
The Comparison of the Efficiency of Emodin and Aloe-Emodin in Photodynamic Therapy


Abstract
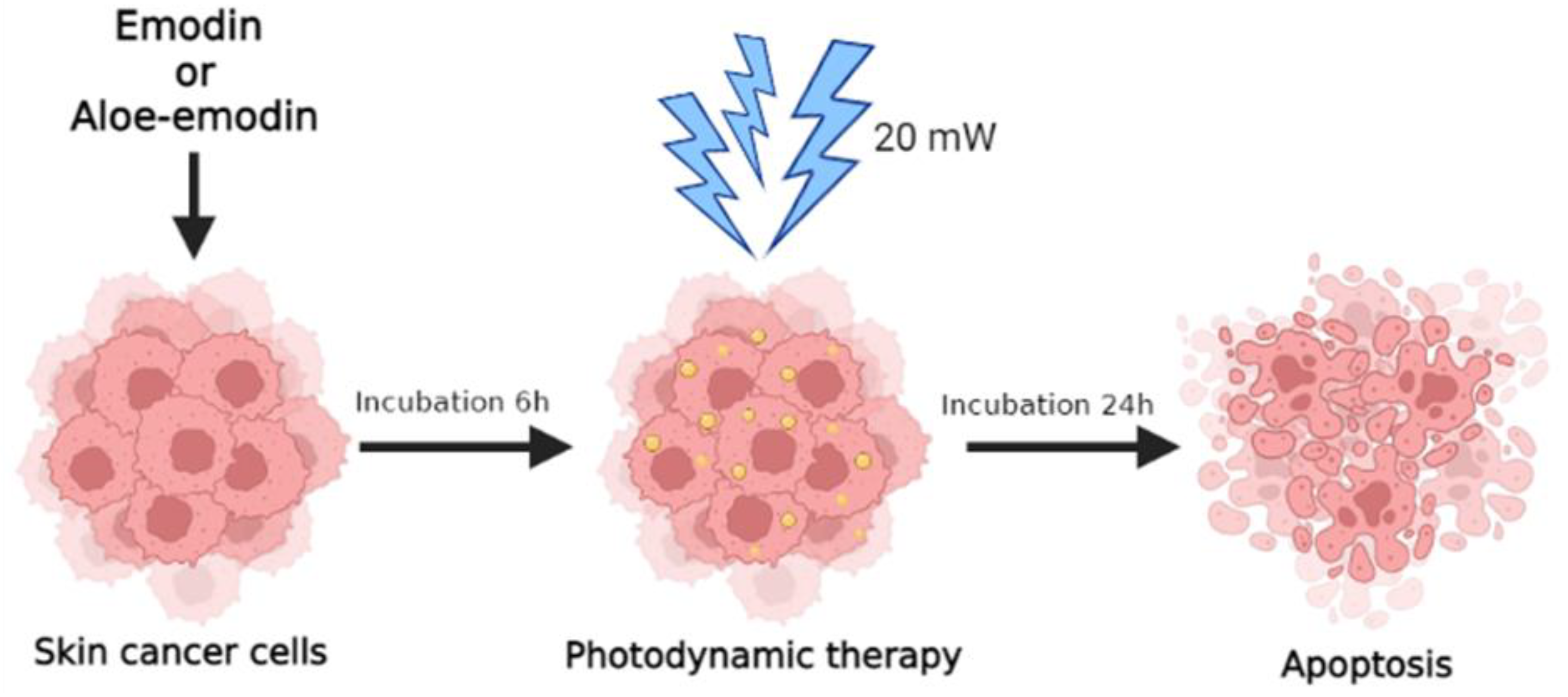
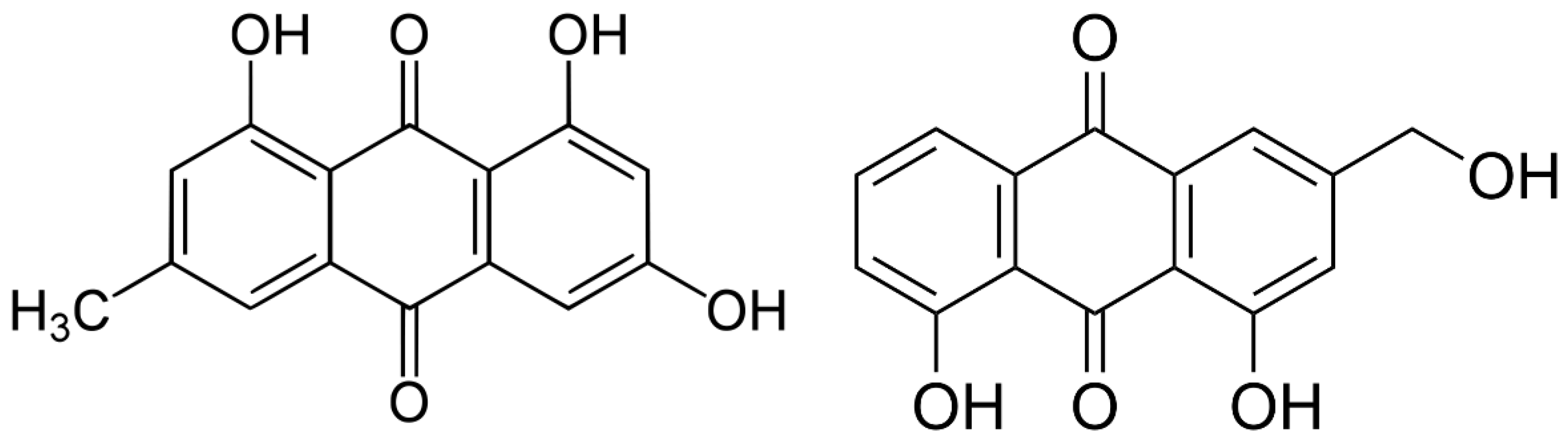
:1. Introduction
2. Results
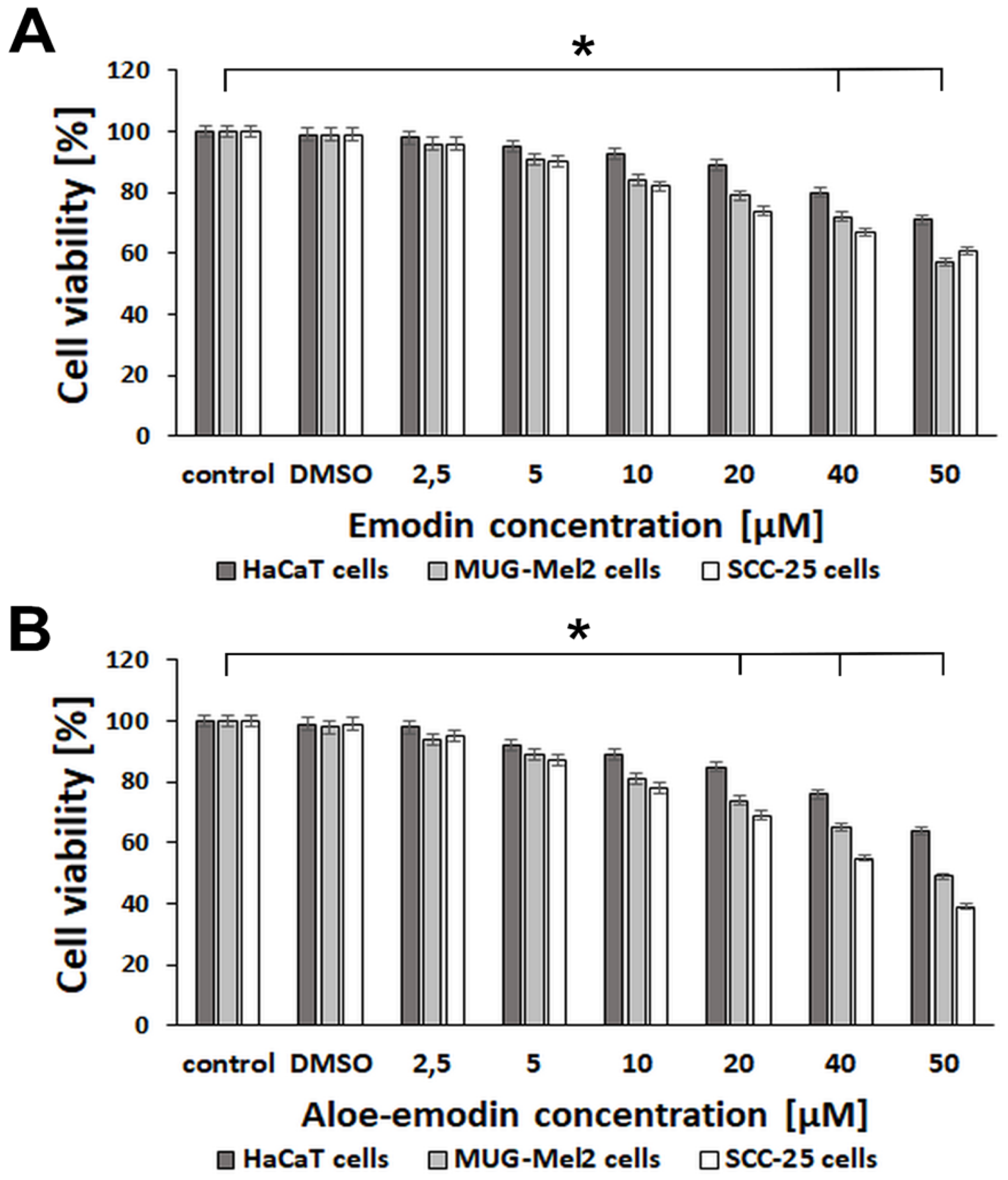
2.1. The Effect of Emodin and Aloe-Emodin on HaCaT, SCC-25, and MUG-Mel2 Cells in MTT Assay
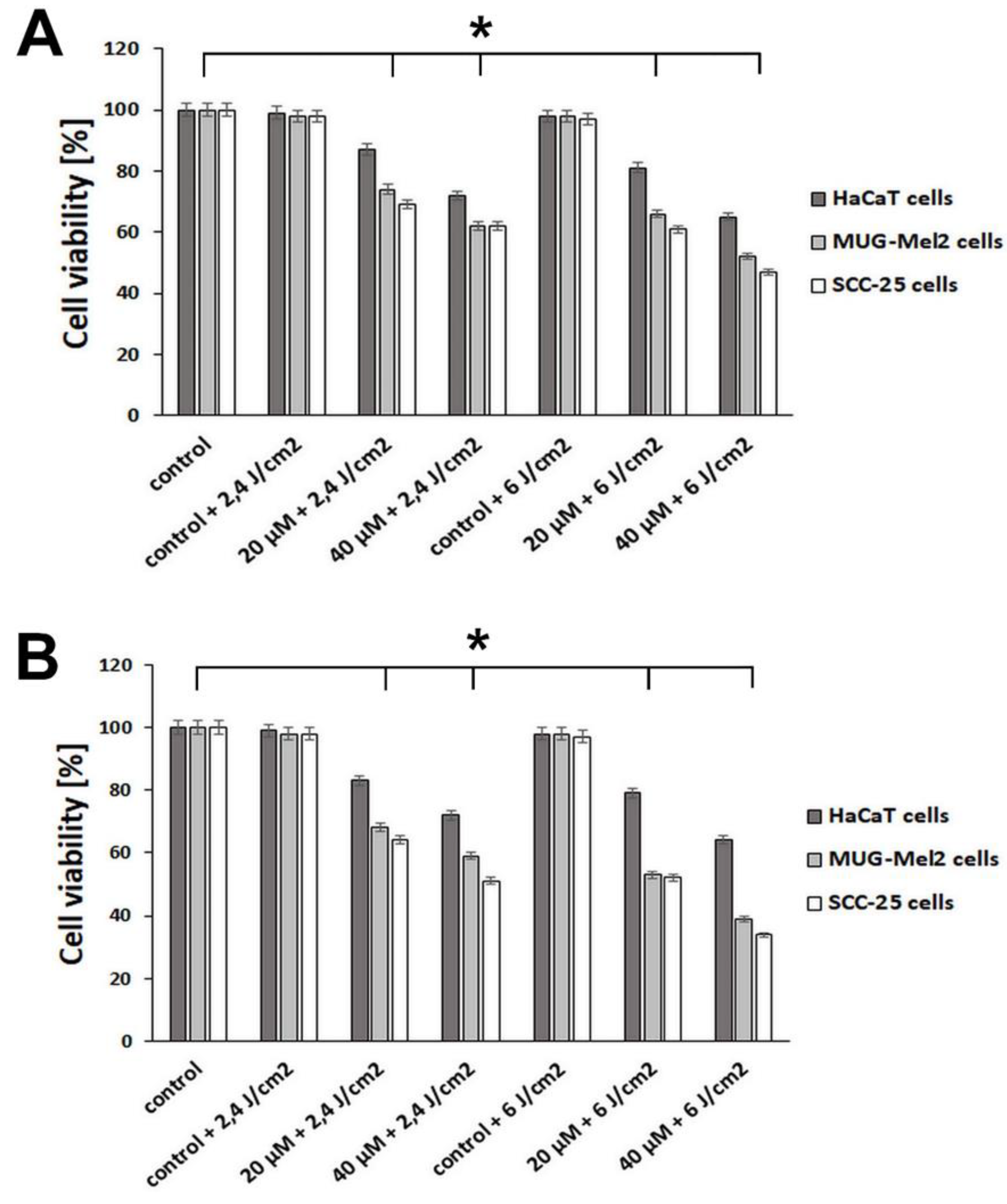
2.2. The Effect of Emodin-Based PDT and Aloe-Emodin-Based PDT on HaCaT, SCC-25, and MUG-Mel2 Cells in MTT Assay
2.3. The Effect of Emodin-Based PDT and Aloe-Emodin-Based PDT on HaCaT, SCC-25, and MUG-Mel2 Cells in the Wound-Healing Process
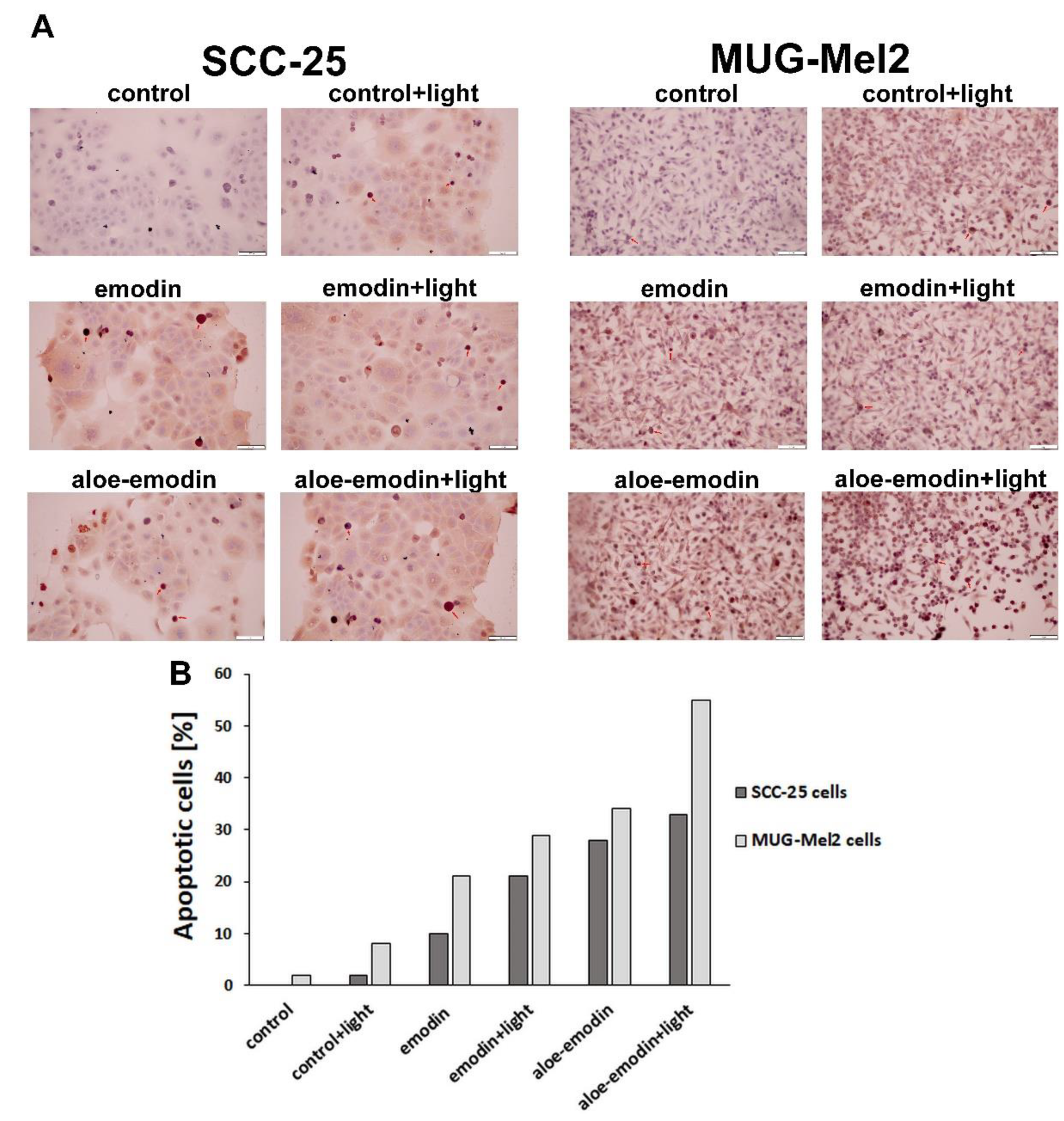
2.4. Tunel Assay—The Impact of Emodin, Emodin-Based PDT, Aloe-Emodin, and Aloe-Emodin-Based PDT on SCC-25 and MUG-Mel2 Cell Lines
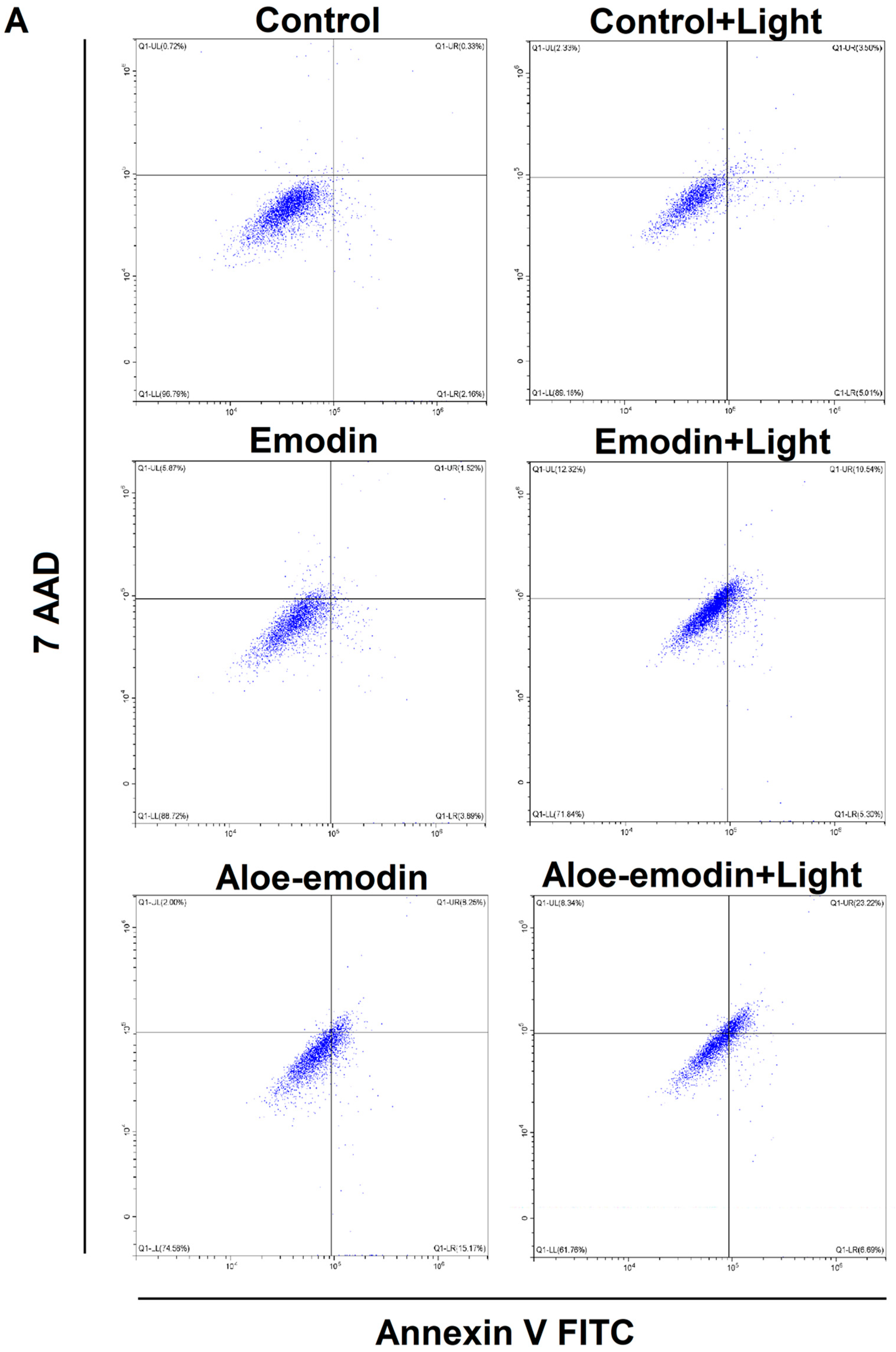
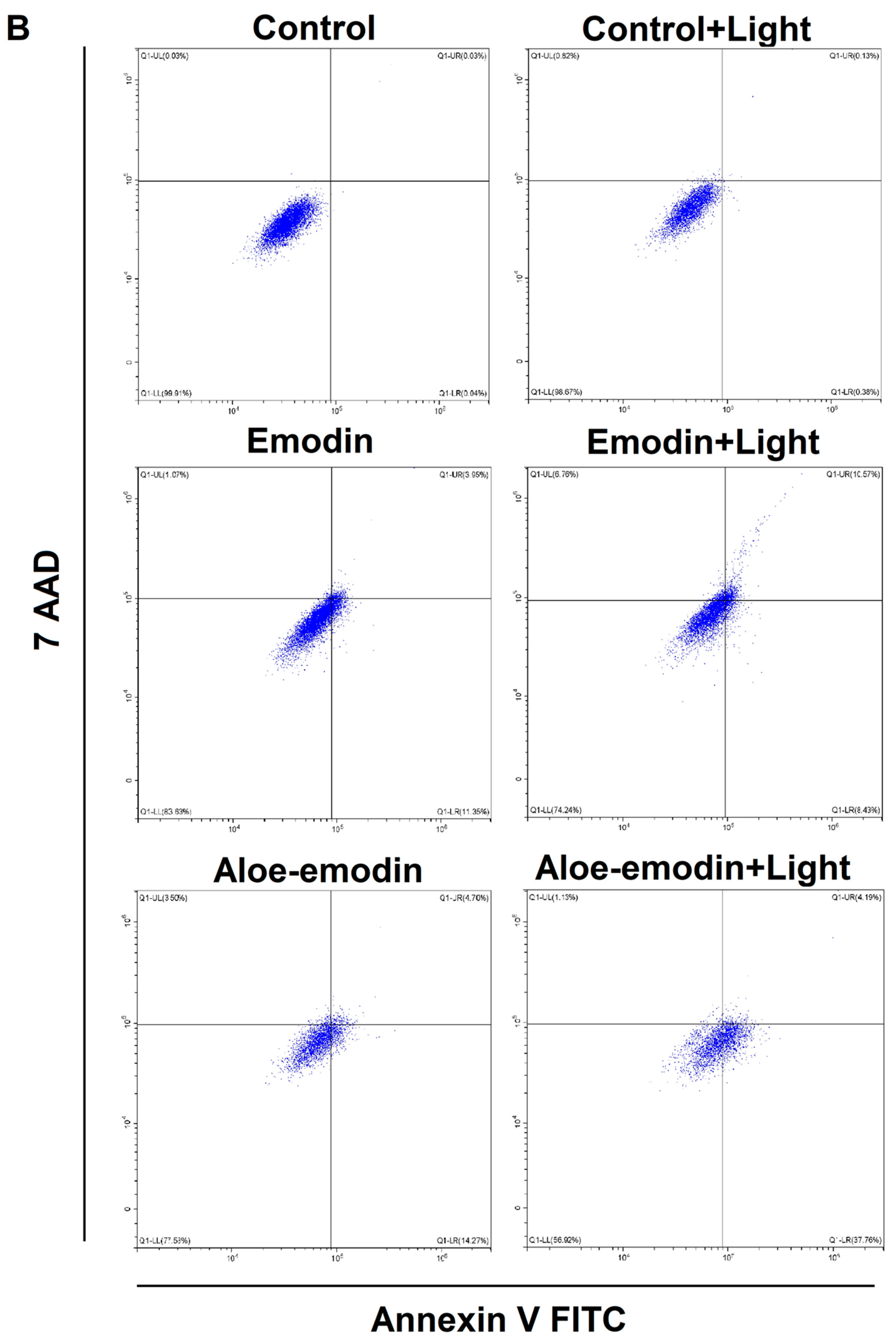
2.5. Apoptosis Assay—The Impact of Emodin, Emodin-Based PDT, Aloe-Emodin, and Aloe-Emodin-Based PDT on SCC-25 and MUG-Mel2 Cell Lines
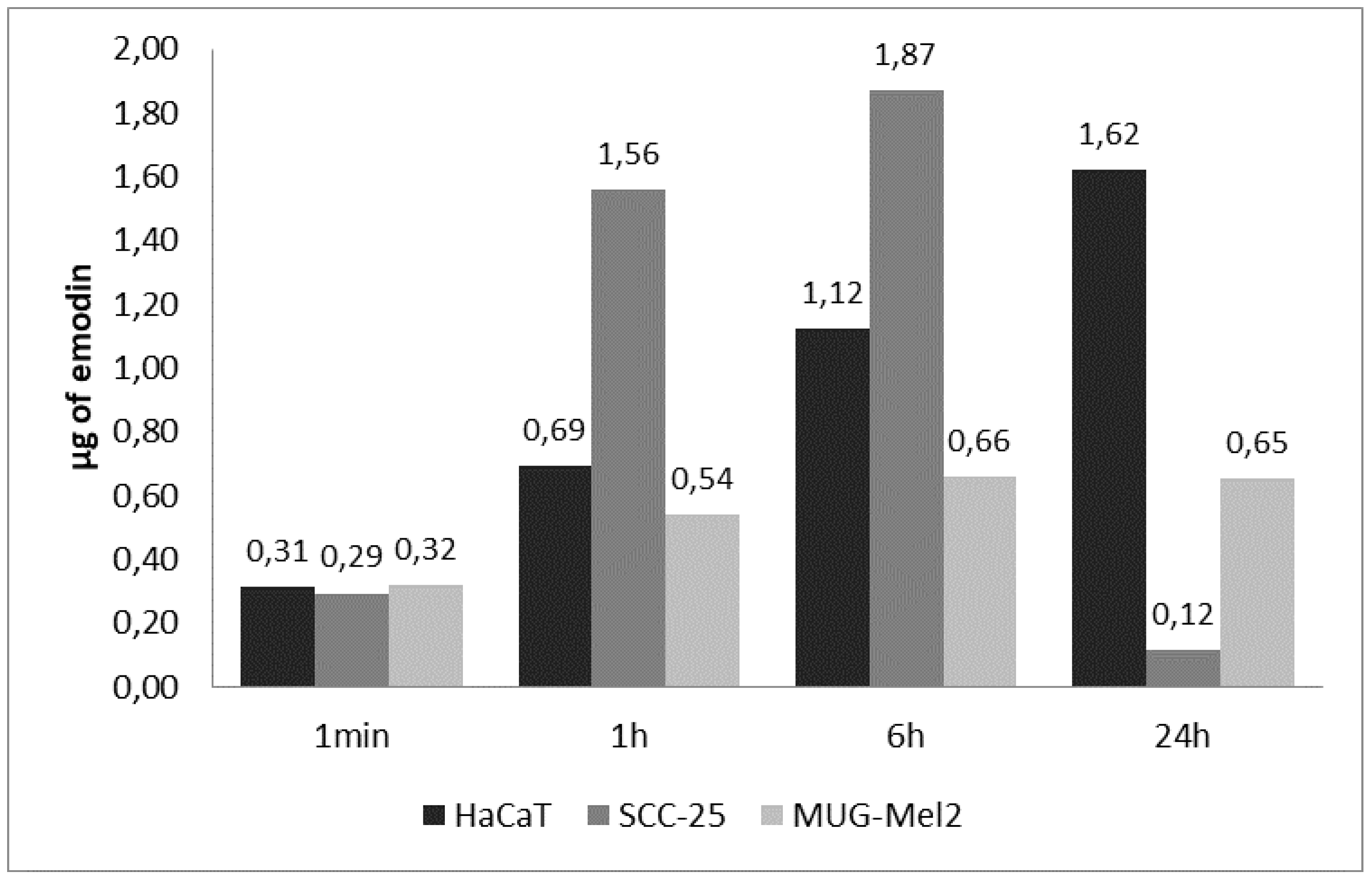
2.6. Results of Cellular Uptake with Emodin and Aloe-Emodin on SCC-25 and MUG-Mel2 Cell Lines
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Emodin Solution Preparation
4.3. Aloe-Emodin Solution Preparation
4.4. PDT Experiment
4.5. Cell Viability Assay—MTT Assay
4.6. Wound Healing Assay
4.7. Tunel Assay—Apoptosis Assay
4.8. Flow Cytometry—Apoptosis Assay
4.9. Cellular Uptake
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Apalla, Z.; Nashan, D.; Weller, R.B. Skin Cancer: Epidemiology, Disease Burden, Pathophysiology, Diagnosis, and Therapeutic Approaches. Dermatol. Ther. 2017, 7, 5–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sajadimajd, S.; Bahramsoltani, R.; Iranpanah, A.; Patra, J.K.; Das, G.; Gouda, S.; Rahimi, R.; Rezaeiamiri, E.; Cao, H.; Giampieri, F.; et al. Advances on Natural Polyphenols as Anticancer Agents for Skin Cancer. Pharmacol. Res. 2020, 151, 104584. [Google Scholar] [CrossRef] [PubMed]
