
Anti-adipogenic and Pro-lipolytic Effects on 3T3-L1 Preadipocytes by CX-4945, an Inhibitor of Casein Kinase 2

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
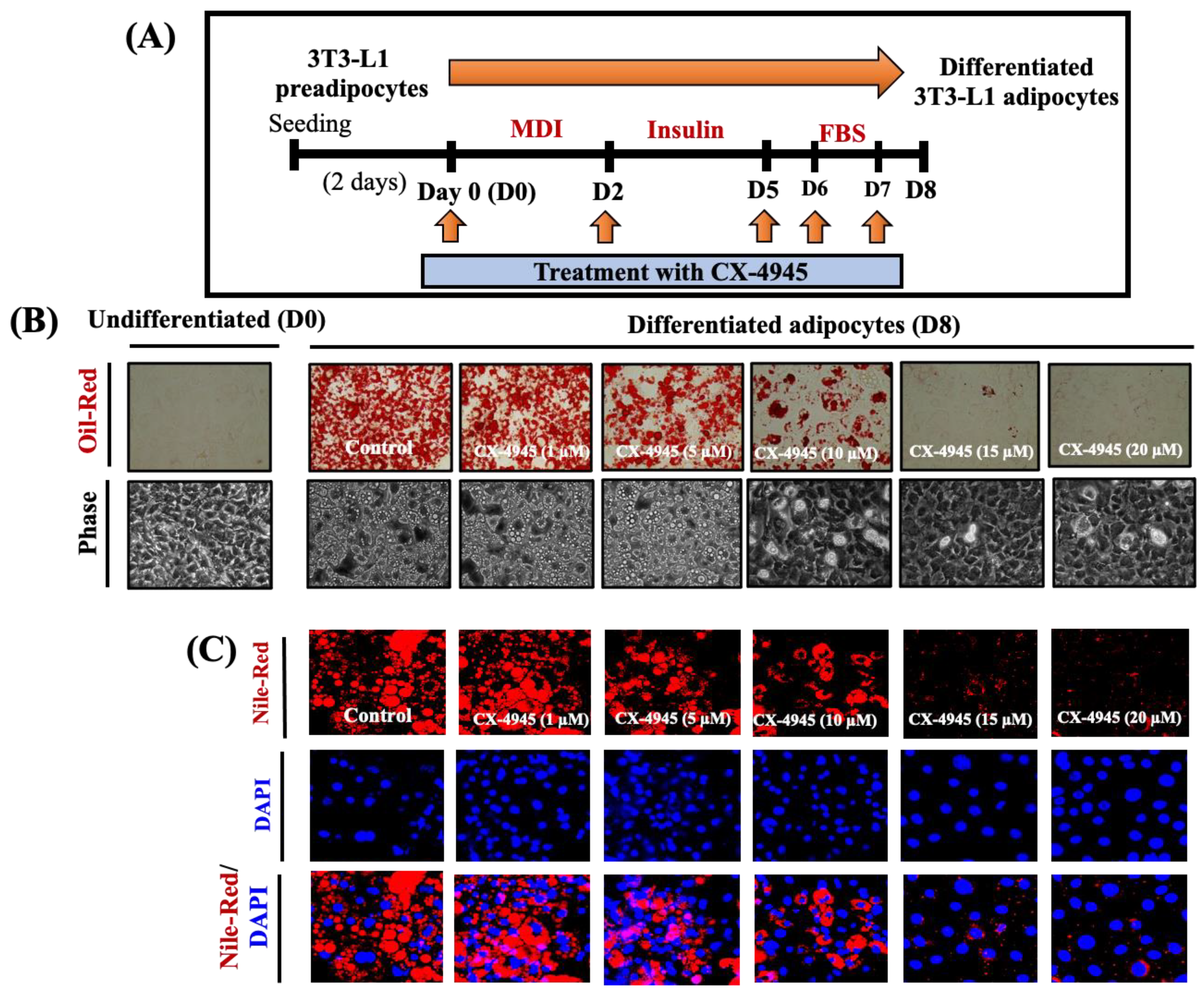
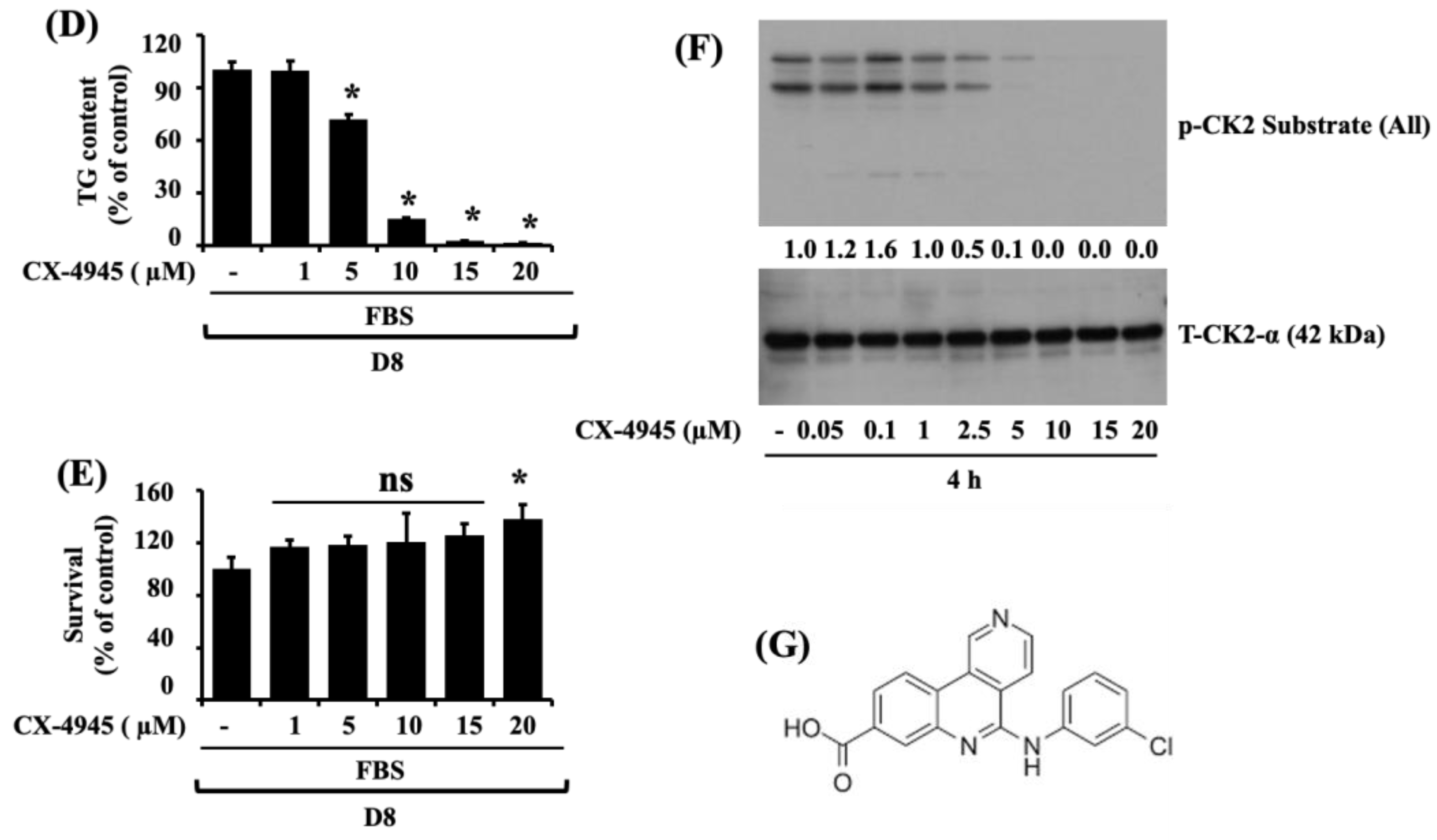
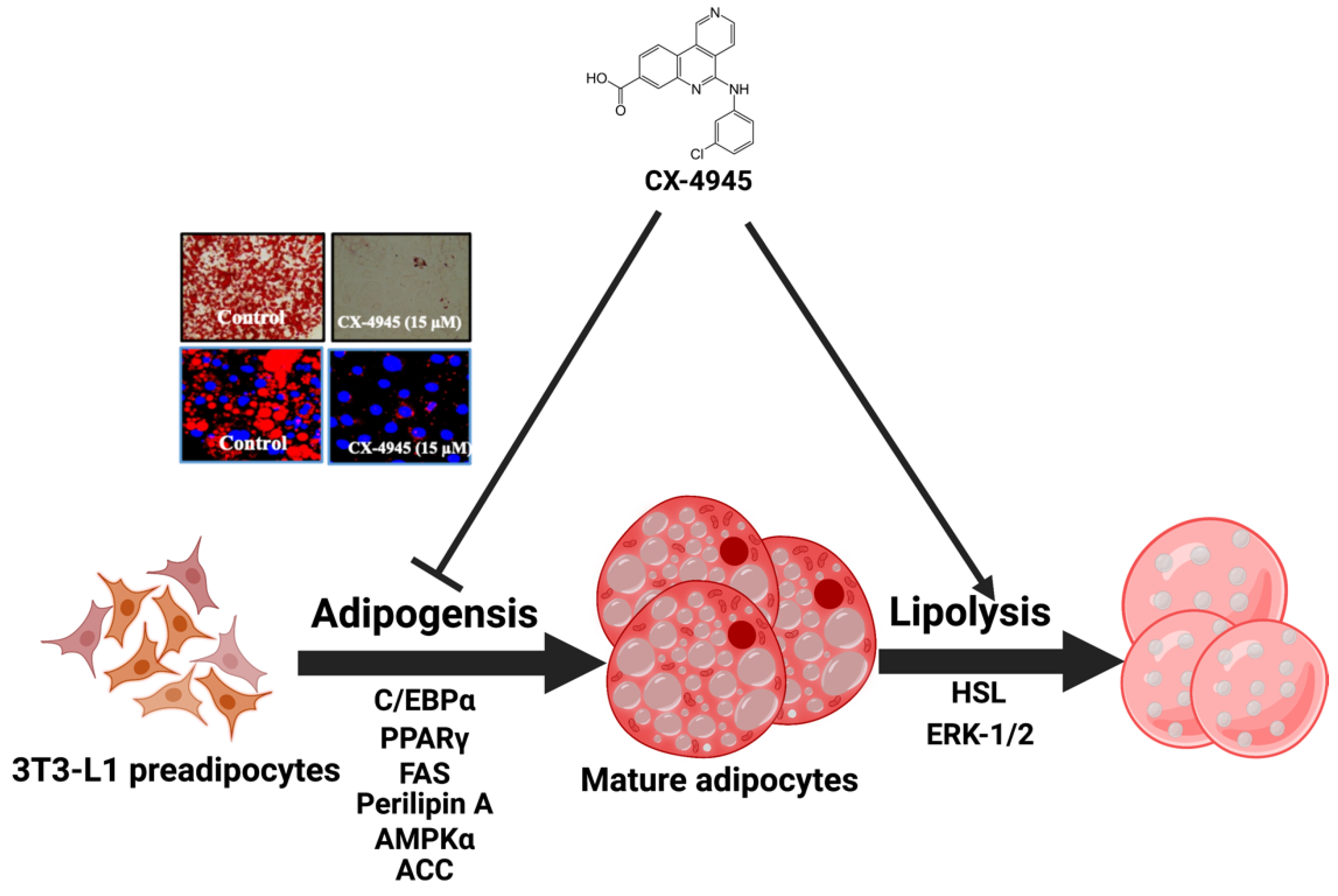
2.1. CX-4945 Markedly Reduces Lipid Accumulation and TG Content in Differentiating 3T3-L1 Preadipocytes with no Significant Cytotoxicity
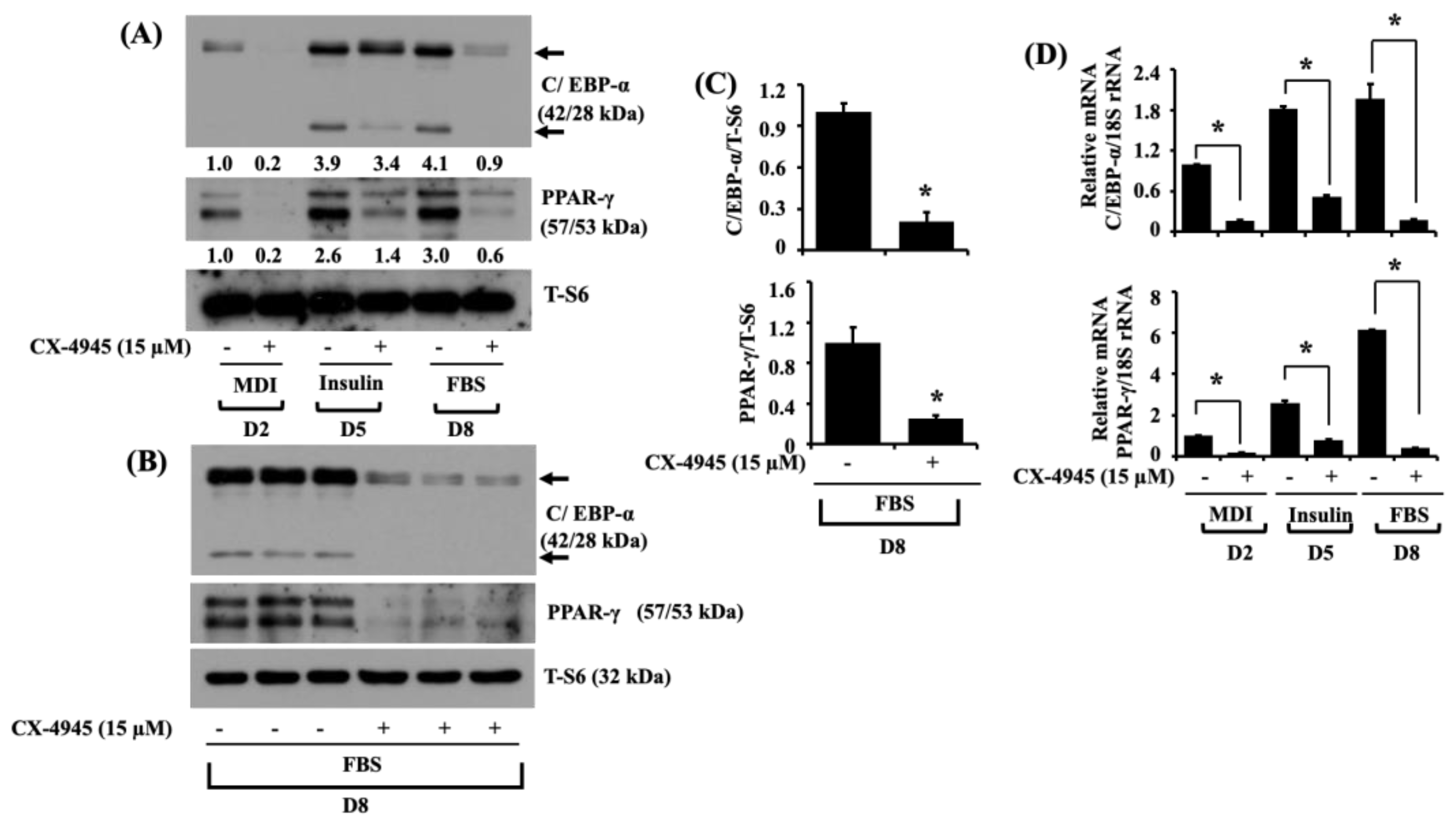
2.2. CX-4945 Decreases the Expression Levels of C/EBP-α and PPAR-γ during 3T3-L1 Preadipocyte Differentiation
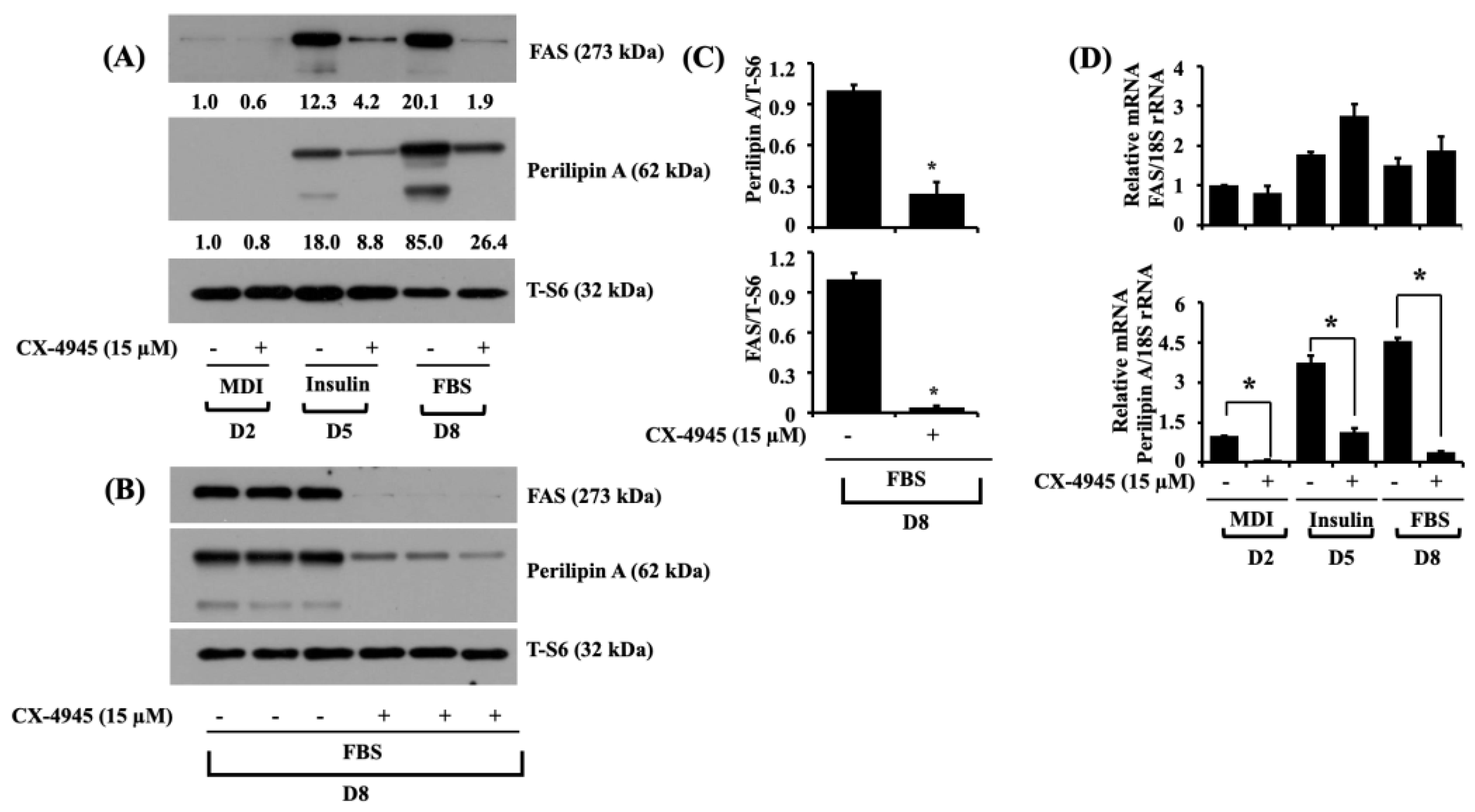
2.3. CX-4945 Reduces the Expression Levels of FAS and Perilipin A during 3T3-L1 Preadipocyte Differentiation
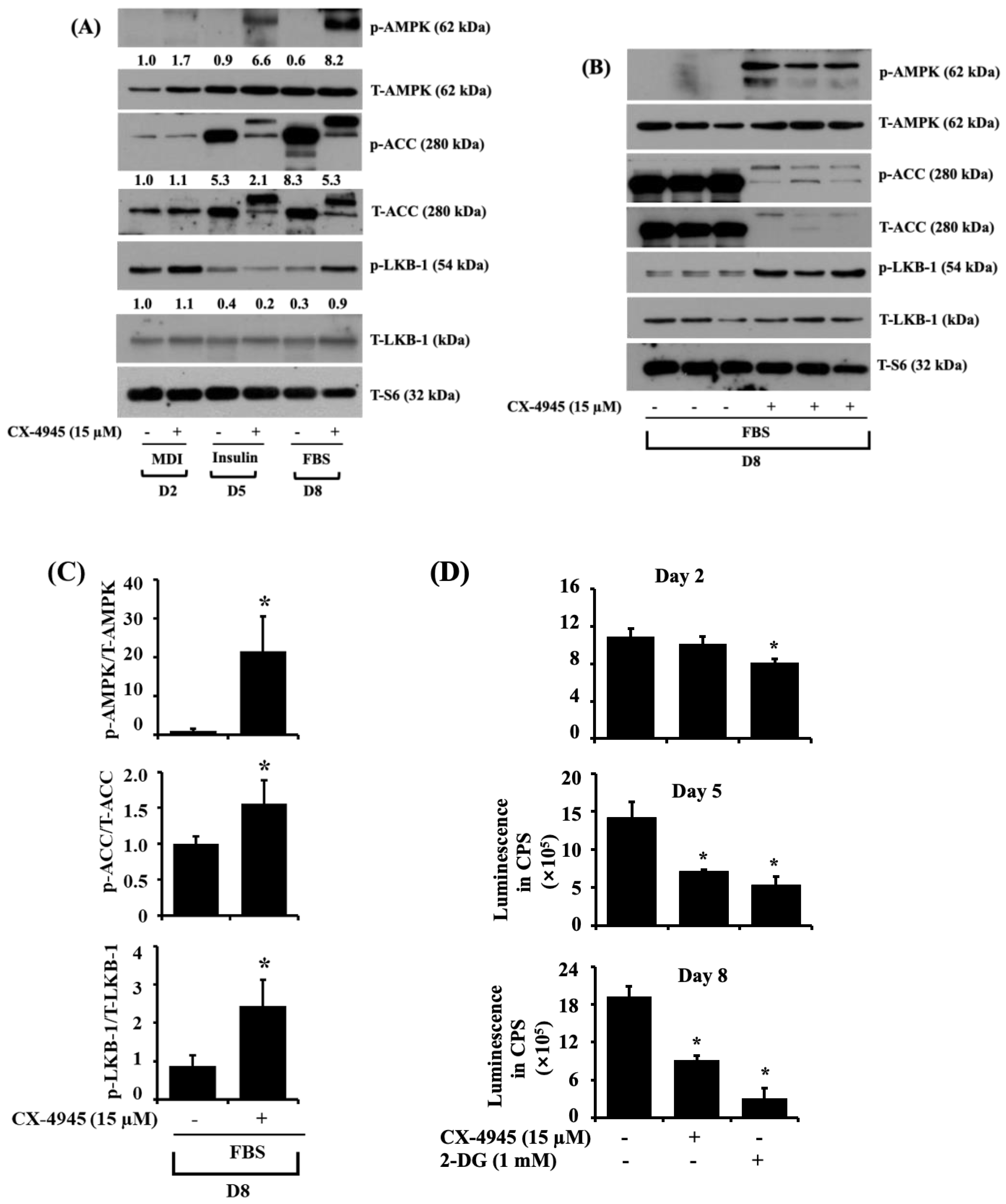
2.4. CX-4945 Increases the Phosphorylation Levels of AMPK, LKB-1, and ACC While Decreasing the Intracellular ATP Content during 3T3-L1 Preadipocyte Differentiation
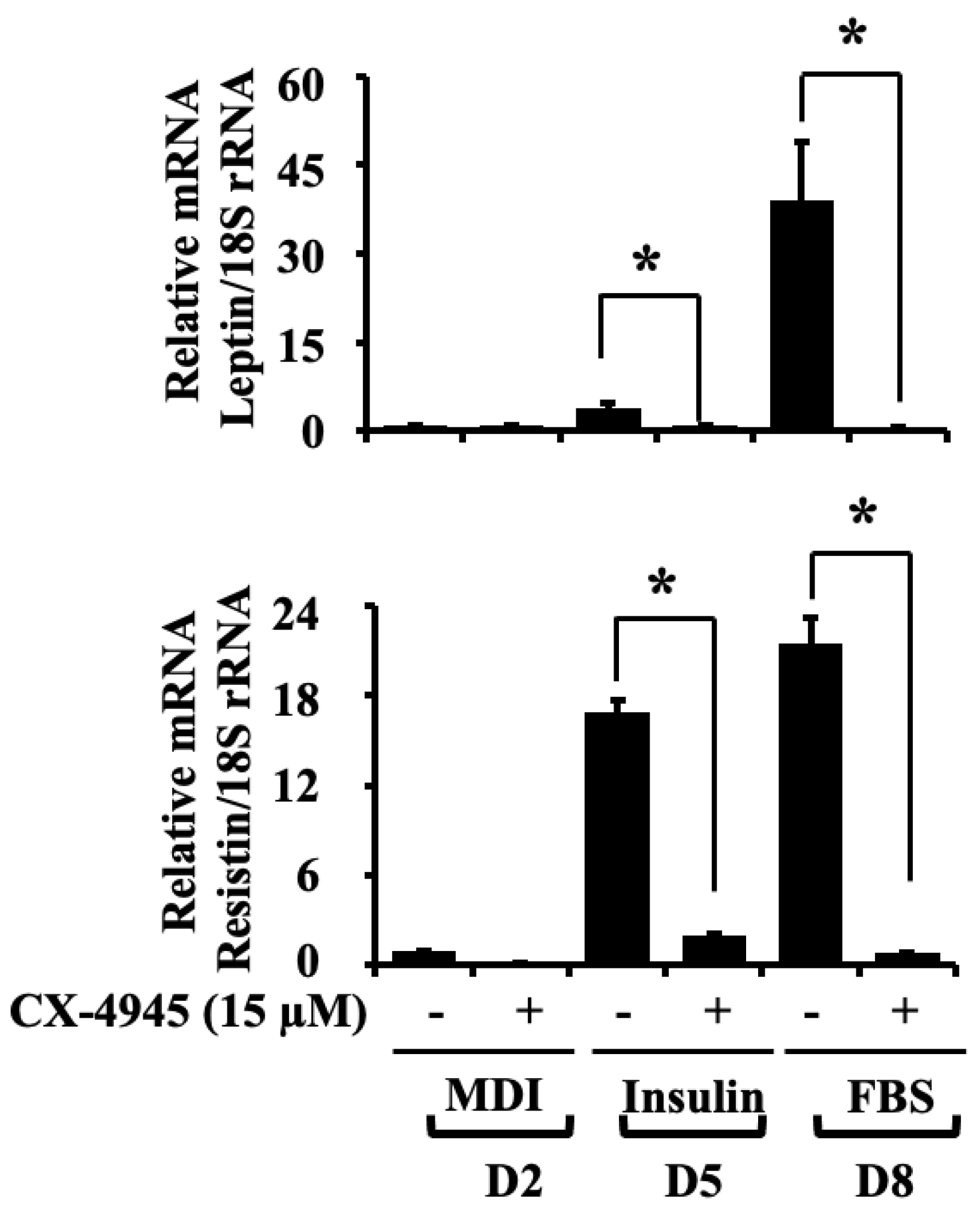
2.5. CX-4945 Down-Regulates the mRNA Expression Levels of Leptin and Resistin during 3T3-L1 Preadipocyte Differentiation
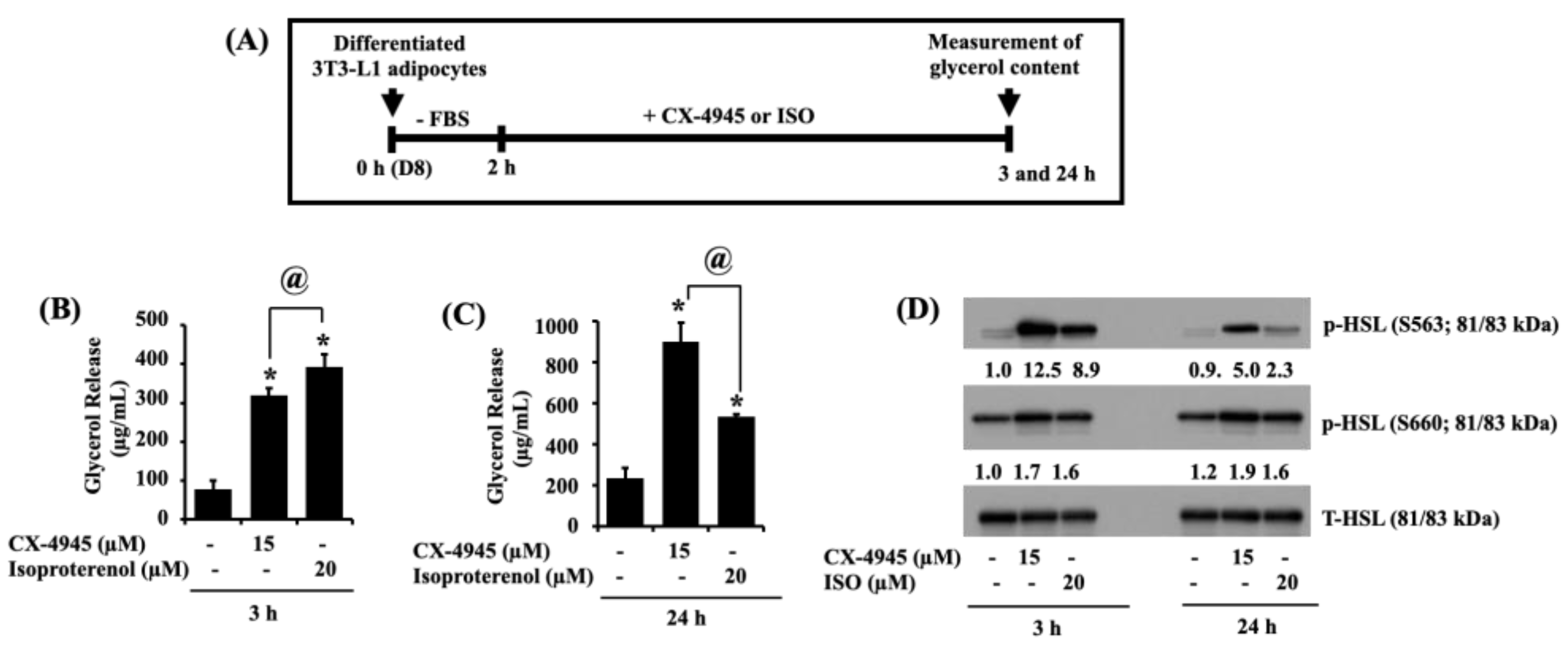
2.6. CX-4945 Can Increase Glycerol Release and the Phosphorylation Levels of HSL in Differentiated 3T3-L1 Adipocytes
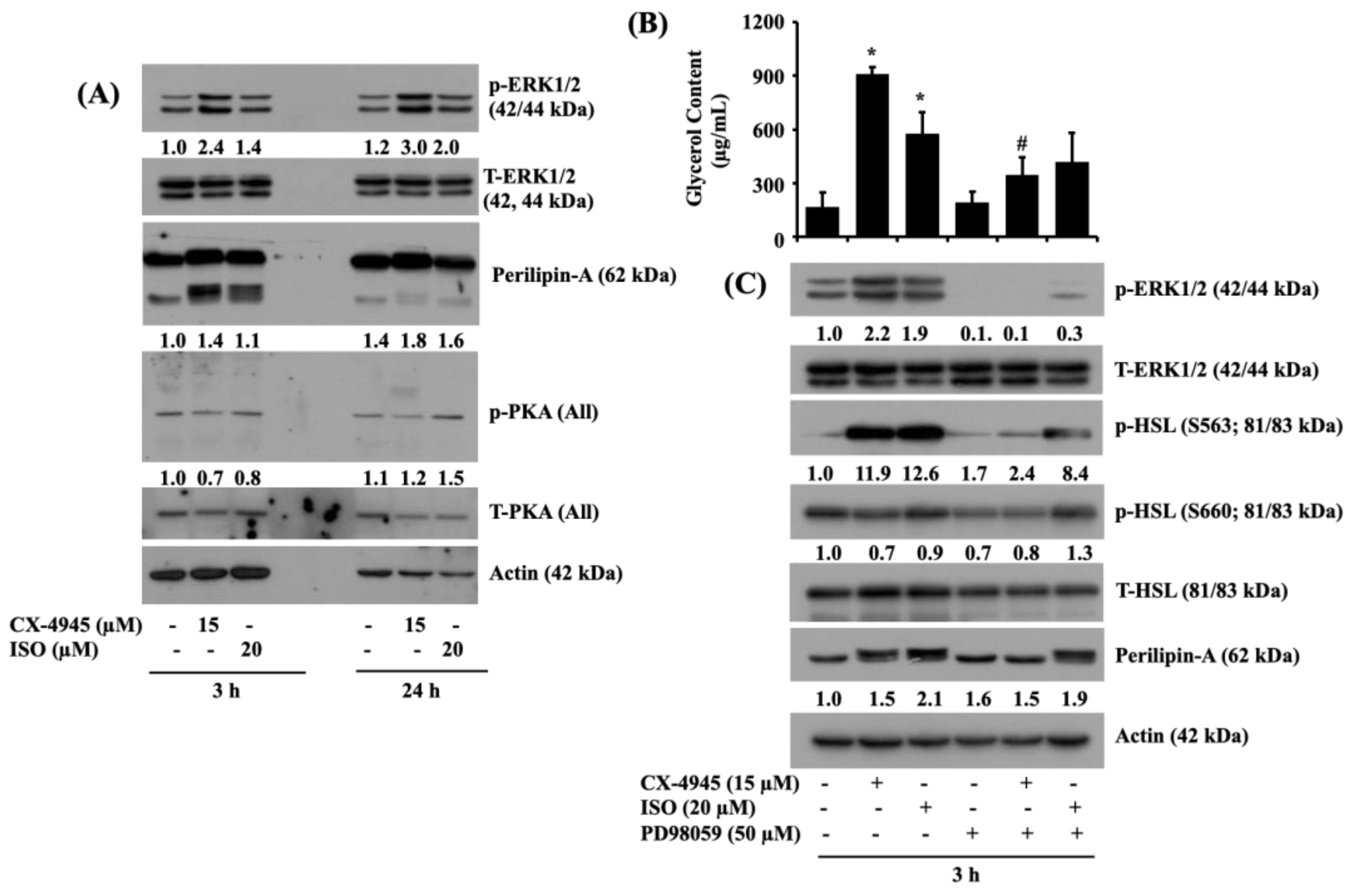
2.7. CX-4945 INDUCES the Phosphorylation of ERK-1/2 and Perilipin A, but Not PKA, and PD98059, an Inhibitor of ERK-1/2, Powerfully Block the CX-4945-Induced Glycerol Release and Phosphorylation of HSL and Perilipin A in Differentiated 3T3-L1 Adipocytes
3. Discussion
4. Materials and Methods
4.1. Drugs and Antibodies
4.2. Cell Culture
4.3. Differentiation of 3T3-L1 Preadipocytes
4.4. Oil Red O Staining
4.5. Nile-Red Staining
4.6. Cell Survival Assay
