
The Small Heat Shock Protein, HSPB1, Interacts with and Modulates the Physical Structure of Membranes

 , , , and
, , , and 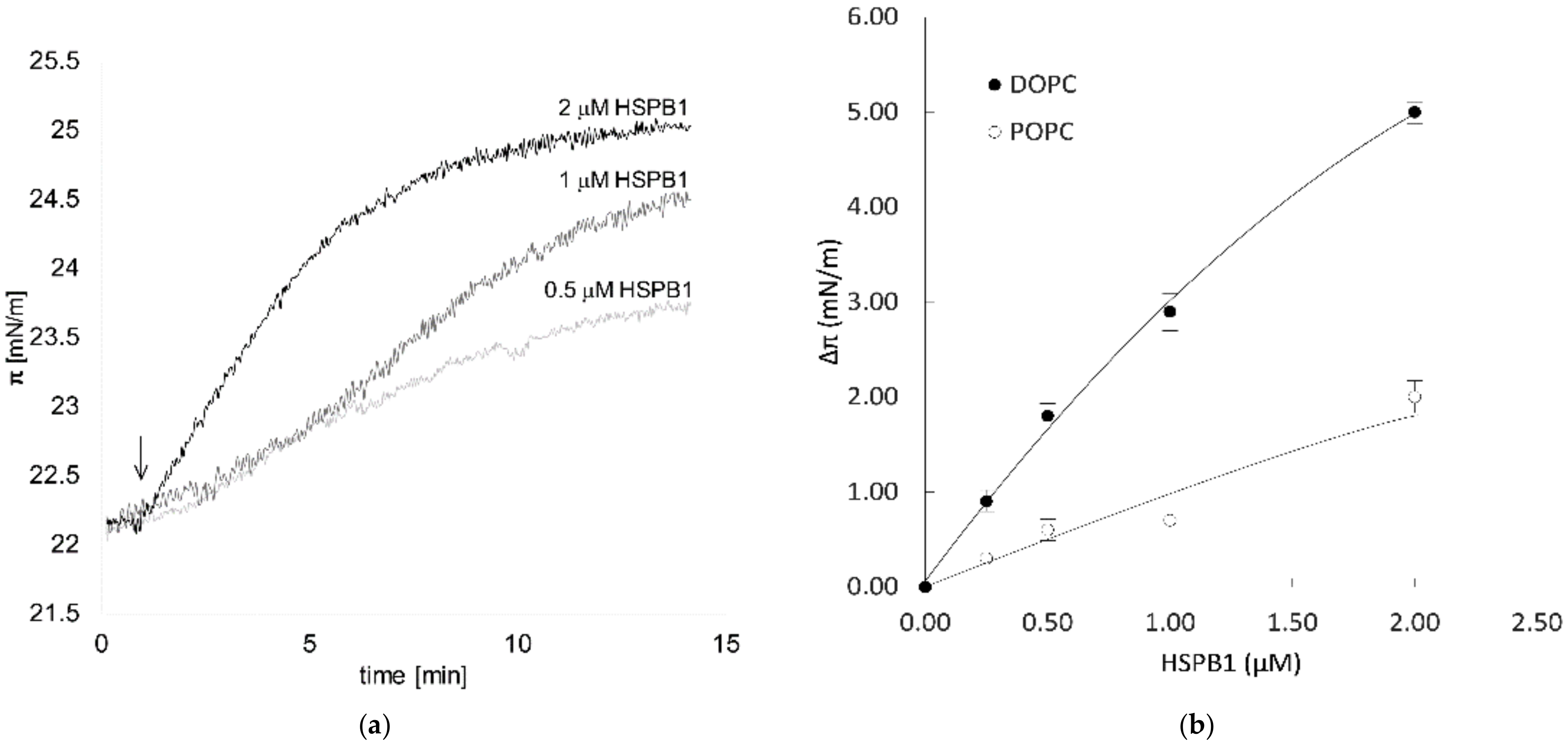{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
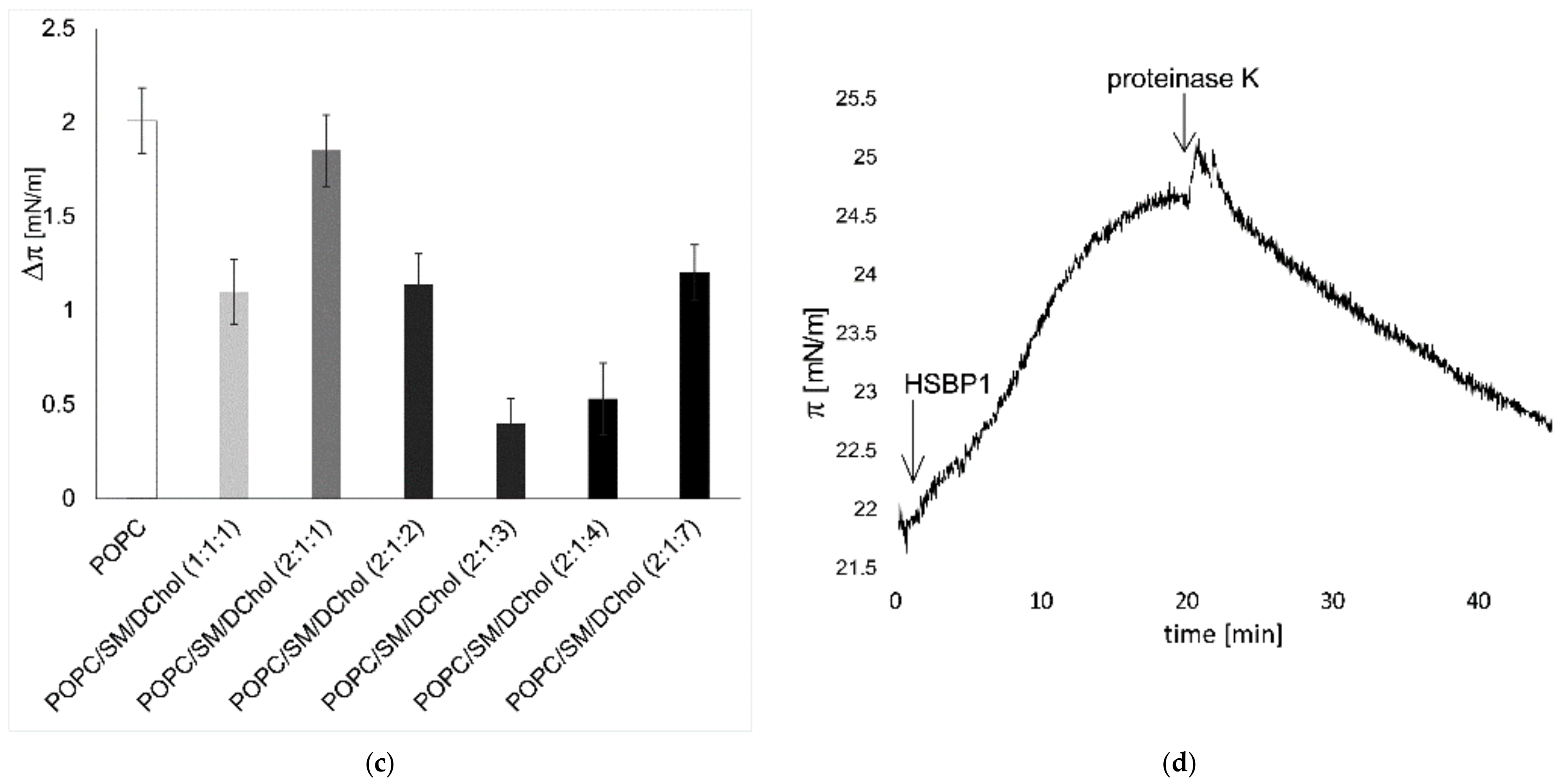
2.1. HSPB1 Interacts with Lipids: A Monolayer Study
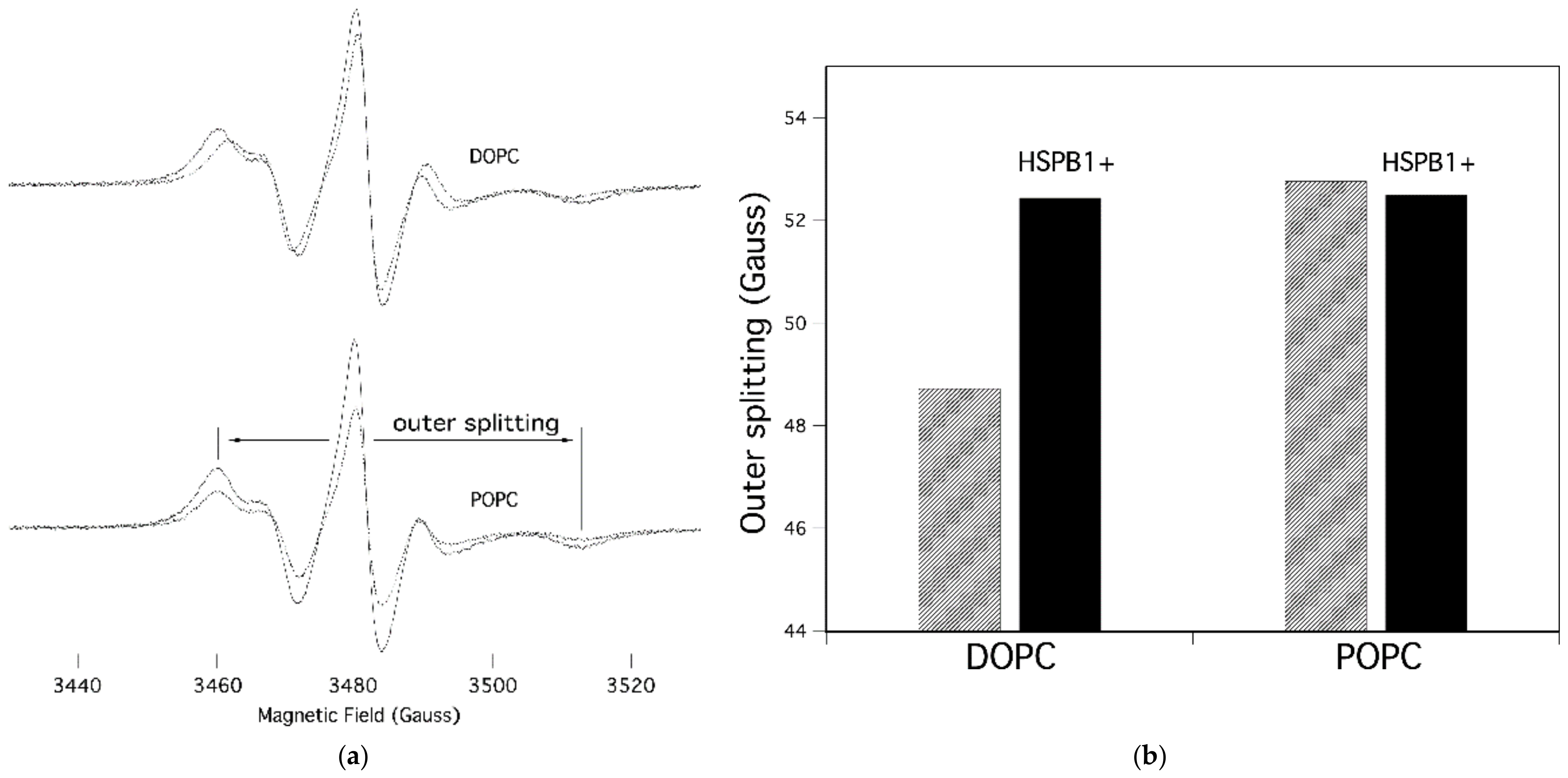
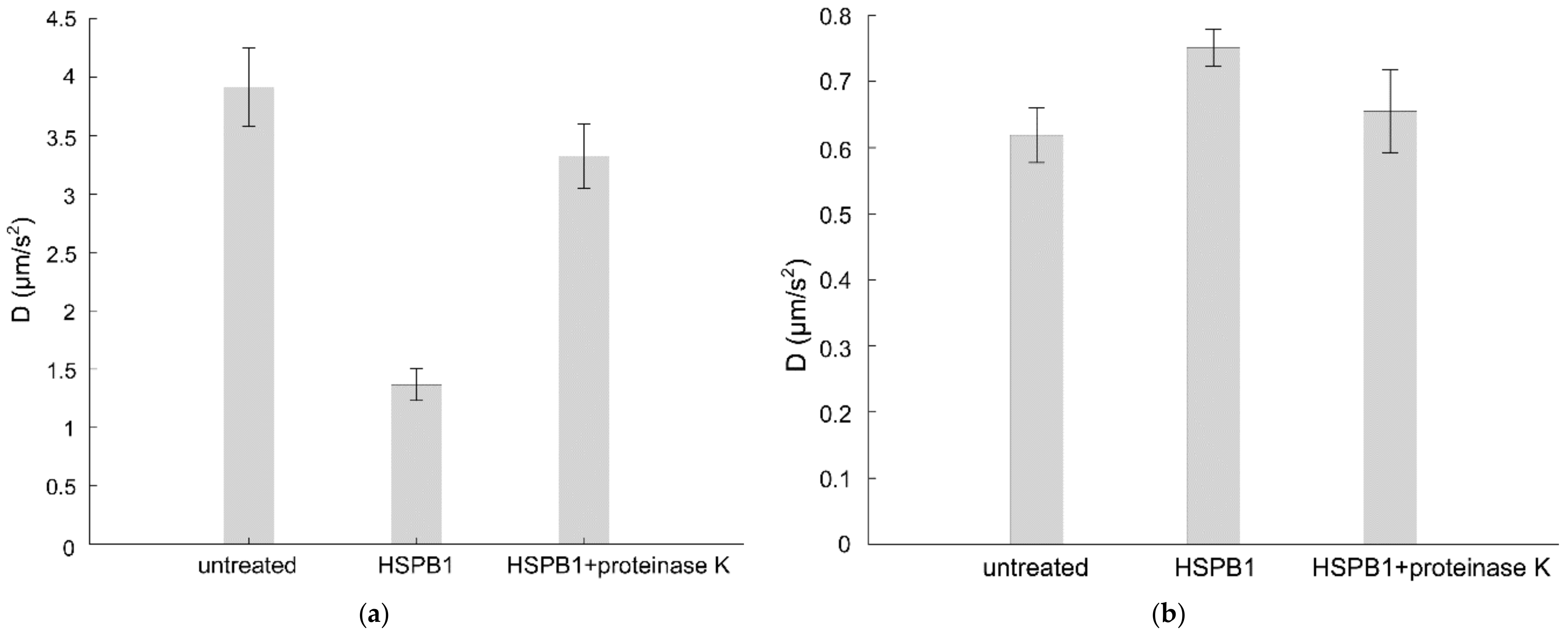
2.2. HSPB1 Interaction Decreases Both Rotational and Lateral Fluidity in Model Membranes
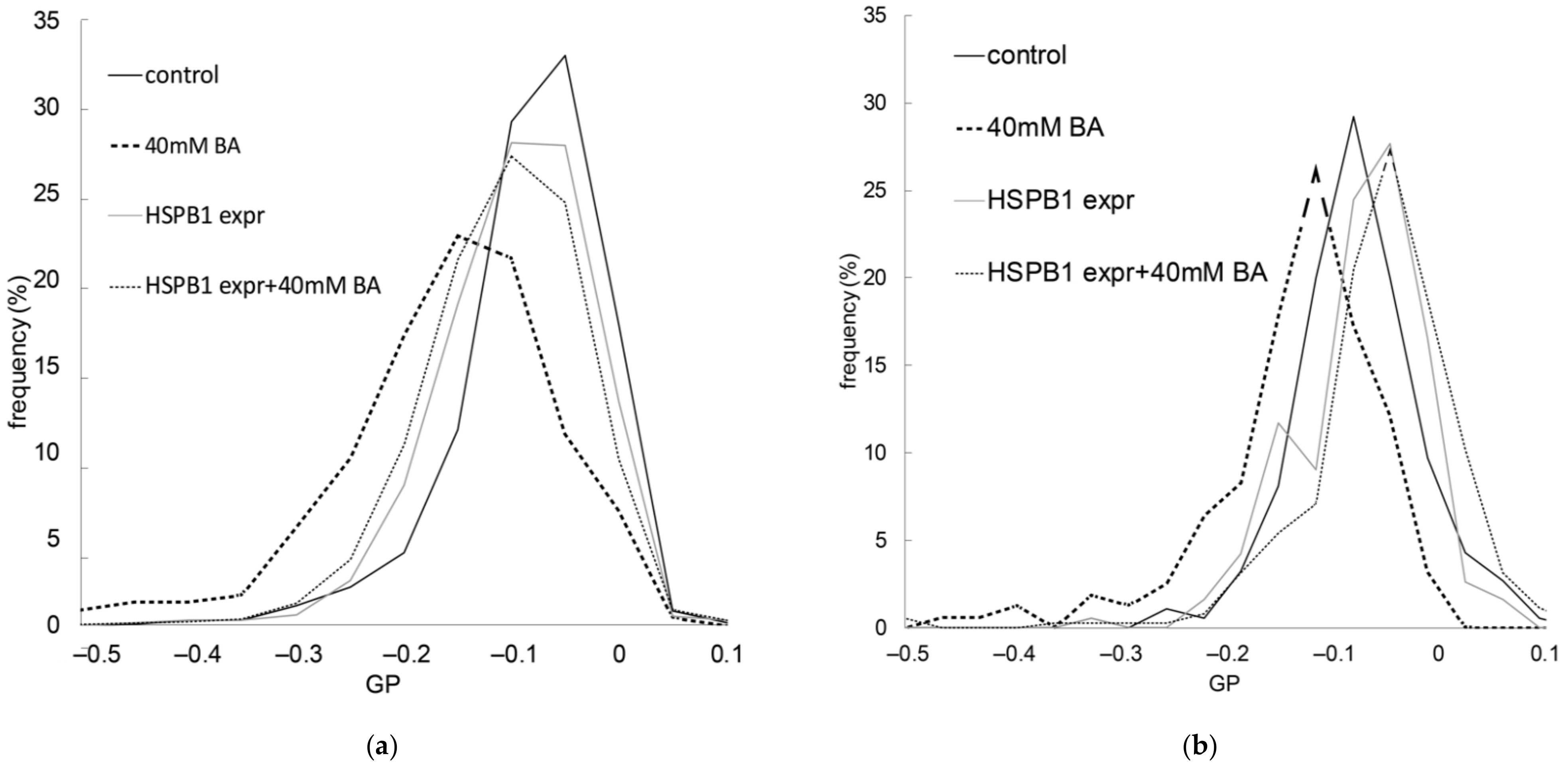
2.3. The Presence of HSPB1 Preserves Membrane Order in Mammalian Cells
3. Discussion
4. Materials and Methods
4.1. Expression and Purification of HSPB1
4.2. Langmuir Monolayer Experiments
4.3. EPR Spectroscopy
4.4. ITIR-FCS
4.5. Imaging of di-4-ANEPPDHQ and Calculation of General Polarization (GP)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Lindquist, S. The Heat-Shock Response. Annu. Rev. Biochem. 1986, 55, 1151–1191. [Google Scholar] [CrossRef]
- Kampinga, H. Chaperones in Preventing Protein Denaturation in Living Cells and Protecting Against Cellular Stress. Handb. Exp. Pharmacol. 2006, 172, 1–42. [Google Scholar] [CrossRef]
- Török, Z.; Crul, T.; Maresca, B.; Schütz, G.J.; Viana, F.; Dindia, L.; Piotto, S.; Brameshuber, M.; Balogh, G.; Péter, M.; et al. Plasma membranes as heat stress sensors: From lipid-controlled molecular switches to therapeutic applications. Biochim. Biophys. Acta 2014, 1838, 1594–1618. [Google Scholar] [CrossRef] [Green Version]
- Horváth, I.; Multhoff, G.; Sonnleitner, A.; Vígh, L. Membrane-associated stress proteins: More than simply chaperones. Biochim. Biophys. Acta 2008, 1778, 1653–1664. [Google Scholar] [CrossRef] [Green Version]
