
Isolation and Characterization of Germline Stem Cells in Protogynous Hermaphroditic Monopterus albus


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Ethics
2.2. Fish and Sample Preparation
2.3. Culturing of Germline Stem Cells
2.4. Alkaline Phosphatase (AP) Staining
2.5. Cell Transplantation
2.6. RT-PCR and Immunofluorescence Analysis of Gonads of Chimeric Fish
2.7. Reverse Transcription Polymerase Chain Reaction (RT-PCR), Quantitative Real-Time PCR (qRT-PCR), and RNA-Sequencing
2.8. Hematoxylin–Eosin (HE) Staining
2.9. Immunofluorescence Assay
2.10. Statistical Analysis
3. Results
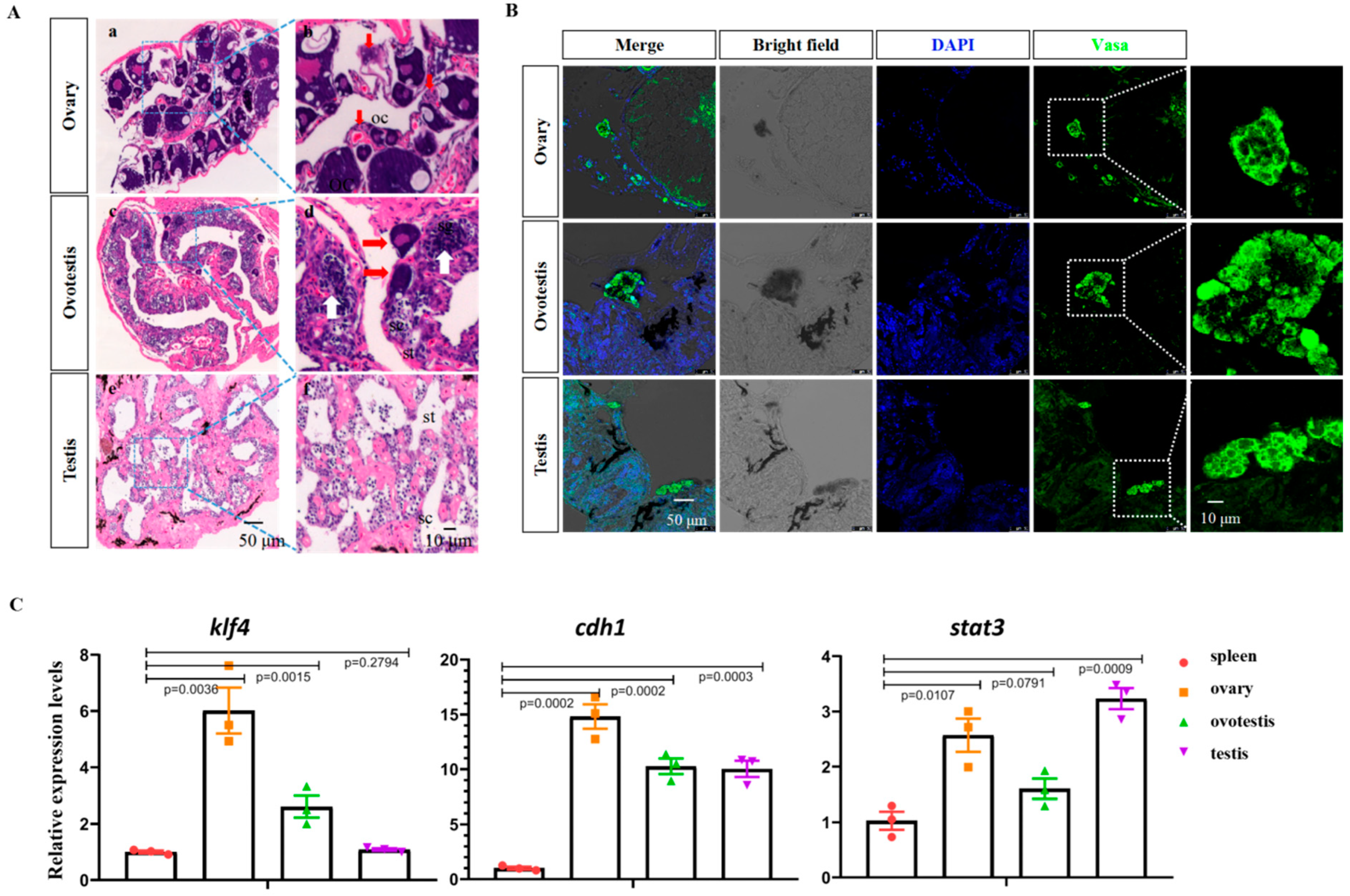
3.1. Characterization of GSCs in Ovary, Ovotestis, and Testis
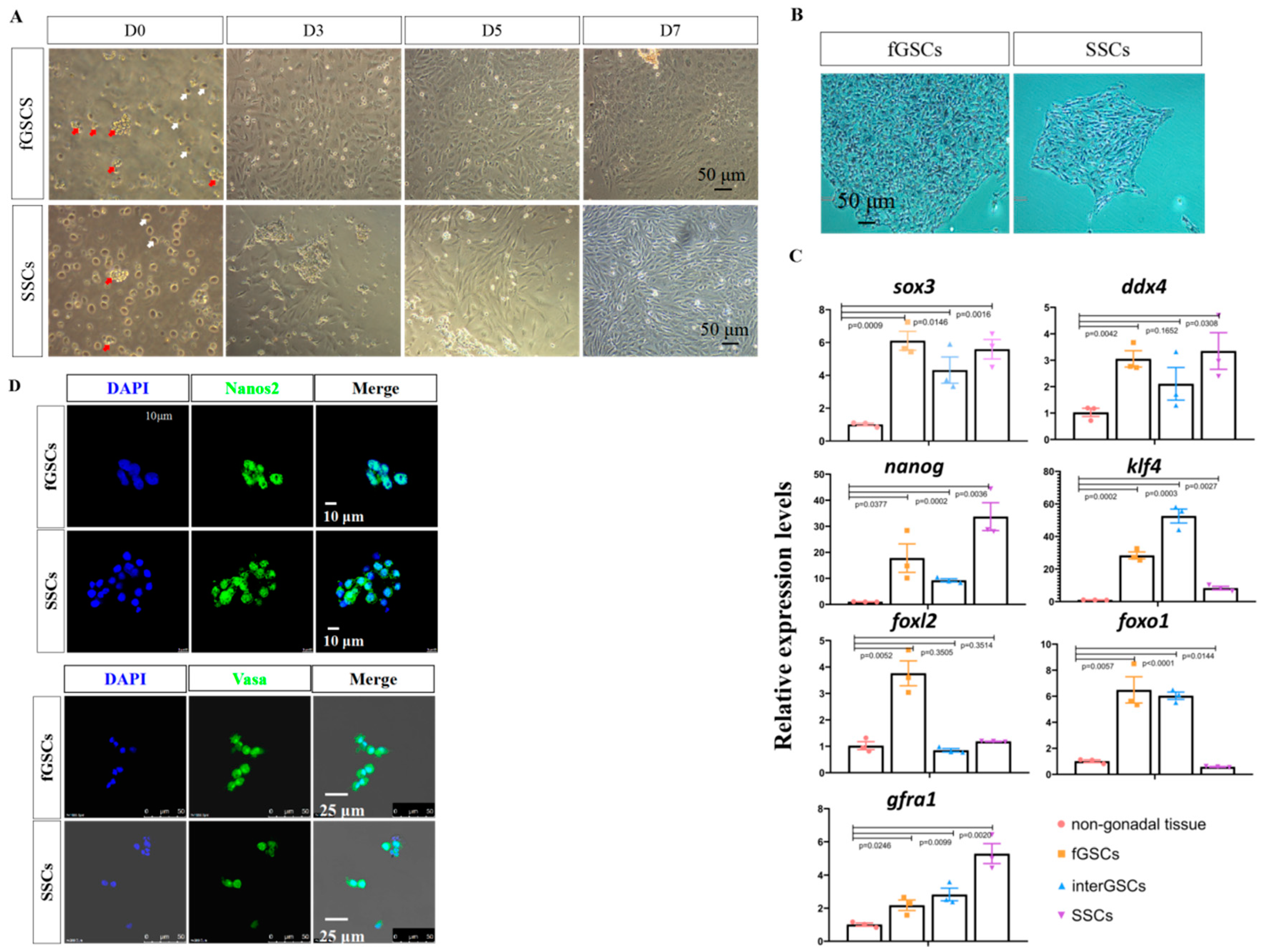
3.2. Isolation and Culture of fGSCs and SSCs
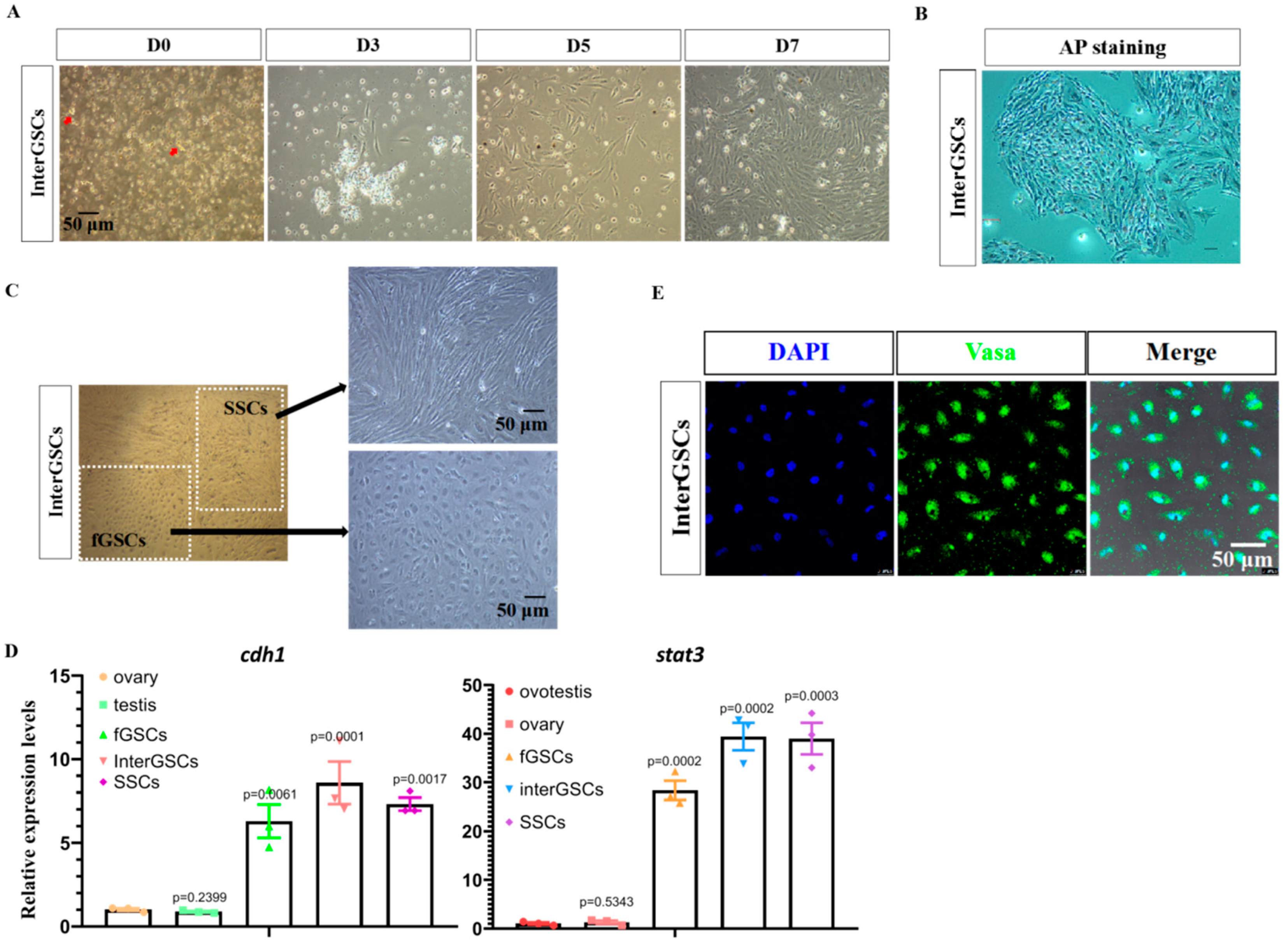
3.3. Isolation and Culturing of interGSCs
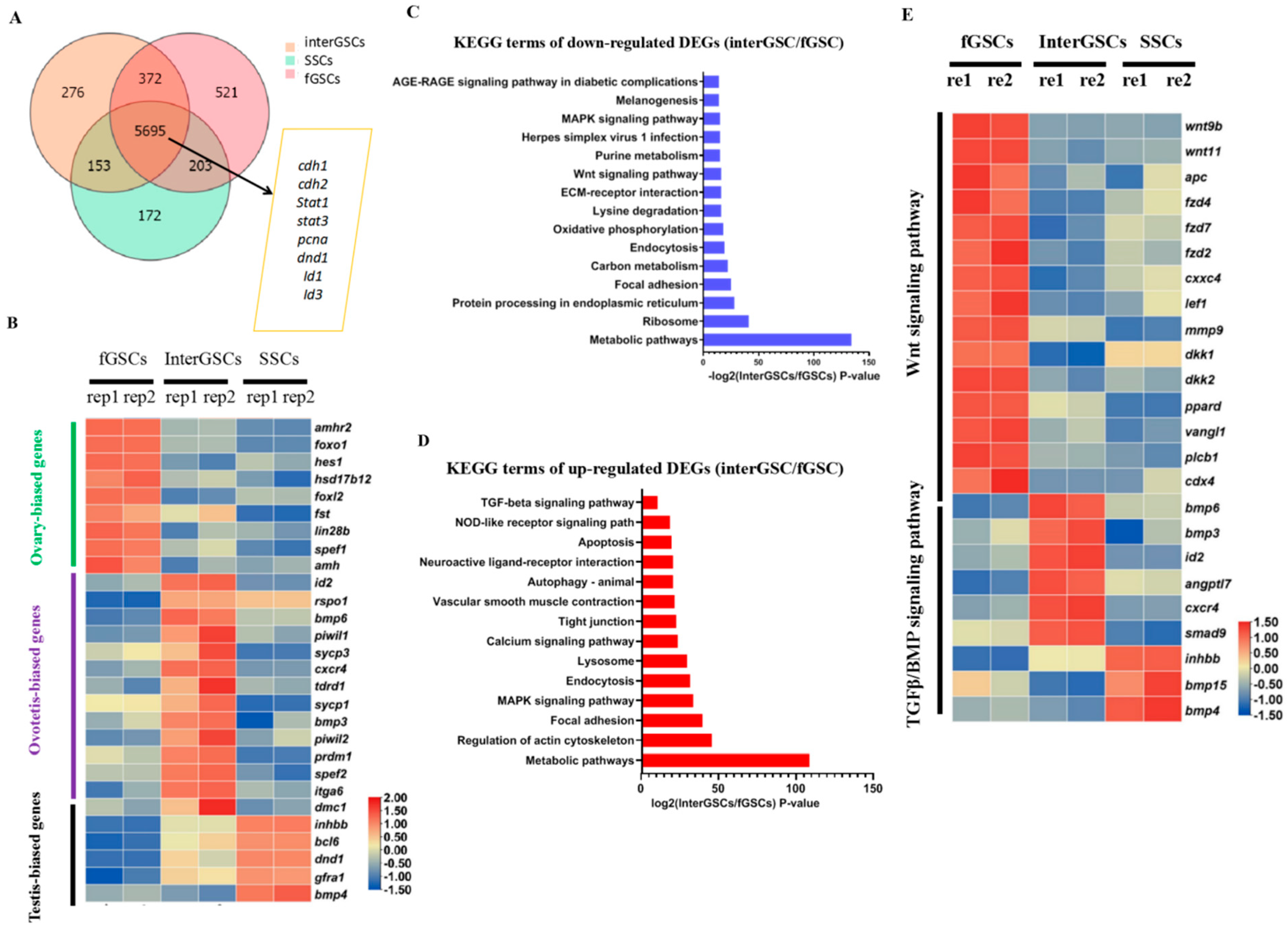
3.4. Transcriptome Comparison of fGSCs, interGSCs, and SSCs
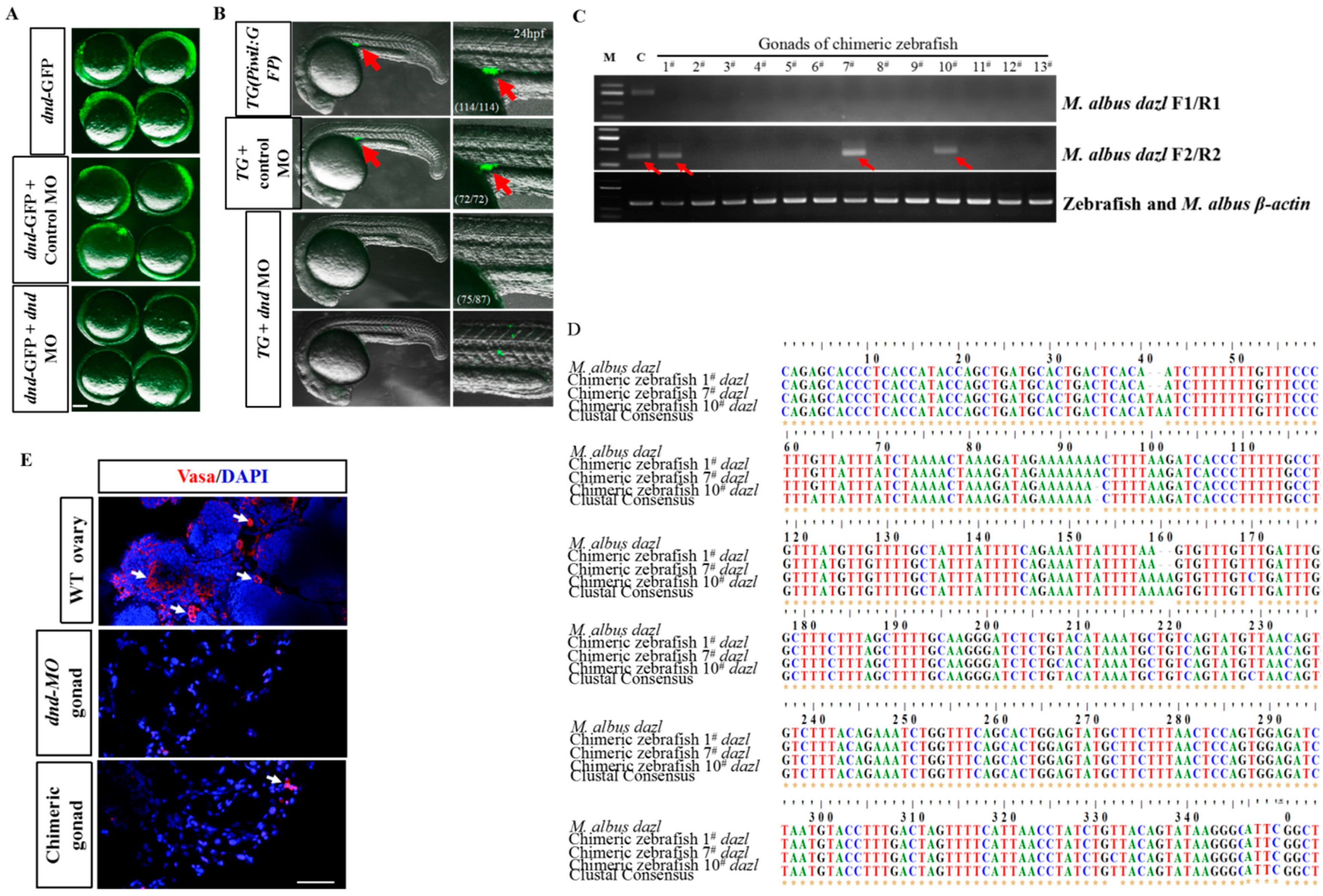
3.5. fGSCs’ Ability to Colonize the Recipient Zebrafish and Contribute to the Germline Cell Lineage after Transplantation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tao, Y.X.; Lin, H.R.; Vanderkraak, G.; Peter, R.E. Hormonal Induction of Precocious Sex Reversal in the Ricefield Eel, Monopterus-Albus. Aquaculture 1993, 118, 131–140. [Google Scholar] [CrossRef]
- Zhou, H.; Hu, Y.; Jiang, H.; Duan, G.; Ling, J.; Pan, T.; Chen, X.; Wang, H.; Zhang, Y. Population genetics of swamp eel in the Yangtze River: Comparative analyses between mitochondrial and microsatellite data provide novel insights. PeerJ 2020, 8, e8415. [Google Scholar] [CrossRef] [PubMed]
- Chi, W.; Gao, Y.; Hu, Q.; Guo, W.; Li, D.P. Genome-wide analysis of brain and gonad transcripts reveals changes of key sex reversal-related genes expression and signaling pathways in three stages of Monopterus albus. PLoS ONE 2017, 12, e0173974. [Google Scholar] [CrossRef] [PubMed]
