
Coexpression of TRPML1 and TRPML2 Mucolipin Channels Affects the Survival of Glioblastoma Patients

 , , , ,
, , , ,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results
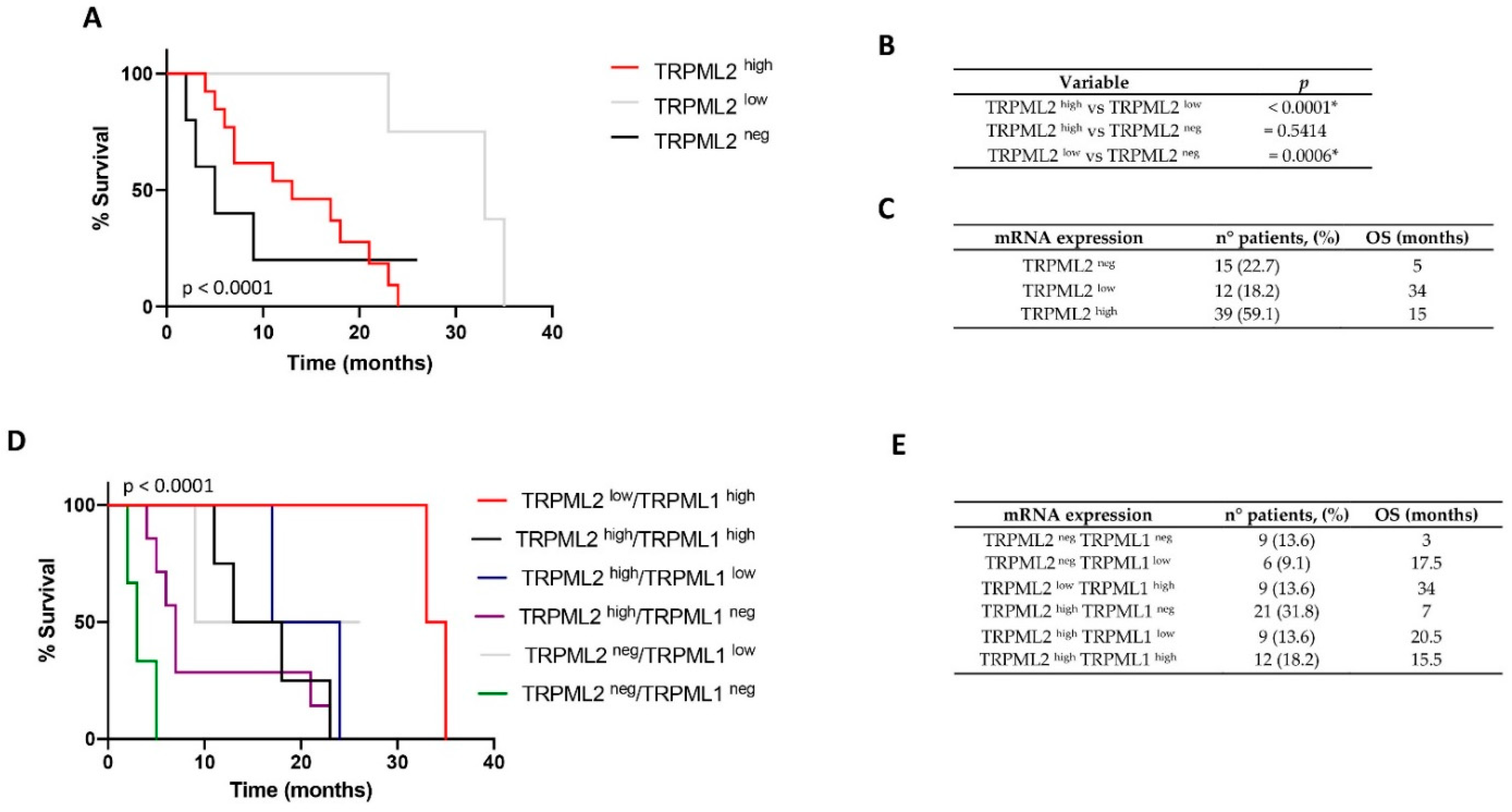
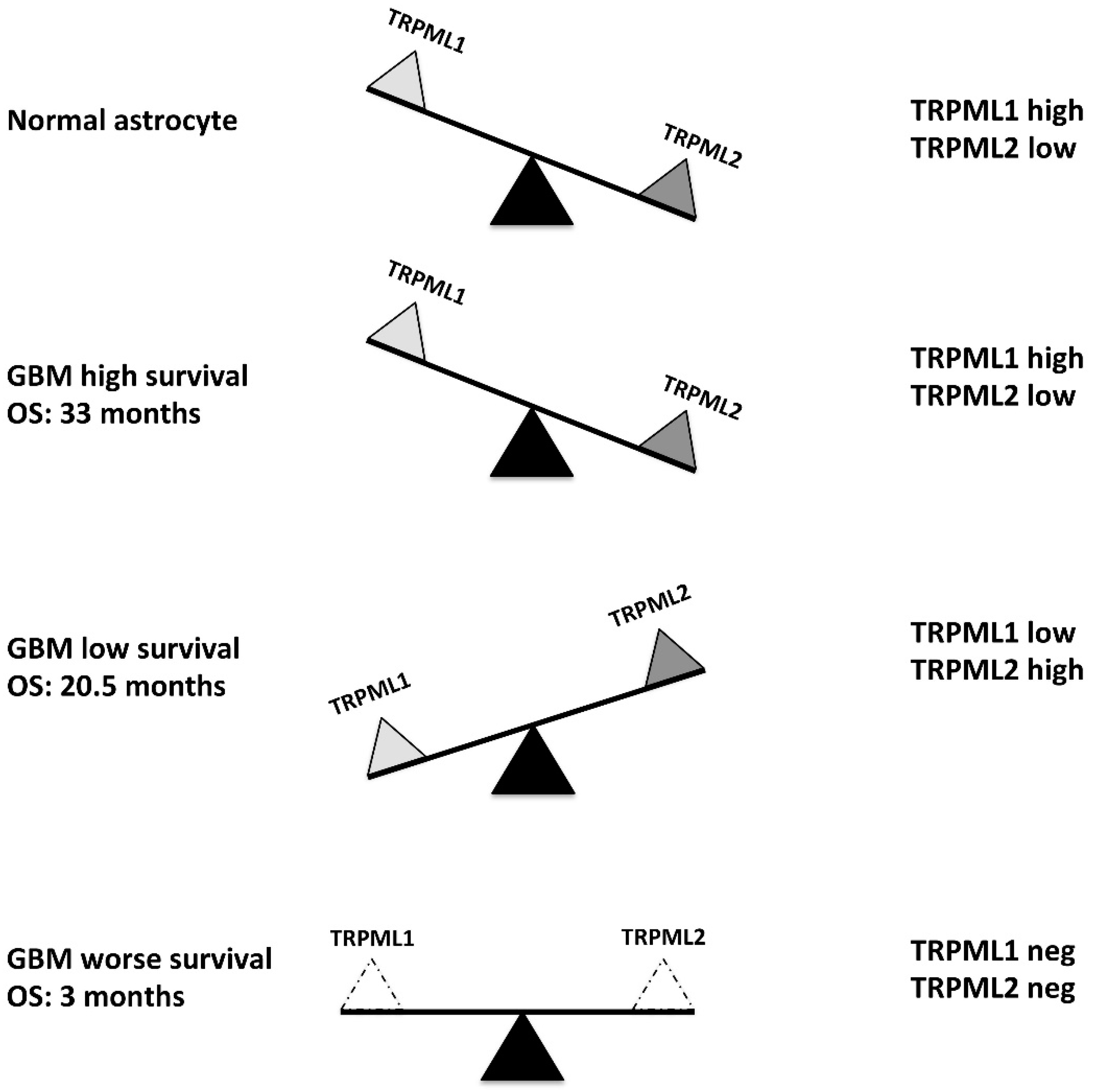
2.1. Low TRPML2 mRNA Expression Enhances the Overall Survival (OS) of GBM Patients
2.2. Correlation of TRPML1 and TRPML2 mRNA Expression with OS of GBM Patients
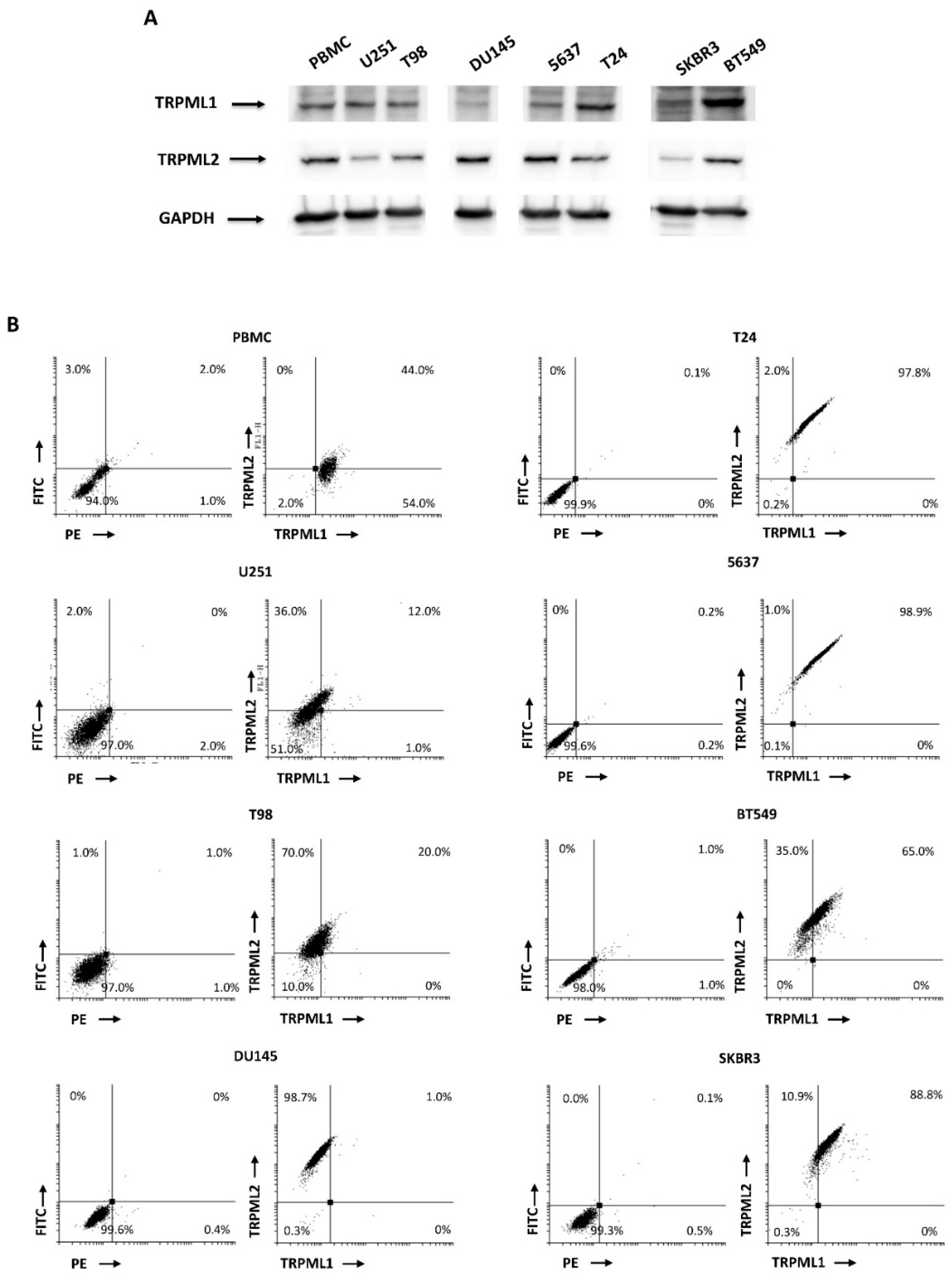
2.3. Coexpression of TRPML1 and TRPML2 Proteins inT98 and U251 Cells
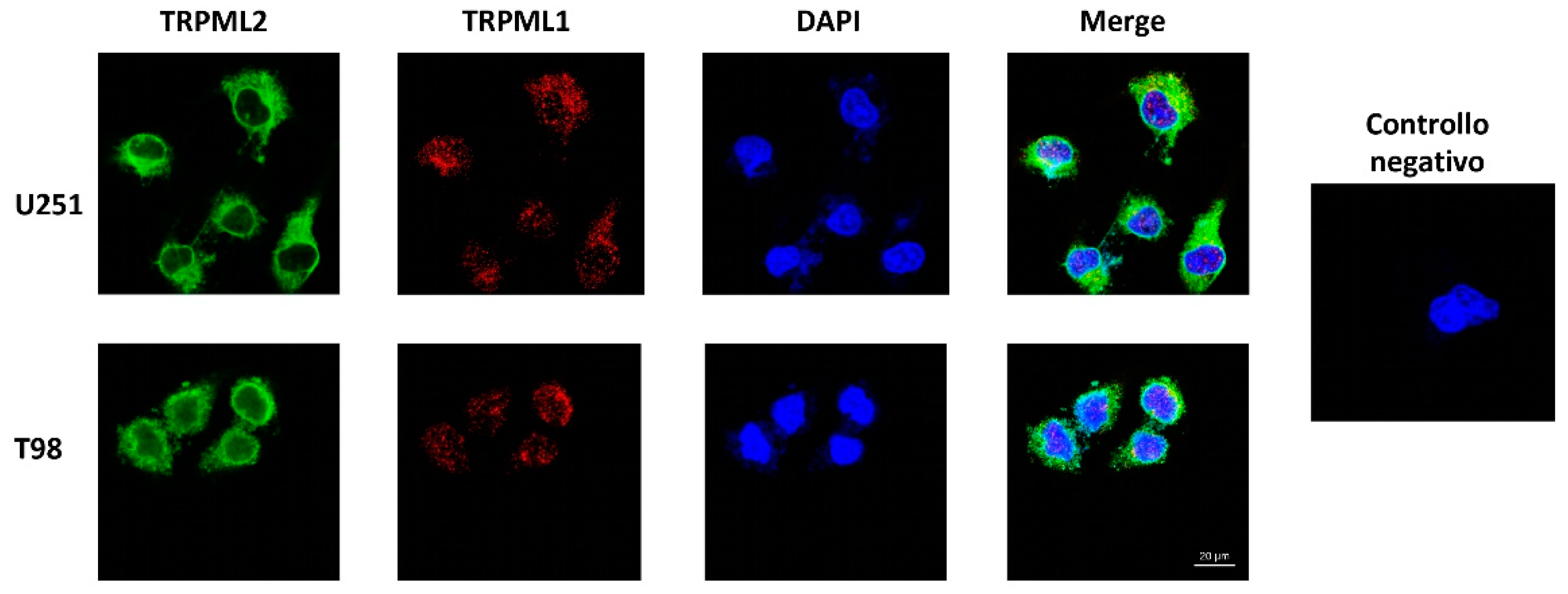
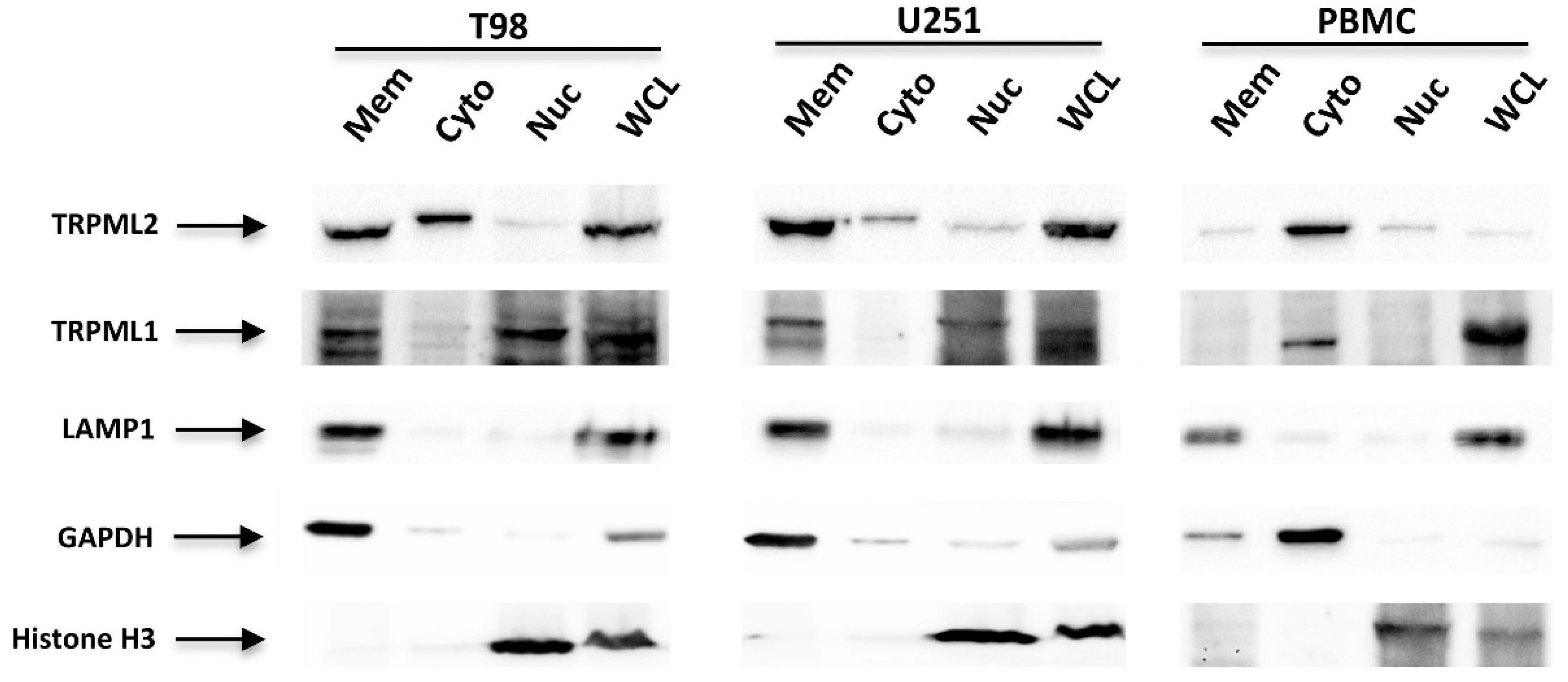
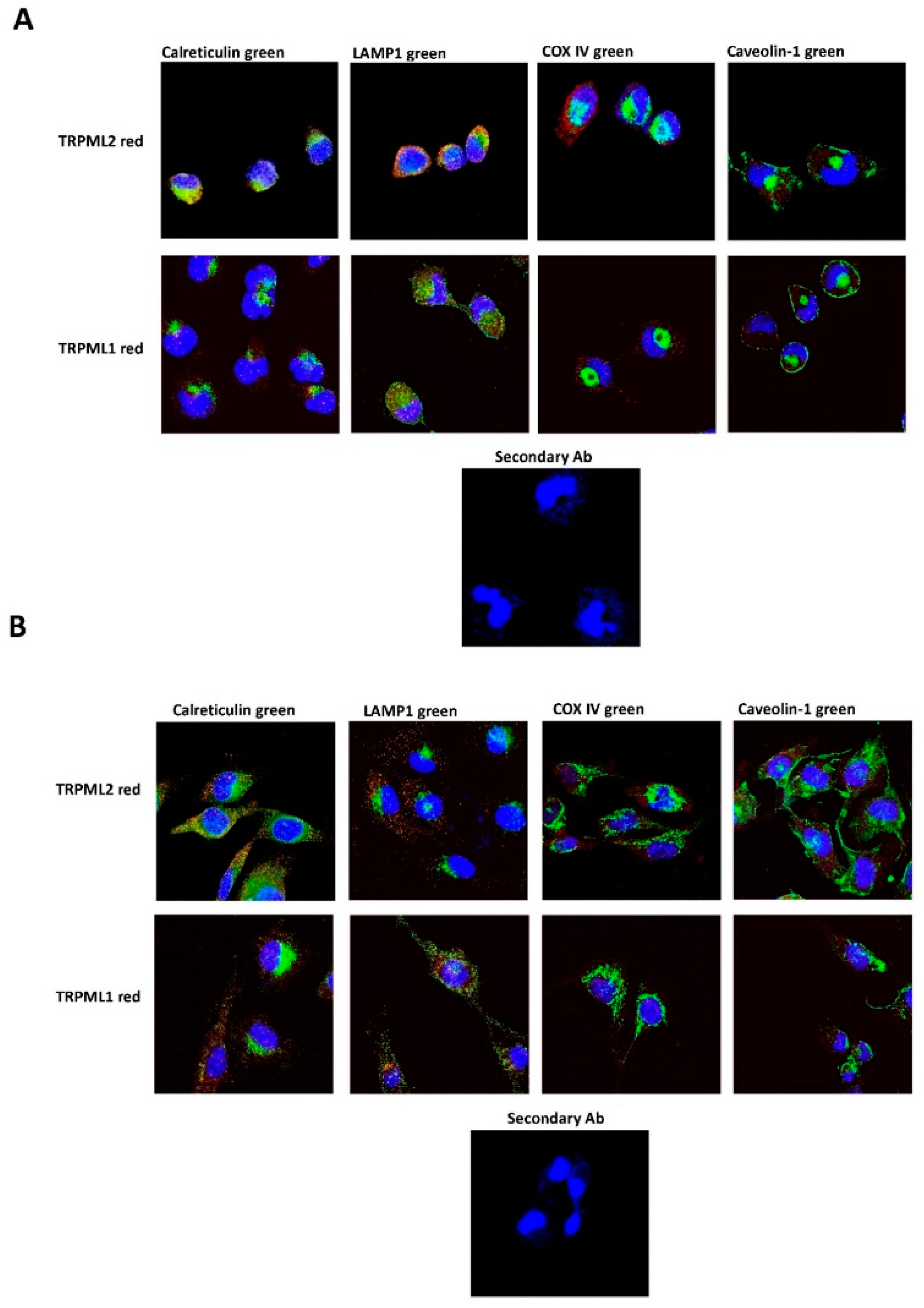
2.4. Subcellular Localization of the TRPML1 and TRPML2 Channels in GBM Cell Lines
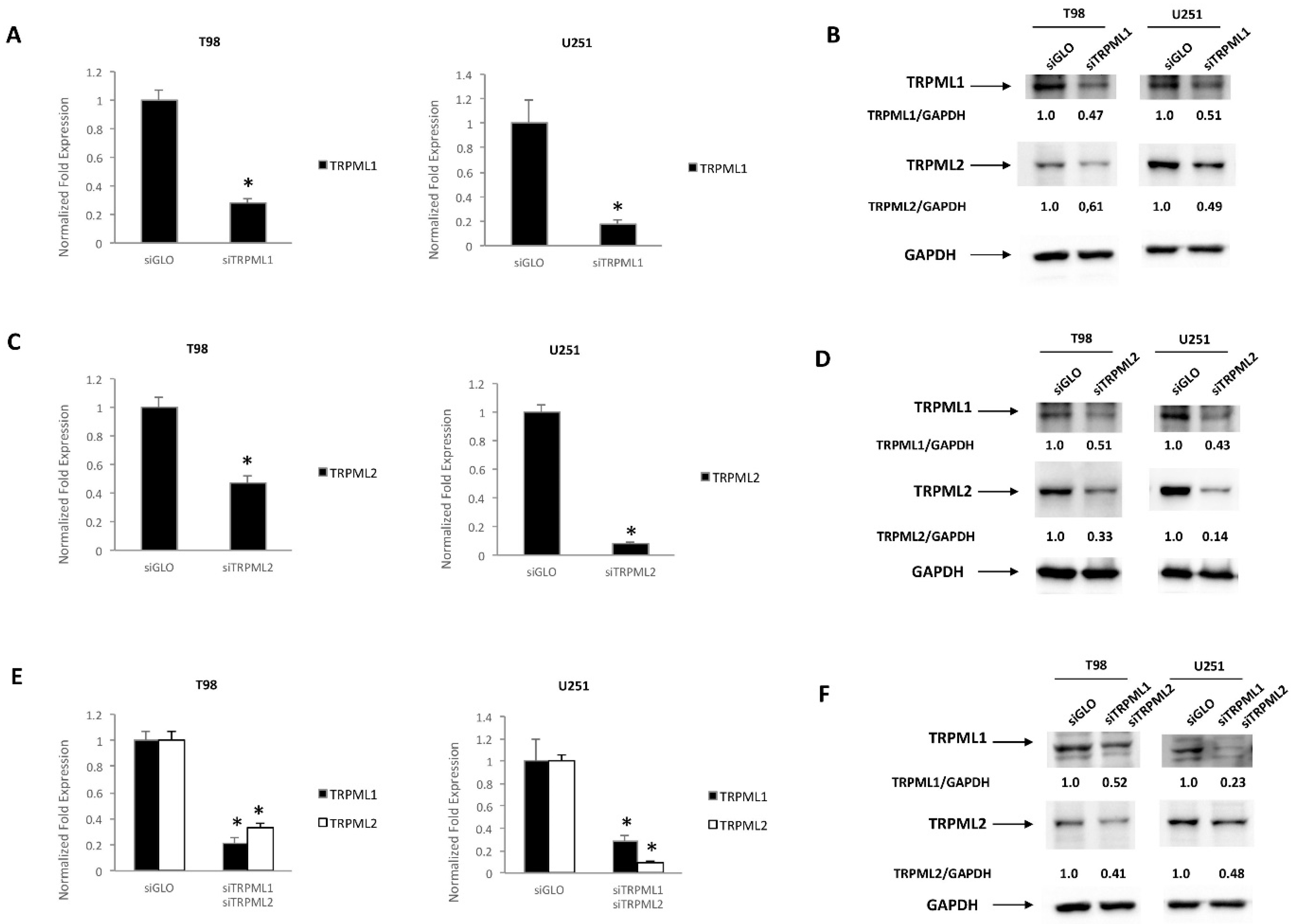
2.5. TRPML1 and TRPML2 Coregulation in T98 and U251 Cells
2.6. Gene Expression Profile in TRPML1/TRPML2 Double-Silenced T98 and U251 Cells
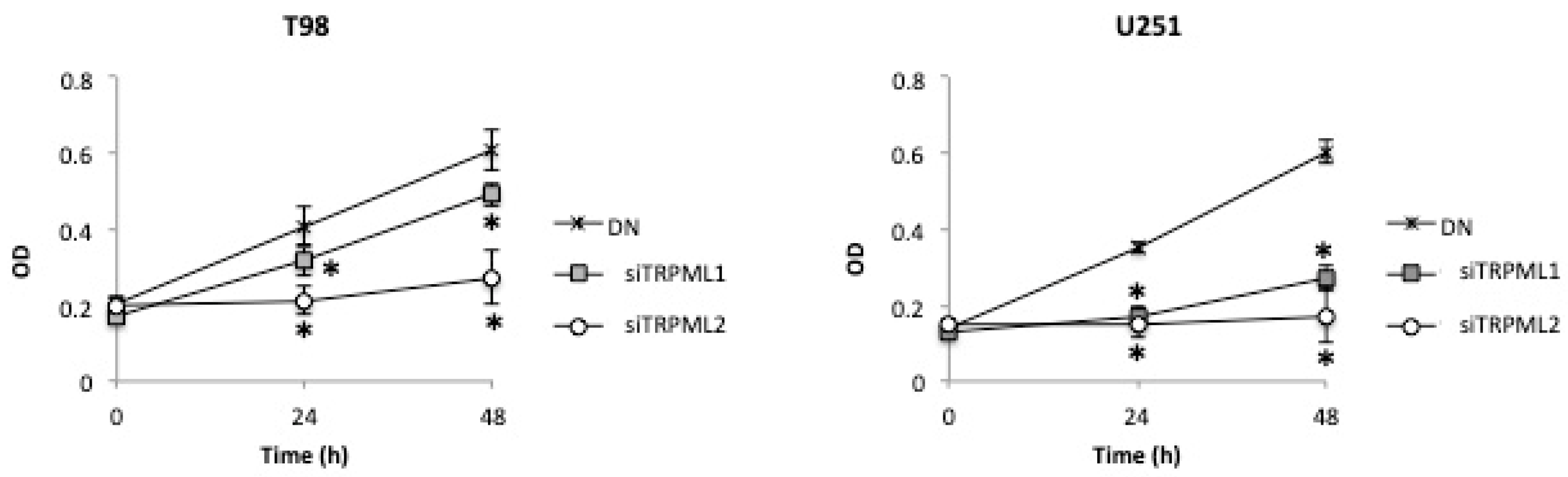