4.7. Measurement of Intracellular TG
4.8. Quantification of Glycerol Content
4.9. Preparation of Whole-Cell Lysates
4.10. Immunoblot Analysis
4.11. Quantitative Real-Time PCR
4.12. Quantification of Intracellular ATP Content
4.13. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stefan, N.; Birkenfeld, A.L.; Schulze, M.B. Global pandemics interconnected–obesity, impaired metabolic health and COVID-19. Nat. Rev. Endocrinol. 2021, 17, 135–149. [Google Scholar] [CrossRef]
- Blüher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef]
- Parra-Peralbo, E.; Talamillo, A.; Barrio, R. Origin and Development of the Adipose Tissue, a Key Organ in Physiology and Disease. Front. Cell Dev. Biol. 2021, 9, 786129. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Z.; Ngandiri, D.A.; Perez, M.L.; Wolf, A.; Wang, Y. The Molecular Brakes of Adipose Tissue Lipolysis. Front. Physiol. 2022, 13, 826314. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, M.; Blundell, J.E. Energy balance, body composition, sedentariness and appetite regulation: Pathways to obesity. Clin. Sci. 2016, 130, 1615–1628. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, D.; Stice, E.; Rodríguez-López, R.; Manco, L.; Nóbrega, C. Current review of genetics of human obesity: From molecular mechanisms to an evolutionary perspective. Mol. Genet. Genom. 2015, 290, 1191–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrillo, L.; Costa, V.; Raciti, G.A.; Longo, M.; Spinelli, R.; Esposito, R.; Nigro, C.; Vastolo, V.; Desiderio, A.; Zatterale, F.; et al. Hoxa5 undergoes dynamic DNA methylation and transcriptional repression in the adipose tissue of mice exposed to high-fat diet. Int. J. Obes. 2016, 40, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Coelho, M.; Oliveira, T.; Fernandes, R. Biochemistry of adipose tissue: An endocrine organ. Arch. Med. Sci. 2013, 9, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Blüher, M. Adipokines-removing road blocks to obesity and diabetes therapy. Mol. Metab. 