
Molecular and Physiological Responses of Citrus sinensis Leaves to Long-Term Low pH Revealed by RNA-Seq Integrated with Targeted Metabolomics

 and
and 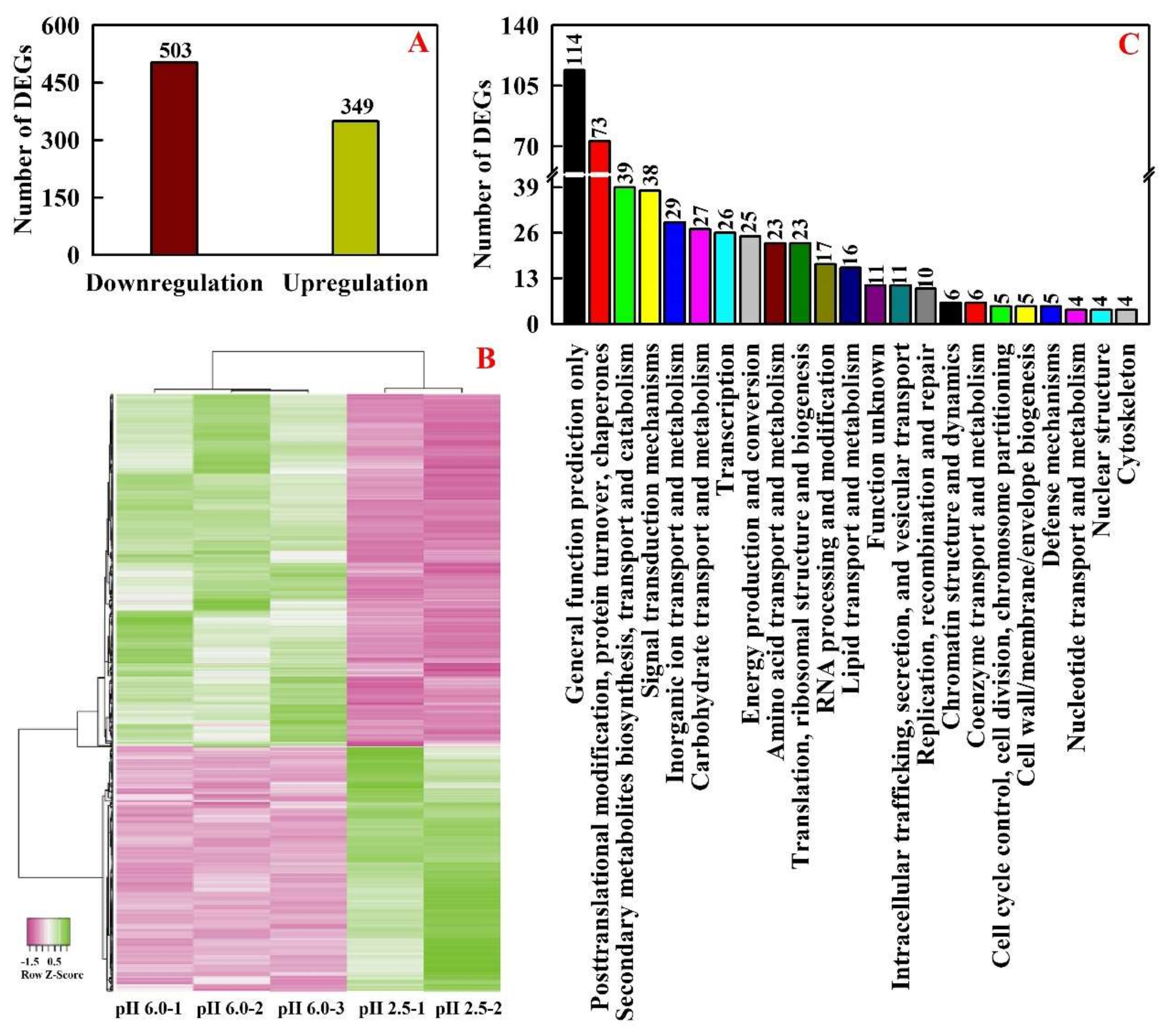{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. RNA-Seq, Mapping and Transcript Assembly in Leaves
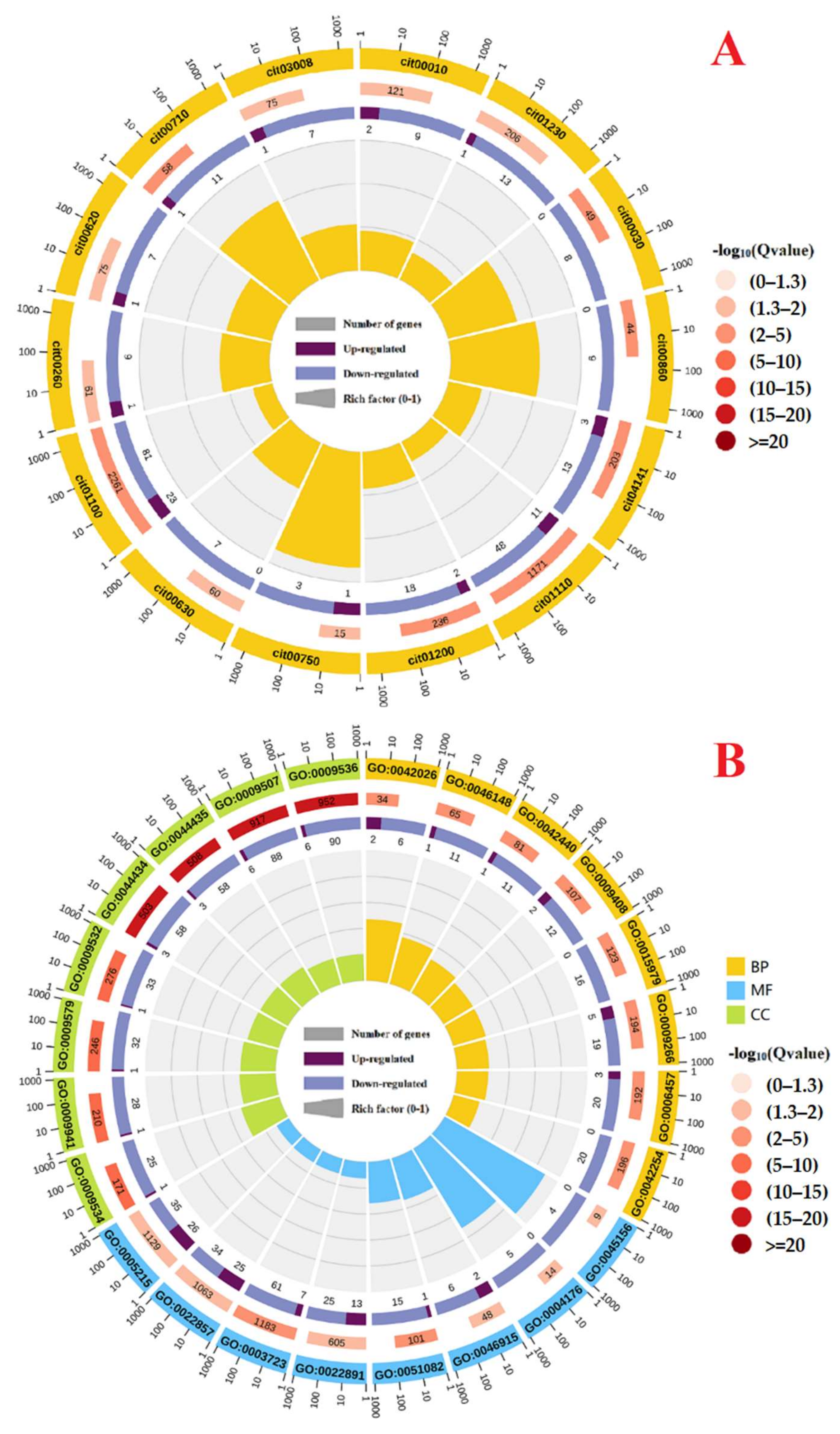
2.2. Low pH-Responsive Genes in Leaves
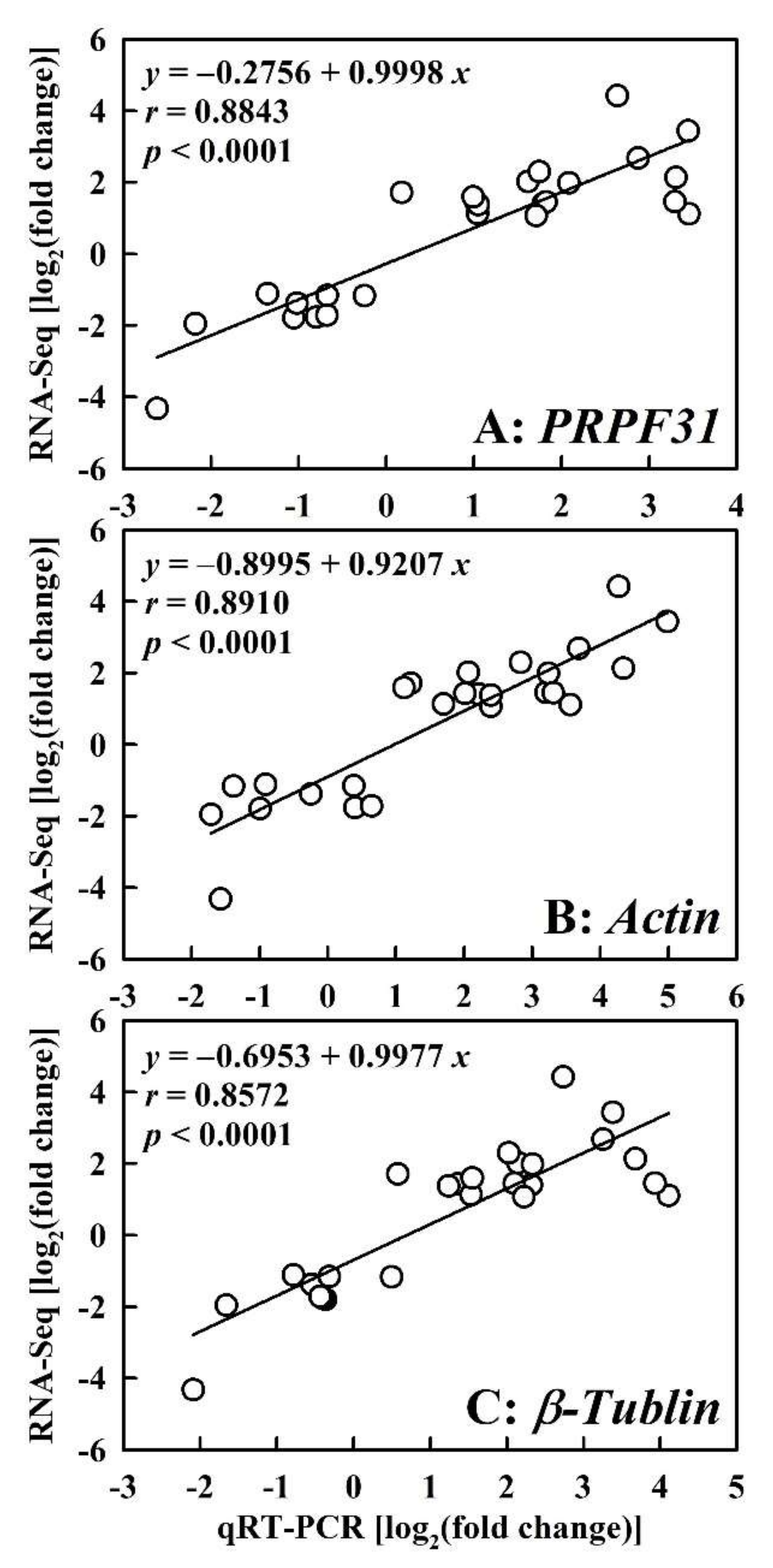
2.3. qRT-PCR Validation of DEGs
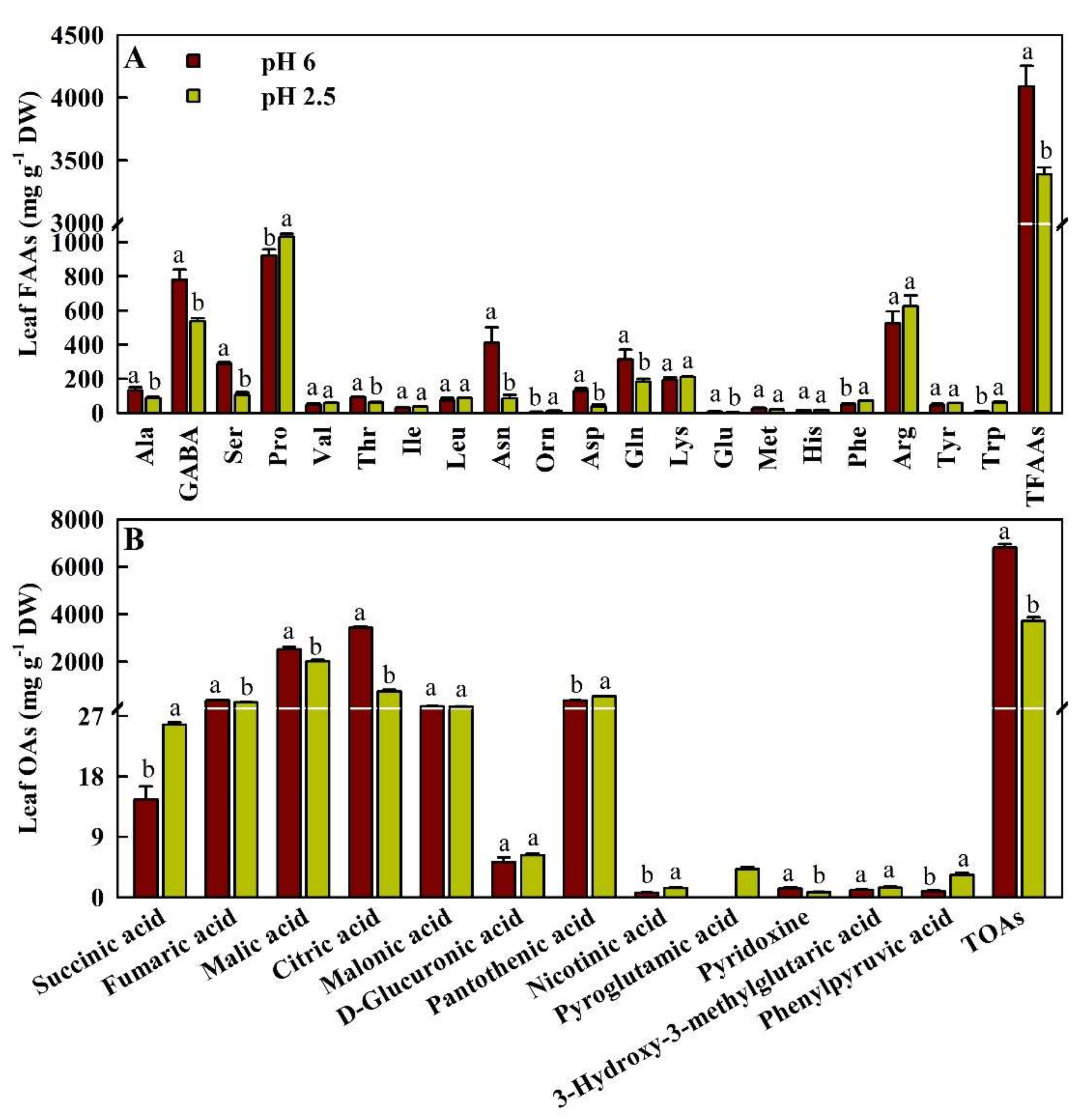
2.4. Effects of Low pH on the Concentrations of FAAs and OAs in Leaves
3. Discussion
3.1. Low pH Altered the Expression of Genes Related to Photosynthesis, Carbon, Carbohydrate, and Energy Metabolisms in Leaves
3.2. Low pH Altered the Expression of Genes Related to N, Protein, and AA Metabolisms in Leaves
3.3. Low Pi-Responsive Genes Displayed Adaptive Responses to Low pH in Leaves
3.4. Low pH Reduced SM Biosynthesis and Accumulation in Leaves
3.5. Low pH Broke the Balance between Production and Detoxification of ROS and Aldehydes, thus Leading to Oxidative Damage in Leaves
4. Materials and Methods
4.1. Seedling Culture
4.2. RNA Extraction, Library Construction, RNA-Seq, and Analysis
4.3. qRT-PCR Validation
4.4. Assay of OAs and FAAs in Leaves
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AAs | amino acids |
| ACO2 | CO2 assimilation |
| AER | alkenal reductase |
| AKR | aldo-keto reductase |
| ALDH | aldehyde dehydrogenase |
| APX | ascorbate peroxidase |
| ASC | ascorbate |
| BP | biological process |
| CRTISO | carotenoid isomerase |
| CC | cellular component |
| DAG | diacylglycerol |
| DAMs | differentially abundant metabolites |
| DEGs | differentially expressed genes |
| DHA | dehydroascorbate |
| FAAs | free amino acids |
| GSH | reduced glutathione |
| GSSG | oxidized glutathione |
| GST | glutathione S-transferase |
| HFAs | hydroxy fatty acids |
| MDAR | monodehydroascorbate reductase |
| MDHA | monodehydroascorbate |
| MF | molecular function |
| NA | nicotinic acid |
| NAM | nicotinamide |
| OAA | oxaloacetate |
| OAs | organic acids |
| PA | pantothenic acid |
| PAP | purple acid phosphatase |
| PC | phytochelatin |
| PCK | phosphoenolpyruvate carboxykinase |
| PDCT | phosphatidylcholine:diacylglycerol cholinephosphotransferase |
| PEP | phosphoenolpyruvate |
| PEPC | phosphoenolpyruvate carboxylase |
| PETC | photosynthetic electron transport chain |
| PG | pyroglutamic acid |
| PK | pyruvate kinase |
| PN | pyridoxine |
| PPi | pyrophosphate |
| PPA | phenylpyruvic acid |
| PPiase | pyrophosphatase |
| PPP | pentose phosphate pathway |
| PTM | posttranslational modification |
| ROS | reactive oxygen species |
| RWC | relative water content |
| SAMDC | S-adenosylmethionine decarboxylase |
| SOD | superoxide dismutase |
| TFAAs | total free amino acids |
| TOAs | total organic acids |
| TNC | total nonstructural carbohydrates |
References
- Shen, R.; Zhao, X. The sustainable use of acid soils. J. Agric. 2019, 9(3), 16–20. [Google Scholar]
- Guo, J.H.; Liu, X.J.; Zhang, Y.; Shen, J.L.; Han, W.X.; Zhang, W.F.; Christie, P.; Goulding, K.W.T.; Vitousek, P.M.; Zhang, F.S. Significant acidification in major Chinese croplands. Science 2010, 327, 1008–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Li, Q.; Qi, Y.-P.; Huang, W.-L.; Yang, L.-T.; Lai, L.-W.; Ye, X.; Chen, L.-S. Low pH-responsive proteins revealed by a 2-DE based MS approach and related physiological responses in Citrus leaves. BMC Plant Biol. 2018, 18, 188. [Google Scholar] [CrossRef] [PubMed]