- Ip, Y.K.; Tay, A.S.; Lee, K.H.; Chew, S.F. Strategies for surviving high concentrations of environmental ammonia in the swamp eel Monopterus albus. Physiol. Biochem. Zool. 2004, 77, 390–405. [Google Scholar] [CrossRef]
- Liang, H.W.; Guo, S.S.; Li, Z.; Luo, X.Z.; Zou, G.W. Assessment of genetic diversity and population structure of swamp eel Monopterus albus in China. Biochem. Syst. Ecol. 2016, 68, 81–87. [Google Scholar] [CrossRef]
- Liu, L.; Zhou, Q.; Lin, C.; He, L.; Wei, L. Histological alterations, oxidative stress, and inflammatory response in the liver of swamp eel (Monopterus albus) acutely exposed to copper. Fish Physiol. Biochem. 2021, 47, 1865–1878. [Google Scholar] [CrossRef]
- Tay, A.S.; Chew, S.F.; Ip, Y.K. The swamp eel Monopterus albus reduces endogenous ammonia production and detoxifies ammonia to glutamine during 144 h of aerial exposure. J. Exp. Biol. 2003, 206, 2473–2486. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, R. Germline stem cells: Origin and destiny. Cell Stem Cell. 2012, 10, 729–739. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Yu, M.; Hu, Y.; Qiu, P.; Liu, W.; Zheng, W.; Peng, S.; Hua, J. Location and characterization of female germline stem cells (FGSCs) in juvenile porcine ovary. Cell Prolif. 2013, 46, 516–528. [Google Scholar] [CrossRef]
- Hong, Y.H.; Liu, T.M.; Zhao, H.B.; Xu, H.Y.; Wang, W.J.; Liu, R.; Chen, T.S.; Deng, J.R.; Gui, J.F. Establishment of a normal medakafish spermatogonial cell line capable of sperm production in vitro. Proc. Natl. Acad. Sci. USA 2004, 101, 8011–8016. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.H.; Chin, C.C.; Ho, H.N.; Chou, C.K.; Shen, C.N.; Kuo, H.C.; Wu, T.J.; Wu, Y.C.; Hung, Y.C.; Chang, C.C.; et al. Pluripotency of mouse spermatogonial stem cells maintained by IGF-1- dependent pathway. FASEB J. 2009, 23, 2076–2087. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Li, C.J.; Yue, H.M.; Du, H.; Yang, X.G.; Yoshino, T.; Hayashida, T.; Takeuchi, Y.; Wei, Q.W. Establishment of intraperitoneal germ cell transplantation for critically endangered Chinese sturgeon Acipenser sinensis. Theriogenology 2017, 94, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Zou, K.; Hou, L.; Sun, K.J.; Xie, W.H.; Wu, J. Improved Efficiency of Female Germline Stem Cell Purification Using Fragilis-Based Magnetic Bead Sorting. Stem Cells Dev. 2011, 20, 2197–2204. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.; Yoshihara, M.; Douagi, I.; Damdimopoulos, A.; Panula, S.; Petropoulos, S.; Lu, H.; Pettersson, K.; Palm, K.; Katayama, S.; et al. Single-cell analysis of human ovarian cortex identifies distinct cell populations but no oogonial stem cells. Nat. Commun. 2020, 11, 1147. [Google Scholar] [CrossRef] [Green Version]
- Shang, M.; Su, B.; Perera, D.A.; Alsaqufi, A.; Lipke, E.A.; Cek, S.; Dunn, D.A.; Qin, Z.; Peatman, E.; Dunham, R.A. Testicular germ line cell identification, isolation, and transplantation in two North American catfish species. Fish Physiol. Biochem. 2018, 44, 717–733. [Google Scholar] [CrossRef]
- Wong, T.T.; Tesfamichael, A.; Collodi, P. Production of zebrafish offspring from cultured female germline stem cells. PLoS ONE 2013, 8, e62660. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Lai, F.L.; Shang, D.T.; Cheng, Y.B.; Lan, T.; Cheng, H.H.; Zhou, R.J. Cellular fate of intersex differentiation. Cell Death Dis. 2021, 12, 388. [Google Scholar] [CrossRef]
- Chau, J.; Kulnane, L.S.; Salz, H.K. Sex-lethal enables germline stem cell differentiation by down-regulating Nanos protein levels during Drosophila oogenesis. Proc. Natl. Acad. Sci. USA 2012, 109, 9465–9470. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Wu, J.; Yang, W.; Xia, C.; Shi, X.; Li, H.; Sun, J.; Shao, Z.; Zhao, X. STAT3 is required for proliferation and exhibits a cell type-specific binding preference in mouse female germline stem cells. Mol. Omics. 2018, 14, 95–102. [Google Scholar] [CrossRef]