2014, 3, 230–240. [Google Scholar] [CrossRef]
- Ali, A.T.; Hochfeld, W.E.; Myburgh, R.; Pepper, M.S. Adipocyte and adipogenesis. Eur. J. Cell Biol. 2013, 92, 229–236. [Google Scholar] [CrossRef]
- Jones, J.E.C.; Rabhi, N.; Orofino, J.; Gamini, R.; Perissi, V.; Vernochet, C.; Farmer, S.R. The Adipocyte Acquires a Fibroblast-like Transcriptional Signature in Response to a High Fat Diet. Sci. Rep. 2020, 10, 2380. [Google Scholar] [CrossRef] [PubMed]
- Sarjeant, K.; Stephens, J.M. Adipogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunn, E.R.; Shinde, A.B.; Zaganjor, E. Weighing in on Adipogenesis. Front Physiol. 2022, 13, 821278. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Umek, R.M.; McKnight, S.L. Regulated expression of three C/EBP isoforms during adipose conversion of 3T3-L1 cells. Genes Dev. 1991, 5, 1538–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Wang, D.; Zhao, W.; Xu, L. Deciphering the Roles of PPARγ in Adipocytes via Dynamic Change of Transcription Complex. Front. Endocrinol. 2018, 9, 473. [Google Scholar] [CrossRef] [Green Version]
- Lakshmanan, M.R.; Nepokroeff, C.M.; Porter, J.W. Control of the synthesis of fatty-acid synthetase in rat liver by insulin, glucagon, and adenosine 3’:5’ cyclic monophosphate. Proc. Natl. Acad. Sci. USA 1972, 69, 3516–3529. [Google Scholar] [CrossRef] [Green Version]
- Janovská, A.; Hatzinikolas, G.; Staikopoulos, V.; McInerney, J.; Mano, M.; Wittert, G.A. AMPK and ACC phosphorylation: Effect of leptin, muscle fibre type and obesity. Mol. Cell Endocrinol. 2008, 284, 1–10. [Google Scholar] [CrossRef]
