
The Profile of MicroRNA Expression and Potential Role in the Regulation of Drug-Resistant Genes in Doxorubicin and Topotecan Resistant Ovarian Cancer Cell Lines

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Gene Chip Quality Assessment
2.2. Gene Expression Evaluation and Gene Expression Lists
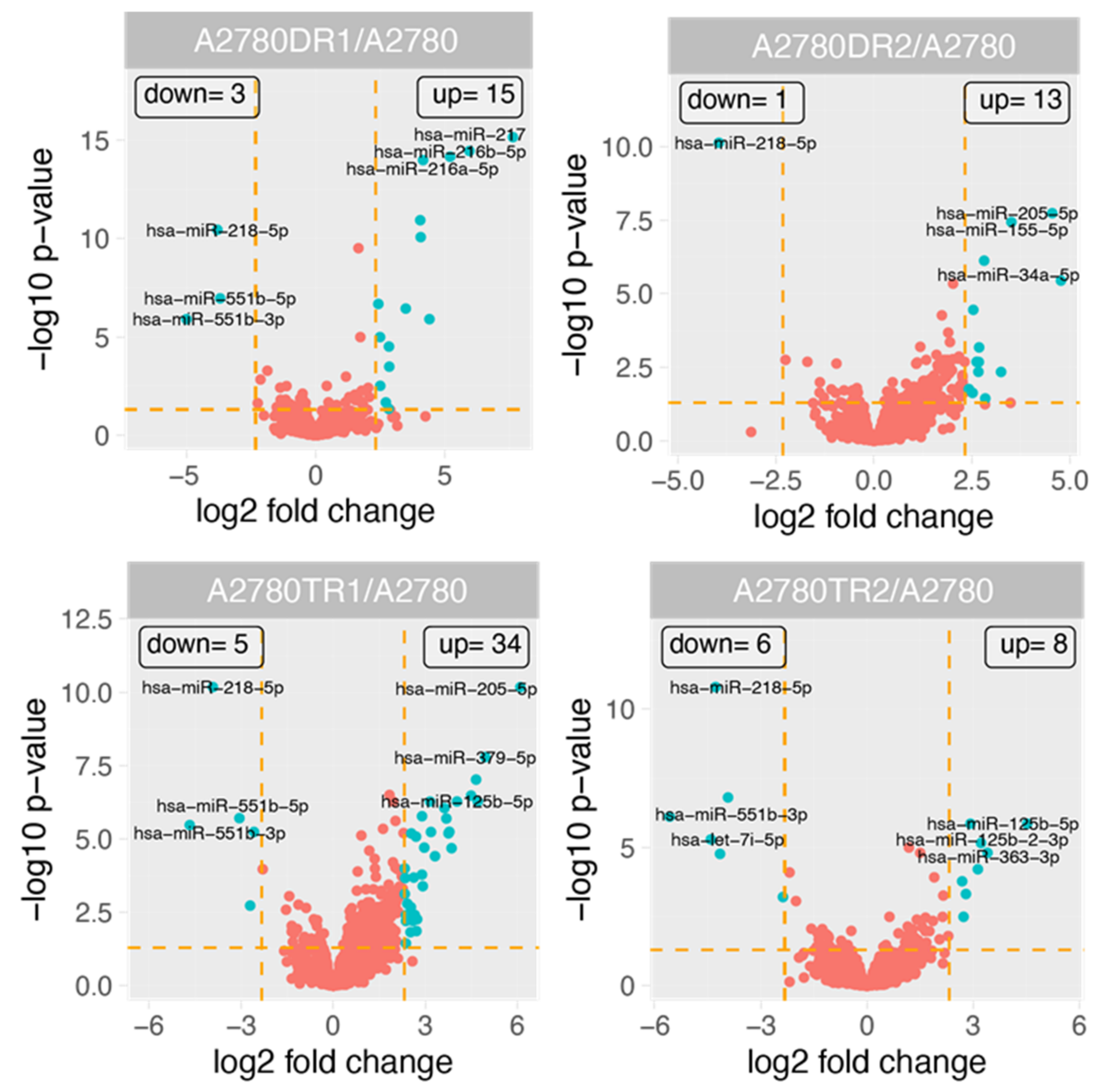
2.3. miRNAs Expression in DOX and TOP Resistant Cell Lines
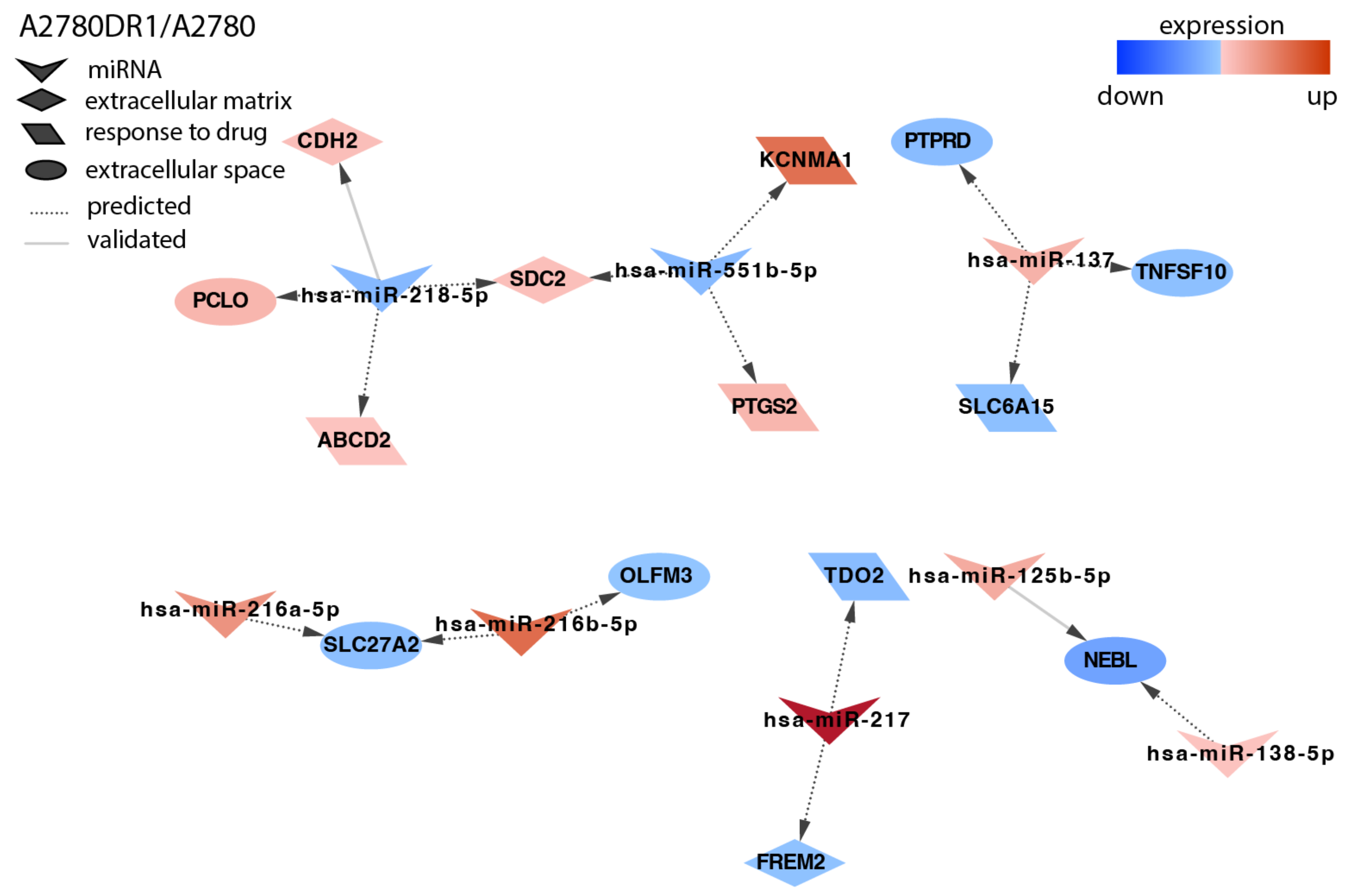
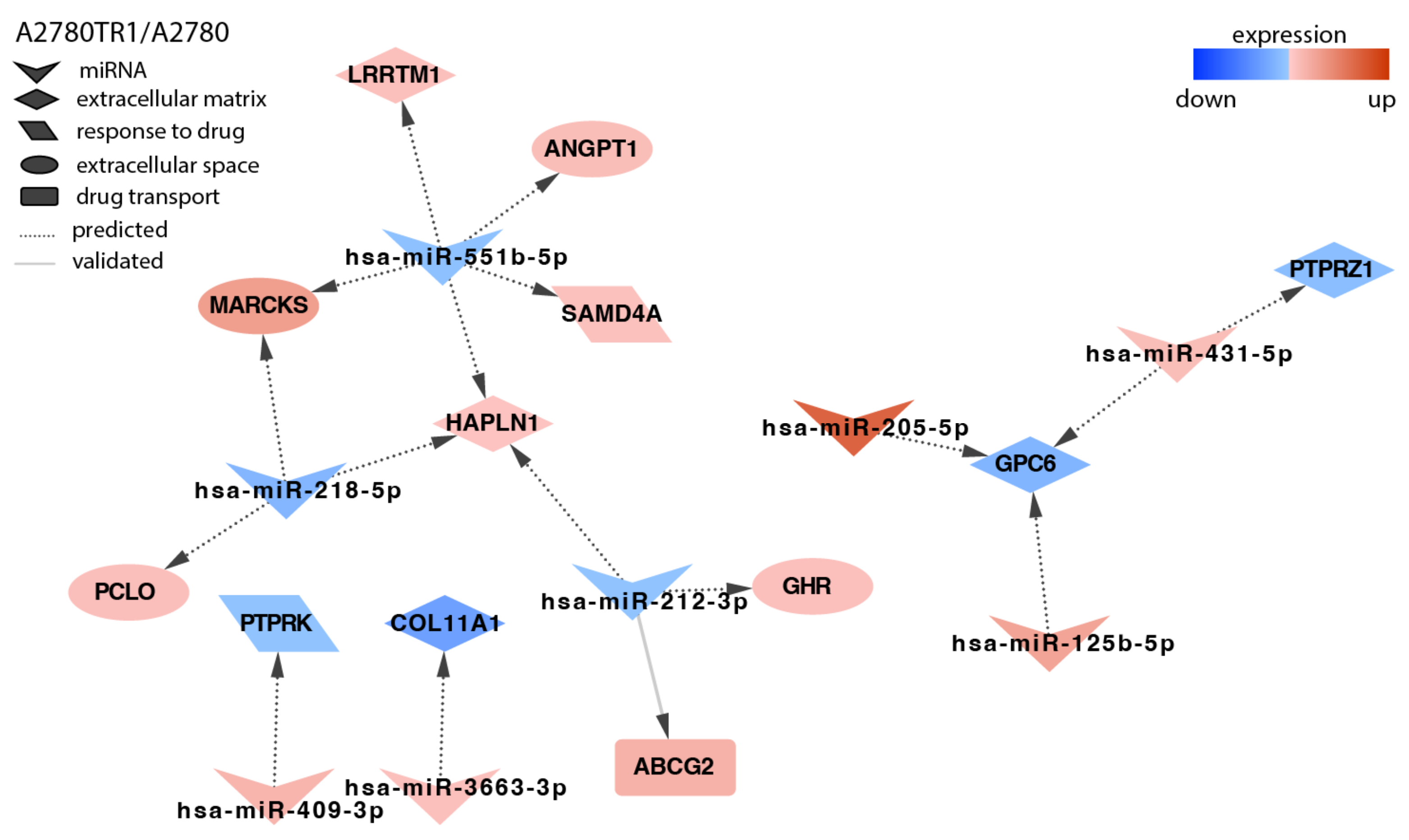
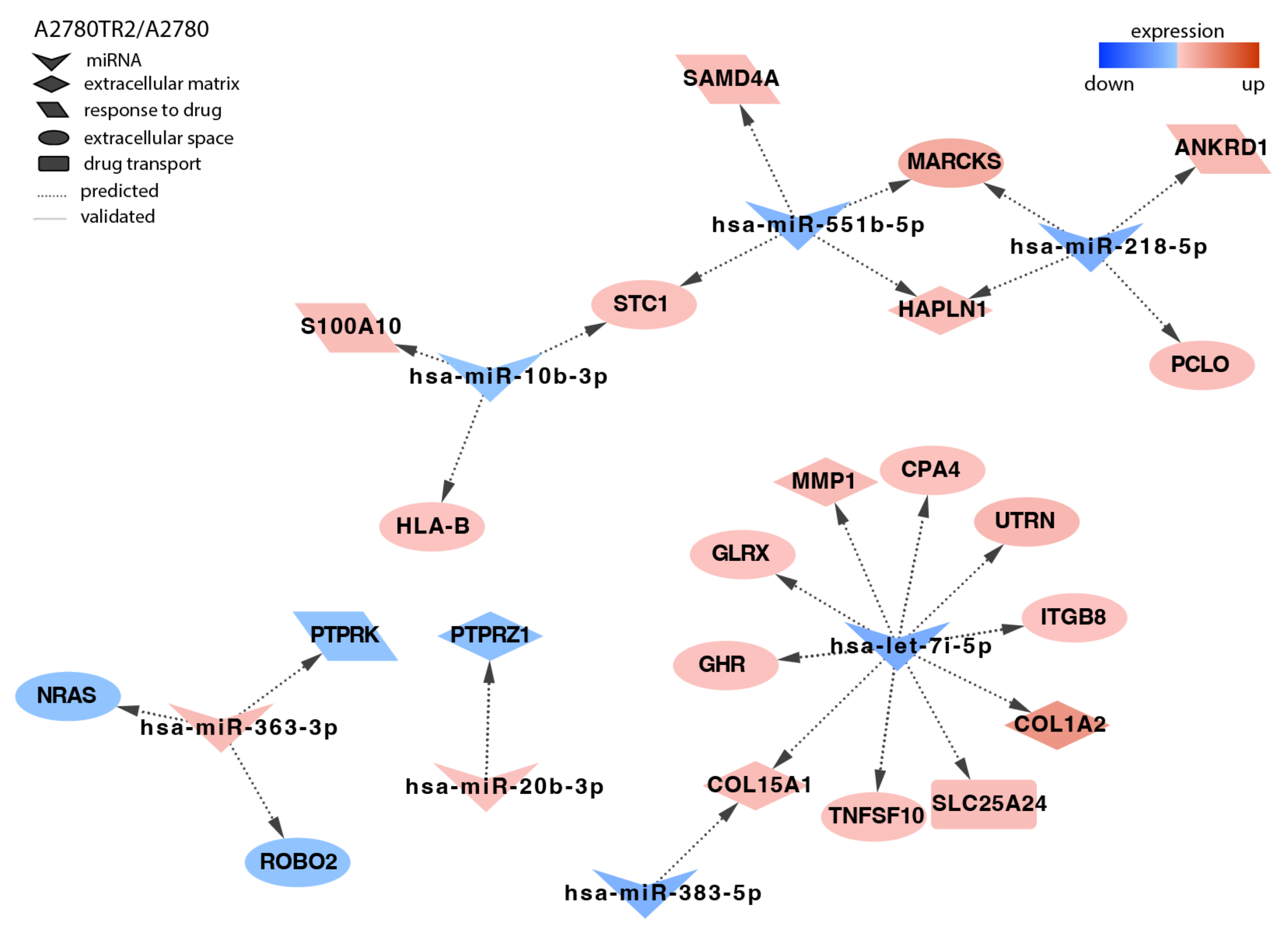
2.4. Analysis of Target Genes Expression
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Lines and Cell Culture
4.3. miRNA Isolation
4.4. Microarray Preparation, Hybridization, and Scanning
4.5. Microarray Analysis and miRNA Gene Screening
4.6. miRNA-Target Gene Prediction
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennessy, B.T.; Coleman, R.L.; Markman, M. Ovarian Cancer. Lancet 2009, 374, 1371–1382. [Google Scholar] [CrossRef]
- Webb, P.M.; Jordan, S.J. Epidemiology of Epithelial Ovarian Cancer. Best Pract. Res. Clin. Obstet. Gynaecol. 2017, 41, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Webber, K.; Friedlander, M. Chemotherapy for Epithelial Ovarian, Fallopian Tube and Primary Peritoneal Cancer. Best Pract. Res. Clin. Obstet. Gynaecol. 2017, 41, 126–138. [Google Scholar] [CrossRef]
- Parmar, M.K.B.; Ledermann, J.A.; Colombo, N.; du Bois, A.; Delaloye, J.-F.; Kristensen, G.B.; Wheeler, S.; Swart, A.M.; Qian, W.; Torri, V.; et al. Paclitaxel plus Platinum-Based Chemotherapy versus Conventional Platinum-Based Chemotherapy in Women with Relapsed Ovarian Cancer: The ICON4/AGO-OVAR-2.2 Trial. Lancet 2003, 361, 2099–2106. [Google Scholar] [CrossRef]
- Ferrandina, G.; Ludovisi, M.; Lorusso, D.; Pignata, S.; Breda, E.; Savarese, A.; Del Medico, P.; Scaltriti, L.; Katsaros, D.; Priolo, D.; et al. Phase III Trial of Gemcitabine Compared with Pegylated Liposomal Doxorubicin in Progressive or Recurrent Ovarian Cancer. J. Clin. Oncol. 2008, 26, 890–896. [Google Scholar] [CrossRef]
- Sehouli, J.; Stengel, D.; Oskay-Oezcelik, G.; Zeimet, A.G.; Sommer, H.; Klare, P.; Stauch, M.; Paulenz, A.; Camara, O.; Keil, E.; et al. Nonplatinum Topotecan Combinations Versus Topotecan Alone for Recurrent Ovarian Cancer: Results of a Phase III Study of the North-Eastern German Society of Gynecological Oncology Ovarian Cancer Study Group. J. Clin. Oncol. 2008, 26, 3176–3182. [Google Scholar] [CrossRef]
- Pommier, Y.; Leo, E.; Zhang, H.; Marchand, C. DNA Topoisomerases and Their Poisoning by Anticancer and Antibacterial Drugs. Chem. Biol. 2010, 17, 421–433. [Google Scholar] [CrossRef] [Green Version]
- Lihua, P.; Chen, X.Y.; Wu, T.X. Topotecan for Ovarian Cancer. Cochrane Database Syst. Rev. 2008, 2008, CD005589. [Google Scholar] [CrossRef]
- Hartwell, D.; Jones, J.; Loveman, E.; Harris, P.; Clegg, A.; Bird, A. Topotecan for Relapsed Small Cell Lung Cancer: A Systematic Review and Economic Evaluation. Cancer Treat. Rev. 2011, 37, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Pommier, Y.; Sun, Y.; Huang, S.-Y.N.; Nitiss, J.L. Roles of Eukaryotic Topoisomerases in Transcription, Replication and Genomic Stability. Nat. Rev. Mol. Cell Biol. 2016, 17, 703–721. [Google Scholar] [CrossRef] [PubMed]
- Staker, B.L.; Hjerrild, K.; Feese, M.D.; Behnke, C.A.; Burgin, A.B.; Stewart, L. The Mechanism of Topoisomerase I Poisoning by a Camptothecin Analog. Proc. Natl. Acad. Sci. USA 2002, 99, 15387–15392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maliepaard, M.; van Gastelen, M.A.; de Jong, L.A.; Pluim, D.; van Waardenburg, R.C.; Ruevekamp-Helmers, M.C.; Floot, B.G.; Schellens, J.H. Overexpression of the BCRP/MXR/ABCP Gene in a Topotecan-Selected Ovarian Tumor Cell Line. Cancer Res. 1999, 59, 4559–4563. [Google Scholar] [PubMed]
- Robey, R.W.; Polgar, O.; Deeken, J.; To, K.W.; Bates, S.E. ABCG2: Determining Its Relevance in Clinical Drug Resistance. Cancer Metastasis Rev. 2007, 26, 39–57. [Google Scholar] [CrossRef] [Green Version]
- Januchowski, R.; Sterzyńska, K.; Zaorska, K.; Sosińska, P.; Klejewski, A.; Brązert, M.; Nowicki, M.; Zabel, M. Analysis of MDR Genes Expression and Cross-Resistance in Eight Drug Resistant Ovarian Cancer Cell Lines. J. Ovarian Res. 2016, 9, 65. [Google Scholar] [CrossRef] [Green Version]
- Klejewski, A.; Świerczewska, M.; Zaorska, K.; Brązert, M.; Nowicki, M.; Zabel, M.; Januchowski, R. New and Old Genes Associated with Topotecan Resistance Development in Ovarian Cancer Cell Lines. Anticancer Res. 2017, 37, 1625–1636. [Google Scholar] [CrossRef] [Green Version]