- Simões, M.; Sousa, J.; Pais, A. Skin Cancer and New Treatment Perspectives: A Review. Cancer Lett. 2015, 357, 8–42. [Google Scholar] [CrossRef] [PubMed]
- Cullen, J.K.; Simmons, J.L.; Parsons, P.G.; Boyle, G.M. Topical Treatments for Skin Cancer. Adv. Drug Deliv. Rev. 2020, 153, 54–64. [Google Scholar] [CrossRef]
- Woźniak, M.; Nowak, M.; Lazebna, A.; Więcek, K.; Jabłońska, I.; Szpadel, K.; Grzeszczak, A.; Gubernator, J.; Ziółkowski, P. The Comparison of In Vitro Photosensitizing Efficacy of Curcumin-Loaded Liposomes Following Photodynamic Therapy on Melanoma MUG-Mel2, Squamous Cell Carcinoma SCC-25, and Normal Keratinocyte HaCaT Cells. Pharmaceuticals 2021, 14, 374. [Google Scholar] [CrossRef]
- Akasov, R.A.; Sholina, N.V.; Khochenkov, D.A. Photodynamic Therapy of Melanoma by Blue-Light Photoactivation of Flavin Mononucleotide. Sci. Rep. 2019, 9, 9679. [Google Scholar] [CrossRef] [Green Version]
- Allegra, A.; Pioggia, G.; Tonacci, A.; Musolino, C.; Gangemi, S. Oxidative Stress and Photodynamic Therapy of Skin Cancers: Mechanisms, Challenges and Promising Developments. Antioxidants 2020, 9, 448. [Google Scholar] [CrossRef]
- Kessel, D.; Oleinick, N.L. Cell Death Pathways Associated with Photodynamic Therapy: An Update. Photochem. Photobiol. 2018, 94, 213–218. [Google Scholar] [CrossRef] [Green Version]
- Simões, J.C.S.; Sarpaki, S.; Papadimitroulas, P.; Therrien, B.; Loudos, G. Conjugated Photosensitizers for Imaging and PDT in Cancer Research. J. Med. Chem. 2020, 63, 14119–14150. [Google Scholar] [CrossRef]