- Long, A.; Zhang, J.; Yang, L.-T.; Ye, X.; Lai, N.-W.; Tan, L.-L.; Lin, D.; Chen, L.-S. Effects of low pH on photosynthesis, related physiological parameters, and nutrient profiles of Citrus. Front. Plant Sci. 2017, 8, 185. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Han, M.-Q.; Lin, F.; Ten, Y.; Lin, J.; Zhu, D.-H.; Guo, P.; Weng, Y.-B.; Chen, L.-S. Soil chemical properties, ‘Guanximiyou’ pummelo leaf mineral nutrient status and fruit quality in the southern region of Fujian province, China. J. Soil Sci. Plant Nutr. 2015, 15, 615–628. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Wu, W.; Zhao, T.; Tan, W.; Tian, J.; Liang, C. Complex gene regulation underlying mineral nutrient homeostasis in soybean root response to acidity stress. Genes 2019, 10, 402. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.-Y.; Cai, L.-Y.; Qi, Y.-P.; Yang, L.-T.; Lai, N.-W.; Chen, L.-S. Increasing nutrient solution pH alleviated aluminum-induced inhibition of growth and impairment of photosynthetic electron transport chain in Citrus sinensis seedlings. BioMed Res. Int. 2019, 2019, 9058715-17. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Tan, L.; Xu, Y.; Zhao, Y.; Cheng, F.; Ye, S.; Jiang, W. Effect of low pH and aluminum toxicity on the photosynthetic characteristics of different fast-growing Eucalyptus vegetatively propagated clones. PLoS ONE 2015, 10, e0130963. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.P.; Meng, P.; Li, J.Z.; Wan, X.C. Interactive effects of soil acidification and phosphorus deficiency on photosynthetic characteristics and growth in Juglans regia seedlings. Chin. J. Plant Ecol. 2014, 38, 1345–1355. [Google Scholar]
- Geng, G.; Wang, G.; Stevanato, P.; Lv, C.; Wang, Q.; Yu, L.; Wang, Y. Physiological and proteomic analysis of different molecular mechanisms of sugar beet response to acidic and alkaline pH environment. Front. Plant Sci. 2021, 12, 682799. [Google Scholar] [CrossRef]
- Long, A.; Huang, W.-L.; Qi, Y.-P.; Yang, L.-T.; Lai, N.-W.; Guo, J.-X.; Chen, L.-S. Low pH effects on reactive oxygen species and methylglyoxal metabolisms in Citrus roots and leaves. BMC Plant Biol. 2019, 19, 477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tóth, B.; Juhász, C.; Labuschagne, M.; Moloi, M.J. The influence of soil acidity on the physiological responses of two bread wheat cultivars. Plants 2020, 9, 1472. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.-Y.; Qi, Y.-P.; Huang, H.-Y.; Wu, F.-L.; Huang, W.-T.; Deng, C.-L.; Yang, L.-T.; Chen, L.S. Interactive effects of pH and aluminum on the secretion of organic acid anions by roots and related metabolic factors in Citrus sinensis roots and leaves. Environ. Pollut. 2020, 262, 114303. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, X.-F.; Huang, W.-T.; Chen, H.-H.; Lai, N.-W.; Yang, L.-T.; Huang, Z.-R.; Guo, J.; Ye, X.; Chen, L.-S. Mechanisms for increased pH-mediated amelioration of copper toxicity in Citrus sinensis leaves using physiology, transcriptomics and metabolomics. Environ. Exp. Bot. 2022, 196, 104812. [Google Scholar] [CrossRef]
- Rossini, B.C.; Bouvet, J.M.; Frouin, J.; Guerrini, I.A.; de Freitas, T.C.M.; da Silva, M.R.; Bouillet, J.-P.; Laclau, J.P.; Marino, C.L. Differential gene expression in Eucalyptus clones in response to nutrient deficiency. Tree Genet. Genomes 2022, 182, 1–17. [Google Scholar]