- Januchowski, R.; Wojtowicz, K.; Sujka-Kordowska, P.; Andrzejewska, M.; Zabel, M. MDR Gene Expression Analysis of Six Drug-Resistant Ovarian Cancer Cell Lines. BioMed Res. Int. 2013, 2013, 241763. [Google Scholar] [CrossRef] [Green Version]
- Mutch, D.G.; Orlando, M.; Goss, T.; Teneriello, M.G.; Gordon, A.N.; McMeekin, S.D.; Wang, Y.; Scribner, D.R.; Marciniack, M.; Naumann, R.W.; et al. Randomized Phase III Trial of Gemcitabine Compared with Pegylated Liposomal Doxorubicin in Patients with Platinum-Resistant Ovarian Cancer. J. Clin. Oncol. 2007, 25, 2811–2818. [Google Scholar] [CrossRef]
- Gordon, A.N.; Tonda, M.; Sun, S.; Rackoff, W.; Doxil Study 30-49 Investigators. Long-Term Survival Advantage for Women Treated with Pegylated Liposomal Doxorubicin Compared with Topotecan in a Phase 3 Randomized Study of Recurrent and Refractory Epithelial Ovarian Cancer. Gynecol. Oncol. 2004, 95, 1–8. [Google Scholar] [CrossRef]
- Uemura, T.; Ohkura, H.; Adachi, Y.; Morino, K.; Shiozaki, K.; Yanagida, M. DNA Topoisomerase II Is Required for Condensation and Separation of Mitotic Chromosomes in S. Pombe. Cell 1987, 50, 917–925. [Google Scholar] [CrossRef]
- Tacar, O.; Sriamornsak, P.; Dass, C.R. Doxorubicin: An Update on Anticancer Molecular Action, Toxicity and Novel Drug Delivery Systems. J. Pharm. Pharmacol. 2013, 65, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, J.I.; Williams, R.T.; Henderson, M.J.; Norris, M.D.; Haber, M. ABC Transporters as Mediators of Drug Resistance and Contributors to Cancer Cell Biology. Drug Resist. Updat. 2016, 26, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Januchowski, R.; Sterzyńska, K.; Zawierucha, P.; Ruciński, M.; Świerczewska, M.; Partyka, M.; Bednarek-Rajewska, K.; Brązert, M.; Nowicki, M.; Zabel, M.; et al. Microarray-Based Detection and Expression Analysis of New Genes Associated with Drug Resistance in Ovarian Cancer Cell Lines. Oncotarget 2017, 8, 49944–49958. [Google Scholar] [CrossRef] [Green Version]
- Borucka, J.; Sterzyńska, K.; Kaźmierczak, D.; Świerczewska, M.; Nowacka, M.; Wojtowicz, K.; Klejewski, A.; Nowicki, M.; Zabel, M.; Ramlau, R.; et al. The Significance of Interferon Gamma Inducible Protein 16 (IFI16) Expression in Drug Resistant Ovarian Cancer Cell Lines. Biomed. Pharmacother. 2022, 150, 113036. [Google Scholar] [CrossRef]
- Świerczewska, M.; Sterzyńska, K.; Wojtowicz, K.; Kaźmierczak, D.; Iżycki, D.; Nowicki, M.; Zabel, M.; Januchowski, R. PTPRK Expression Is Downregulated in Drug Resistant Ovarian Cancer Cell Lines, and Especially in ALDH1A1 Positive CSCs-Like Populations. Int. J. Mol. Sci. 2019, 20, 2053. [Google Scholar] [CrossRef] [Green Version]
- Wojtowicz, K.; Sterzyńska, K.; Świerczewska, M.; Nowicki, M.; Zabel, M.; Januchowski, R. Piperine Targets Different Drug Resistance Mechanisms in Human Ovarian Cancer Cell Lines Leading to Increased Sensitivity to Cytotoxic Drugs. Int. J. Mol. Sci. 2021, 22, 4243. [Google Scholar] [CrossRef]
- Correia, A.L.; Bissell, M.J. The Tumor Microenvironment Is a Dominant Force in Multidrug Resistance. Drug Resist. Updat. 2012, 15, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Sterzyńska, K.; Kaźmierczak, D.; Klejewski, A.; Świerczewska, M.; Wojtowicz, K.; Nowacka, M.; Brązert, J.; Nowicki, M.; Januchowski, R. Expression of Osteoblast-Specific Factor 2 (OSF-2, Periostin) Is Associated with Drug Resistance in Ovarian Cancer Cell Lines. Int. J. Mol. Sci. 2019, 20, 3927. [Google Scholar] [CrossRef] [Green Version]
- Klejewski, A.; Sterzyńska, K.; Wojtowicz, K.; Świerczewska, M.; Partyka, M.; Brązert, M.; Nowicki, M.; Zabel, M.; Januchowski, R. The Significance of Lumican Expression in Ovarian Cancer Drug-Resistant Cell Lines. Oncotarget 2017, 8, 74466–74478. [Google Scholar] [CrossRef] [Green Version]
- Januchowski, R.; Świerczewska, M.; Sterzyńska, K.; Wojtowicz, K.; Nowicki, M.; Zabel, M. Increased Expression of Several Collagen Genes Is Associated with Drug Resistance in Ovarian Cancer Cell Lines. J. Cancer 2016, 7, 1295–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowacka, M.; Ginter-Matuszewska, B.; Świerczewska, M.; Sterzyńska, K.; Nowicki, M.; Januchowski, R. Effect of ALDH1A1 Gene Knockout on Drug Resistance in Paclitaxel and Topotecan Resistant Human Ovarian Cancer Cell Lines in 2D and 3D Model. Int. J. Mol. Sci. 2022, 23, 3036. [Google Scholar] [CrossRef] [PubMed]