- Kern, P.A.; Di Gregorio, G.; Lu, T.; Rassouli, N.; Ranganathan, G. Perilipin expression in human adipose tissue is elevated with obesity. J. Clin. Endocrinol. Metab. 2004, 89, 1352–1358. [Google Scholar] [CrossRef] [Green Version]
- Wolins, N.E.; Brasaemle, D.L.; Bickel, P.E. A proposed model of fat packaging by exchangeable lipid droplet proteins. FEBS Lett. 2006, 580, 5484–5491. [Google Scholar] [CrossRef] [Green Version]
- Lage, R.; Diéguez, C.; Vidal-Puig, A.; López, M. AMPK: A metabolic gauge regulating whole-body energy homeostasis. Trends Mol. Med. 2008, 14, 539–549. [Google Scholar] [CrossRef]
- Martini, C.N.; Plaza, M.V.; Vila Mdel, C. PKA-dependent and independent cAMP signaling in 3T3-L1 fibroblasts differentiation. Mol. Cell Endocrinol. 2009, 298, 42–47. [Google Scholar] [CrossRef]
- Prusty, D.; Park, B.H.; Davis, K.E.; Farmer, S.R. Activation of MEK/ERK signaling promotes adipogenesis by enhancing peroxisome proliferator-activated receptor gamma (PPARgamma) and C/EBPalpha gene expression during the differentiation of 3T3-L1 preadipocytes. J. Biol. Chem. 2002, 277, 46226–46232. [Google Scholar] [CrossRef] [Green Version]
- Engelman, J.A.; Lisanti, M.P.; Scherer, P.E. Specific inhibitors of p38 mitogen-activated protein kinase block 3T3-L1 adipogenesis. J. Biol. Chem. 1998, 273, 32111–32120. [Google Scholar] [CrossRef] [Green Version]
- Lass, A.; Zimmermann, R.; Oberer, M.; Zechner, R. Lipolysis-a highly regulated multi-enzyme complex mediates the catabolism of cellular fat stores. Prog. Lipid Res. 2011, 50, 14–27. [Google Scholar] [CrossRef] [Green Version]
- Yang, A.; Mottillo, E.P. Adipocyte lipolysis: From molecular mechanisms of regulation to disease and therapeutics. Biochem. J. 2020, 477, 985–1008. [Google Scholar] [CrossRef]