- Guo, P.; Qi, Y.-P.; Huang, W.-L.; Yang, L.-T.; Huang, Z.-R.; Lai, N.-W.; Chen, L.-S. Aluminum-responsive genes revealed by RNA-Seq and related physiological responses in leaves of two Citrus species with contrasting aluminum-tolerance. Ecotoxicol. Environ. Saf. 2018, 158, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Huang, H.; Peng, M.; Lai, Y.; Ren, Q.; Zhang, J.; Huang, Z.; Yang, L.; Rensing, C.; Chen, L. Adaptive Responses of Citrus grandis leaves to copper toxicity revealed by RNA-Seq and physiology. Int. J. Mol. Sci. 2021, 22, 12023. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.-T.; Pan, J.-F.; Hu, N.-J.; Chen, H.-H.; Jiang, H.-X.; Lu, Y.-B.; Chen, L.-S. Citrus physiological and molecular response to boron stresses. Plants 2022, 11, 40. [Google Scholar] [CrossRef]
- Wu, B.-S.; Zhang, J.; Huang, W.-L.; Yang, L.-T.; Huang, Z.-R.; Guo, J.; Wu, J.-C.; Chen, L.-S. Molecular mechanisms for pH-mediated amelioration of aluminum-toxicity revealed by conjoint analysis of transcriptome and metabolome in Citrus sinensis roots. Chemosphere 2022, 299, 134335. [Google Scholar] [CrossRef]
- Zheng, J.; Johnson, M.; Mandal, R.; Wishart, D.S. A Comprehensive targeted metabolomics assay for crop plant sample analysis. Metabolites 2021, 11, 303. [Google Scholar] [CrossRef]
- Cai, L.-Y.; Zhang, J.; Ren, Q.-Q.; Lai, Y.-H.; Peng, M.-Y.; Deng, C.-L.; Ye, X.; Yang, L.-T.; Huang, Z.-R.; Chen, L.-S. Increased pH-mediated alleviation of copper-toxicity and growth response function in Citrus sinensis seedlings. Sci. Horti. 2021, 288, 110310. [Google Scholar] [CrossRef]
- Yang, T.-Y.; Huang, W.-T.; Zhang, J.; Yang, L.-T.; Wu, B.-S.; Lai, N.-W.; Chen, L.-S. Raised pH conferred the ability to maintain a balance between production and detoxification of reactive oxygen species and methylglyoxal in aluminum-toxic Citrus sinensis leaves and roots. Environ. Pollut. 2021, 268 Pt B, 115676. [Google Scholar] [CrossRef]
- Li, R.; Kang, C.; Song, X.; Yu, L.; Liu, D.; He, S.; Zhai, H.; Liu, Q. A ζ-carotene desaturase gene, IbZDS, increases β-carotene and lutein contents and enhances salt tolerance in transgenic sweetpotato. Plant Sci. 2017, 262, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Liu, Y.J.; Huang, L.Q.; Cui, G.H.; Fu, G.F. Soil acidity elevates some phytohormone and β-eudesmol contents in roots of Atractylodes lancea. Russ. J. Plant Physiol. 2009, 56, 133–137. [Google Scholar] [CrossRef]
- Yan, F.; Schubert, S.; Mengel, K. Effect of low root medium pH on net proton release, root respiration, and root growth of corn (Zea mays L.) and broad bean (Vicia faba L.). Plant Physiol. 1992, 99, 415–421. [Google Scholar]
- Chen, Z.H.; Nimmo, G.A.; Jenkins, G.I.; Nimmo, H.G. BHLH32 modulates several biochemical and morphological processes that respond to Pi starvation in Arabidopsis. Biochem. J. 2007, 405, 191–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Z.-H.; Chen, L.-S.; Chen, R.-B.; Zhang, F.-Z.; Jiang, H.-X.; Tang, N.; Smith, B.R. Root release and metabolism of organic acids in tea plants in response to phosphorus supply. J. Plant Physiol. 2011, 168, 644–652. [Google Scholar] [CrossRef]
- Leegood, R.C.; Walker, R.P. Regulation and roles of phosphoenolpyruvate carboxykinase in plants. Arch. Biochem. Biophys. 2003, 414, 204–210. [Google Scholar] [CrossRef]