- Nakamoto, H.; Vígh, L. The small heat shock proteins and their clients. Cell. Mol. Life Sci. 2006, 64, 294–306. [Google Scholar] [CrossRef]
- Carra, S.; Alberti, S.; Arrigo, P.A.; Benesch, J.L.; Benjamin, I.J.; Boelens, W.; Bartelt-Kirbach, B.; Brundel, B.J.J.M.; Buchner, J.; Bukau, B.; et al. The growing world of small heat shock proteins: From structure to functions. Cell Stress Chaperones 2017, 22, 601–611. [Google Scholar] [CrossRef] [Green Version]
- Horváth, I.; Glatz, A.; Varvasovszki, V.; Török, Z.; Páli, T.; Balogh, G.; Kovács, E.; Nádasdi, L.; Benkö, S.; Joó, F.; et al. Membrane physical state controls the signaling mechanism of the heat shock response in Synechocystis PCC 6803: Identification of hsp17 as a “fluidity gene”. Proc. Natl. Acad. Sci. USA 1998, 95, 3513–3518. [Google Scholar] [CrossRef] [Green Version]
- Balogi, Z.; Cheregi, O.; Giese, K.C.; Juhász, K.; Vierling, E.; Vass, I.; Vígh, L.; Horváth, I. A Mutant Small Heat Shock Protein with Increased Thylakoid Association Provides an Elevated Resistance Against UV-B Damage in Synechocystis 6803. J. Biol. Chem. 2008, 283, 22983–22991. [Google Scholar] [CrossRef] [Green Version]
- Török, Z.; Goloubinoff, P.; Horváth, I.; Tsvetkova, N.M.; Glatz, A.; Balogh, G.; Varvasovszki, V.; Los, D.A.; Vierling, E.; Crowe, J.H.; et al. Synechocystis HSP17 is an amphitropic protein that stabilizes heat-stressed membranes and binds denatured proteins for subsequent chaperone-mediated refolding. Proc. Natl. Acad. Sci. USA 2001, 98, 3098–3103. [Google Scholar] [CrossRef] [Green Version]
- Tsvetkova, N.M.; Horváth, I.; Török, Z.; Wolkers, W.F.; Balogi, Z.; Shigapova, N.; Crowe, L.M.; Tablin, F.; Vierling, E.; Crowe, J.H.; et al. Small heat-shock proteins regulate membrane lipid polymorphism. Proc. Natl. Acad. Sci. USA 2002, 99, 13504–13509. [Google Scholar] [CrossRef] [Green Version]