- Borgo, C.; D’Amore, C.; Sarno, S.; Salvi, M.; Ruzzene, M. Protein kinase CK2: A potential therapeutic target for diverse human diseases. Sig. Transduct. Target Ther. 2021, 6, 183. [Google Scholar] [CrossRef]
- Wilhelm, N.; Kostelnik, K.; Götz, C.; Montenarh, M. Protein kinase CK2 is implicated in early steps of the differentiation of pre-adipocytes into adipocytes. Mol. Cell Biochem. 2012, 365, 37–45. [Google Scholar] [CrossRef]
- Ma, Y.; Sender, S.; Sekora, A.; Kong, W.; Bauer, P.; Ameziane, N.; Krake, S.; Radefeldt, M.; Al-Ali, R.; Weiss, F.U.; et al. Inhibitory Response to CK II Inhibitor Silmitasertib and CDKs Inhibitor Dinaciclib Is Related to Genetic Differences in Pancreatic Ductal Adenocarcinoma Cell Lines. Int. J. Mol. Sci. 2022, 23, 4409. [Google Scholar] [CrossRef]
- Wang, S.; Yadav, A.K.; Han, J.Y.; Ahn, K.S.; Jang, B.C. Anti-Growth, Anti-Angiogenic, and Pro-Apoptotic Effects by CX-4945, an Inhibitor of Casein Kinase 2, on HuCCT-1 Human Cholangiocarcinoma Cells via Control of Caspase-9/3, DR-4, STAT-3/STAT-5, Mcl-1, eIF-2α, and HIF-1α. Int. J. Mol. Sci. 2022, 23, 6353. [Google Scholar] [CrossRef]
- Klink, M.; Rahman, M.A.; Song, C.; Dhanyamraju, P.K.; Ehudin, M.; Ding, Y.; Steffens, S.; Bhadauria, P.; Iyer, S.; Aliaga, C.; et al. Mechanistic Basis for In Vivo Therapeutic Efficacy of CK2 Inhibitor CX-4945 in Acute Myeloid Leukemia. Cancers 2021, 13, 1127. [Google Scholar] [CrossRef]
- Schwind, L.; Schetting, S.; Montenarh, M. Inhibition of Protein Kinase CK2 Prevents Adipogenic Differentiation of Mesenchymal Stem Cells Like C3H/10T1/2 Cells. Pharmaceuticals. 2017, 10, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, B.; Serpell, C.J.; Fong, I.L.; Wong, E.H. Molecular Mechanisms of Adipogenesis: The Anti-adipogenic Role of AMP-Activated Protein Kinase. Front. Mol. Biosci. 2020, 7, 76. [Google Scholar] [CrossRef] [PubMed]