- Kraemer, B.; Crittenden, S.; Gallegos, M.; Moulder, G.; Barstead, R.; Kimble, J.; Wickens, M. NANOS-3 and FBF proteins physically interact to control the sperm-oocyte switch in Caenorhabditis elegans. Curr. Biol. 1999, 9, 1009–1018. [Google Scholar] [CrossRef] [Green Version]
- Li, M.Y.; Zhu, F.; Li, Z.D.; Hong, N.; Hong, Y.H. Dazl is a critical player for primordial germ cell formation in medaka. Sci. Rep. 2016, 6, 28317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mcaninch, D.; Makela, J.A.; La, H.M.; Hughes, J.N.; Lovell-Badge, R.; Hobbs, R.M.; Thomas, P.Q. SOX3 promotes generation of committed spermatogonia in postnatal mouse testes. Sci. Rep. 2020, 10, 6751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oatley, J.M.; Kaucher, A.V.; Avarbock, M.R.; Brinster, R.L. Regulation of mouse spermatogonial stem cell differentiation by STAT3 signaling. Biol. Reprod. 2010, 83, 427–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Presslauer, C.; Nagasawa, K.; Fernandes, J.M.O.; Babiak, I. Expression of vasa and nanos3 during primordial germ cell formation and migration in Atlantic cod (Gadus morhua L.). Theriogenology 2012, 78, 1262–1277. [Google Scholar] [CrossRef] [PubMed]
- Sada, A.; Suzuki, A.; Suzuki, H.; Saga, Y. The RNA-binding protein NANOS2 is required to maintain murine spermatogonial stem cells. Science 2009, 325, 1394–1398. [Google Scholar] [CrossRef] [Green Version]
- Su, R.; Cao, S.; Ma, J.; Liu, Y.; Liu, X.; Zheng, J.; Chen, J.; Liu, L.; Cai, H.; Li, Z.; et al. Knockdown of SOX2OT inhibits the malignant biological behaviors of glioblastoma stem cells via up-regulating the expression of miR-194-5p and miR-122. Mol. Cancer 2017, 16, 171. [Google Scholar] [CrossRef]
- Tokuda, M.; Kadokawa, Y.; Kurahashi, H.; Marunouchi, T. CDH1 is a specific marker for undifferentiated spermatogonia in mouse testes. Biol. Reprod. 2007, 76, 130–141. [Google Scholar] [CrossRef] [Green Version]