- Miyake, T.; Araki, S.; Tsuchido, T. Synthesis and Sedimentation of a Subset of 15-kDa Heat Shock Proteins in Escherichia coli Cells Recovering from Sublethal Heat Stress. Biosci. Biotechnol. Biochem. 1993, 57, 578–583. [Google Scholar] [CrossRef]
- Lee, B.Y.; A Hefta, S.; Brennan, P.J. Characterization of the major membrane protein of virulent Mycobacterium tuberculosis. Infect. Immun. 1992, 60, 2066–2074. [Google Scholar] [CrossRef] [Green Version]
- Coucheney, F.; Gal, L.; Beney, L.; Lherminier, J.; Gervais, P.; Guzzo, J. A small HSP, Lo18, interacts with the cell membrane and modulates lipid physical state under heat shock conditions in a lactic acid bacterium. Biochim. Biophys. Acta-Biomembr. 2005, 1720, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Bellyei, S.; Szigeti, A.; Pozsgai, E.; Boronkai, A.; Gomori, E.; Hocsak, E.; Farkas, R.; Sumegi, B.; Gallyas, F. Preventing apoptotic cell death by a novel small heat shock protein. Eur. J. Cell Biol. 2007, 86, 161–171. [Google Scholar] [CrossRef]
- Nakagawa, M.; Tsujimoto, N.; Nakagawa, H.; Iwaki, T.; Fukumaki, Y.; Iwaki, A. Association of HSPB2, a Member of the Small Heat Shock Protein Family, with Mitochondria. Exp. Cell Res. 2001, 271, 161–168. [Google Scholar] [CrossRef]
- Cobb, B.A.; Petrash, J.M. Characterization of α-Crystallin-Plasma Membrane Binding. J. Biol. Chem. 2000, 275, 6664–6672. [Google Scholar] [CrossRef] [Green Version]
- Guay, J.; Lambert, H.; Gingras-Breton, G.; Lavoie, J.N.; Huot, J.; Landry, J. Regulation of actin filament dynamics by p38 map kinase-mediated phosphorylation of heat shock protein 27. J. Cell Sci. 1997, 110, 357–368. [Google Scholar] [CrossRef]
- Clarke, J.P.; Mearow, K.M. Cell Stress Promotes the Association of Phosphorylated HspB1 with F-Actin. PLoS ONE 2013, 8, e68978. [Google Scholar] [CrossRef] [Green Version]
