Anti-necrotic | In vivo (mice) In vitro | Drug-induced AKI | CCNB1/CDK8/CDC6 mRNA HGF/IGF1 miR | [49] | |
| Anti-oxidant | In vivo (rat) In vitro | I/R and drug-induced AKI | [50,51,52] | ||
| Mitochondrial protection | In vivo (rat) | I/R and drug-induced AKI | miR-30 | [53] | |
| Autophagy stimulation | In vivo (rat) In vitro | Drug-induced AKI | miR-145 | [48,54,55] | |
| Anti-fibrotic | In vivo (mice, rat) In vitro | I/R-induced AKI; renovascular stenosis and unilateral ureteral obstruction-induced CKD; DN; 5/6 subtotal nephrectomy | miR-451a miR-let7c miR-133b-3p miR-294 miR-29b | [56,57,58,59,60,61,62,63] | |
| Cell regeneration | De-differentiation | In vivo (rat) In vitro | I/R-induced AKI | HGF mRNA | [64] |
| Proliferation | In vivo (rat) | I/R-induced AKI | HGF mRNA | [65] | |
| Angiogenesis | In vivo (rat) In vitro | I/R-induced AKI and CKD | miR-210 | [65,66] | |
| Immunomodulation | Anti-inflammation | In vivo (rat) | I/R and drug-induced AKI; renal artery stenosis; unilateral ureteral obstruction; 5/6 subtotal nephrectomy | miR-210/-378 miR-21 miR-199a | [67] |
| Macrophage inhibition and switch from M1 to M2 anti-inflammatory phenotype | In vivo (rat) In vitro | EVs released by LPS-preconditioned UC-MSC | miR-let7b | [68] | |
| Treg expansion and apoptosis of activated T cells | In vivo (canine) In vitro | Canine model | HGF, IL10, TGFβ, Indoleamine 2,3-dioxygenase, Prostaglandin E2, adenosine, PD-L1, Galectin-1 | [69,70,71] | |
| B lymphocyte inhibition | In vitro | CpG-stimulated PBMC coculture | [72,73] |
| MSC Type | Study Type | No. of pts (LN/SLE) | Median F/U (Months) | Main Results | References |
|---|---|---|---|---|---|
| Allogenic BM-MSCs | Pilot clinical study | 15/15 | 17,2 | MSC infusion safe and well tolerated. Decrease in SLEDAI and proteinuria. | [117] |
| Allogenic UC-MSCs | Clinical trial phase I/II | 15/16 | 8,25 | MSC infusion safe and well tolerated. No mortality reported. Improvement in SLEDAI, autoantibodies, serum albumin and complement C3 levels; increase in glomerular filtration rate and reduction in proteinuria. | [118] |
| Allogenic UC-MSCs | Multicenter clinical trial phase II | 38/40 | 12 | MSC infusion safe and well tolerated. No treatment related mortality. Significant decrease in SLEDAI. 32% and 28% of pts achieved complete and partial clinical response, respectively. 16% pts relapsed at 12 mo. | [120] |
| Allogenic BM-MSCs or UC-MSCs | Clinical trial phase II | 66/81 | 12 | No treatment-related mortality. Remission at 12 months: 60.5% Relapse: 22.4% Improvement in SLEDAI score, renal function, proteinuria | [122] |
| Allogenic UC-MSCs | RCT | 18/18 | 12 | MSC infusion safe and well tolerated. No difference in LN remission between MSC-treated and placebo. | [124] |
| Allogenic BM-MSCs Or UC-MSCs | Clinical trial phase II | 66/81 | 60 | Complete/partial remission: 27/7% Relapse: 24% 5-year mortality 16% | [119] |
| MSC/EV Type | Study Type | Experimental Model | Main Effects and Mechanisms | References |
|---|---|---|---|---|
| BM-MSCs | In vivo (mice) | STZ-induced type I C57BL/6 mice | Increase in β-pancreatic cells and reversal of hyperglycemia; improved mesangial expansion and glomerular hyalinosis | [133] |
| BM-MSCs | In vivo (mice) | STZ-induced type I NOD/SCID mice | Increase in β-pancreatic cells; decreased mesangial expansion, macrophage infiltration and GBM thickening | [134] |
| BM-MSCs | In vivo (rat) | STZ-induced type I Sprague Dawley rats | BM-MSC differentiate into islet-like cells through miR-124a; BM-MSC combined with miR-124a inhibit podocyte apoptosis | [135] |
| BM-MSCs | In vivo (rat) | STZ-induced type I Sprague Dawley rats | Reduced accumulation of ECM through restored fibrinolytic activity (decreases expression of PAI-1 and inhibition of TGFβ/Smad pathway); reduced renal fibrosis | [136] |
| BM-MSCs | In vivo (rat) | STZ-induced type I Sprague Dawley rats | Blunted diabetic glomerulosclerosis through inhibition of TGFβ/Smad pathway, reduced Lipoxin A4 and pro-inflammatory cytokines | [137] |