- Dong, X.; Fu, J.; Yin, X.; Cao, S.; Li, X.; Lin, L.; Ni, J. Emodin: A Review of Its Pharmacology, Toxicity and Pharmacokinetics. Phytother. Res. 2016, 30, 1207–1218. [Google Scholar] [CrossRef]
- Semwal, R.B.; Semwal, D.K.; Combrinck, S.; Viljoen, A. Emodin—A Natural Anthraquinone Derivative with Diverse Pharmacological Activities. Phytochemistry 2021, 190, 112854. [Google Scholar] [CrossRef]
- Cui, Y.; Chen, L.-J.; Huang, T.; Ying, J.-Q.; Li, J. The pharmacology, toxicology and therapeutic potential of anthraquinone derivative emodin. Chin. J. Nat. Med. 2020, 18, 425–435. [Google Scholar] [CrossRef]
- De Oliveira, M.R.; de Souza, I.C.C.; Brasil, F.B. Mitochondrial Protection and Anti-Inflammatory Effects Induced by Emodin in the Human Neuroblastoma SH-SY5Y Cells Exposed to Hydrogen Peroxide: Involvement of the AMPK/Nrf2 Signaling Pathway. Neurochem. Res. 2021, 46, 482–493. [Google Scholar] [CrossRef]
- Yang, N.; Li, C.; Li, H.; Liu, M.; Cai, X.; Cao, F.; Feng, Y.; Li, M.; Wang, X. Emodin Induced SREBP1-Dependent and SREBP1-Independent Apoptosis in Hepatocellular Carcinoma Cells. Front. Pharmacol. 2019, 10, 709. [Google Scholar] [CrossRef] [Green Version]
- Teng, X.; Wang, S.Y.; Shi, Y.Q.; Fan, X.F.; Liu, S.; Xing, Y.; Guo, Y.Y.; Dong, M. The Role of Emodin on Cisplatin Resistance Reversal of Lung Adenocarcinoma A549/DDP Cell. Anti-Cancer Drugs 2021, 32, 939–949. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, H.; Chen, J.; Hong, Z.; Liao, Y.; Zhang, Q.; Tong, H. Emodin Sensitizes Human Pancreatic Cancer Cells to EGFR Inhibitor through Suppressing Stat3 Signaling Pathway. Cancer Manag. Res. 2019, 11, 8463–8473. