
Proteomics and Phosphoproteomics of Circulating Extracellular Vesicles Provide New Insights into Diabetes Pathobiology

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Study Design and Clinical Characteristics of the Study Cohort
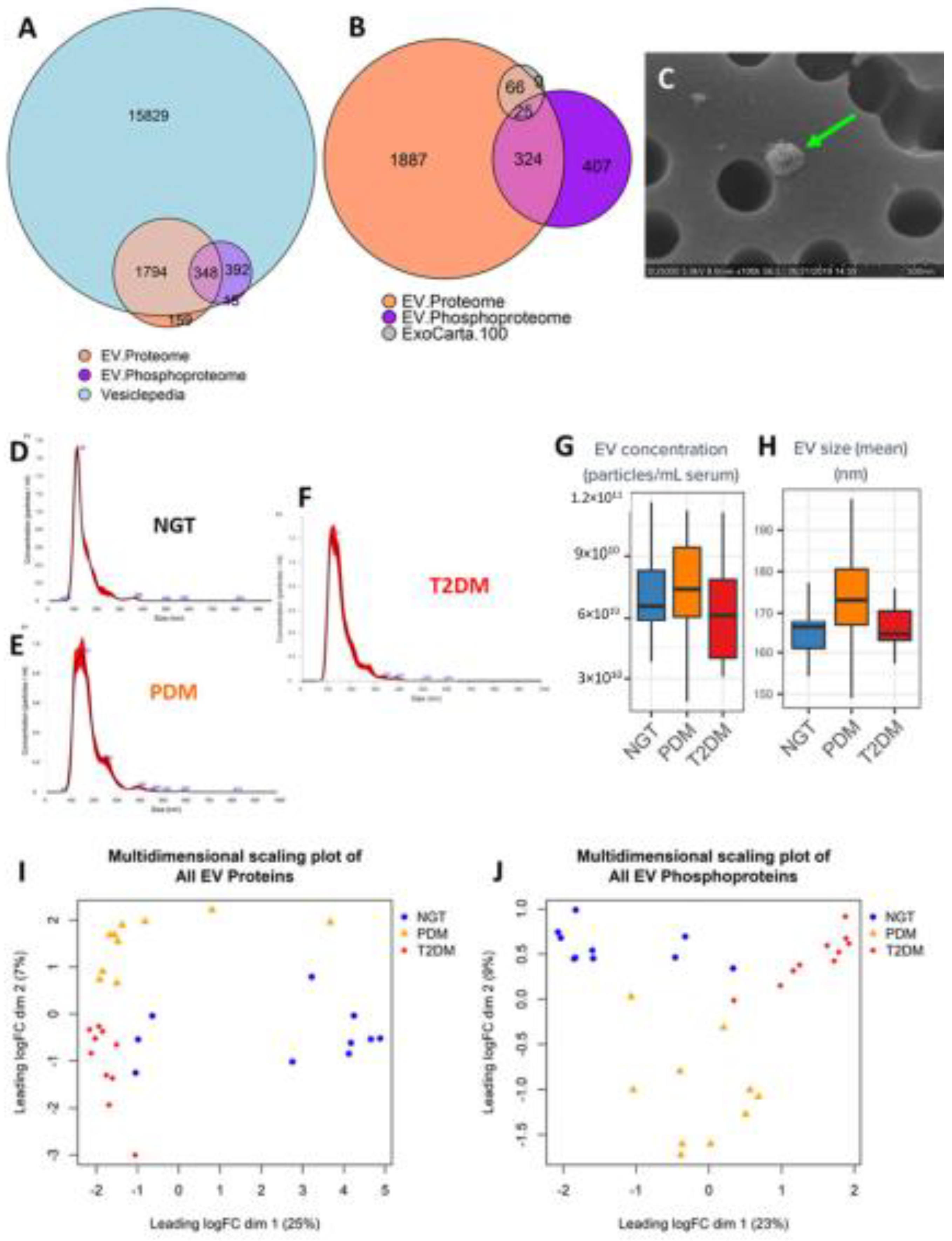
2.2. EVs from Serum Are Highly Enriched with Exosomal Proteins and Phosphoproteins
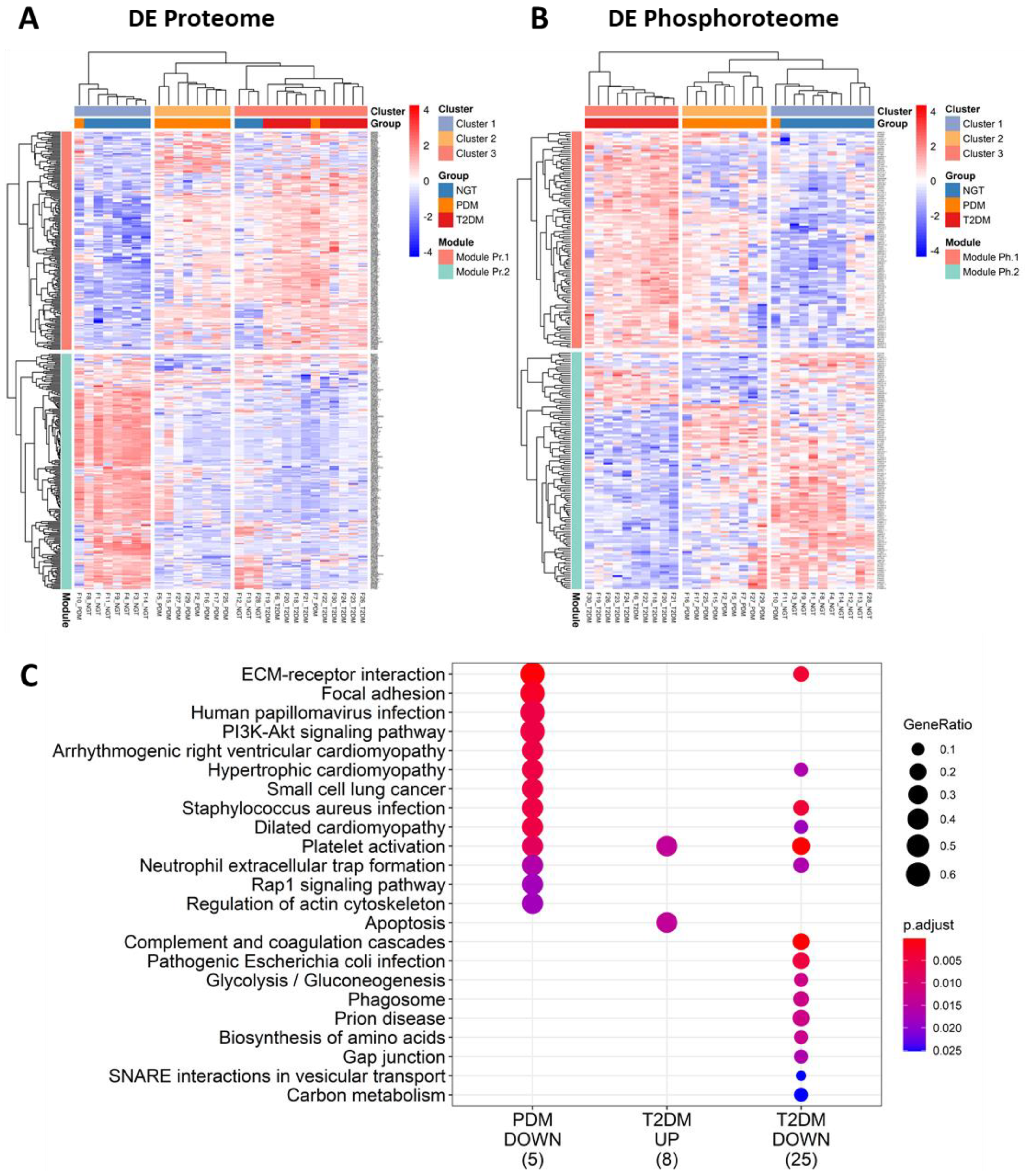
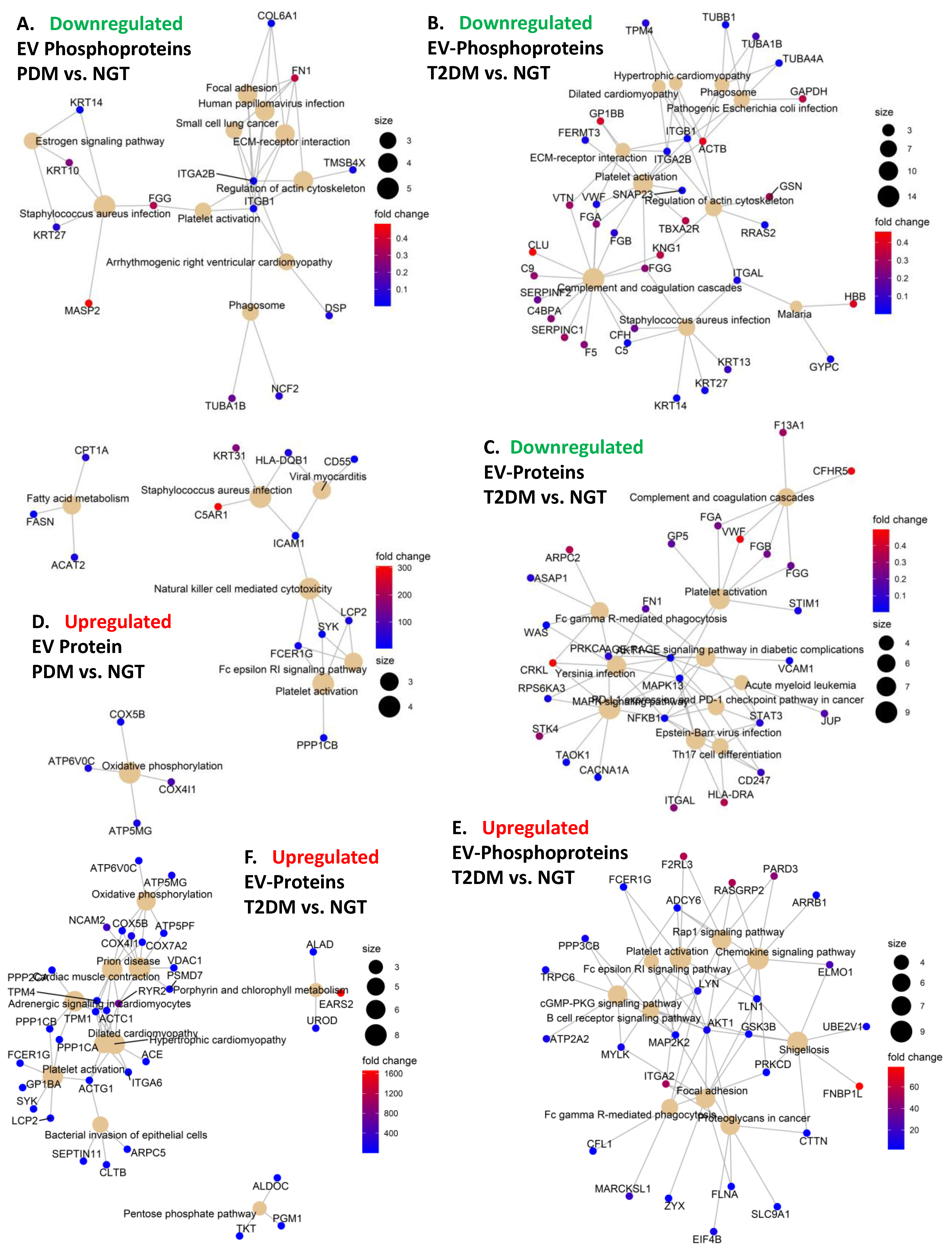
2.3. Differential Expression Analysis Provides Insight on Potential Tissue-Specific Mechanisms
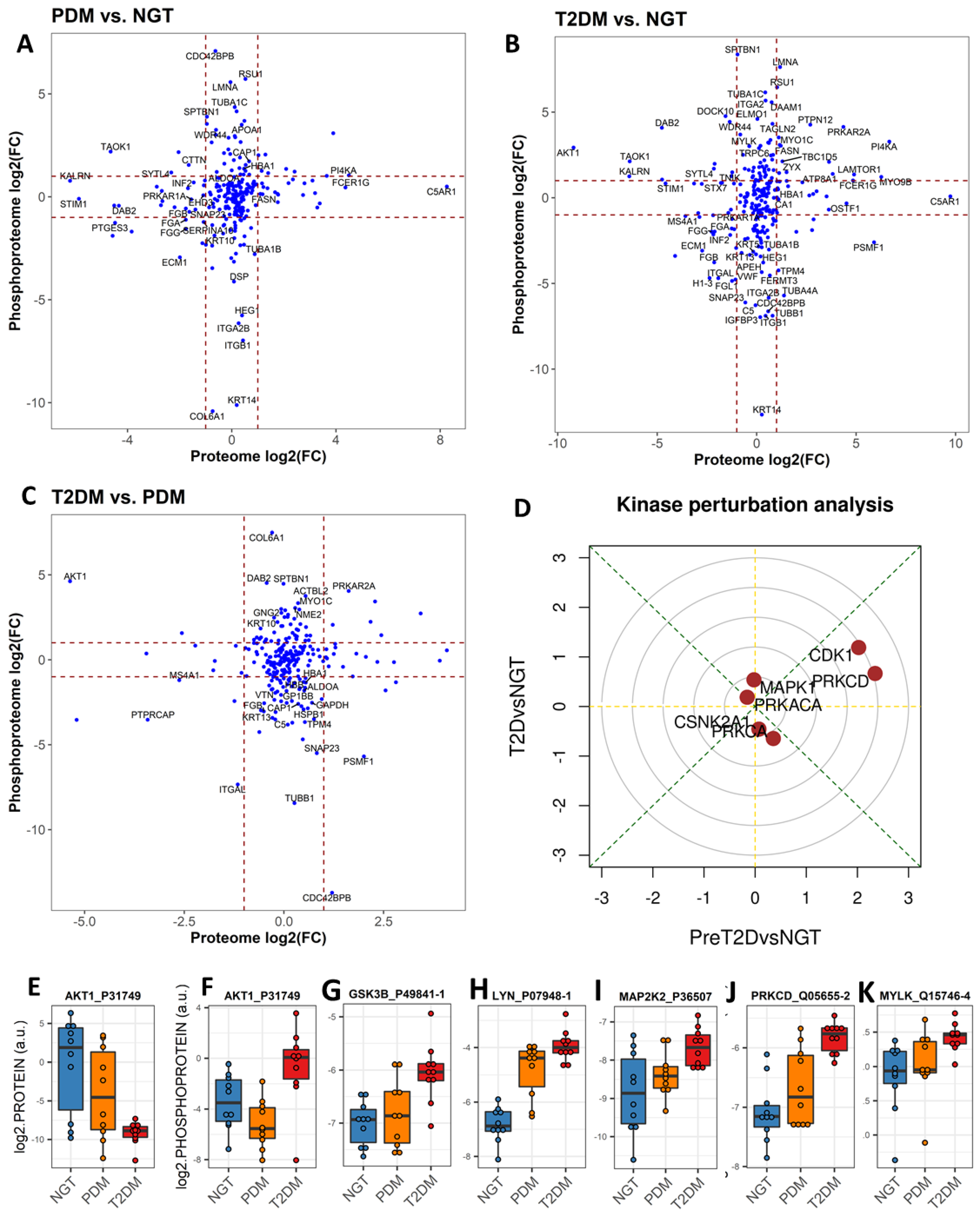
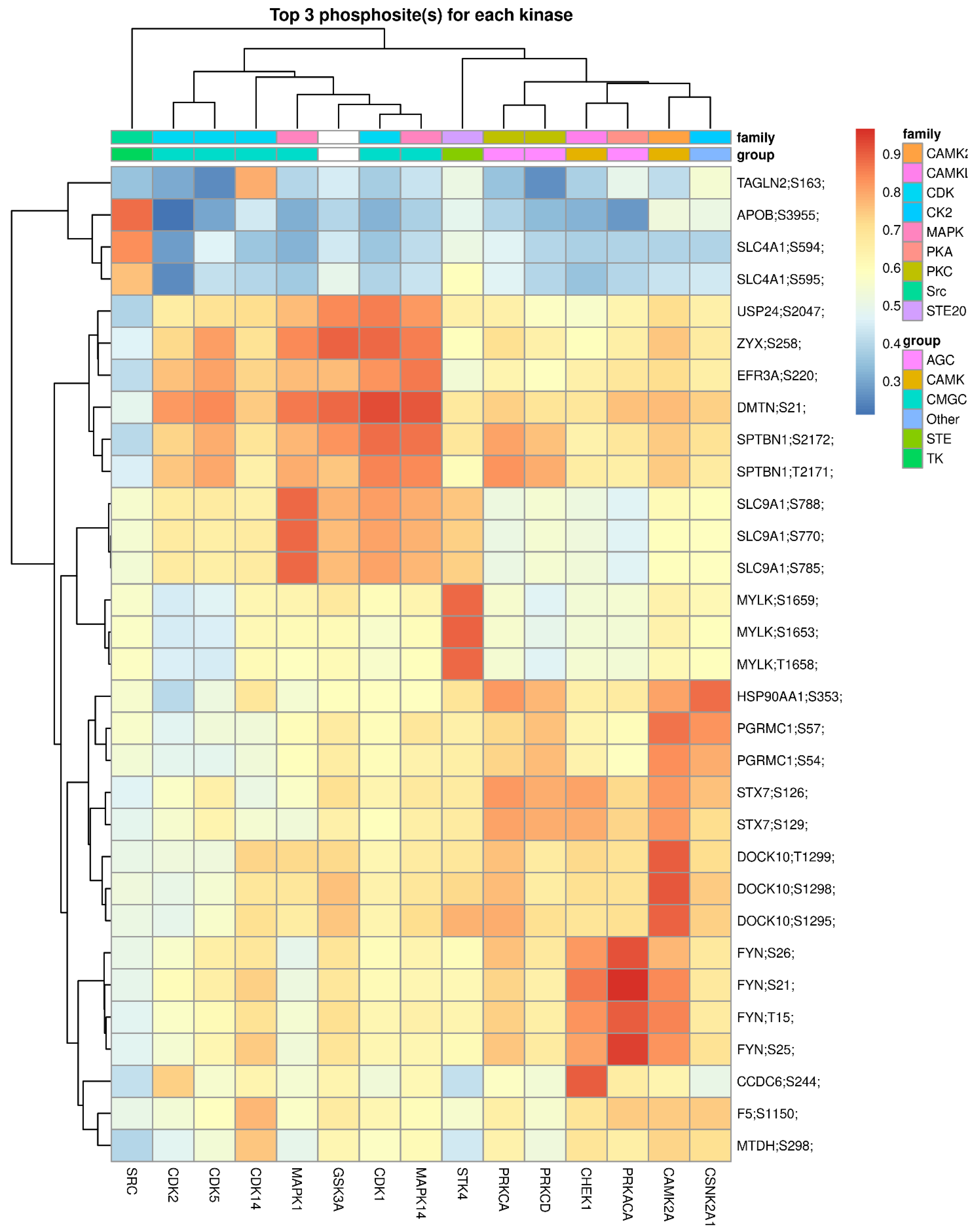
2.4. Phosphosite-Centric Analyses Identify Upstream Kinases Driving Phosphoproteomic Signatures in PDM and T2DM
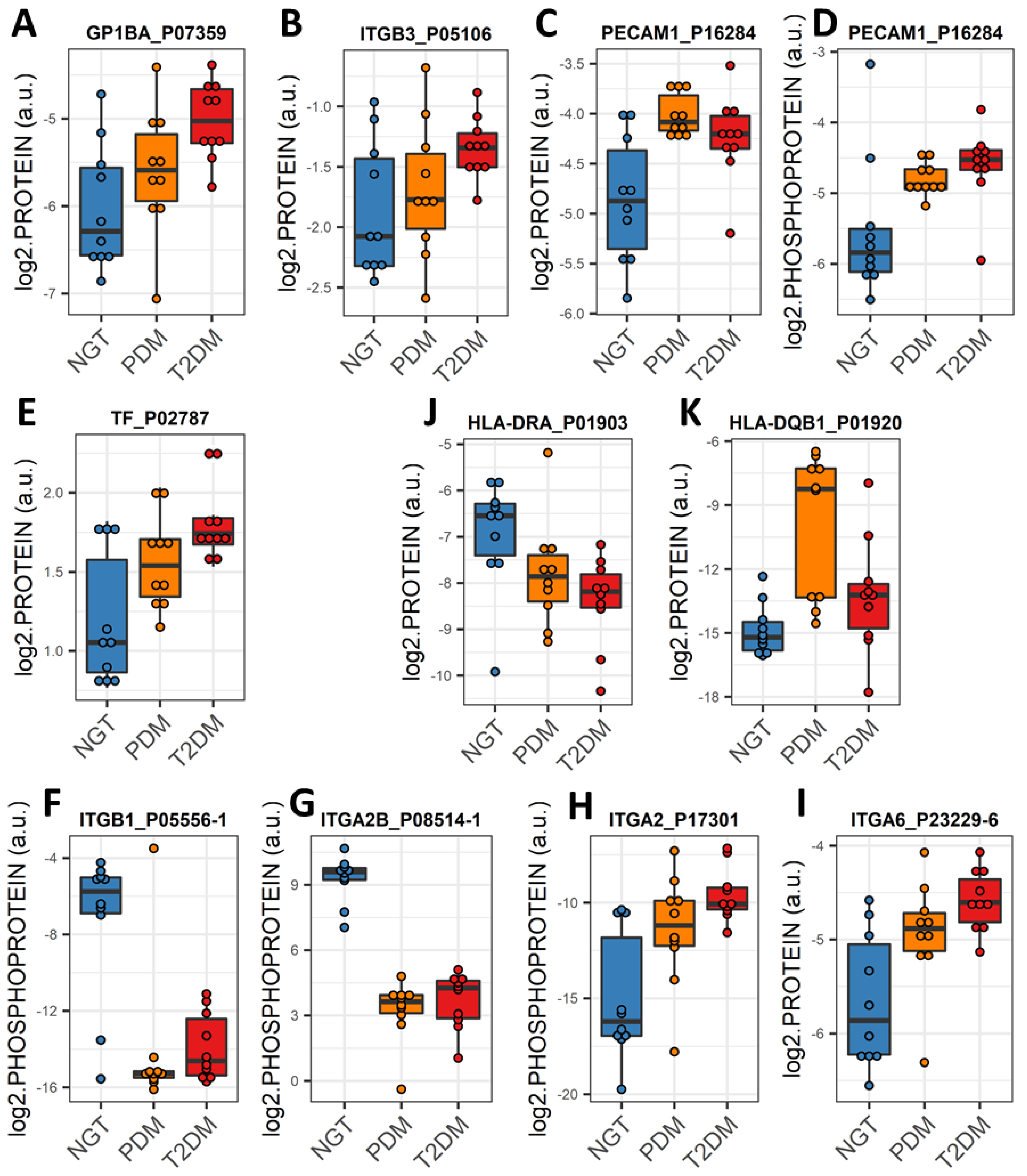
2.5. Altered Expression of Platelet and Immune Activation and Coagulation Markers in Circulating EVs Is Common in PDM and T2DM
2.6. An “Integrin Switching” Signature in Circulating EVs Is Characteristic of PDM and T2DM
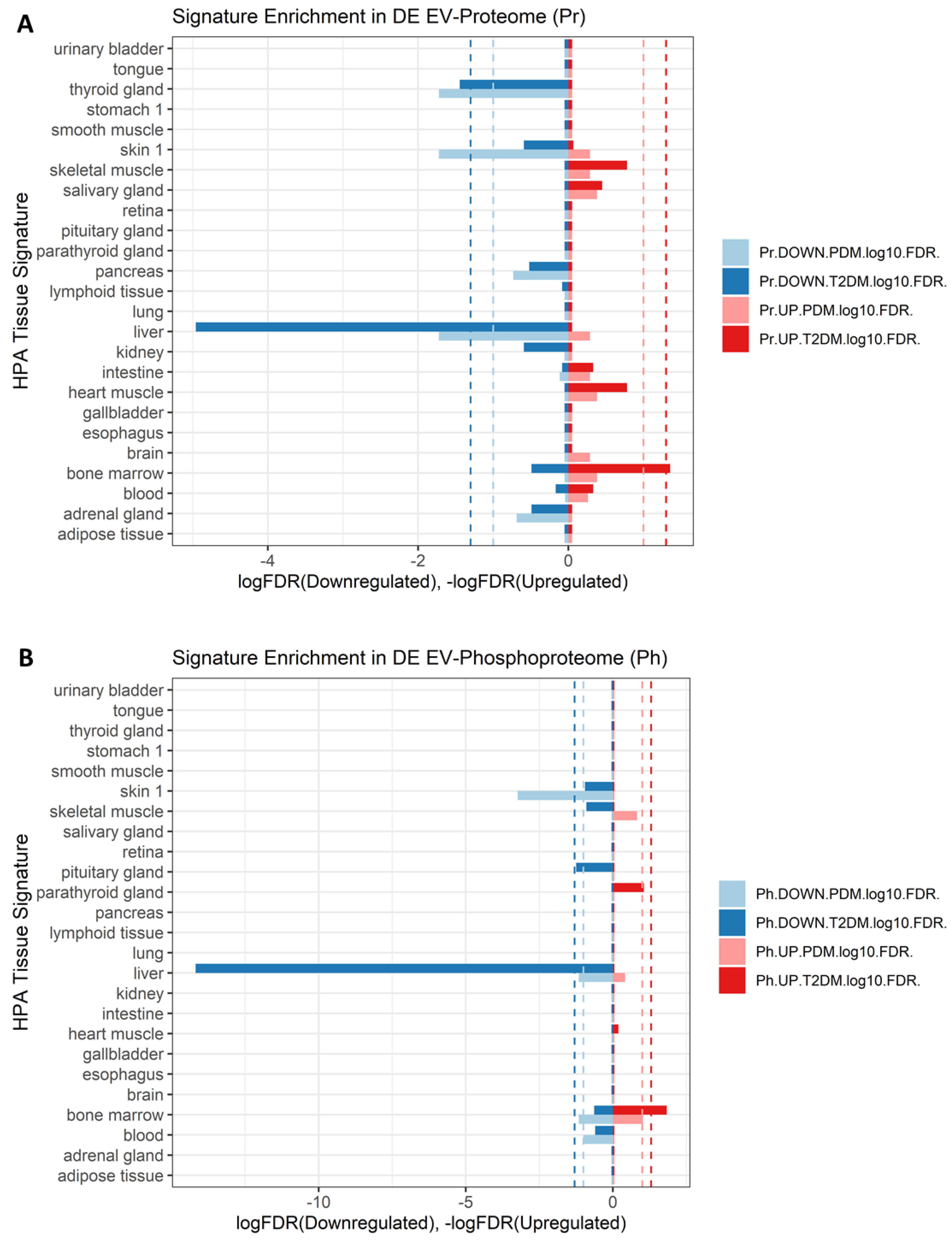
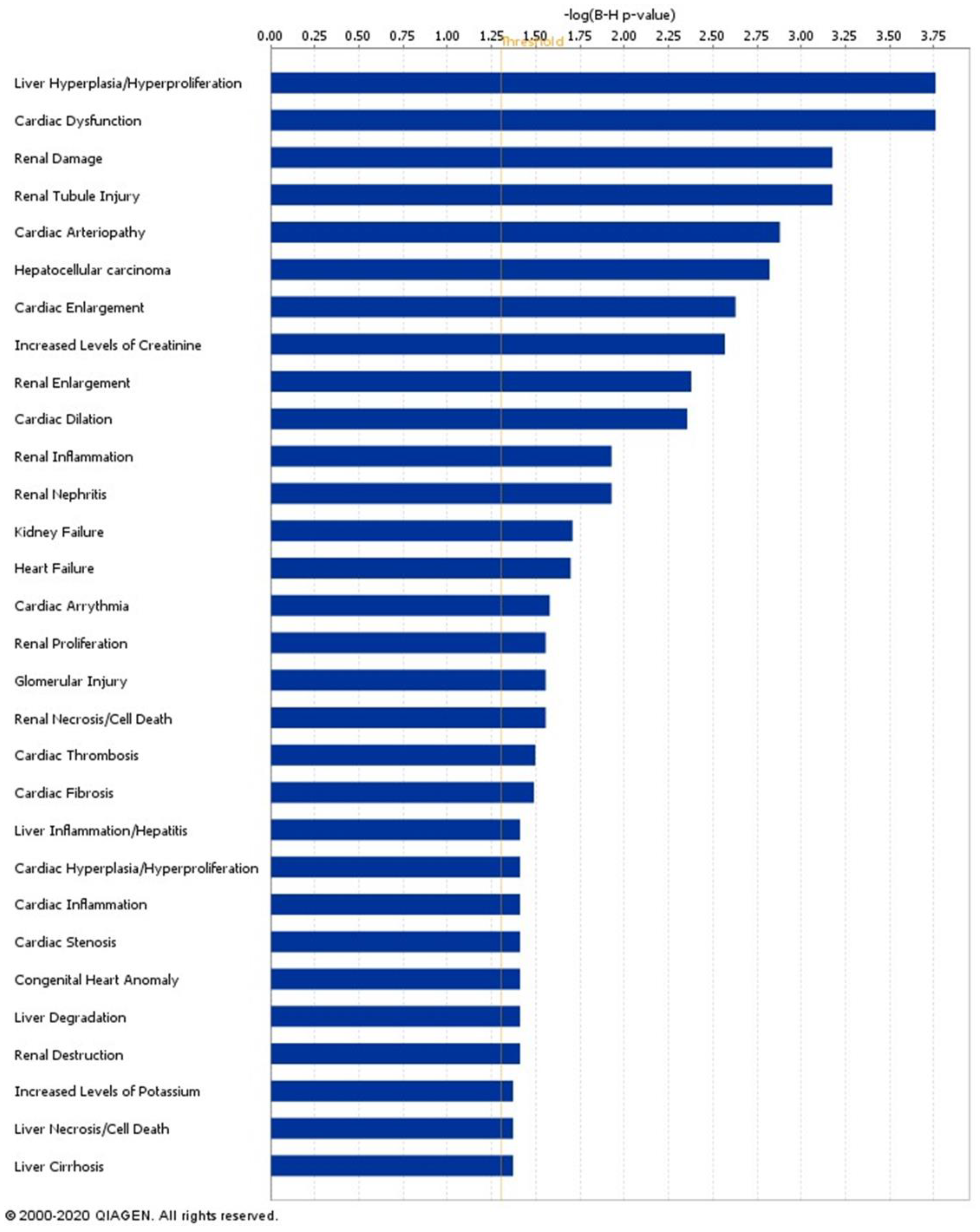
2.7. Signatures of Liver Proteins and Phosphoproteins Are Downregulated in EVs as Early as the Prediabetes Stage
3. Discussion
4. Materials and Methods
4.1. Samples
4.2. Clinical and Metabolic Measurements
4.3. EV Isolation
4.4. Mass Spectrometry (LC-MS/MS)-Based Methods Developed by Tymora Were Used to Detect the Global EV Proteome and Phosphoproteome
4.5. Bioinformatic Analysis of MS/MS Data
4.6. Nanoparticle Tracking Analysis (NTA)
4.7. Scanning Electron Microscopy (SEM)
4.8. Phosphosite-Centric Analyses
4.9. Detection of Tissue-Specific Signatures and KEGG Pathway Enrichment Analysis
4.10. Downstream Effect Analysis (DEA)
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the In-ternational Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, E.Y.F.; Chin, W.Y.; Yu, E.Y.T.; Wong, I.C.K.; Chan, E.W.Y.; Li, S.X.; Cheung, N.K.L.; Wang, Y.; Lam, C.L.K. The Impact of Cardiovascular Disease and Chronic Kidney Disease on Life Expectancy and Direct Medical Cost in a 10-Year Diabetes Cohort Study. Diabetes Care 2020, 43, 1750–1758. [Google Scholar] [CrossRef] [PubMed]
- Tuomilehto, J.; Schwarz, P.; Lindström, J. Long-Term Benefits From Lifestyle Interventions for Type 2 Diabetes Prevention. Diabetes Care 2011, 34 (Suppl. 2), S210–S214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeFronzo, R.A.; Abdul-Ghani, M. Type 2 Diabetes Can Be Prevented With Early Pharmacological Intervention. Diabetes Care 2011, 34 (Suppl. 2), S202–S209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, E.R. Type 2 diabetes: A multifaceted disease. Diabetologia 2019, 62, 1107–1112. [Google Scholar] [CrossRef] [Green Version]
- Li, C.-J.; Fang, Q.-H.; Liu, M.-L.; Lin, J.-N. Current understanding of the role of Adipose-derived Extracellular Vesicles in Metabolic Homeostasis and Diseases: Communication from the distance between cells/tissues. Theranostics 2020, 10, 7422–7435. [Google Scholar] [CrossRef] [PubMed]
- Romero, A.; Eckel, J. Organ Crosstalk and the Modulation of Insulin Signaling. Cells 2021, 10, 2082. [Google Scholar] [CrossRef]
- Guay, C.; Regazzi, R. Exosomes as new players in metabolic organ cross-talk. Diabetes Obes. Metab. 2017, 19, 137–146. [Google Scholar] [CrossRef]