- Tannock, I.F.; Lee, C.M.; Tunggal, J.K.; Cowan, D.S.M.; Egorin, M.J. Limited Penetration of Anticancer Drugs through Tumor Tissue: A Potential Cause of Resistance of Solid Tumors to Chemotherapy. Clin. Cancer Res. 2002, 8, 878–884. [Google Scholar] [PubMed]
- Di Paolo, A.; Bocci, G. Drug Distribution in Tumors: Mechanisms, Role in Drug Resistance, and Methods for Modification. Curr. Oncol. Rep. 2007, 9, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Wantoch von Rekowski, K.; König, P.; Henze, S.; Schlesinger, M.; Zawierucha, P.; Januchowski, R.; Bendas, G. The Impact of Integrin-Mediated Matrix Adhesion on Cisplatin Resistance of W1 Ovarian Cancer Cells. Biomolecules 2019, 9, 788. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans Heterochronic Gene Lin-4 Encodes Small RNAs with Antisense Complementarity to Lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Friedman, R.C.; Farh, K.K.-H.; Burge, C.B.; Bartel, D.P. Most Mammalian MRNAs Are Conserved Targets of MicroRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Fischer, S.; Handrick, R.; Aschrafi, A.; Otte, K. Unveiling the Principle of MicroRNA-Mediated Redundancy in Cellular Pathway Regulation. RNA Biol. 2015, 12, 238–247. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.-H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA Genes Are Transcribed by RNA Polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef]
- Mandilaras, V.; Vernon, M.; Meryet-Figuière, M.; Karakasis, K.; Lambert, B.; Poulain, L.; Oza, A.; Denoyelle, C.; Lheureux, S. Updates and Current Challenges in MicroRNA Research for Personalized Medicine in Ovarian Cancer. Expert Opin. Biol. Ther. 2017, 17, 927–943. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Shoorei, H.; Taheri, M. MiRNA Profile in Ovarian Cancer. Exp. Mol. Pathol. 2020, 113, 104381. [Google Scholar] [CrossRef] [PubMed]
- Deb, B.; Uddin, A.; Chakraborty, S. MiRNAs and Ovarian Cancer: An Overview. J. Cell. Physiol. 2018, 233, 3846–3854. [Google Scholar] [CrossRef] [PubMed]
- Hammond, S.M. An Overview of MicroRNAs. Adv. Drug Deliv. Rev. 2015, 87, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.S.; Dutta, A. MicroRNAs in Cancer. Annu. Rev. Pathol. Mech. Dis. 2009, 4, 199–227. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, X.; Cobb, G.P.; Anderson, T.A. MicroRNAs as Oncogenes and Tumor Suppressors. Dev. Biol. 2007, 302, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Xu, S.; Liu, X. MicroRNA Profiling of Plasma Exosomes from Patients with Ovarian Cancer Using High throughput Sequencing. Oncol. Lett. 2019, 17, 5601–5607. [Google Scholar] [CrossRef] [Green Version]
- Penyige, A.; Márton, É.; Soltész, B.; Szilágyi-Bónizs, M.; Póka, R.; Lukács, J.; Széles, L.; Nagy, B. Circulating MiRNA Profiling in Plasma Samples of Ovarian Cancer Patients. Int. J. Mol. Sci. 2019, 20, 4533. [Google Scholar] [CrossRef] [Green Version]
- Mihanfar, A.; Fattahi, A.; Nejabati, H.R. MicroRNA-Mediated Drug Resistance in Ovarian Cancer. J. Cell. Physiol. 2019, 234, 3180–3191. [Google Scholar] [CrossRef]
- Acunzo, M.; Romano, G.; Wernicke, D.; Croce, C.M. MicroRNA and Cancer—A Brief Overview. Adv. Biol. Regul. 2015, 57, 1–9. [Google Scholar] [CrossRef]
- Kazmierczak, D.; Jopek, K.; Sterzynska, K.; Ginter-Matuszewska, B.; Nowicki, M.; Rucinski, M.; Januchowski, R. The Significance of MicroRNAs Expression in Regulation of Extracellular Matrix and Other Drug Resistant Genes in Drug Resistant Ovarian Cancer Cell Lines. Int. J. Mol. Sci. 2020, 21, 2619. [Google Scholar] [CrossRef] [Green Version]
- Kazmierczak, D.; Jopek, K.; Sterzynska, K.; Nowicki, M.; Rucinski, M.; Januchowski, R. The Profile of MicroRNA Expression and Potential Role in the Regulation of Drug-Resistant Genes in Cisplatin- and Paclitaxel-Resistant Ovarian Cancer Cell Lines. Int. J. Mol. Sci. 2022, 23, 526. [Google Scholar] [CrossRef] [PubMed]
- Mitamura, T.; Watari, H.; Wang, L.; Kanno, H.; Hassan, M.K.; Miyazaki, M.; Katoh, Y.; Kimura, T.; Tanino, M.; Nishihara, H.; et al. Downregulation of MiRNA-31 Induces Taxane Resistance in Ovarian Cancer Cells through Increase of Receptor Tyrosine Kinase MET. Oncogenesis 2013, 2, e40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Januchowski, R.; Zawierucha, P.; Andrzejewska, M.; Ruciński, M.; Zabel, M. Microarray-Based Detection and Expression Analysis of ABC and SLC Transporters in Drug-Resistant Ovarian Cancer Cell Lines. Biomed. Pharmacother. 2013, 67, 240–245. [Google Scholar] [CrossRef]