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Hodge, J.; Wang, J.; Wang, Y.; Wang, L.; Singh, U.; Li, Y.; Yao, Y.; Wang, D.; Ai, W.; et al. Emodin Reduces Breast Cancer Lung Metastasis by Suppressing Macrophage-Induced Breast Cancer Cell Epithelial-Mesenchymal Transition and Cancer Stem Cell Formation. Theranostics 2020, 10, 8365–8381. [Google Scholar] [CrossRef]
- Liu, C.; Chen, L.; Wang, W.; Qin, D.; Jia, C.; Yuan, M.; Wang, H.; Guo, Y.; Zhu, J.; Zhou, Y.; et al. Emodin Suppresses the Migration and Invasion of Melanoma Cells. Biol. Pharm. Bull. 2021, 44, 771–779. [Google Scholar] [CrossRef]
- Way, T.-D.; Huang, J.-T.; Chou, C.-H.; Huang, C.-H.; Yang, M.-H.; Ho, C.-T. Emodin Represses TWIST1-Induced Epithelial–Mesenchymal Transitions in Head and Neck Squamous Cell Carcinoma Cells by Inhibiting the β-Catenin and Akt Pathways. Eur. J. Cancer 2014, 50, 366–378. [Google Scholar] [CrossRef]
- Sanders, B.; Ray, A.M.; Goldberg, S.; Tyler Clark, H.R.M.; Atlas, S.E.; Farooqi, A.; Konefal, J.; Lages, L.C.; Lopez, J.; Rasul, A.; et al. Anti-Cancer Effects of Aloe-Emodin: A Systematic Review. J. Clin. Transl. Res. 2018, 3, 283–296. [Google Scholar]
- Du, M.; Shen, P.; Tan, R.; Wu, D.; Tu, S. Aloe-Emodin Inhibits the Proliferation, Migration, and Invasion of Melanoma Cells via Inactivation of the Wnt/Beta-Catenin Signaling Pathway. Ann. Transl. Med. 2021, 9, 1722. [Google Scholar] [CrossRef] [PubMed]
- Pecere, T.; Gazzola, M.V.; Mucignat, C.; Parolin, C.; Vecchia, F.D.; Cavaggioni, A.; Basso, G.; Diaspro, A.; Salvato, B.; Carli, M.; et al. Aloe-Emodin Is a New Type of Anticancer Agent with Selective Activity against Neuroectodermal Tumors. Cancer Res. 2000, 60, 2800–2804. [Google Scholar] [PubMed]