- Huang, Z.; Xu, A. Adipose Extracellular Vesicles in Intercellular and Inter-Organ Crosstalk in Metabolic Health and Diseases. Front. Immunol. 2021, 12, 608680. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, biologic function and clinical potential. Cell Biosci. 2019, 9, 19. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Bedina Zavec, A.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowal, J.; Arras, G.; Colombo, M.; Jouve, M.; Morath, J.P.; Primdal-Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Théry, C. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc. Natl. Acad. Sci. USA 2016, 113, E968–E977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boukouris, S.; Mathivanan, S. Exosomes in bodily fluids are a highly stable resource of disease biomarkers. Proteom. Clin. Appl. 2015, 9, 358–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iliuk, A.B.; Tao, W.A. Is phosphoproteomics ready for clinical research? Clin. Chim. Acta 2013, 420, 23–27. [Google Scholar] [CrossRef] [Green Version]
- Andaluz Aguilar, H.A.; Iliuk, A.B.; Chen, I.-H.; Tao, W.A. Sequential phosphoproteomics and N-glycoproteomics of plasma-derived extracellular vesicles. Nat. Protoc. 2019, 15, 161–180. [Google Scholar] [CrossRef]
- Iliuk, A.; Wu, X.; Li, L.; Sun, J.; Hadisurya, M.; Boris, R.S.; Tao, W.A. Plasma-Derived Extracellular Vesicle Phosphoproteomics through Chemical Affinity Purification. J. Proteome Res. 2020, 19, 2563–2574. [Google Scholar] [CrossRef]
- Wu, X.; Li, L.; Iliuk, A.; Tao, W.A. Highly Efficient Phosphoproteome Capture and Analysis from Urinary Extracellular Vesicles. J. Proteome Res. 2018, 17, 3308–3316. [Google Scholar] [CrossRef]
- Chen, I.-H.; Xue, L.; Hsu, C.-C.; Paez, J.S.P.; Pan, L.; Andaluz, H.; Wendt, M.K.; Iliuk, A.B.; Zhu, J.-K.; Tao, W.A. Phosphoproteins in extracellular vesicles as candidate markers for breast cancer. Proc. Natl. Acad. Sci. USA 2017, 114, 3175–3180. [Google Scholar] [CrossRef] [Green Version]
- Hooten, N.N.; Evans, M.K. Extracellular vesicles as signaling mediators in type 2 diabetes mellitus. Am. J. Physiol. Physiol. 2020, 318, C1189–C1199. [Google Scholar] [CrossRef]
- Guay, C.; Menoud, V.; Rome, S.; Regazzi, R. Horizontal transfer of exosomal microRNAs transduce apoptotic signals between pancreatic beta-cells. Cell Commun. Signal. 2015, 13, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Freeman, D.W.; Noren Hooten, N.N.; Eitan, E.; Green, J.; Mode, N.A.; Bodogai, M.; Zhang, Y.; Lehrmann, E.; Zonderman, A.B.; Biragyn, A.; et al. Altered Extracellular Vesicle Concentration, Cargo, and Function in Diabetes. Diabetes 2018, 67, 2377–2388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Liu, J.; Qu, D.; Wang, L.; Wong, C.M.; Lau, C.-W.; Huang, Y.; Wang, Y.F.; Huang, H.; Xia, Y.; et al. Serum exosomes mediate delivery of arginase 1 as a novel mechanism for endothelial dysfunction in diabetes. Proc. Natl. Acad. Sci. USA 2018, 115, E6927–E6936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, M.; Wen, J.; Ma, Z.; Xiao, L.; Liu, Y.; Kwon, S.; Liu, Y.; Dong, Z. FOXO1-Mediated Downregulation of RAB27B Leads to Decreased Exosome Secretion in Diabetic Kidneys. Diabetes 2021, 70, 1536–1548. [Google Scholar] [CrossRef] [PubMed]
- Prattichizzo, F.; De Nigris, V.; Sabbatinelli, J.; Giuliani, A.; Castaño, C.; Párrizas, M.; Crespo, I.; Grimaldi, A.; Baranzini, N.; Spiga, R.; et al. CD31+ Extracellular Vesicles From Patients With Type 2 Diabetes Shuttle a miRNA Signature Associated With Cardiovascular Complications. Diabetes 2020, 70, 240–254. [Google Scholar] [CrossRef]
- Prattichizzo, F.; Matacchione, G.; Giuliani, A.; Sabbatinelli, J.; Olivieri, F.; de Candia, P.; De Nigris, V.; Ceriello, A. Extracellular vesicle-shuttled miRNAs: A critical appraisal of their potential as nano-diagnostics and nano-therapeutics in type 2 diabetes mellitus and its cardiovascular complications. Theranostics 2021, 11, 1031–1045. [Google Scholar] [CrossRef]