- Galic, S.; Loh, K.; Murray-Segal, L.; Steinberg, G.R.; Andrews, Z.B.; Kemp, B.E. AMPK signaling to acetyl-CoA carboxylase is required for fasting- and cold-induced appetite but not thermogenesis. Elife 2018, 7, e32656. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.K.; Obiang-Obounou, B.W.; Lee, K.B.; Choi, J.S.; Jang, B.C. AZD1208, a pan-Pim kinase inhibitor, inhibits adipogenesis and induces lipolysis in 3T3-L1 adipocytes. J. Cell Mol. Med. 2018, 22, 2488–2497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurley, R.L.; Barré, L.K.; Wood, S.D.; Anderson, K.A.; Kemp, B.E.; Means, A.R.; Witters, L.A. Regulation of AMP-activated protein kinase by multisite phosphorylation in response to agents that elevate cellular cAMP. J. Biol. Chem. 2006, 281, 36662–36672. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; He, J.; Xu, C.; Zu, L.; Jiang, H.; Pu, S.; Guo, X.; Xu, G. Mechanisms of metformin inhibiting lipolytic response to isoproterenol in primary rat adipocytes. J. Mol. Endocrinol. 2009, 42, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Yadav, A.K.; Jang, B.-C. Inhibition of Lipid Accumulation and Cyclooxygenase-2 Expression in Differentiating 3T3-L1 Preadipocytes by Pazopanib, a Multikinase Inhibitor. Int. J. Mol. Sci. 2021, 22, 4884. [Google Scholar] [CrossRef]
- Achari, A.E.; Jain, S.K. Adiponectin, a Therapeutic Target for Obesity, Diabetes, and Endothelial Dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef] [Green Version]