- Lavoie, J.N.; Lambert, H.; Hickey, E.; Weber, L.A.; Landry, J. Modulation of cellular thermoresistance and actin filament stability accompanies phosphorylation-induced changes in the oligomeric structure of heat shock protein 27. Mol. Cell. Biol. 1995, 15, 505–516. [Google Scholar] [CrossRef] [Green Version]
- De Maio, A.; Cauvi, D.M.; Capone, R.; Bello, I.; Egberts, W.V.; Arispe, N.; Boelens, W. The small heat shock proteins, HSPB1 and HSPB5, interact differently with lipid membranes. Cell Stress Chaperones 2019, 24, 947–956. [Google Scholar] [CrossRef]
- Nagy, E.; Balogi, Z.; Gombos, I.; Åkerfelt, M.; Björkbom, A.; Balogh, G.; Török, Z.; Maslyanko, A.; Fiszer-Kierzkowska, A.; Lisowska, K.; et al. Hyperfluidization-coupled membrane microdomain reorganization is linked to activation of the heat shock response in a murine melanoma cell line. Proc. Natl. Acad. Sci. USA 2007, 104, 7945–7950. [Google Scholar] [CrossRef] [Green Version]
- Csoboz, B.; Balogh, G.; Kusz, E.; Gombos, I.; Peter, M.; Crul, T.; Gungor, B.; Haracska, L.; Bogdanovics, G.; Torok, Z.; et al. Membrane fluidity matters: Hyperthermia from the aspects of lipids and membranes. Int. J. Hyperth. 2013, 29, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Ionova, I.V.; Livshits, V.A.; Marsh, D. Phase Diagram of Ternary Cholesterol/Palmitoylsphingomyelin/Palmitoyloleoyl-Phosphatidylcholine Mixtures: Spin-Label EPR Study of Lipid-Raft Formation. Biophys. J. 2012, 102, 1856–1865. [Google Scholar] [CrossRef] [Green Version]
- Eckstein, J.; Berndt, N.; Holzhütter, H.-G. Computer Simulations Suggest a Key Role of Membranous Nanodomains in Biliary Lipid Secretion. PLoS Comput. Biol. 2015, 11, e1004033. [Google Scholar] [CrossRef] [Green Version]