- Feng, Z.; Ding, C.; Li, W.; Wang, D.; Cui, D. Applications of metabolomics in the research of soybean plant under abiotic stress. Food Chem. 2020, 310, 125914. [Google Scholar] [CrossRef]
- Wan, H.; Du, J.; He, J.; Lyu, D.; Li, H. Copper accumulation, subcellular partitioning and physiological and molecular responses in relation to different copper tolerance in apple rootstocks. Tree Physiol. 2019, 39, 1215–1234. [Google Scholar] [CrossRef]
- Chen, G.X.; Liu, S.H.; Zhang, C.J.; Lu, C.J. Effects of drought on photosynthetic characteristics of flag leaves of a newly-developed super-high-yield rice hybrid. Photosynthetica 2004, 42, 573–578. [Google Scholar] [CrossRef]
- Zhang, K.; Liu, H.; Tao, P.; Chen, H. Comparative proteomic analyses provide new insights into low phosphorus stress responses in maize leaves. PLoS ONE 2014, 9, e98215. [Google Scholar] [CrossRef] [PubMed]
- Oestreicher, G.; Hogue, P.; Singer, T.P. Regulation of succinate dehydrogenase in higher Plants: II. Activation by substrates, reduced coenzyme Q, nucleotides, and anions. Plant Physiol. 1973, 52, 622–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, N.; Yüce, M.; Neslihan Öztürk Gökçe, Z.; Budak, H. Comparative metabolite profiling of drought stress in roots and leaves of seven triticeae species. BMC Genomics 2017, 18, 969. [Google Scholar] [CrossRef] [PubMed]
- Zandalinas, S.I.; Sales, C.; Beltrán, J.; Gómez-Cadenas, A.; Arbona, V. Activation of secondary metabolism in Citrus plants is associated to sensitivity to combined drought and high temperatures. Front. Plant Sci. 2017, 7, 1954. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Arias, D.; García-Machado, F.J.; Morales-Sierra, S.; Luis, J.C.; Suarez, E.; Hernández, M.; Valdés, F.; Borges, A.A. Lettuce plants treated with L-pyroglutamic acid increase yield under water deficit stress. Environ. Exp. Bot. 2019, 158, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Balfagón, D.; Rambla, J.L.; Granell, A.; Arbona, V.; Gómez-Cadenas, A. Grafting improves tolerance to combined drought and heat stresses by modifying metabolism in citrus scion. Environ. Exp. Bot. 2022, 195, 104793. [Google Scholar] [CrossRef]
- Liu, D.K.; Xu, C.C.; Zhang, L.; Ma, H.; Chen, X.J.; Sui, Y.C.; Zhang, H.Z. Evaluation of bioactive components and antioxidant capacity of four celery (Apium graveolens L.) leaves and petioles. Int. J. Food Prop. 2020, 23, 1097–1109. [Google Scholar] [CrossRef]
- Tilton, G.B.; Wedemeyer, W.J.; Browse, J.; Ohlrogge, J. Plant coenzyme A biosynthesis: Characterization of two pantothenate kinases from Arabidopsis. Plant Mol. Biol. 2006, 61, 629–642. [Google Scholar] [CrossRef]
- Berglund, T.; Wallström, A.; Nguyen, T.V.; Laurell, C.; Ohlsson, A.B. Nicotinamide; antioxidative and DNA hypomethylation effects in plant cells. Plant Physiol Biochem. 2017, 118, 551–560. [Google Scholar] [CrossRef]
- Ahmad, Z.; Bashir, K.; Matsui, A.; Tanaka, M.; Sasaki, R.; Oikawa, A.; Hirai, M.Y.; Chaomurilege; Zu, Y.; Kawai-Yamada, M.; et al. Overexpression of nicotinamidase 3 (NIC3) gene and the exogenous application of nicotinic acid (NA) enhance drought tolerance and increase biomass in Arabidopsis. Plant Mol. Biol. 2021, 107, 63–84. [Google Scholar] [CrossRef]