- Nunez Lopez, Y.O.; Garufi, G.; Seyhan, A.A. Altered levels of circulating cytokines and microRNAs in lean and obese individuals with prediabetes and type 2 diabetes. Mol. BioSyst. 2016, 13, 106–121. [Google Scholar] [CrossRef]
- Yang, P.; Patrick, E.; Humphrey, S.; Ghazanfar, S.; James, D.; Jothi, R.; Yang, J.Y.H. KinasePA: Phosphoproteomics data annotation using hypothesis driven kinase perturbation analysis. Proteomics 2016, 16, 1868–1871. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, T.; Xiao, D.; Yang, P. Protocol for the processing and downstream analysis of phosphoproteomic data with PhosR. STAR Protoc. 2021, 2, 100585. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, T.; Hoffman, N.J.; Xiao, D.; James, D.E.; Humphrey, S.J.; Yang, P. PhosR enables processing and functional analysis of phosphoproteomic data. Cell Rep. 2021, 34, 108771. [Google Scholar] [CrossRef]
- Kilanowska, A.; Ziółkowska, A. Apoptosis in Type 2 Diabetes: Can It Be Prevented? Hippo Pathway Prospects. Int. J. Mol. Sci. 2022, 23, 636. [Google Scholar] [CrossRef]
- Grover, S.P.; Mackman, N. Tissue Factor. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 709–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sliker, B.H.; Goetz, B.T.; Barnes, R.; King, H.; Maurer, H.C.; Olive, K.P.; Solheim, J.C. HLA-B influences integrin beta-1 expression and pancreatic cancer cell migration. Exp. Cell Res. 2020, 390, 111960. [Google Scholar] [CrossRef] [PubMed]
- Krämer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]
- Apostolopoulou, M.; Mastrototaro, L.; Hartwig, S.; Pesta, D.; Straßburger, K.; de Filippo, E.; Jelenik, T.; Karusheva, Y.; Gancheva, S.; Markgraf, D.; et al. Metabolic responsiveness to training depends on insulin sensitivity and protein content of exosomes in insulin-resistant males. Sci. Adv. 2021, 7, eabi9551. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, P.A.; Pisitkun, T.; Hoffert, J.D.; Tchapyjnikov, D.; Star, R.A.; Kleta, R.; Wang, N.S.; Knepper, M.A. Large-Scale Proteomics and Phosphoproteomics of Urinary Exosomes. J. Am. Soc. Nephrol. 2008, 20, 363–379. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Han, S.; Ma, L.; Zhang, H.; Zhan, Z.; Aguilar, H.A.; Zhang, H.; Xiao, K.; Gu, Y.; Gu, Z.; et al. Synergistically Bifunctional Paramagnetic Separation Enables Efficient Isolation of Urine Extracellular Vesicles and Downstream Phosphoproteomic Analysis. ACS Appl. Mater. Interfaces 2021, 13, 3622–3630. [Google Scholar] [CrossRef]
- Zheng, H.; Guan, S.; Wang, X.; Zhao, J.; Gao, M.; Zhang, X. Deconstruction of Heterogeneity of Size-Dependent Exosome Subpopulations from Human Urine by Profiling N-Glycoproteomics and Phosphoproteomics Simultaneously. Anal. Chem. 2020, 92, 9239–9246. [Google Scholar] [CrossRef]
- Huang, X.; Liu, G.; Guo, J.; Su, Z. The PI3K/AKT pathway in obesity and type 2 diabetes. Int. J. Biol. Sci. 2018, 14, 1483–1496. [Google Scholar] [CrossRef] [Green Version]
- Kelly, P.; Bailey, C.L.; Fueger, P.T.; Newgard, C.B.; Casey, P.; Kimple, M.E. Rap1 Promotes Multiple Pancreatic Islet Cell Functions and Signals through Mammalian Target of Rapamycin Complex 1 to Enhance Proliferation. J. Biol. Chem. 2010, 285, 15777–15785. [Google Scholar] [CrossRef] [Green Version]