2.7. High Cell Growth Capability in DN Compared to siTRPML2 and siTRPML1 T98 and U251 Cells
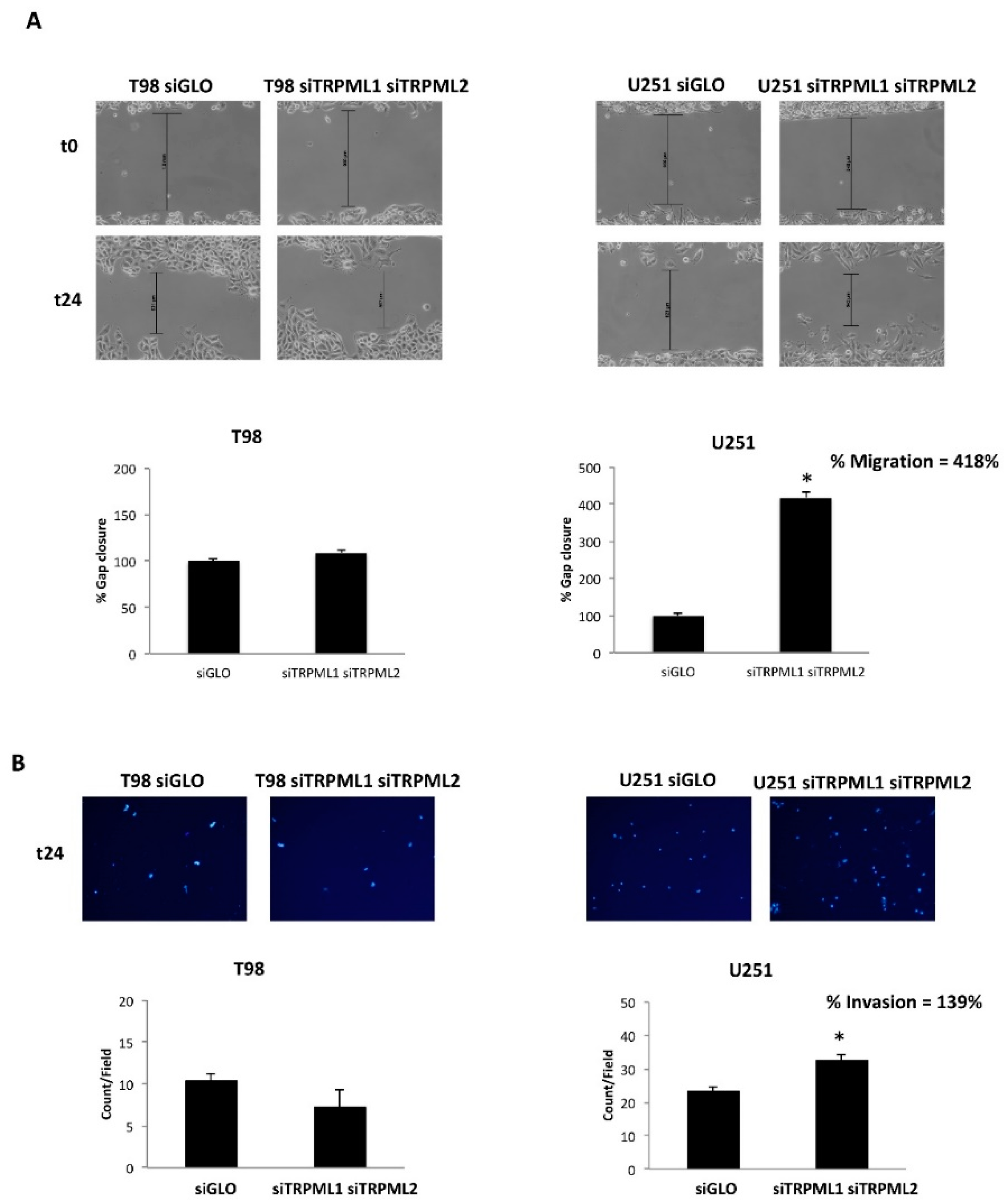
2.8. The TRPML1/TRPML2 Double Knockdown Increases the Migration/Invasion Capability in U251 GBM Cells
3. Discussion
4. Materials and Methods
4.1. Cells and Tissues
4.2. Chemical and Reagents
4.3. Western Blot Analysis
4.4. TRPML1 and/or TRPML2 Transfection Models
4.5. MTT Assay
4.6. Cell Layer Wound Assays
4.7. Invasion Assay
4.8. Confocal Laser Scanning Microscopy Analysis
4.9. Immunofluorescence and FACS Analysis
4.10. Gene Expression Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Taylor, O.G.; Brzozowski, J.S.; Skelding, K.A. Glioblastoma Multiforme: An Overview of Emerging Therapeutic Targets. Front. Oncol. 2019, 9, 963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tirosh, I.; Suvà, M.L. Tackling the Many Facets of Glioblastoma Heterogeneity. Cell Stem. Cell 2020, 26, 303–304. [Google Scholar] [CrossRef] [PubMed]