- Tawaraya, K.; Horie, R.; Shinano, T.; Wagatsuma, T.; Saito, K.; Oikawa, A. Metabolite profiling of soybean root exudates under phosphorus deficiency. Soil Sci. Plant Nutr. 2014, 60, 679–694. [Google Scholar] [CrossRef]
- Brown, S.; Clastre, M.; Courdavault, V.; O’Connor, S.E. De novo production of the plant-derived alkaloid strictosidine in yeast. Proc. Natl. Acad. Sci. USA. 2015, 112, 3205–3210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, W.; Yang, B.; Komatsu, S.; Lu, X.; Li, X.; Tian, J. Binary stress induces an increase in indole alkaloid biosynthesis in Catharanthus roseus. Front. Plant Sci. 2015, 6, 582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Z.-H.; Chen, L.-S.; Chen, R.-B.; Zhang, F.-Z.; Yang, L.-T.; Tang, N. Expression of genes for two phosphofructokinases, tonoplast ATPase subunit A, and pyrophosphatase of tea roots in response to phosphorus-deficiency. J. Hortic. Sci. Biotech. 2010, 85, 449–453. [Google Scholar] [CrossRef]
- Yang, L.-T.; Yang, G.-H.; You, X.; Zhou, C.-P.; Lu, Y.-B.; Chen, L.-S. Magnesium deficiency-induced changes in organic acid metabolism of Citrus sinensis roots and leaves. Biol. Plant. 2013, 57, 481–486. [Google Scholar] [CrossRef]
- Araújo, W.L.; Tohge, T.; Ishizaki, K.; Leaver, C.J.; Fernie, A.R. Protein degradation-an alternative respiratory substrate for stressed plants. Trends Plant Sci. 2011, 16, 489–498. [Google Scholar] [CrossRef]
- Li, H.; Yang, L.-T.; Qi, Y.-P.; Guo, P.; Lu, Y.-B.; Chen, L.-S. Aluminum toxicity-induced alterations of leaf proteome in two Citrus species differing in aluminum tolerance. Int. J. Mol. Sci. 2016, 17, 1180. [Google Scholar] [CrossRef] [Green Version]
- Mazzucotellin, E.; Mastrangelo, A.M.; Crosatti, C.; Guerra, D.; Stanca, A.M.; Cattivelli, L. Abiotic stress response in plants: When post-transcriptional and post-translational regulations control transcription. Plant Sci. 2008, 174, 420–431. [Google Scholar] [CrossRef]
- Santiago, J.; Rodrigues, A.; Saez, A.; Rubio, S.; Antoni, R.; Dupeux, F.; Park, S.Y.; Márquez, J.A.; Cutler, S.R.; Rodriguez, P.L. Modulation of drought resistance by the abscisic acid receptor PYL5 through inhibition of clade A PP2Cs. Plant J. 2009, 60, 575–588. [Google Scholar] [CrossRef]
- Sidari, M.; Panuccio, M.; Muscolo, A. Influence of acidity on growth and biochemistry of Pennisetum clandestinum. Biol. Plant. 2004, 48, 133–136. [Google Scholar] [CrossRef]
- Ferreira, T.C.; Aguilar, J.V.; Souza, L.A.; Justino, G.C.; Aguiar, L.F.; Camargos, L.S. pH effects on nodulation and biological nitrogen fixation in Calopogonium mucunoides. Braz. J. Bot. 2016, 39, 1015–1020. [Google Scholar] [CrossRef] [Green Version]
- McCue, P.; Zheng, Z.; Pinkham, J.L.; Shetty, K. A model for enhanced pea seedling vigour following low pH and salicylic acid treatments. Process Biochem. 2000, 35, 603–613. [Google Scholar] [CrossRef]
- Witt, S.; Galicia, L.; Lisec, J.; Cairns, J.; Tiessen, A.; Araus, J.L.; Palacios-Rojas, N.; Fernie, A.R. Metabolic and phenotypic responses of greenhouse-grown maize hybrids to experimentally controlled drought stress. Mol. Plant 2012, 5, 401–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.-Y.; Ren, Q.-Q.; Lai, N.-W.; Peng, M.-Y.; Zhang, J.; Yang, L.-T.; Huang, Z.-R.; Chen, L.-S. Metabolomics combined with physiology and transcriptomics reveals how Citrus grandis leaves cope with copper-toxicity. Ecotoxicol. Environ. Saf. 2021, 223, 112579. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liu, P.; Qi, D.; Peng, X.; Liu, G. Enhancement of cold and salt tolerance of Arabidopsis by transgenic expression of the S-adenosylmethionine decarboxylase gene from Leymus chinensis. J. Plant Physiol. 2017, 211, 90–99. [Google Scholar] [CrossRef]