- Shibasaki, T.; Takahashi, H.; Miki, T.; Sunaga, Y.; Matsumura, K.; Yamanaka, M.; Zhang, C.; Tamamoto, A.; Satoh, T.; Miyazaki, J.I.; et al. Essential role of Epac2/Rap1 signaling in regulation of insulin granule dynamics by cAMP. Proc. Natl. Acad. Sci. USA 2007, 104, 19333–19338. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, H.; Shibasaki, T.; Park, J.-H.; Hidaka, S.; Takahashi, T.; Ono, A.; Song, D.-K.; Seino, S. Role of Epac2A/Rap1 Signaling in Interplay Between Incretin and Sulfonylurea in Insulin Secretion. Diabetes 2014, 64, 1262–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camaya, I.; Donnelly, S.; O’Brien, B. Targeting the PI3K /Akt signaling pathway in pancreatic β-cells to enhance their survival and function: An emerging therapeutic strategy for type 1 diabetes. J. Diabetes 2022, 14, 247–260. [Google Scholar] [CrossRef] [PubMed]
- James, D.E.; Stöckli, J.; Birnbaum, M.J. The aetiology and molecular landscape of insulin resistance. Nat. Rev. Mol. Cell Biol. 2021, 22, 751–771. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-Y.; Deng, W.; Chen, Y.; Fan, W.; Baldwin, K.M.; Jope, R.S.; Wallace, D.C.; Wang, P.H. Impaired translocation and activation of mitochondrial Akt1 mitigated mitochondrial oxidative phosphorylation Complex V activity in diabetic myocardium. J. Mol. Cell. Cardiol. 2013, 59, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.-Y.; Yeh, H.-Y.; Lin, K.; Wang, P.H. Insulin stimulates Akt translocation to mitochondria: Implications on dysregulation of mitochondrial oxidative phosphorylation in diabetic myocardium. J. Mol. Cell. Cardiol. 2009, 46, 919–926. [Google Scholar] [CrossRef] [Green Version]
- Montagnani, M.; Chen, H.; Barr, V.A.; Quon, M.J. Insulin-stimulated activation of eNOS is independent of Ca2+ but requires phosphorylation by Akt at Ser1179. J. Biol. Chem. 2001, 276, 30392–30398. [Google Scholar] [CrossRef] [Green Version]
- Kuzmenko, D.I.; Udintsev, S.N.; Klimentyeva, T.K.; Serebrov, V.Y. Oxidative stress in adipose tissue as a primary link in pathogenesis of insulin resistance. Biochem. Mosc. Suppl. Ser. B Biomed. Chem. 2016, 10, 212–219. [Google Scholar] [CrossRef]
- Olsson, A.H.; Yang, B.T.; Hall, E.; Taneera, J.; Salehi, A.; Nitert, M.D.; Ling, C. Decreased expression of genes involved in oxidative phosphorylation in human pancreatic islets from patients with type 2 diabetes. Eur. J. Endocrinol. 2011, 165, 589–595. [Google Scholar] [CrossRef]
- Wang, M.; Wang, X.C.; Zhang, Z.Y.; Mou, B.; Hu, R.M. Impaired mitochondrial oxidative phosphorylation in multiple insulin-sensitive tissues of humans with type 2 diabetes mellitus. J. Int. Med Res. 2010, 38, 769–781. [Google Scholar]
- Mootha, V.K.; Handschin, C.; Arlow, D.; Xie, X.; St, P.J.; Sihag, S. Erralpha and Gabpa/b specify PGC-1alpha-dependent oxidative phosphorylation gene expression that is altered in diabetic muscle. Proc. Natl. Acad. Sci. USA 2004, 101, 6570–6575. [Google Scholar] [CrossRef] [Green Version]
- Mootha, V.K.; Lindgren, C.M.; Eriksson, K.-F.; Subramanian, A.; Sihag, S.; Lehar, J.; Puigserver, P.; Carlsson, E.; Ridderstråle, M.; Laurila, E.; et al. PGC-1α-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 2003, 34, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Ying, W.; Tseng, A.; Chang, R.C.-A.; Wang, H.; Lin, Y.-L.; Kanameni, S.; Brehm, T.; Morin, A.; Jones, B.; Splawn, T.; et al. miR-150 regulates obesity-associated insulin resistance by controlling B cell functions. Sci. Rep. 2016, 6, 20176. [Google Scholar] [CrossRef] [PubMed]