- Török, Z.; Pilbat, A.-M.; Gombos, I.; Hocsák, E.; Sümegi, B.; Horváth, I.; Vígh, L. Evidence on Cholesterol-Controlled Lipid Raft Interaction of the Small Heat Shock Protein HSPB11. Heat Shock Proteins Cell. Traffick. Cell Stress Proteins Health Dis. 2012, 6, 75–85. [Google Scholar] [CrossRef]
- Kessel, A.; Ben-Tal, N.; May, S. Interactions of Cholesterol with Lipid Bilayers: The Preferred Configuration and Fluctuations. Biophys. J. 2001, 81, 643–658. [Google Scholar] [CrossRef] [Green Version]
- Ng, X.W.; Bag, N.; Wohland, T. Characterization of Lipid and Cell Membrane Organization by the Fluorescence Correlation Spectroscopy Diffusion Law. Chimia (Aarau) 2015, 69, 112–119. [Google Scholar] [CrossRef]
- Bag, N.; Sankaran, J.; Paul, A.; Kraut, R.S.; Wohland, T. Calibration and Limits of Camera-Based Fluorescence Correlation Spectroscopy: A Supported Lipid Bilayer Study. ChemPhysChem 2012, 13, 2784–2794. [Google Scholar] [CrossRef]
- De Maio, A.; Hightower, L. The interaction of heat shock proteins with cellular membranes: A historical perspective. Cell Stress Chaperones 2021, 26, 769–783. [Google Scholar] [CrossRef]
- Vigh, L.; Nakamoto, H.; Landry, J.; Muñoz, A.G.; Harwood, J.; Horvath, I. Membrane Regulation of the Stress Response from Prokaryotic Models to Mammalian Cells. Ann. N. Y. Acad. Sci. 2007, 1113, 40–51. [Google Scholar] [CrossRef]
- Burke, K.A.; Yates, E.A.; Legleiter, J. Biophysical Insights into How Surfaces, Including Lipid Membranes, Modulate Protein Aggregation Related to Neurodegeneration. Front. Neurol. 2013, 4, 17. [Google Scholar] [CrossRef] [Green Version]
- Krausser, J.; Knowles, T.P.J.; Šarić, A. Physical mechanisms of amyloid nucleation on fluid membranes. Proc. Natl. Acad. Sci. USA 2020, 117, 33090–33098. [Google Scholar] [CrossRef]