- Qazi, M.A.; Vora, P.; Venugopal, C.; Sidhu, S.S.; Moffat, J.; Swanton, C.; Singh, S.K. Intratumoral Heterogeneity: Pathways to Treatment Resistance and Relapse in Human Glioblastoma. Ann. Oncol. 2017, 28, 1448–1456. [Google Scholar] [CrossRef]
- Friedmann-Morvinski, D.; Verma, I.M. Dedifferentiation and Reprogramming: Origins of Cancer Stem Cells. EMBO Rep. 2014, 15, 244–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilius, B.; Voets, T. TRP Channels: A TR(I)P through a World of Multifunctional Cation Channels. Pflügers Arch.-Eur. J. Physiol. 2005, 451, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Santoni, G.; Farfariello, V. TRP Channels and Cancer: New Targets for Diagnosis and Chemotherapy. Endocr. Metab. Immune Disord. Drug Targets 2011, 11, 54–67. [Google Scholar] [CrossRef]
- Liberati, S.; Morelli, M.; Amantini, C.; Farfariello, V.; Santoni, M.; Conti, A.; Nabissi, M.; Cascinu, S.; Santoni, G. Loss of TRPV2 Homeostatic Control of Cell Proliferation Drives Tumor Progression. Cells 2014, 3, 112–128. [Google Scholar] [CrossRef] [Green Version]