- Winer, D.A.; Winer, S.; Shen, L.; Wadia, P.P.; Yantha, J.; Paltser, G.; Tsui, H.; Wu, P.; Davidson, M.G.; Alonso, M.N.; et al. B cells promote insulin resistance through modulation of T cells and production of pathogenic IgG antibodies. Nat. Med. 2011, 17, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Winer, D.A.; Winer, S.; Shen, L.; Chng, M.H.Y.; Engleman, E.G. B lymphocytes as emerging mediators of insulin resistance. Int. J. Obes. Suppl. 2012, 2, S4–S7. [Google Scholar] [CrossRef]
- Gregg, T.; Sdao, S.M.; Dhillon, R.S.; Rensvold, J.W.; Lewandowski, S.L.; Pagliarini, D.J.; Denu, J.M.; Merrins, M.J. Obesity-dependent CDK1 signaling stimulates mitochondrial respiration at complex I in pancreatic β-cells. J. Biol. Chem. 2019, 294, 4656–4666. [Google Scholar] [CrossRef] [Green Version]
- Sdao, S.M.; Ho, T.; Poudel, C.; Foster, H.R.; De Leon, E.R.; Adams, M.T.; Lee, J.-H.; Blum, B.; Rane, S.G.; Merrins, M.J. CDK2 limits the highly energetic secretory program of mature β cells by restricting PEP cycle-dependent KATP channel closure. Cell Rep. 2021, 34, 108690. [Google Scholar] [CrossRef]
- Yu, W.; Niwa, T.; Fukasawa, T.; Hidaka, H.; Senda, T.; Sasaki, Y.; Niki, I. Synergism of protein kinase A, protein kinase C, and myosin light-chain kinase in the secretory cascade of the pancreatic beta-cell. Diabetes 2000, 49, 945–952. [Google Scholar] [CrossRef] [Green Version]
- Bezy, O.; Tran, T.T.; Pihlajamäki, J.; Suzuki, R.; Emanuelli, B.; Winnay, J.; Mori, M.; Haas, J.; Biddinger, S.B.; Leitges, M.; et al. PKCδ regulates hepatic insulin sensitivity and hepatosteatosis in mice and humans. J. Clin. Investig. 2011, 121, 2504–2517. [Google Scholar] [CrossRef]
- Li, M.; Vienberg, S.G.; Bezy, O.; O’Neill, B.T.; Kahn, C.R. Role of PKCδ in Insulin Sensitivity and Skeletal Muscle Metabolism. Diabetes 2015, 64, 4023–4032. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Sasse, K.C.; Bayguinov, Y.; Ward, S.M.; Perrino, B.A. Contractile Protein Expression and Phosphorylation and Contractility of Gastric Smooth Muscles from Obese Patients and Patients with Obesity and Diabetes. J. Diabetes Res. 2018, 2018, 8743874. [Google Scholar] [CrossRef]
- Hu, W.; Feng, P. Myosin Light Chain Kinase Is Involved in the Mechanism of Gastrointestinal Dysfunction in Diabetic Rats. Am. J. Dig. Dis. 2012, 57, 1197–1202. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Rigor, R.R.; Pivetti, C.D.; Wu, M.H.; Yuan, S.Y. Myosin light chain kinase in microvascular endothelial barrier function. Cardiovasc. Res. 2010, 87, 272–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foley, J.; Conway, E.M. Cross Talk Pathways Between Coagulation and Inflammation. Circ. Res. 2016, 118, 1392–1408. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, R.B. Cytokine and Cytokine-Like Inflammation Markers, Endothelial Dysfunction, and Imbalanced Coagulation in Development of Diabetes and Its Complications. J. Clin. Endocrinol. Metab. 2009, 94, 3171–3182. [Google Scholar] [CrossRef] [Green Version]
- Aso, Y.; Yoshida, N.; Okumura, K.-I.; Wakabayashi, S.; Matsutomo, R.; Takebayashi, K.; Inukai, T. Coagulation and inflammation in overt diabetic nephropathy: Association with hyperhomocysteinemia. Clin. Chim. Acta 2004, 348, 139–145. [Google Scholar] [CrossRef]
- Hoshino, A.; Costa-Silva, B.; Shen, T.-L.; Rodrigues, G.; Hashimoto, A.; Mark, M.T.; Molina, H.; Kohsaka, S.; Di Giannatale, A.; Ceder, S.; et al. Tumour exosome integrins determine organotropic metastasis. Nature 2015, 527, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Wortzel, I.; Dror, S.; Kenific, C.M.; Lyden, D. Exosome-Mediated Metastasis: Communication from a Distance. Dev. Cell 2019, 49, 347–360. [Google Scholar] [CrossRef]