- Caillon, L.; Lequin, O.; Khemtémourian, L. Evaluation of membrane models and their composition for islet amyloid polypeptide-membrane aggregation. Biochim. Biophys. Acta-Biomembr. 2013, 1828, 2091–2098. [Google Scholar] [CrossRef]
- Viennet, T.; Wördehoff, M.M.; Uluca, B.; Poojari, C.; Shaykhalishahi, H.; Willbold, D.; Strodel, B.; Heise, H.; Buell, A.K.; Hoyer, W.; et al. Structural insights from lipid-bilayer nanodiscs link α-Synuclein membrane-binding modes to amyloid fibril formation. Commun. Biol. 2018, 1, 44. [Google Scholar] [CrossRef] [Green Version]
- Dukay, B.; Walter, F.R.; Vigh, J.P.; Barabási, B.; Hajdu, P.; Balassa, T.; Migh, E.; Kincses, A.; Hoyk, Z.; Szögi, T.; et al. Neuroinflammatory processes are augmented in mice overexpressing human heat-shock protein B1 following ethanol-induced brain injury. J. Neuroinflammation 2021, 18, 22. [Google Scholar] [CrossRef]
- Ciocca, D.R.; Cuello-Carrión, F.D.; Natoli, A.L.; Restall, C.; Anderson, R.L. Absence of caveolin-1 alters heat shock protein expression in spontaneous mammary tumors driven by Her-2/neu expression. Histochem. Cell Biol. 2012, 137, 187–194. [Google Scholar]
- Black, A.T.; Hayden, P.J.; Casillas, R.P.; Heck, D.E.; Gerecke, D.R.; Sinko, P.J.; Laskin, D.L.; Laskin, J.D. Regulation of Hsp27 and Hsp70 expression in human and mouse skin construct models by caveolae following exposure to the model sulfur mustard vesicant, 2-chloroethyl ethyl sulfide. Toxicol. Appl. Pharmacol. 2011, 253, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Crul, T.; Csoboz, B.; Gombos, I.; Marton, A.; Peter, M.; Balogh, G.; Vizler, C.; Szente, L.; Vigh, L. Modulation of Plasma Membrane Composition and Microdomain Organization Impairs Heat Shock Protein Expression in B16-F10 Mouse Melanoma Cells. Cells 2020, 9, 951. [Google Scholar] [CrossRef]