- Naing, A.H.; Lee, D.B.; Ai, T.N.; Lim, K.B.; Kim, C.K. Enhancement of low pH stress tolerance in anthocyanin-enriched transgenetic petunia overexpressing RsMYB1 gene. Front. Plant Sci. 2018, 9, 1124. [Google Scholar] [CrossRef]
- Alotaibi, M.O.; Saleh, A.M.; Sobrinho, R.L.; Sheteiwy, M.S.; El-Sawah, A.M.; Mohammed, A.E.; AbdElgawad, H. Arbuscular mycorrhizae mitigate aluminum toxicity and regulate proline metabolism in plants grown in acidic soil. J. Fungi 2021, 7, 531. [Google Scholar] [CrossRef]
- Tzin, V.; Galili, G. New insights into the shikimate and aromatic amino acids biosynthesis pathways in plants. Mol. Plant 2010, 3, 956–972. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Moradi, M.; Razavi, F.; Rabiei, V. Exogenous phenylalanine application promotes chilling tolerance in tomato fruits during cold storage by ensuring supply of NADPH for activation of ROS scavenging systems. Sci. Hortic. 2019, 246, 818–825. [Google Scholar] [CrossRef]
- Rao, S.R.; Qayyum, A.; Razzaq, A.; Ahmad, M.; Mahmood, I.; Sher, A. Role of foliar application of salicylic acid and L-Tryptophan in drought tolerance of maize Japs. J. Anim. Plant Sci. 2012, 22, 768–772. [Google Scholar]
- Schlüter, U.; Mascher, M.; Colmsee, C.; Scholz, U.; Bräutigam, A.; Fahnenstich, H.; Sonnewald, U. Maize source leaf adaptation to nitrogen deficiency affects not only nitrogen and carbon metabolism but also control of phosphate homeostasis. Plant Physiol. 2012, 160, 1384–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.-S.; Yang, L.-T.; Lin, Z.-H.; Tang, N. Roles of organic acid metabolism in plant tolerance to phosphorus-deficiency. Prog. Bot. 2013, 74, 213–237. [Google Scholar]
- Cheng, Y.; Zhou, W.; El Sheery, N.I.; Peters, C.; Li, M.; Wang, X.; Huang, J. Characterization of the Arabidopsis glycerophosphodiester phosphodiesterase (GDPD) family reveals a role of the plastid-localized AtGDPD1 in maintaining cellular phosphate homeostasis under phosphate starvation. Plant J. 2011, 66, 781–795. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Ren, Z.; Lu, C. The phosphatidylcholine diacylglycerol cholinephosphotransferase is required for efficient hydroxy fatty acid accumulation in transgenic Arabidopsis. Plant Physiol. 2012, 158, 1944–1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- May, A.; Berger, S.; Hertel, T.; Köck, M. The Arabidopsis thaliana phosphate starvation responsive gene AtPPsPase1 encodes a novel type of inorganic pyrophosphatase. Biochim. Biophys. Acta-Gen. Subj. 2011, 1810, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ruan, W.; Shi, J.; Zhang, L.; Xiang, D.; Yang, C.; Li, C.; Wu, Z.; Liu, Y.; Yu, Y.; et al. Rice SPX1 and SPX2 inhibit phosphate starvation responses through interacting with PHR2 in a phosphate-dependent manner. Proc. Natl. Acad. Sci. USA 2014, 111, 14953–14958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.Y.; Aung, K.; Tseng, C.Y.; Chang, T.Y.; Chen, Y.S.; Chiou, T.J. Vacuolar Ca2+/H+ transport activity is required for systemic phosphate homeostasis involving shoot-to-root signaling in Arabidopsis. Plant Physiol. 2011, 156, 1176–1189. [Google Scholar] [CrossRef] [Green Version]