- Gahmberg, C.G.; Grönholm, M. How integrin phosphorylations regulate cell adhesion and signaling. Trends Biochem. Sci. 2021, 47, 265–278. [Google Scholar] [CrossRef]
- Zong, H.; Bastie, C.C.; Xu, J.; Fassler, R.; Campbell, K.P.; Kurland, I.J.; Pessin, J.E. Insulin resistance in striated muscle-specific integrin receptor beta1-deficient mice. J. Biol. Chem. 2009, 284, 4679–4688. [Google Scholar] [CrossRef] [Green Version]
- Kang, L.; Ayala, J.E.; Lee-Young, R.S.; Zhang, Z.; James, F.D.; Neufer, P.D.; Pozzi, A.; Zutter, M.M.; Wasserman, D.H. Diet-induced muscle insulin resistance is associated with extracellular matrix remodeling and interaction with integrin alpha2beta1 in mice. Diabetes 2011, 60, 416–426. [Google Scholar] [CrossRef] [Green Version]
- de Fougerolles, A.R.; Sprague, A.G.; Nickerson-Nutter, C.L.; Chi-Rosso, G.; Rennert, P.D.; Gardner, H.; Gotwals, P.J.; Lobb, R.R.; Koteliansky, V.E. Regulation of inflammation by collagen-binding integrins alpha1beta1 and alpha2beta1 in models of hy-persensitivity and arthritis. J. Clin. Investig. 2000, 105, 721–729. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Ramirez, N.E.; Yankeelov, T.E.; Li, Z.; Ford, L.E.; Qi, Y.; Pozzi, A.; Zutter, M.M. alpha2beta1 integrin expression in the tumor microenvironment enhances tumor angiogenesis in a tumor cell-specific manner. Blood 2008, 111, 1980–1988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roest, M.; Banga, J.D.; Grobbee, D.E.; de Groot, P.G.; Sixma, J.J.; Tempelman, M.J.; van der Schouw, Y.T. Homozygosity for 807 T polymorphism in alpha(2) subunit of platelet alpha(2)beta(1) is associated with increased risk of cardiovascular mortality in high-risk women. Circulation 2000, 102, 1645–1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wertheimer, E.; Taylor, S.I.; Tennenbaum, T. Insulin receptor regulation of cell surface integrins: A possible mechanism contrib-uting to the development of diabetic complications. Proc. Assoc. Am. Physicians 1998, 110, 333–339. [Google Scholar] [PubMed]
- Yu, L.; Su, Y.; Paueksakon, P.; Cheng, H.; Chen, X.; Wang, H.; Harris, R.C.; Zent, R.; Pozzi, A. Integrin alpha1/Akita double-knockout mice on a Balb/c background develop advanced features of human diabetic nephropathy. Kidney Int. 2012, 81, 1086–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, T.; Podestá, F.; Stepp, M.A.; Boeri, D.; Lorenzi, M. Integrin overexpression induced by high glucose and by human diabetes: Potential pathway to cell dysfunction in diabetic microangiopathy. Proc. Natl. Acad. Sci. USA 1993, 90, 9640–9644. [Google Scholar] [CrossRef] [Green Version]
- Lay, A.C.; Hale, L.J.; Stowell-Connolly, H.; Pope, R.J.; Nair, V.; Ju, W.; Marquez, E.; Rollason, R.; Hurcombe, J.A.; Hayes, B.; et al. IGFBP-1 expression is reduced in human type 2 diabetic glomeruli and modulates beta1-integrin/FAK signalling in human podocytes. Diabetologia 2021, 64, 1690–1702. [Google Scholar] [CrossRef]
- Kanasaki, K.; Kanda, Y.; Palmsten, K.; Tanjore, H.; Lee, S.B.; Lebleu, V.S.; Gattone, V.H.; Kalluri, R. Integrin β1-mediated matrix assembly and signaling are critical for the normal development and function of the kidney glomerulus. Dev. Biol. 2008, 313, 584–593. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.-C.; Fornoni, A.; Weins, A.; Hakroush, S.; Maiguel, D.; Sageshima, J.; Chen, L.; Ciancio, G.; Faridi, M.H.; Behr, D.; et al. Abatacept in B7-1–Positive Proteinuric Kidney Disease. N. Engl. J. Med. 2013, 369, 2416–2423. [Google Scholar] [CrossRef] [Green Version]