- Balogh, G.; Maulucci, G.; Gombos, I.; Horváth, I.; Török, Z.; Péter, M.; Fodor, E.; Páli, T.; Benkő, S.; Parasassi, T.; et al. Heat Stress Causes Spatially-Distinct Membrane Re-Modelling in K562 Leukemia Cells. PLoS ONE 2011, 6, e21182. [Google Scholar] [CrossRef] [Green Version]
- Vega, V.L.; Rodríguez-Silva, M.; Frey, T.; Gehrmann, M.; Diaz, J.C.; Steinem, C.; Multhoff, G.; Arispe, N.; De Maio, A. Hsp70 Translocates into the Plasma Membrane after Stress and Is Released into the Extracellular Environment in a Membrane-Associated Form that Activates Macrophages. J. Immunol. 2008, 180, 4299–4307. [Google Scholar] [CrossRef] [Green Version]
- Jakob, U.; Gaestel, M.; Engel, K.; Buchner, J. Small heat shock proteins are molecular chaperones. J. Biol. Chem. 1993, 268, 1517–1520. [Google Scholar] [CrossRef]
- Buchner, J.; Ehrnsperger, M.; Gaestel, M.; Walke, S. Purification and characterization of small heat shock proteins. Methods Enzymol. 1998, 290, 339–349. [Google Scholar] [CrossRef]
- Glatz, A.; Pilbat, A.-M.; Németh, G.L.; Vince-Kontár, K.; Jósvay, K.; Hunya, Á.; Udvardy, A.; Gombos, I.; Péter, M.; Balogh, G.; et al. Involvement of small heat shock proteins, trehalose, and lipids in the thermal stress management in Schizosaccharomyces pombe. Cell Stress Chaperones 2015, 21, 327–338. [Google Scholar] [CrossRef] [Green Version]
- Peksel, B.; Gombos, I.; Péter, M.; Vigh, L.; Tiszlavicz, Á; Brameshuber, M.; Balogh, G.; Schütz, G.J.; Horváth, I.; Török, Z. Mild heat induces a distinct “eustress” response in Chinese Hamster Ovary cells but does not induce heat shock protein synthesis. Sci. Rep. 2017, 7, 15643. [Google Scholar] [CrossRef]