- Santoni, G.; Santoni, M.; Maggi, F.; Marinelli, O.; Morelli, M.B. Emerging Role of Mucolipins TRPML Channels in Cancer. Front. Oncol. 2020, 10, 659. [Google Scholar] [CrossRef]
- Grimm, C.; Bartel, K.; Vollmar, A.M.; Biel, M. Endolysosomal Cation Channels and Cancer-A Link with Great Potential. Pharmaceuticals 2018, 11, 4. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.; Cho, K.; Naji, A.K.; Clemons, K.N.; Wong, C.O.; Villanueva, M.; Gregory, S.; Karagas, N.E.; Tan, L.; Liang, H.; et al. HRAS-driven Cancer Cells Are Vulnerable to TRPML1 Inhibition. EMBO Rep. 2019, 20, e46685. [Google Scholar] [CrossRef]
- Xu, M.; Almasi, S.; Yang, Y.; Yan, C.; Sterea, A.M.; Rizvi Syeda, A.K.; Shen, B.; Richard Derek, C.; Huang, P.; Gujar, S.; et al. The Lysosomal TRPML1 Channel Regulates Triple Negative Breast Cancer Development by Promoting MTORC1 and Purinergic Signaling Pathways. Cell Calcium 2019, 79, 80–88. [Google Scholar] [CrossRef]