- Januchowski, R.; Zawierucha, P.; Ruciński, M.; Nowicki, M.; Zabel, M. Extracellular Matrix Proteins Expression Profiling in Chemoresistant Variants of the A2780 Ovarian Cancer Cell Line. BioMed Res. Int. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Świerczewska, M.; Klejewski, A.; Wojtowicz, K.; Brązert, M.; Iżycki, D.; Nowicki, M.; Zabel, M.; Januchowski, R. New and Old Genes Associated with Primary and Established Responses to Cisplatin and Topotecan Treatment in Ovarian Cancer Cell Lines. Molecules 2017, 22, 1717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, B.; Li, C.; Cai, P.; Yu, L.; Zhao, B.; Chen, G. Knockdown of MiR 935 Increases Paclitaxel Sensitivity via Regulation of SOX7 in Non small cell Lung Cancer. Mol. Med. Rep. 2018, 18, 3397–3402. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, J.; Yu, Z.; Li, J.; Sun, R.; Kan, Q. MiR-935 Promotes Liver Cancer Cell Proliferation and Migration by Targeting SOX7. Oncol. Res. 2017, 25, 427–435. [Google Scholar] [CrossRef]
- Liu, F.; Chen, Y.; Chen, B.; Liu, C.; Xing, J. MiR-935 Promotes Clear Cell Renal Cell Carcinoma Migration and Invasion by Targeting IREB2. Cancer Manag. Res. 2019, 11, 10891–10900. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Xiao, W.; Jiang, X.; Li, H. MicroRNA-935 Acts as a Prognostic Marker and Promotes Cell Proliferation, Migration, and Invasion in Colorectal Cancer. Cancer Biomark. 2019, 26, 229–237. [Google Scholar] [CrossRef]
- Chen, Z.; Guo, X.; Sun, S.; Lu, C.; Wang, L. Serum MiR-125b Levels Associated with Epithelial Ovarian Cancer (EOC) Development and Treatment Responses. Bioengineered 2020, 11, 311–317. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Wang, L.; Tan, G.; Guo, Z.; Liu, L.; Yang, M.; He, J. MicroRNA-218 Inhibits Proliferation and Invasion in Ovarian Cancer by Targeting Runx2. Oncotarget 2017, 8, 91530–91541. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, A.; Sawada, K.; Nakamura, K.; Kinose, Y.; Nakatsuka, E.; Kobayashi, M.; Miyamoto, M.; Ishida, K.; Matsumoto, Y.; Kodama, M.; et al. Exosomal MiR-99a-5p Is Elevated in Sera of Ovarian Cancer Patients and Promotes Cancer Cell Invasion by Increasing Fibronectin and Vitronectin Expression in Neighboring Peritoneal Mesothelial Cells. BMC Cancer 2018, 18, 1065. [Google Scholar] [CrossRef] [PubMed]
- Samli, H.; Samli, M.; Vatansever, B.; Ardicli, S.; Aztopal, N.; Dincel, D.; Sahin, A.; Balci, F. Paclitaxel resistance and the role of miRNAs in prostate cancer cell lines. World J. Urol. 2019, 37, 1117–1126. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, C.; Zhang, A.; Wang, Q.; Ge, J.; Li, Q.; Xiao, J. Long Non-Coding RNA SDCBP2-AS1 Delays the Progression of Ovarian Cancer via MicroRNA-100-5p-Targeted EPDR1. World J. Surg. Oncol. 2021, 19, 199. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.-L.; Lu, J.-H.; Wang, Y.; Sheng, H.; Wang, Y.-N.; Chen, Z.-H.; Wu, Q.-N.; Zheng, J.-B.; Chen, Y.-X.; Yang, D.-D.; et al. The LncRNA XIST/MiR-125b-2-3p Axis Modulates Cell Proliferation and Chemotherapeutic Sensitivity via Targeting Wee1 in Colorectal Cancer. Cancer Med. 2021, 10, 2423–2441. [Google Scholar] [CrossRef]
- Huang, H.-Q.; Chen, G.; Xiong, D.-D.; Lai, Z.-F.; Liu, L.-M.; Fang, Y.-Y.; Shen, J.-H.; Gan, X.-Y.; Liao, L.-F.; Dang, Y.-W. Down-Regulation of MicroRNA-125b-2-3p Is a Risk Factor for a Poor Prognosis in Hepatocellular Carcinoma. Bioengineered 2021, 12, 1627–1641. [Google Scholar] [CrossRef]
- Qi, C.-L.; Sheng, J.-F.; Huang, M.-L.; Zou, Y.; Wang, Y.-P.; Wang, F.; Zeng, F.; Hua, Q.-Q.; Chen, S.-M. Integrated Analysis of Deregulation MicroRNA Expression in Head and Neck Squamous Cell Carcinoma. Medicine 2021, 100, e24618. [Google Scholar] [CrossRef]
- Qi, L.; Sun, B.; Yang, B.; Lu, S. LINC00665 Stimulates Breast Cancer Progression via Regulating MiR-551b-5p. Cancer Manag. Res. 2021, 13, 1113–1121. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, X.; Hu, Z.; Wang, Y.; Liu, M.; Liu, X.; Li, H.; Ji, R.; Guo, Q.; Zhou, Y. Identification and Characterization of Tumor Suppressor and Oncogenic MiRNAs in Gastric Cancer. Oncol. Lett. 2015, 10, 329–336. [Google Scholar] [CrossRef]
- Chang, W.; Wang, Y.; Li, W.; Shi, L.; Geng, Z. MicroRNA-551b-3p Inhibits Tumour Growth of Human Cholangiocarcinoma by Targeting Cyclin D1. J. Cell. Mol. Med. 2019, 23, 4945–4954. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Xie, C.; Liu, Y.; Shi, Q.; Chen, Y. Up-Regulation of MiR-383-5p Suppresses Proliferation and Enhances Chemosensitivity in Ovarian Cancer Cells by Targeting TRIM27. Biomed. Pharmacother. 2019, 109, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Tu, C.; Chen, W.; Wang, S.; Tan, W.; Guo, J.; Shao, C.; Wang, W. MicroRNA-383 Inhibits Doxorubicin Resistance in Hepatocellular Carcinoma by Targeting Eukaryotic Translation Initiation Factor 5A2. J. Cell. Mol. Med. 2019, 23, 7190–7199. [Google Scholar] [CrossRef] [PubMed]