- Stirling, D.R.; Swain-Bowden, M.J.; Lucas, A.M.; Carpenter, A.E.; Cimini, B.A.; Goodman, A. CellProfiler 4: Improvements in speed, utility and usability. BMC Bioinform. 2021, 22, 433. [Google Scholar] [CrossRef]
- Berg, S.; Kutra, D.; Kroeger, T.; Straehle, C.N.; Kausler, B.X.; Haubold, C.; Schiegg, M.; Ales, J.; Beier, T.; Rudy, M.; et al. ilastik: Interactive machine learning for (bio)image analysis. Nat. Methods 2019, 16, 1226–1232. [Google Scholar] [CrossRef]
- Csoboz, B.; Gombos, I.; Tatrai, E.; Tovari, J.; Kiss, A.L.; Horvath, I.; Vigh, L. Chemotherapy induced PRL3 expression promotes cancer growth via plasma membrane remodeling and specific alterations of caveolae-associated signaling. Cell Commun. Signal. 2018, 16, 51. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Csoboz, B.; Gombos, I.; Kóta, Z.; Dukic, B.; Klement, É.; Varga-Zsíros, V.; Lipinszki, Z.; Páli, T.; Vígh, L.; Török, Z. The Small Heat Shock Protein, HSPB1, Interacts with and Modulates the Physical Structure of Membranes. Int. J. Mol. Sci. 2022, 23, 7317. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137317
Csoboz B, Gombos I, Kóta Z, Dukic B, Klement É, Varga-Zsíros V, Lipinszki Z, Páli T, Vígh L, Török Z. The Small Heat Shock Protein, HSPB1, Interacts with and Modulates the Physical Structure of Membranes. International Journal of Molecular Sciences. 2022; 23(13):7317. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137317
Chicago/Turabian StyleCsoboz, Balint, Imre Gombos, Zoltán Kóta, Barbara Dukic, Éva Klement, Vanda Varga-Zsíros, Zoltán Lipinszki, Tibor Páli, László Vígh, and Zsolt Török. 2022. "The Small Heat Shock Protein, HSPB1, Interacts with and Modulates the Physical Structure of Membranes" International Journal of Molecular Sciences 23, no. 13: 7317. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137317