- Kasitinon, S.Y.; Eskiocak, U.; Martin, M.; Bezwada, D.; Khivansara, V.; Tasdogan, A.; Zhao, Z.; Mathews, T.; Aurora, A.B.; Morrison, S.J. TRPML1 Promotes Protein Homeostasis in Melanoma Cells by Negatively Regulating MAPK and MTORC1 Signaling. Cell Rep. 2019, 28, 2293–2305.e9. [Google Scholar] [CrossRef] [Green Version]
- Yin, C.; Zhang, H.; Liu, X.; Zhang, H.; Zhang, Y.; Bai, X.; Wang, L.; Li, H.; Li, X.; Zhang, S.; et al. Downregulated MCOLN1 Attenuates The Progression Of Non-Small-Cell Lung Cancer By Inhibiting Lysosome-Autophagy. Cancer Manag. Res. 2019, 11, 8607–8617. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.-D.; Yan, J.; Cao, K.-Y.; Yin, Z.-Q.; Xin, W.-W.; Zhang, M.-F. MCOLN1 Promotes Proliferation and Predicts Poor Survival of Patients with Pancreatic Ductal Adenocarcinoma. Dis. Markers 2019, 2019, 9436047. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.; Liao, H.; Coker, S.A.; Liang, H.; Hancock, J.F.; Denicourt, C.; Venkatachalam, K. P53 Mitigates the Effects of Oncogenic HRAS in Urothelial Cells via the Repression of MCOLN1. iScience 2021, 24, 102701. [Google Scholar] [CrossRef]