- Lee, H.Z.; Hsu, S.L.; Liu, M.C.; Wu, C.H. Effects and mechanisms of aloe-emodin on cell death in human lung squamous cell carcinoma. Eur. J. Pharmacol. 2001, 431, 287–295. [Google Scholar] [CrossRef]
- Kuo, P.L.; Lin, T.C.; Lin, C.C. The Antiproliferative Activity of Aloe-Emodin Is through P53-Dependent and P21-Dependent Apoptotic Pathway in Human Hepatoma Cell Lines. Life Sci. 2002, 71, 1879–1892. [Google Scholar] [CrossRef]
- Lin, K.-Y.; Uen, Y.-H. Aloe-Emodin, an anthraquinone, in vitro inhibits proliferation and induces apoptosis in human colon carcinoma cells. Oncol. Lett. 2010, 1, 541–547. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wen, J.; Yu, K.; Shu, Y.; He, W.; Chu, H.; Zhang, B.; Ge, C. Aloe-Emodin Induces Apoptosis in Human Oral Squamous Cell Carcinoma SCC15 Cells. BMC Complement. Altern. Med. 2018, 18, 296. [Google Scholar] [CrossRef]
- Mpiana, P.T.; Tshibangu, D.S.; Kilembe, J.T.; Gbolo, B.Z.; Mwanangombo, D.T.; Inkoto, C.L.; Lengbiye, E.M.; Mbadiko, C.M.; Matondo, A.; Bongo, G.N.; et al. Identification of Potential Inhibitors of SARS-CoV-2 Main Protease from Aloe Vera Compounds: A Molecular Docking Study. Chem. Phys. Lett. 2020, 754, 137751. [Google Scholar] [CrossRef]
- Du, H.-X. Revealing the Therapeutic Targets and Molecular Mechanisms of Emodin-Treated Coronavirus Disease 2019 via a Systematic Study of Network Pharmacology. Aging 2021, 13, 14571–14589. [Google Scholar] [CrossRef]
- Liu, Y.; Meng, P.; Zhang, H.; Liu, X.; Wang, M.; Cao, W.; Hu, Z.; Zhang, Z. Inhibitory Effect of Aloe Emodin Mediated Photodynamic Therapy on Human Oral Mucosa Carcinoma in Vitro and in Vivo. Biomed. Pharmacother. 2018, 97, 697–707. [Google Scholar] [CrossRef]