- Liu, D. Root developmental responses to phosphorus nutrition. J. Integr. Plant Biol. 2021, 63, 1065–1090. [Google Scholar] [CrossRef]
- Huang, C.F.; Yamaji, N.; Mitani, N.; Yano, M.; Nagamura, Y.; Ma, J.F. A bacterial-type ABC transporter is involved in aluminum tolerance in rice. Plant Cell 2009, 21, 655–667. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Piñeros, M.A.; Li, X.; Yang, H.; Liu, Y.; Murphy, A.S.; Kochian, L.V.; Liu, D. An Arabidopsis ABC transporter mediates phosphate deficiency-induced remodeling of root architecture by modulating iron homeostasis in roots. Mol. Plant. 2017, 10, 244–259. [Google Scholar] [CrossRef] [Green Version]
- Spetea, C.; Lundin, B. Evidence for nucleotide-dependent processes in the thylakoid lumen of plant chloroplasts–an update. FEBS Lett. 2012, 586, 2946–2954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson, P.M.; Herdean, A.; Adolfsson, L.; Beebo, A.; Nziengui, H.; Irigoyen, S.; Ünnep, R.; Zsiros, O.; Nagy, G.; Garab, G.; et al. The Arabidopsis thylakoid transporter PHT4;1 influences phosphate availability for ATP synthesis and plant growth. Plant J. 2015, 84, 99–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gho, Y.-S.; Choi, H.; Moon, S.; Song, M.Y.; Park, H.E.; Kim, D.-H.; Ha, S.-H.; Jung, K.-H. Phosphate-starvation-inducible S-like RNase genes in rice are involved in phosphate source recycling by RNA decay. Front. Plant Sci. 2020, 11, 585561. [Google Scholar] [CrossRef] [PubMed]
- Rojas, H.J.; Roldán, J.A.; Goldraij, A. NnSR1, a class III non-S-RNase constitutively expressed in styles, is induced in roots and stems under phosphate deficiency in Nicotiana alata. Ann. Bot. 2013, 112, 1351–1360. [Google Scholar] [CrossRef] [Green Version]
- Daniel, B.; Pavkov-Keller, T.; Steiner, B.; Dordic, A.; Gutmann, A.; Nidetzky, B.; Sensen, C.W.; van der Graaff, E.; Wallner, S.; Gruber, K.; et al. Oxidation of monolignols by members of the berberine bridge enzyme family suggests a role in plant cell wall metabolism. J. Biol. Chem. 2015, 290, 18770–18781. [Google Scholar] [CrossRef] [Green Version]
- Goujon, T.; Sibout, R.; Pollet, B.; Maba, B.; Nussaume, L.; Bechtold, N.; Lu, F.; Ralph, J.; Mila, I.; Barrière, Y.; et al. A new Arabidopsis thaliana mutant deficient in the expression of O-methyltransferase impacts lignins and sinapoyl esters. Plant Mol. Biol. 2003, 51, 973–989. [Google Scholar] [CrossRef]
- Besseau, S.; Hoffmann, L.; Geoffroy, P.; Lapierre, C.; Pollet, B.; Legrand, M. Flavonoid accumulation in Arabidopsis repressed in lignin synthesis affects auxin transport and plant growth. Plant Cell 2007, 19, 148–162. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Zeng, H.; Zhang, J.; Wei, S.; Huang, L. Characterization of enzymes involved in the interconversions of different forms of vitamin B6 in tobacco leaves. Plant Physiol. Biochem. 2011, 49, 1299–1305. [Google Scholar] [CrossRef]
- Guo, P.; Qi, Y.-P.; Cai, Y.-T.; Yang, T.-Y.; Yang, L.-T.; Huang, Z.-R.; Chen, L.-S. Aluminum effects on photosynthesis, reactive oxygen species and methylglyoxal detoxification in two Citrus species differing in aluminum tolerance. Tree Physiol. 2018, 38, 1548–1565. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, H.; Watanabe-Takahashi, A.; Smith, F.W.; Blake-Kalff, M.; Hawkesford, M.J.; Saito, K. The roles of three functional sulphate transporters involved in uptake and translocation of sulphate in Arabidopsis thaliana. Plant J. 2000, 23, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Awazuhara, M.; Fujiwara, T.; Hayashi, H.; Watanabe-Takahashi, A.; Takahashi, H.; Saito, K. The function of SULTR2; 1 sulfate transporter during seed development in Arabidopsis thaliana. Physiol. Plant. 2005, 125, 95–105. [Google Scholar] [CrossRef]