- Almamun, M.; Levinson, B.T.; van Swaay, A.C.; Johnson, N.T.; McKay, S.D.; Arthur, G.L.; Davis, J.W.; Taylor, K.H. Integrated Methylome and Transcriptome Analysis Reveals Novel Regulatory Elements in Pediatric Acute Lymphoblastic Leukemia. Epigenetics 2015, 10, 882–890. [Google Scholar] [CrossRef] [Green Version]
- Xie, D.; Luo, X. Identification of Four Methylation-Driven Genes as Candidate Biomarkers for Monitoring Single-Walled Carbon Nanotube-Induced Malignant Transformation of the Lung. Toxicol. Appl. Pharm. 2021, 412, 115391. [Google Scholar] [CrossRef]
- Han, P.; Liu, G.; Lu, X.; Cao, M.; Yan, Y.; Zou, J.; Li, X.; Wang, G. CDH1 Rs9929218 Variant at 16q22.1 Contributes to Colorectal Cancer Susceptibility. Oncotarget 2016, 7, 47278–47286. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.-C.; Tu, S.-H.; Lien, H.-H.; Jeng, J.-Y.; Huang, C.-S.; Huang, C.-J.; Lai, L.-C.; Chuang, E.Y. Concurrent Gene Signatures for Han Chinese Breast Cancers. PLoS ONE 2013, 8, e76421. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Xie, M.; Meng, Z.; Lo, C.-Y.; Chan, F.L.; Jiang, L.; Meng, X.; Yao, X. Endolysosomal Ion Channel MCOLN2 (Mucolipin-2) Promotes Prostate Cancer Progression via IL-1β/NF-ΚB Pathway. Br. J. Cancer 2021, 125, 1420–1431. [Google Scholar] [CrossRef]
- Wu, M.; Li, X.; Zhang, T.; Liu, Z.; Zhao, Y. Identification of a Nine-Gene Signature and Establishment of a Prognostic Nomogram Predicting Overall Survival of Pancreatic Cancer. Front. Oncol. 2019, 9, 996. [Google Scholar] [CrossRef] [Green Version]
- Morelli, M.B.; Nabissi, M.; Amantini, C.; Tomassoni, D.; Rossi, F.; Cardinali, C.; Santoni, M.; Arcella, A.; Oliva, M.A.; Santoni, A.; et al. Overexpression of Transient Receptor Potential Mucolipin-2 Ion Channels in Gliomas: Role in Tumor Growth and Progression. Oncotarget 2016, 7, 43654–43668. [Google Scholar] [CrossRef] [Green Version]