- Galiardi-Campoy, A.E.B.; Machado, F.C.; Carvalho, T.; Tedesco, A.C.; Rahal, P.; Calmon, M.F. Effects of Photodynamic Therapy Mediated by Emodin in Cervical Carcinoma Cells. Photodiagn. Photodyn. Ther. 2021, 35, 102394. [Google Scholar] [CrossRef]
- Mansoori, B.; Mohammadi, A.; Amin Doustvandi, M.; Mohammadnejad, F.; Kamari, F.; Gjerstorff, M.F.; Baradaran, B.; Hamblin, M.R. Photodynamic Therapy for Cancer: Role of Natural Products. Photodiagn. Photodyn. Ther. 2019, 26, 395–404. [Google Scholar] [CrossRef]
- Szlasa, W.; Supplitt, S.; Drąg-Zalesińska, M.; Przystupski, D.; Kotowski, K.; Szewczyk, A.; Kasperkiewicz, P.; Saczko, J.; Kulbacka, J. Effects of Curcumin Based PDT on the Viability and the Organization of Actin in Melanotic (A375) and Amelanotic Melanoma (C32)– in Vitro Studies. Biomed. Pharmacother. 2020, 132, 110883. [Google Scholar] [CrossRef]
- Zang, L.; Zhao, H.; Ji, X.; Cao, W.; Zhang, Z.; Meng, P. Photophysical Properties, Singlet Oxygen Generation Efficiency and Cytotoxic Effects of Aloe Emodin as a Blue Light Photosensitizer for Photodynamic Therapy in Dermatological Treatment. Photochem. Photobiol. Sci. 2017, 16, 1088–1094. [Google Scholar] [CrossRef] [Green Version]
- Vargas, F.; Fraile, G.; Velásquez, M.; Correia, H.; Fonseca, G.; Marín, M.; Marcano, E.; Sánchez, Y. Studies on the Photostability and Phototoxicity of Aloe-Emodin, Emodin and Rhein. Pharmazie 2002, 57, 399–404. [Google Scholar]
- Maytin, E.V.; Kaw, U.; Ilyas, M.; Mack, J.A.; Hu, B. Blue Light versus Red Light for Photodynamic Therapy of Basal Cell Carcinoma in Patients with Gorlin Syndrome: A Bilaterally Controlled Comparison Study. Photodiagn. Photodyn. Ther. 2018, 22, 7–13. [Google Scholar] [CrossRef]