- Tattini, M.; Loreto, F.; Fini, A.; Guidi, L.; Brunetti, C.; Velikova, V.; Gori, A.; Ferrini, F. Isoprenoids and phenylpropanoids are part of the antioxidant defense orchestrated daily by drought-stressed Platanus × acerifolia plants during Mediterranean summers. New Phytol. 2015, 207, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Stincone, A.; Prigione, A.; Cramer, T.; Wamelink, M.M.; Campbell, K.; Cheung, E.; Olin Sandoval, V.; Grüning, N.M.; Krüger, A.; TauqeerAlam, M.; et al. The return of metabolism: Biochemistry and physiology of the pentose phosphate pathway. Biol. Rev. 2015, 90, 927–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mano, J.; Belles-Boix, E.; Babiychuk, E.; Inzé, D.; Torii, Y.; Hiraoka, E.; Takimoto, K.; Slooten, L.; Asada, K.; Kushnir, S. Protection against photooxidative injury of tobacco leaves by 2-alkenal reductase. Detoxication of lipid peroxide-derived reactive carbonyls. Plant Physiol. 2005, 139, 1773–1783. [Google Scholar] [CrossRef] [Green Version]
- Yalcinkaya, T.; Uzilday, B.; Ozgur, R.; Turkan, I.; Mano, J. Lipid peroxidation-derived reactive carbonyl species (RCS): Their interaction with ROS and cellular redox during environmental stresses. Environ. Exp. Bot. 2019, 165, 139–149. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]
- Bu, D.; Luo, H.; Huo, P.; Wang, Z.; Zhang, S.; He, Z.; Wu, Y.; Zhao, L.; Liu, J.; Guo, J.; et al. KOBAS-i: Intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res. 2021, 49, W317–W325. [Google Scholar] [CrossRef]
- Klupczynska, A.; Plewa, S.; Sytek, N.; Sawicki, W.; Dereziński, P.; Matysiak, J.; Kokot, Z.J. A study of low-molecular-weight organic acid urinary profiles in prostate cancer by a new liquid chromatography-tandem mass spectrometry method. J. Pharm. Biomed. Anal. 2018, 159, 229–236. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, N.-W.; Zheng, Z.-C.; Hua, D.; Zhang, J.; Chen, H.-H.; Ye, X.; Huang, Z.-R.; Guo, J.; Yang, L.-T.; Chen, L.-S. Molecular and Physiological Responses of Citrus sinensis Leaves to Long-Term Low pH Revealed by RNA-Seq Integrated with Targeted Metabolomics. Int. J. Mol. Sci. 2022, 23, 5844. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105844
Lai N-W, Zheng Z-C, Hua D, Zhang J, Chen H-H, Ye X, Huang Z-R, Guo J, Yang L-T, Chen L-S. Molecular and Physiological Responses of Citrus sinensis Leaves to Long-Term Low pH Revealed by RNA-Seq Integrated with Targeted Metabolomics. International Journal of Molecular Sciences. 2022; 23(10):5844. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105844
Chicago/Turabian StyleLai, Ning-Wei, Zhi-Chao Zheng, Dan Hua, Jiang Zhang, Huan-Huan Chen, Xin Ye, Zeng-Rong Huang, Jiuxin Guo, Lin-Tong Yang, and Li-Song Chen. 2022. "Molecular and Physiological Responses of Citrus sinensis Leaves to Long-Term Low pH Revealed by RNA-Seq Integrated with Targeted Metabolomics" International Journal of Molecular Sciences 23, no. 10: 5844. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105844