- Yamaji, M.; Jishage, M.; Meyer, C.; Suryawanshi, H.; Der, E.; Garzia, A.; Morozov, P.; Manickavel, S.; Mcfarland, H.L.; Roeder, R.G.; et al. DND1 maintains germline stem cells via recruitment of the CCR4-NOT complex to target mRNAs. Nature 2017, 543, 568–572. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, S.; Luo, F.H.; Baiyinbatu; Liu, L.H.; Hu, T.Y.; Yu, B.Y.; Li, G.P.; Wu, Y.J. CDH1, a Novel Surface Marker of Spermatogonial Stem Cells in Sheep Testis. J. Integr. Agric. 2014, 13, 1759–1765. [Google Scholar] [CrossRef]
- La, H.M.; Liao, J.; Legrand, J.M.D.; Rossello, F.J.; Chan, A.L.; Vaghjiani, V.; Cain, J.E.; Papa, A.; Lee, T.L.; Hobbs, R.M. Distinctive molecular features of regenerative stem cells in the damaged male germline. Nat. Commun. 2022, 13, 2500. [Google Scholar] [CrossRef]
- Huang, X.; Guo, Y.; Shui, Y.; Gao, S.; Yu, H.; Cheng, H.; Zhou, R. Multiple alternative splicing and differential expression of dmrt1 during gonad transformation of the rice field eel. Biol. Reprod. 2005, 73, 1017–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.; Cai, H.; Zhang, G.; Zhang, H.; Bao, H.; Wang, L.; Ye, J.; Qian, G.; Ge, C. Dmrt1 is required for primary male sexual differentiation in Chinese soft-shelled turtle Pelodiscus sinensis. Sci. Rep. 2017, 7, 4433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, K.A.; Schach, U.; Ordaz, A.; Steinfeld, J.S.; Draper, B.W.; Siegfried, K.R. Dmrt1 is necessary for male sexual development in zebrafish. Dev. Biol. 2017, 422, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, M.; Ma, H.; Liu, X.; Shi, H.; Wang, D. Mutation of foxl2 or cyp19a1a Results in Female to Male Sex Reversal in XX Nile Tilapia. Endocrinology 2017, 158, 2634–2647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulanger, L.; Pannetier, M.; Gall, L.; Allais-Bonnet, A.; Elzaiat, M.; Le Bourhis, D.; Daniel, N.; Richard, C.; Cotinot, C.; Ghyselinck, N.B.; et al. FOXL2 Is a Female Sex-Determining Gene in the Goat. Curr. Biol. 2014, 24, 404–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biason-Lauber, A. WNT4, RSPO1, and FOXL2 in sex development. Semin. Reprod. Med. 2012, 30, 387–395. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.Q.; Zhang, L.J.; Hu, Y.Q.; Chen, M.; Han, F.; Qin, Y.; Chen, M.; Cui, X.H.; Duo, S.G.; Tang, F.C.; et al. Beta-catenin directs the transformation of testis Sertoli cells to ovarian granulosa-like cells by inducing Foxl2 expression. J. Biol. Chem. 2017, 292, 17577–17586. [Google Scholar] [CrossRef] [Green Version]
- Ottolenghi, C.; Pelosi, E.; Tran, J.; Colombino, M.; Douglass, E.; Nedorezov, T.; Cao, A.; Forabosco, A.; Schlessinger, D. Loss of Wnt4 and Foxl2 leads to female-to-male sex reversal extending to germ cells. Hum. Mol. Genet. 2007, 16, 2795–2804. [Google Scholar] [CrossRef] [Green Version]
- Ye, D.; Sun, Y. Manipulating and Visualizing the Germline with Transgenic Lines. Methods Mol. Biol. 2021, 2218, 265–276. [Google Scholar]
- Maatouk, D.M.; Dinapoli, L.; Alvers, A.; Parker, K.L.; Taketo, M.M.; Capel, B. Stabilization of beta-catenin in XY gonads causes male-to-female sex-reversal. Hum. Mol. Genet. 2008, 17, 2949–2955. [Google Scholar] [CrossRef] [Green Version]
- Tomizuka, K.; Horikoshi, K.; Kitada, R.; Sugawara, Y.; Iba, Y.; Kojima, A.; Yoshitome, A.; Yamawaki, K.; Amagai, M.; Inoue, A.; et al. R-spondin1 plays an essential role in ovarian development through positively regulating Wnt-4 signaling. Hum. Mol. Genet. 2008, 17, 1278–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chassot, A.A.; Ranc, F.; Gregoire, E.P.; Roepers-Gajadien, H.L.; Taketo, M.M.; Camerino, G.; De Rooij, D.G.; Schedl, A.; Chaboissier, M.C. Activation of beta-catenin signaling by Rspo1 controls differentiation of the mammalian ovary. Hum. Mol. Genet. 2008, 17, 1264–1277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Charkraborty, T.; Zhou, Q.; Mohapatra, S.; Nagahama, Y.; Zhang, Y. Rspo1-activated signalling molecules are sufficient to induce ovarian differentiation in XY medaka (Oryzias latipes). Sci. Rep. 2016, 6, 19543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyer, A.; Goff, A.K.; Boerboom, D. WNT signaling in ovarian follicle biology and tumorigenesis. Trends Endocrinol. Metab. 2010, 21, 25–32. [Google Scholar] [CrossRef]