- Han, R.L.; Wang, F.P.; Zhang, P.A.; Zhou, X.Y.; Li, Y. MiR-383 Inhibits Ovarian Cancer Cell Proliferation, Invasion and Aerobic Glycolysis by Targeting LDHA. Neoplasma 2017, 64, 244–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilming Elgaaen, B.; Olstad, O.K.; Haug, K.B.F.; Brusletto, B.; Sandvik, L.; Staff, A.C.; Gautvik, K.M.; Davidson, B. Global MiRNA Expression Analysis of Serous and Clear Cell Ovarian Carcinomas Identifies Differentially Expressed MiRNAs Including MiR-200c-3p as a Prognostic Marker. BMC Cancer 2014, 14, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Qin, R.; Guan, A.; Yao, Y.; Huang, Y.; Jia, H.; Huang, W.; Gao, J. HOTAIR Enhanced Paclitaxel and Doxorubicin Resistance in Gastric Cancer Cells Partly through Inhibiting MiR-217 Expression. J. Cell. Biochem. 2018, 119, 7226–7234. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Deng, T.; Su, C.; Shang, Z. MicroRNA 217 Inhibits Cell Proliferation and Enhances Chemosensitivity to Doxorubicin in Acute Myeloid Leukemia by Targeting KRAS. Oncol. Lett. 2017, 13, 4986–4994. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, D.; Zhang, W. Tumor Suppressor Role of MiR-217 in Human Epithelial Ovarian Cancer by Targeting IGF1R. Oncol. Rep. 2016, 35, 1671–1679. [Google Scholar] [CrossRef]
- Pei, M.L.; Zhao, Z.X.; Shuang, T. Dysregulation of Lnc-SNHG1 and MiR-216b-5p Correlate with Chemoresistance and Indicate Poor Prognosis of Serous Epithelial Ovarian Cancer. J. Ovarian Res. 2020, 13, 144. [Google Scholar] [CrossRef]
- Zhang, J.; Gao, S.; Zhang, Y.; Yi, H.; Xu, M.; Xu, J.; Liu, H.; Ding, Z.; He, H.; Wang, H.; et al. MiR-216a-5p Inhibits Tumorigenesis in Pancreatic Cancer by Targeting TPT1/MTORC1 and Is Mediated by LINC01133. Int. J. Biol. Sci. 2020, 16, 2612–2627. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, P.; Zou, J.-Y.; Zou, G.; Wang, W.-Z.; Liu, Y.-L.; Zhao, H.-W.; Fang, A.-P. MiR-216a-5p Act as a Tumor Suppressor, Regulating the Cell Proliferation and Metastasis by Targeting PAK2 in Breast Cancer. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 2469–2475. [Google Scholar] [CrossRef]
- Zeng, X.; Liu, Y.; Zhu, H.; Chen, D.; Hu, W. Downregulation of MiR-216a-5p by Long Noncoding RNA PVT1 Suppresses Colorectal Cancer Progression via Modulation of YBX1 Expression. Cancer Manag. Res. 2019, 11, 6981–6993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Yin, Y.; Shan, X.; Zhou, X.; Liu, P.; Cao, Q.; Zhu, D.; Zhang, J.; Zhu, W. The Value of Plasma-Based MicroRNAs as Diagnostic Biomarkers for Ovarian Cancer. Am. J. Med. Sci. 2019, 358, 256–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Zhu, P.; Gao, Y.; Wang, A. MiR 379 5p Inhibits Cell Proliferation and Promotes Cell Apoptosis in Non small Cell Lung Cancer by Targeting Β arrestin 1. Mol. Med. Rep. 2020, 22, 4499–4508. [Google Scholar] [CrossRef]
- Khan, S.; Brougham, C.L.; Ryan, J.; Sahrudin, A.; O’Neill, G.; Wall, D.; Curran, C.; Newell, J.; Kerin, M.J.; Dwyer, R.M. MiR-379 Regulates Cyclin B1 Expression and Is Decreased in Breast Cancer. PLoS ONE 2013, 8, e68753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fletcher, J.I.; Haber, M.; Henderson, M.J.; Norris, M.D. ABC Transporters in Cancer: More than Just Drug Efflux Pumps. Nat. Rev. Cancer 2010, 10, 147–156. [Google Scholar] [CrossRef] [PubMed]
- St Croix, B.; Kerbel, R.S. Cell Adhesion and Drug Resistance in Cancer. Curr. Opin. Oncol. 1997, 9, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Morin, P.J. Drug Resistance and the Microenvironment: Nature and Nurture. Drug Resist. Updat. 2003, 6, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Işeri, O.D.; Kars, M.D.; Arpaci, F.; Gündüz, U. Gene Expression Analysis of Drug-Resistant MCF-7 Cells: Implications for Relation to Extracellular Matrix Proteins. Cancer Chemother. Pharmacol. 2010, 65, 447–455. [Google Scholar] [CrossRef]
- Sherman-Baust, C.A.; Weeraratna, A.T.; Rangel, L.B.A.; Pizer, E.S.; Cho, K.R.; Schwartz, D.R.; Shock, T.; Morin, P.J. Remodeling of the Extracellular Matrix through Overexpression of Collagen VI Contributes to Cisplatin Resistance in Ovarian Cancer Cells. Cancer Cell 2003, 3, 377–386. [Google Scholar] [CrossRef] [Green Version]