- Taylor, R. Pathogenesis of type 2 diabetes: Tracing the reverse route from cure to cause. Diabetologia 2008, 51, 1781–1789. [Google Scholar] [CrossRef] [Green Version]
- Taylor, R. Banting Memorial Lecture 2012 Reversing the twin cycles of Type 2 diabetes. Diabet. Med. 2013, 30, 267–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gastaldelli, A.; Cusi, K. From NASH to diabetes and from diabetes to NASH: Mechanisms and treatment options. JHEP Rep. 2019, 1, 312–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Targher, G.; Chonchol, M.; Zoppini, G.; Abaterusso, C.; Bonora, E. Risk of chronic kidney disease in patients with non-alcoholic fatty liver disease: Is there a link? J. Hepatol. 2011, 54, 1020–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alicic, R.Z.; Rooney, M.T.; Tuttle, K.R. Diabetic Kidney Disease: Challenges, Progress, and Possibilities. Clin. J. Am. Soc. Nephrol. CJASN 2017, 12, 2032–2045. [Google Scholar] [CrossRef] [PubMed]
- Le, M.H.; Yeo, Y.H.; Henry, L.; Nguyen, M.H. Nonalcoholic Fatty Liver Disease and Renal Function Impairment: A Cross-Sectional Population-Based Study on Its Relationship From 1999 to 2016. Hepatol. Commun. 2019, 3, 1334–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pais, R.; Bourron, O. Fatty liver and renal function impairment—Time for awareness? J. Hepatol. 2017, 68, 13–15. [Google Scholar] [CrossRef] [Green Version]
- Association, A.D. Standards of Medical Care in Diabetes—2014. Diabetes Care 2014, 37, S14–S80. [Google Scholar] [CrossRef] [Green Version]
- Ho, D.; Imai, K.; King, G.; Stuart, E.A. MatchIt: Nonparametric Preprocessing for Parametric Causal Inference. J. Stat. Softw. 2011, 42, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Welch, S.; Gebhart, S.S.P.; Bergman, R.N.; Phillips, L.S. Minimal Model Analysis of Intravenous Glucose Tolerance Test-Derived Insulin Sensitivity in Diabetic Subjects *. J. Clin. Endocrinol. Metab. 1990, 71, 1508–1518. [Google Scholar] [CrossRef]
- Pacini, G.; Bergman, R.N. MINMOD: A computer program to calculate insulin sensitivity and pancreatic responsivity from the frequently sampled intravenous glucose tolerance test. Comput. Methods Programs Biomed. 1986, 23, 113–122. [Google Scholar] [CrossRef]
- Masuda, T.; Tomita, M.; Ishihama, Y. Phase Transfer Surfactant-Aided Trypsin Digestion for Membrane Proteome Analysis. J. Proteome Res. 2008, 7, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Smyth, G.K. Linear Models and Empirical Bayes Methods for Assessing Differential Expression in Microarray Experiments. Stat. Appl. Genet. Mol. Biol. 2004, 3, 1–25. [Google Scholar] [CrossRef]
- van Ooijen, M.P.; Jong, V.L.; Eijkemans, M.J.; Heck, A.J.; Andeweg, A.C.; Binai, N.A.; van den Ham, H.J. Identification of differentially expressed peptides in high-throughput proteomics data. Brief. Bioinform. 2018, 19, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Hornbeck, P.V.; Kornhauser, J.M.; Latham, V.; Murray, B.; Nandhikonda, V.; Nord, A.; Skrzypek, E.; Wheeler, T.; Zhang, B.; Gnad, F. 15 years of PhosphoSitePlus(R): Integrating post-translationally modified sites, disease variants and isoforms. Nucleic Acids Res. 2019, 47, D433–D441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-Based Map of the Human Proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Pathan, M.; Fonseka, P.; Chitti, S.V.; Kang, T.; Sanwlani, R.; Van Deun, J.; Hendrix, A.; Mathivanan, S. Vesiclepedia 2019: A compendium of RNA, proteins, lipids and metabolites in extracellular vesicles. Nucleic Acids Res. 2018, 47, D516–D519. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy | PreT2D | T2D | p | |