- Colombo, I.; Sangiovanni, E.; Maggio, R.; Mattozzi, C.; Zava, S.; Corbett, Y.; Fumagalli, M.; Carlino, C.; Corsetto, P.A.; Scaccabarozzi, D. HaCaT Cells as a Reliable in Vitro Differentiation Model to Dissect the Inflammatory/Repair Response of Human Keratinocytes. Mediat. Inflamm. 2017, 2017, 7435621. [Google Scholar] [CrossRef]
- Micallef, L.; Belaubre, F.; Pinon, A.; Jayat-Vignoles, C.; Delage, C.; Charveron, M.; Simon, A. Effects of Extracellular Calcium on the Growth-Differentiation Switch in Immortalized Keratinocyte HaCaT Cells Compared with Normal Human Keratinocytes. Exp. Dermatol. 2009, 18, 143–151. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, T.; Wu, L.; Huang, Y.; Mao, Z.; Zhan, Z.; Chen, W.; Dai, F.; Cao, W.; Cao, Y. Metabolism and Toxicity of Emodin: Genome-Wide Association Studies Reveal Hepatocyte Nuclear Factor 4α Regulates UGT2B7 and Emodin Glucuronidation. Chem. Res. Toxicol. 2020, 33, 1798–1808. [Google Scholar] [CrossRef]
- Wu, W.; Hu, N.; Zhang, Q.; Li, Y.; Li, P.; Yan, R.; Wang, Y. In Vitro Glucuronidation of Five Rhubarb Anthraquinones by Intestinal and Liver Microsomes from Humans and Rats. Chem. Biol. Interact. 2014, 219, 18–27. [Google Scholar] [CrossRef]
- Sumida, K.; Kawana, M.; Kouno, E.; Itoh, T.; Takano, S.; Narawa, T.; Tukey, R.H.; Fujiwara, R. Importance of UDP-Glucuronosyltransferase 1A1 Expression in Skin and Its Induction by UVB in Neonatal Hyperbilirubinemia. Mol. Pharmacol. 2013, 84, 679–686. [Google Scholar] [CrossRef] [Green Version]
- Dellinger, R.W.; Matundan, H.H.; Ahmed, A.S.; Duong, P.H.; Meyskens, F.L. Anti-Cancer Drugs Elicit Re-Expression of UDP-Glucuronosyltransferases in Melanoma Cells. PLoS ONE 2012, 7, e47696. [Google Scholar] [CrossRef]