| BM-MSCs | In vivo (mice) | STZ-induced type I diabetes in BALB/c mice | Mitochondrial transfer of MSC to macrophages stimulated polarization towards M2 phenotype | [138] |
| BM-MSCs | In vivo (rat) | STZ-induced type I diabetes rats | Early administration prevents DN; systemic anti-inflammatory effect through increased EGF and IL 10 serum levels and downregulation of pro-inflammatory cytokines | [139] |
| BM-MSCs | In vivo (rat) | STZ-induced type I Sprague Dawley rats | Inhibition of intra-renal macrophage infiltration and oxidative stress through increased HGF and suppressed MCP-1 levels | [140] |
| BM-MSC-EVs | In vivo (mice) | STZ-induced type I diabetic mice and high-fat diet-induced type 2 diabetic mice | MSC-EVs preserved tight junctions in RTECs and exerted an anti-apoptotic effect | [141] |
| BM-MSC-EVs | In vivo (rat) | STZ-induced type I diabetic albino rats | Autophagy induction through mTOR inhibition and reduced renal fibrosis | [142] |
| BM-MSC-EVs | In vivo (rat) | STZ-induced type I Sprague Dawley rats | Inhibition of renal cell apoptosis through transfer of miR let7a | [143] |
| AD-MSCs | In vivo (rat) | STZ-induced type I Sprague Dawley rats | Anti-apoptotic effect; reduction of Wnt/βcatenin and elevation of Bcl-2 and klotho levels | [144] |
| AD-MSCs | In vivo (rat) | STZ-induced type I Sprague Dawley rats | Inhibition of oxidative stress and p38-MAPK signaling pathway | [145] |
| AD-MSCs | In vivo (rat) | STZ-induced type I Sprague Dawley rats | Attenuation of glomerular hypertrophy and tubule-interstitial damage in overt DN; downregulation of WT-1 and synaptopodin expression. | [146] |
| AD-MSC conditioned medium | In vitro | Hyperglycemia-injured podocytes | Protection of podocytes from hyperglycemia-induced apoptosis occurs via secretion of EGF by AD-MSCs. | [147] |
| AD-MSC-EVs | In vivo (mice) | Mouse podocytes and C57BL/KsJ db/db spontaneous diabetic mice | Protection of RTECs from apoptosis through interaction with TLR 4 and transfer of miR-26a-5p | [148] |
| Human UCB-MSCs | In vivo (rhesus macaque) | Non human primate model of early DN | Reduction in insulin requirement; anti-inflammatory effect in blood and in the kidney (reduced IL 16); improved histology and reduced in vitro sodium glucose cotransporter 2 expression in RTECs. | [149] |
| UCB-MSCs | In vivo (mice) | STZ-induced type I diabetic mice | Attenuation of podocyte injury through inhibition of hyperglycemia-activated TLR 2 and 4 signaling pathways | [150] |
| UCB-MSC conditioned medium | In vivo (rat) | STZ-induced type I Sprague Dawley rats | Reduction in mRNA expression of TGFβ-1, αSMA, collagen I and increased expression of E-cadherin and BMP7; inhibition of TGFβ-1-induced extracellular matrix upregulation and EMT. | [151] |
| UCB-MSC conditioned medium | In vivo (mice) | STZ-induced type I diabetic mice | Upregulation of MMP 2 and 9 in mesangial cells; inhibition of TGFβ-1-induced myofibroblast differentiation | [152] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quaglia, M.; Merlotti, G.; Fornara, L.; Colombatto, A.; Cantaluppi, V. Extracellular Vesicles Released from Stem Cells as a New Therapeutic Strategy for Primary and Secondary Glomerulonephritis. Int. J. Mol. Sci. 2022, 23, 5760. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105760
Quaglia M, Merlotti G, Fornara L, Colombatto A, Cantaluppi V. Extracellular Vesicles Released from Stem Cells as a New Therapeutic Strategy for Primary and Secondary Glomerulonephritis. International Journal of Molecular Sciences. 2022; 23(10):5760. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105760
Chicago/Turabian StyleQuaglia, Marco, Guido Merlotti, Laura Fornara, Andrea Colombatto, and Vincenzo Cantaluppi. 2022. "Extracellular Vesicles Released from Stem Cells as a New Therapeutic Strategy for Primary and Secondary Glomerulonephritis" International Journal of Molecular Sciences 23, no. 10: 5760. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105760