- Munger, S.C.; Natarajan, A.; Looger, L.L.; Ohler, U.; Capel, B. Fine time course expression analysis identifies cascades of activation and repression and maps a putative regulator of mammalian sex determination. PLoS Genet. 2013, 9, e1003630. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Bai, Y.; Chu, Z.; Wang, J.; Wang, L.; Yu, M.; Lian, Z.; Hua, J. GSK3 inhibitor-BIO regulates proliferation of female germline stem cells from the postnatal mouse ovary. Cell Prolif. 2012, 45, 287–298. [Google Scholar] [CrossRef]
- Kleber, M.; Lee, H.Y.; Wurdak, H.; Buchstaller, J.; Riccomagno, M.M.; Ittner, L.M.; Suter, U.; Epstein, D.J.; Sommer, L. Neural crest stem cell maintenance by combinatorial Wnt and BMP signaling. J. Cell. Biol. 2005, 169, 309–320. [Google Scholar] [CrossRef] [Green Version]
- Mottier-Pavie, V.I.; Palacios, V.; Eliazer, S.; Scoggin, S.; Buszczak, M. The Wnt pathway limits BMP signaling outside of the germline stem cell niche in Drosophila ovaries. Dev. Biol. 2016, 417, 50–62. [Google Scholar] [CrossRef]
- Wei, X.F.; Chen, Q.L.; Fu, Y.; Zhang, Q.K. Wnt and BMP signaling pathways co-operatively induce the differentiation of multiple myeloma mesenchymal stem cells into osteoblasts by upregulating EMX2. J. Cell. Biochem. 2019, 120, 6515–6527. [Google Scholar] [CrossRef]
- Song, X.; Wong, M.D.; Kawase, E.; Xi, R.; Ding, B.C.; Mccarthy, J.J.; Xie, T. Bmp signals from niche cells directly repress transcription of a differentiation-promoting gene, bag of marbles, in germline stem cells in the Drosophila ovary. Development 2004, 131, 1353–1364. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Hao, Y.; Li, X.; Li, Y.; Ye, D.; Zhang, R.; Wang, X.; He, M.; Wang, H.; Zhu, Z.; et al. Surrogate production of genome-edited sperm from a different subfamily by spermatogonial stem cell transplantation. Sci. China Life Sci. 2021, 64, 969–987. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward (5’-3’) | Reverse (5’-3’) |