- Sher, Y.-P.; Wang, L.-J.; Chuang, L.-L.; Tsai, M.-H.; Kuo, T.-T.; Huang, C.-C.; Chuang, E.Y.; Lai, L.-C. ADAM9 Up-Regulates N-Cadherin via MiR-218 Suppression in Lung Adenocarcinoma Cells. PLoS ONE 2014, 9, e94065. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Yang, R.; Chen, J.; Qi, E.; Zhou, S.; Wang, Y.; Fu, Q.; Chen, R.; Fang, X. Let-7i-5p Regulation of Cell Morphology and Migration Through Distinct Signaling Pathways in Normal and Pathogenic Urethral Fibroblasts. Front. Bioeng. Biotechnol. 2020, 8, 428. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, H.-X.; Li, Y.-L.; Zhang, C.-C.; Zhou, C.-Y.; Wang, L.; Xia, Y.-L.; Du, J.; Li, H.-H. MicroRNA Let-7i Negatively Regulates Cardiac Inflammation and Fibrosis. Hypertension 2015, 66, 776–785. [Google Scholar] [CrossRef] [PubMed]
- Ardito, F.; Giuliani, M.; Perrone, D.; Troiano, G.; Lo Muzio, L. The Crucial Role of Protein Phosphorylation in Cell Signaling and Its Use as Targeted Therapy (Review). Int. J. Mol. Med. 2017, 40, 271–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.; Grandis, J.R. Receptor-Type Protein Tyrosine Phosphatases in Cancer. Chin. J. Cancer 2015, 34, 61–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abukhdeir, A.M.; Park, B.H. P21 and P27: Roles in Carcinogenesis and Drug Resistance. Expert Rev. Mol. Med. 2008, 10, e19. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.-H.; Ye, L.; Mason, M.D.; Jiang, W.G. Protein Tyrosine Phosphatase Kappa (PTPRK) Is a Negative Regulator of Adhesion and Invasion of Breast Cancer Cells, and Associates with Poor Prognosis of Breast Cancer. J. Cancer Res. Clin. Oncol. 2013, 139, 1129–1139. [Google Scholar] [CrossRef]
- Chen, Y.-W.; Guo, T.; Shen, L.; Wong, K.-Y.; Tao, Q.; Choi, W.W.L.; Au-Yeung, R.K.H.; Chan, Y.-P.; Wong, M.L.Y.; Tang, J.C.O.; et al. Receptor-Type Tyrosine-Protein Phosphatase κ Directly Targets STAT3 Activation for Tumor Suppression in Nasal NK/T-Cell Lymphoma. Blood 2015, 125, 1589–1600. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R. The Eph Family Receptors and Ligands. Pharmacol. Ther. 1998, 77, 151–181. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, G.; Zhao, Y.; Wu, Y.; Li, J.; Chai, Y. Expression of EphA7 protein in primary hepatocellular carcinoma and its clinical significance. Zhonghua Wai Ke Za Zhi 2010, 48, 53–56. [Google Scholar]
- Huang, J.; Jiang, W.; Tong, X.; Zhang, L.; Zhang, Y.; Fan, H. Identification of Gene and MicroRNA Changes in Response to Smoking in Human Airway Epithelium by Bioinformatics Analyses. Medicine 2019, 98, e17267. [Google Scholar] [CrossRef]
- Dickinson, R.E.; Fegan, K.S.; Ren, X.; Hillier, S.G.; Duncan, W.C. Glucocorticoid Regulation of SLIT/ROBO Tumour Suppressor Genes in the Ovarian Surface Epithelium and Ovarian Cancer Cells. PLoS ONE 2011, 6, e27792. [Google Scholar] [CrossRef] [Green Version]
- Pinder, B.D.; Smibert, C.A. Smaug: An Unexpected Journey into the Mechanisms of Post-Transcriptional Regulation. Fly 2013, 7, 142–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, D.N.; Faruq, O.; Zhang, L.; Rastgoo, N.; Liu, A.; Chang, H. Pathophysiological Roles of Myristoylated Alanine-Rich C-Kinase Substrate (MARCKS) in Hematological Malignancies. Biomark. Res. 2021, 9, 34. [Google Scholar] [CrossRef] [PubMed]
- Fong, L.W.R.; Yang, D.C.; Chen, C.-H. Myristoylated Alanine-Rich C Kinase Substrate (MARCKS): A Multirole Signaling Protein in Cancers. Cancer Metastasis Rev. 2017, 36, 737–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-H.; Cheng, C.-T.; Yuan, Y.; Zhai, J.; Arif, M.; Fong, L.W.R.; Wu, R.; Ann, D.K. Elevated MARCKS Phosphorylation Contributes to Unresponsiveness of Breast Cancer to Paclitaxel Treatment. Oncotarget 2015, 6, 15194–15208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Chen, Y.; Saha, M.N.; Chen, J.; Evans, K.; Qiu, L.; Reece, D.; Chen, G.A.; Chang, H. Targeting Phospho-MARCKS Overcomes Drug-Resistance and Induces Antitumor Activity in Preclinical Models of Multiple Myeloma. Leukemia 2015, 29, 715–726. [Google Scholar] [CrossRef]
- Kaehler, M.; Ruemenapp, J.; Gonnermann, D.; Nagel, I.; Bruhn, O.; Haenisch, S.; Ammerpohl, O.; Wesch, D.; Cascorbi, I.; Bruckmueller, H. MicroRNA-212/ABCG2-Axis Contributes to Development of Imatinib-Resistance in Leukemic Cells. Oncotarget 2017, 8, 92018–92031. [Google Scholar] [CrossRef] [Green Version]
- Reustle, A.; Fisel, P.; Renner, O.; Büttner, F.; Winter, S.; Rausch, S.; Kruck, S.; Nies, A.T.; Hennenlotter, J.; Scharpf, M.; et al. Characterization of the Breast Cancer Resistance Protein (BCRP/ABCG2) in Clear Cell Renal Cell Carcinoma. Int. J. Cancer 2018, 143, 3181–3193. [Google Scholar] [CrossRef] [Green Version]