|---|---|---|---|---|
| n | 10 | 10 | 10 | |
| Sex = Male (%) | 5 (50.0) | 5 (50.0) | 5 (50.0) | 1.000 |
| Age (years) | 45.9 (10.5) | 48.0 (7.8) | 50.7 (11.0) | 0.560 |
| BMI (kg/m2) | 33.1 (6.5) | 34.1 (5.6) | 34.7 (5.2) | 0.825 |
| Weight Average (kg) | 89.7 (16.7) | 101.0 (15.6) | 97.8 (21.4) | 0.368 |
| Height Average (cm) | 165.4 (7.8) | 172.8 (11.1) | 167.5 (9.6) | 0.223 |
| Waist Circumference (cm) | 99.6 (16.4) | 110.8 (15.8) | 109.63 (16.2) | 0.252 |
| DEXA Lean Mass (g) | 52,541.6 (8827.6) | 56,993.2 (12,429.7) | 54,111.7 (12,128.5) | 0.672 |
| DEXA Fat Mass (g) | 34,737.9 (16,103.2) | 41,785.8 (15,705.2) | 41,624.5 (12,800.2) | 0.494 |
| DEXA Fat Percentage (%) | 38.3 (14.0) | 41.7 (12.2) | 43.0 (9.3) | 0.672 |
| HDL-c (mg/dL) | 55.9 (11.8) | 42.2 (8.9) | 48.7 (16.6) | 0.076 |
| LDL-c (mg/dL) | 113.7 (26.5) | 113.6 (46.7) | 102.3 (37.0) | 0.741 |
| Triglycerides (mg/dL) | 105.0 (60.4) | 159.7 (134.1) | 139.5 (53.4) | 0.404 |
| TSH (mIU/L) | 1.6 (0.7) | 2.1 (1.1) | 2.4 (1.1) | 0.180 |
| Temperature (F) | 98.0 (0.3) | 97.9 (0.4) | 97.8 (0.2) | 0.605 |
| Respiration Rate (breaths per min) | 14.7 (2.1) | 15.5 (1.8) | 15.2 (1.9) | 0.646 |
| Systolic Blood Pressure (mmHg) | 124.4 (10.1) | 125.2 (10.4) | 126.0 (11.9) | 0.950 |
| Diastolic Blood Pressure (mmHg) | 78.30 (9.1) | 77.2 (9.9) | 79.80 (8.2) | 0.815 |
| Heart Rate (beats per min) | 63.4 (8.4) | 71.0 (11.5) | 67.6 (13.4) | 0.340 |
| HbA1C (%) | 5.4 [5.3, 5.5] | 5.9 [5.8, 6.1] | 6.3 [6.1, 7.0] | 0.001 |
| Glucose baseline (mg/dL) | 90.3 (6.7) | 100.6 (8.2) | 122.2 (20.3) | <0.001 |
| Insulin baseline (µIU/mL) | 3.6 (2.2) | 8.6 (6.8) | 5.8 (5.8) | 0.134 |
| Glucose AUC (mg/dL∙min) | 14,797.3 [13,302.1, 16,433.5] | 17,202.3 [16,488.6, 18,909.1] | 25,796.2 [23,196.5, 29,180.5] | <0.001 |
| Insulin AUC (µIU/mL∙min) | 3594.0 [2560.8, 7438.8] | 6271.0 [2964.6, 11,626.3] | 3463.6 [2254.2, 4915.9] | 0.237 |
| C-peptide AUC (ng/mL∙min) | 694.2 [569.6, 820.0] | 950.3 [573.5, 1262.8] | 816.4 [687.6, 887.7] | 0.783 |
| Insulinogenic Index (ΔIns0–30 min/ΔGlu0–30 min) | 0.9 (0.6) | 0.3 (3.0) | 0.2 (0.2) | 0.623 |
| HOMA-IR | 0.8 (0.5) | 2.1 (1.5) | 2.0 (2.2) | 0.155 |
| HOMA_B | 51.5 (36.4) | 96.0 (107.6) | 30.7 (22.5) | 0.102 |
| MATSUDA | 12.2 (9.4) | 6.3 (5.5) | 9.7 (8.9) | 0.276 |
| AIRg | 468.6 (349.8) | 549.32 (530.1) | 48.75 (64.6) | 0.041 |
| Si | 4.6 (4.5) | 2.7 (2.9) | 4.6 (2.6) | 0.434 |
| DI | 1150.7 (494.2) | 890.3 (569.4) | 249.5 (348.3) | 0.005 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nunez Lopez, Y.O.; Iliuk, A.; Petrilli, A.M.; Glass, C.; Casu, A.; Pratley, R.E. Proteomics and Phosphoproteomics of Circulating Extracellular Vesicles Provide New Insights into Diabetes Pathobiology. Int. J. Mol. Sci. 2022, 23, 5779. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105779
Nunez Lopez YO, Iliuk A, Petrilli AM, Glass C, Casu A, Pratley RE. Proteomics and Phosphoproteomics of Circulating Extracellular Vesicles Provide New Insights into Diabetes Pathobiology. International Journal of Molecular Sciences. 2022; 23(10):5779. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105779
Chicago/Turabian StyleNunez Lopez, Yury O., Anton Iliuk, Alejandra M. Petrilli, Carley Glass, Anna Casu, and Richard E. Pratley. 2022. "Proteomics and Phosphoproteomics of Circulating Extracellular Vesicles Provide New Insights into Diabetes Pathobiology" International Journal of Molecular Sciences 23, no. 10: 5779. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105779