|---|---|---|
| elfa | CGCTGCTGTTTCCTTCGTCC | TTGCGTTCAATCTTCCATCCC |
| nanog | TACGAACTGGGTGTGTGAGC | GGCCAGATAAAAGGCCAGGT |
| klf4 | TACCACCATAGCTCCCCACA | GTAGGTTTTCCCACAGCCGA |
| sox2 | TCCATGTCCTACTCCCAGCA | CATGTCCCTCAGATCTCCGC |
| sox3 | ATGAACGCAGCTTCCACGTA | AGTCCCTGCGGTCTGATAGT |
| gfra1 | TGGCTTCTCGTTCCAGATGT | AAGTTGCTCTCCTTACCGCC |
| dazl | GTCCCATCTGGTTGGTCCAG | TCTGGAAATGGGGTGCAACA |
| stat3 | ATCCAGTCAGTGGAGCCCTA | AGTCCATGAACACGGAGGGA |
| cdh1 | ACTCCCACAAAGAATGACTTCAC | AAAAGGGAATCTTGTGCGGC |
| ddx4 | GCTGGATGAAGCTGACAGGAT | AAGGGCCAGAGTTTACCAGT |
| dmrt1a | CTCGCTGGTTAGCTCTGAT | GGAATATGAAACTATCACAAG |
| foxl2 | TGACAACAACACGAACAAGGAG | GGCAATGAGAGCGACATAGGA |
| sox9a | GTGAAGAACGGACAGAGCGA | TCGCTGCTGAACTCACCAAT |
| foxo1 | CACAGCAGAGCAGCCTCC | CATCTGCCTCCCCAAGATCG |
| prdm1 | AGTGTTCAGTTGAAGCCCCC | GGTGAACGAGTTGCAGGGTA |
| tdrd1 | TGCAAGCGCTGCAAGAAAAT | ATGCCTGGATGCCTGTTCTC |
| smad9 | CCAGAGCACTGGTGTTCCAT | TTGCCTATGTGTCTGCGTGT |
| id2 | ACGACTGCTACTCCAAGCTG | GTCCGGCTGTCATCTGTCAT |
| bmp6 | CGATCCACACAACCACAGGA | TGTCTGCACAATAGCGTGGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.; Tao, B.; Wang, Y.; Hu, W.; Sun, Y. Isolation and Characterization of Germline Stem Cells in Protogynous Hermaphroditic Monopterus albus. Int. J. Mol. Sci. 2022, 23, 5861. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23115861
Sun X, Tao B, Wang Y, Hu W, Sun Y. Isolation and Characterization of Germline Stem Cells in Protogynous Hermaphroditic Monopterus albus. International Journal of Molecular Sciences. 2022; 23(11):5861. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23115861
Chicago/Turabian StyleSun, Xiaoyun, Binbin Tao, Yongxin Wang, Wei Hu, and Yuhua Sun. 2022. "Isolation and Characterization of Germline Stem Cells in Protogynous Hermaphroditic Monopterus albus" International Journal of Molecular Sciences 23, no. 11: 5861. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23115861