- Dietel, M.; Bals, U.; Schaefer, B.; Herzig, I.; Arps, H.; Zabel, M. In Vitro Prediction of Cytostatic Drug Resistance in Primary Cell Cultures of Solid Malignant Tumours. Eur. J. Cancer 1993, 29A, 416–420. [Google Scholar] [CrossRef]
- Stelcer, E.; Kulcenty, K.; Rucinski, M.; Jopek, K.; Richter, M.; Trzeciak, T.; Suchorska, W.M. The Role of MicroRNAs in Early Chondrogenesis of Human Induced Pluripotent Stem Cells (HiPSCs). Int. J. Mol. Sci. 2019, 20, 4371. [Google Scholar] [CrossRef] [Green Version]
- Kulcenty, K.; Wroblewska, J.P.; Rucinski, M.; Kozlowska, E.; Jopek, K.; Suchorska, W.M. MicroRNA Profiling During Neural Differentiation of Induced Pluripotent Stem Cells. Int. J. Mol. Sci. 2019, 20, 3651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautier, L.; Cope, L.; Bolstad, B.M.; Irizarry, R.A. Affy—Analysis of Affymetrix GeneChip Data at the Probe Level. Bioinformatics 2004, 20, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Cava, C.; Colaprico, A.; Bertoli, G.; Graudenzi, A.; Silva, T.C.; Olsen, C.; Noushmehr, H.; Bontempi, G.; Mauri, G.; Castiglioni, I. SpidermiR: An R/Bioconductor Package for Integrative Analysis with MiRNA Data. Int. J. Mol. Sci. 2017, 18, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, M.; Brägelmann, J.; Kryukov, I.; Saraiva-Agostinho, N.; Perner, S. FirebrowseR: An R Client to the Broad Institute’s Firehose Pipeline. Database J. Biol. Databases Curation 2017, 2017, baw160. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRBase Accession Number | Gene Name | Fold Change (adj.p.val) | |||
|---|---|---|---|---|---|
| A2780DR1 | A2780DR2 | A2780TR1 | A2780TR2 | ||
| MIMAT0000097 | hsa-miR-99a-5p | 7.23575965 | N.S | 13.4763932 | 6.94690251 |
| MIMAT0000098 | hsa-miR-100-5p | 5.64229368 | N.S | 5.82903562 | 7.58322275 |
| MIMAT0000252 | hsa-miR-7-5p | N.S | N.S | 5.76802822 | N.S |
| MIMAT0000255 | hsa-miR-34a-5p | N.S | 27.3069918 | N.S | N.S |
| MIMAT0000266 | hsa-miR-205-5p | N.S | 23.4285218 | 68.1826341 | N.S |
| MIMAT0000269 | hsa-miR-212-3p | N.S | NS | −5.98258494 | N.S |
| MIMAT0000273 | hsa-miR-216a-5p | 37.0885659 | N.S | N.S | N.S |
| MIMAT0004959 | hsa-miR-216b-5p | 62.2848826 | N.S | N.S | N.S |
| MIMAT0000274 | hsa-miR-217 | 200.01382 | N.S | N.S | N.S |
| MIMAT0000275 | hsa-miR-218-5p | −13.9723251 | −15.4598435 | −15.1300271 | −19.3883641 |
| MIMAT0000415 | hsa-let-7i-5p | N.S | N.S | N.S | −21.2652763 |
| MIMAT0000423 | hsa-miR-125b-5p | 21.2850391 | 6.14958848 | 26.0504456 | 23.089459 |
| MIMAT0004603 | hsa-miR-125b-2-3p | 7.17389557 | N.S | 12.3020071 | 9.34279559 |
| MIMAT0003233 | hsa-miR-551b-3p | −32.0993258 | N.S | −25.4904329 | −47.6292686 |
| MIMAT0004794 | hsa-miR-551b-5p | −12.9832503 | N.S | −8.30875013 | −15.1855837 |
| MIMAT0000429 | hsa-miR-137 | 16.8235232 | N.S | N.S | N.S |
| MIMAT0000430 | hsa-miR-138-5p | 5.67137662 | N.S | N.S | N.S |
| MIMAT0000446 | hsa-miR-127-3p | N.S | N.S | 22.4289639 | N.S |
| MIMAT0000447 | hsa-miR-134-5p | N.S | N.S | 9.93993656 | N.S |
| MIMAT0000646 | hsa-miR-155-5p | N.S | 11.3736149 | N.S | N.S |
| MIMAT0000707 | hsa-miR-363-3p | N.S | N.S | N.S | 10.6015891 |
| MIMAT0000722 | hsa-miR-370-3p | N.S | N.S | 6.47195247 | N.S |
| MIMAT0000733 | hsa-miR-379-5p | N.S | N.S | 30.945542 | N.S |
| MIMAT0000737 | hsa-miR-382-5p | N.S | N.S | 9.07152577 | N.S |
| MIMAT0000738 | hsa-miR-383-5p | N.S | N.S | −6.51726481 | −17.7612762 |
| MIMAT0001625 | hsa-miR-431-5p | N.S | N.S | 8.87926645 | N.S |
| MIMAT0001639 | hsa-miR-409-3p | N.S | N.S | 14.4397796 | N.S |
| MIMAT0002178 | hsa-miR-487a-3p | N.S | N.S | 7.4124047 | N.S |
| MIMAT0002814 | hsa-miR-432-5p | N.S | N.S | 13.744849 | N.S |
| MIMAT0002816 | hsa-miR-494-3p | N.S | N.S | 16.3141937 | N.S |
| MIMAT0003161 | hsa-miR-493-3p | N.S | N.S | 12.8494251 | N.S |
| MIMAT0003180 | hsa-miR-487b-3p | N.S | N.S | 25.1135622 | N.S |
| MIMAT0003239 | hsa-miR-574-3p | N.S | 5.79962256 | N.S | N.S |
| MIMAT0003297 | hsa-miR-628-3p | N.S | 9.47532594 | N.S | N.S |
| MIMAT0004556 | hsa-miR-10b-3p | −2.842098067 | −4.763634832 | −2.347192792 | −5.183265974 |
| MIMAT0004679 | hsa-miR-296-3p | 11.258982 | N.S | N.S | N.S |
| MIMAT0004752 | hsa-miR-20b-3p | N.S | N.S | N.S | 6.45001658 |
| MIMAT0004776 | hsa-miR-505-5p | 6.59829128 | N.S | N.S | NS |
| MIMAT0004951 | hsa-miR-887-3p | NS | 6.42842883 | N.S | 8.77616954 |
| MIMAT0004978 | hsa-miR-935 | 7.10687862 | 7.1679309 | N.S | N.S |
| MIMAT0005865 | hsa-miR-1202 | N.S | N.S | 7.40030689 | N.S |
| MIMAT0009201 | hsa-miR-196b-3p | 16.6120623 | N.S | N.S | N.S |