- Yadav, A.; Kataria, M.A.; Saini, V.; Yadav, A. Role of leptin and adiponectin in insulin resistance. Clin. Chim. Acta Int. J. Clin. Chem. 2013, 417, 80–84. [Google Scholar] [CrossRef]
- Rajala, M.W.; Obici, S.; Scherer, P.E.; Rossetti, L. Adipose-derived resistin and gut-derived resistin-like molecule- beta selectively impair insulin action on glucose production. J. Clin. Investig. 2003, 111, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Towler, M.C.; Hardie, D.G. AMP-activated protein kinase in metabolic control and insulin signaling. Circ. Res. 2007, 100, 328–341. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, G.R.; Macaulay, S.L.; Febbraio, M.A.; Kemp, B.E. AMP-activated protein kinase—The fat controller of the energy railroad. Can. J. Physiol. Pharmacol. 2006, 84, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Li, X.; Liu, Y.; Hu, Y.; Yang, R. Arctigenin improves lipid metabolism by regulating AMP-activated protein kinase and downstream signaling pathways. J. Cell Biochem. 2019, 120, 13275–13288. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.K.; Ruderman, N.B. Malonyl-CoA and AMP-activated protein kinase: An expanding partnership. Mol. Cell Biochem. 2003, 253, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Sztalryd, C.; Xu, G.; Dorward, H.; Tansey, J.T.; Contreras, J.A.; Kimmel, A.R.; Londos, C. Perilipin A is essential for the translocation of hormone-sensitive lipase during lipolytic activation. J. Cell. Biol. 2003, 161, 1093–1103. [Google Scholar] [CrossRef] [PubMed]
- Bergan, H.E.; Kittilson, J.D.; Sheridan, M.A. PKC and ERK mediate GH-stimulated lipolysis. J. Mol. Endocrinol. 2013, 51, 213–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roshanzadeh, A.; Yadav, A.K.; Pydi, S.P.; Kimura, T.; Jang, B.C. Expression and Role of β3-Adrenergic Receptor during the Differentiation of 3T3-L1 Preadipocytes into Adipocytes. Biology 2022, 11, 772. [Google Scholar] [CrossRef]