- Chen, Q.; Li, K.T.; Tian, S.; Yu, T.H.; Yu, L.H.; Lin, H.D.; Bai, D.Q. Photodynamic Therapy Mediated by Aloe-Emodin Inhibited Angiogenesis and Cell Metastasis Through Activating MAPK Signaling Pathway on HUVECs. Technol. Cancer Res. Treat. 2018, 17, 1533033818785512. [Google Scholar] [CrossRef] [Green Version]
- Glavnik, V.; Vovk, I. Extraction of Anthraquinones from Japanese Knotweed Rhizomes and Their Analyses by High Performance Thin-Layer Chromatography and Mass Spectrometry. Plants 2020, 9, 1753. [Google Scholar] [CrossRef]
- Li, K.T.; Duan, Q.Q.; Chen, Q.; He, J.W.; Tian, S.; Lin, H.D.; Gao, Q.; Bai, D.Q. The Effect of Aloe Emodin-Encapsulated Nanoliposome-Mediated r-Caspase-3 Gene Transfection and Photodynamic Therapy on Human Gastric Cancer Cells. Cancer Med. 2016, 5, 361–369. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Emodin [ng] | Aloe-Emodin [ng] |
|---|---|---|
| HaCaT | 0.90 | 1.59 |
| SCC-25 | 1.14 | 1.32 |
| MUG-Mel2 | 0.75 | 1.98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nowak-Perlak, M.; Bromke, M.A.; Ziółkowski, P.; Woźniak, M. The Comparison of the Efficiency of Emodin and Aloe-Emodin in Photodynamic Therapy. Int. J. Mol. Sci. 2022, 23, 6276. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116276
Nowak-Perlak M, Bromke MA, Ziółkowski P, Woźniak M. The Comparison of the Efficiency of Emodin and Aloe-Emodin in Photodynamic Therapy. International Journal of Molecular Sciences. 2022; 23(11):6276. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116276
Chicago/Turabian StyleNowak-Perlak, Martyna, Mariusz A. Bromke, Piotr Ziółkowski, and Marta Woźniak. 2022. "The Comparison of the Efficiency of Emodin and Aloe-Emodin in Photodynamic Therapy" International Journal of Molecular Sciences 23, no. 11: 6276. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116276