| MIMAT0010251 | hsa-miR-449c-5p | N.S | N.S | 1.87888757 | 6.63203316 |
| MIMAT0015070 | hsa-miR-3188 | N.S | N.S | 6.29125598 | N.S |
| MIMAT0015079 | hsa-miR-3195 | N.S | 7.03029052 | N.S | N.S |
| MIMAT0018085 | hsa-miR-3663-3p | 1.956795219 | 6.39351738 | 7.544158456 | 4.146984658 |
| MIMAT0018198 | hsa-miR-3923 | 17.9229767 | N.S | N.S | N.S |
| MIMAT0018968 | hsa-miR-4449 | N.S | 5.72863128 | 5.72224812 | N.S |
| MIMAT0019691 | hsa-miR-4634 | N.S | N.S | 5.06702643 | N.S |
| MIMAT0019979 | hsa-miR-4800-3p | N.S | N.S | 5.32867026 | N.S |
| MIMAT0022838 | hsa-miR-1185-1-3p | N.S | N.S | 6.15646443 | N.S |
| MIMAT0023693 | hsa-miR-6068 | N.S | N.S | 5.15633061 | N.S |
| MIMAT0023700 | hsa-miR-6075 | N.S | N.S | 5.02749704 | N.S |
| MIMAT0023705 | hsa-miR-6080 | N.S | 5.36404532 | 6.00292053 | N.S |
| MIMAT0027480 | hsa-miR-6790-5p | N.S | N.S | 6.63324861 | N.S |
| MIMAT0027678 | hsa-miR-6889-5p | N.S | N.S | 5.1618654 | N.S |
| MIMAT0028121 | hsa-miR-7112-5p | N.S | 6.3493949 | 5.16702158 | N.S |
| MIMAT0028213 | hsa-miR-7151-3p | 5.39257884 | N.S | N.S | N.S |
| MIMAT0030420 | hsa-miR-7845-5p | N.S | N.S | 5.02021002 | N.S |
| MIMAT0030990 | hsa-miR-8063 | N.S | N.S | 6.53874406 | N.S |
| MIMAT0030991 | hsa-miR-8064 | N.S | N.S | 7.80350724 | N.S |
| miRBase Accession Number | Gene Name | Fold Change (adj.p.val) | |||
|---|---|---|---|---|---|
| A2780DR1 | A2780DR2 | A2780TR1 | A2780TR2 | ||
| MIMAT0000097 | hsa-miR-99a-5p | 7.23575965 | N.S | 13.4763932 | 6.94690251 |
| MIMAT0000098 | hsa-miR-100-5p | 5.64229368 | N.S | 5.82903562 | 7.58322275 |
| MIMAT0000255 | hsa-miR-34a-5p | N.S | 27.3069918 | N.S | N.S |
| MIMAT0000266 | hsa-miR-205-5p | N.S | 23.4285218 | 68.1826341 | N.S |
| MIMAT0000273 | hsa-miR-216a-5p | 37.0885659 | N.S | N.S | N.S |
| MIMAT0004959 | hsa-miR-216b-5p | 62.2848826 | N.S | N.S | N.S |
| MIMAT0000274 | hsa-miR-217 | 200.01382 | N.S | N.S | N.S |
| MIMAT0000275 | hsa-miR-218-5p | −13.9723251 | −15.4598435 | −15.1300271 | −19.3883641 |
| MIMAT0000415 | hsa-let-7i-5p | N.S | N.S | N.S | −21.2652763 |
| MIMAT0000423 | hsa-miR-125b-5p | 21.2850391 | 6.14958848 | 26.0504456 | 23.089459 |
| MIMAT0004603 | hsa-miR-125b-2-3p | 7.17389557 | N.S | 12.3020071 | 9.34279559 |
| MIMAT0003233 | hsa-miR-551b-3p | −32.0993258 | N.S | −25.4904329 | −47.6292686 |
| MIMAT0004794 | hsa-miR-551b-5p | −12.9832503 | N.S | −8.30875013 | −15.1855837 |
| MIMAT0000429 | hsa-miR-137 | 16.8235232 | N.S | N.S | N.S |
| MIMAT0000446 | hsa-miR-127-3p | N.S | N.S | 22.4289639 | N.S |
| MIMAT0000646 | hsa-miR-155-5p | N.S | 11.3736149 | N.S | N.S |
| MIMAT0000707 | hsa-miR-363-3p | N.S | N.S | N.S | 10.6015891 |
| MIMAT0000733 | hsa-miR-379-5p | N.S | N.S | 30.945542 | N.S |
| MIMAT0000738 | hsa-miR-383-5p | N.S | N.S | −6.51726481 | −17.7612762 |
| MIMAT0001639 | hsa-miR-409-3p | N.S | N.S | 14.4397796 | N.S |
| MIMAT0002814 | hsa-miR-432-5p | N.S | N.S | 13.744849 | N.S |
| MIMAT0002816 | hsa-miR-494-3p | N.S | N.S | 16.3141937 | N.S |
| MIMAT0003161 | hsa-miR-493-3p | N.S | N.S | 12.8494251 | N.S |
| MIMAT0003180 | hsa-miR-487b-3p | N.S | N.S | 25.1135622 | N.S |
| MIMAT0004679 | hsa-miR-296-3p | 11.258982 | N.S | N.S | N.S |
| MIMAT0004978 | hsa-miR-935 | 7.10687862 | 7.1679309 | N.S | N.S |
| MIMAT0009201 | hsa-miR-196b-3p | 16.6120623 | N.S | N.S | N.S |
| MIMAT0018198 | hsa-miR-3923 | 17.9229767 | N.S | N.S | N.S |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stasiak, P.; Kaźmierczak, D.; Jopek, K.; Nowicki, M.; Rucinski, M.; Januchowski, R. The Profile of MicroRNA Expression and Potential Role in the Regulation of Drug-Resistant Genes in Doxorubicin and Topotecan Resistant Ovarian Cancer Cell Lines. Int. J. Mol. Sci. 2022, 23, 5846. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105846
Stasiak P, Kaźmierczak D, Jopek K, Nowicki M, Rucinski M, Januchowski R. The Profile of MicroRNA Expression and Potential Role in the Regulation of Drug-Resistant Genes in Doxorubicin and Topotecan Resistant Ovarian Cancer Cell Lines. International Journal of Molecular Sciences. 2022; 23(10):5846. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105846
Chicago/Turabian StyleStasiak, Piotr, Dominika Kaźmierczak, Karol Jopek, Michał Nowicki, Marcin Rucinski, and Radosław Januchowski. 2022. "The Profile of MicroRNA Expression and Potential Role in the Regulation of Drug-Resistant Genes in Doxorubicin and Topotecan Resistant Ovarian Cancer Cell Lines" International Journal of Molecular Sciences 23, no. 10: 5846. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105846