- Yadav, A.K.; Jang, B.C. Anti-Survival and Pro-Apoptotic Effects of 6-Shogaol on SW872 Human Liposarcoma Cells via Control of the Intrinsic Caspase Pathway, STAT-3, AMPK, and ER Stress. Biomolecules 2020, 10, 1380. [Google Scholar] [CrossRef]
- Yadav, A.K.; Kumar, V.; Bailey, D.B.; Jang, B.C. AZD1208, a Pan-Pim Kinase Inhibitor, Has Anti-Growth Effect on 93T449 Human Liposarcoma Cells via Control of the Expression and Phosphorylation of Pim-3, mTOR, 4EBP-1, S6, STAT-3 and AMPK. Int. J. Mol. Sci. 2019, 20, 363. [Google Scholar] [CrossRef] [Green Version]
- Park, N.S.; Park, Y.K.; Yadav, A.K.; Shin, Y.M.; Bishop-Bailey, D.; Choi, J.S.; Park, J.W.; Jang, B.C. Anti-growth and pro-apoptotic effects of dasatinib on human oral cancer cells through multi-targeted mechanisms. J. Cell. Mol. Med. 2021, 25, 8300–8311. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yadav, A.K.; Jang, B.-C. Anti-adipogenic and Pro-lipolytic Effects on 3T3-L1 Preadipocytes by CX-4945, an Inhibitor of Casein Kinase 2. Int. J. Mol. Sci. 2022, 23, 7274. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137274
Yadav AK, Jang B-C. Anti-adipogenic and Pro-lipolytic Effects on 3T3-L1 Preadipocytes by CX-4945, an Inhibitor of Casein Kinase 2. International Journal of Molecular Sciences. 2022; 23(13):7274. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137274
Chicago/Turabian StyleYadav, Anil Kumar, and Byeong-Churl Jang. 2022. "Anti-adipogenic and Pro-lipolytic Effects on 3T3-L1 Preadipocytes by CX-4945, an Inhibitor of Casein Kinase 2" International Journal of Molecular Sciences 23, no. 13: 7274. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137274