- Santoni, G.; Amantini, C.; Nabissi, M.; Arcella, A.; Maggi, F.; Santoni, M.; Morelli, M.B. Functional In Vitro Assessment of VEGFA/NOTCH2 Signaling Pathway and PRB Proteasomal Degradation and the Clinical Relevance of Mucolipin TRPML2 Overexpression in Glioblastoma Patients. Int. J. Mol. Sci. 2022, 23, 688. [Google Scholar] [CrossRef]
- Morelli, M.B.; Amantini, C.; Tomassoni, D.; Nabissi, M.; Arcella, A.; Santoni, G. Transient Receptor Potential Mucolipin-1 Channels in Glioblastoma: Role in Patient’s Survival. Cancers 2019, 11, 525. [Google Scholar] [CrossRef] [Green Version]
- Santoni, G.; Amantini, C.; Nabissi, M.; Maggi, F.; Arcella, A.; Marinelli, O.; Eleuteri, A.M.; Santoni, M.; Morelli, M.B. Knock-Down of Mucolipin 1 Channel Promotes Tumor Progression and Invasion in Human Glioblastoma Cell Lines. Front. Oncol. 2021, 11, 578928. [Google Scholar] [CrossRef]
- Lev, S.; Zeevi, D.A.; Frumkin, A.; Offen-Glasner, V.; Bach, G.; Minke, B. Constitutive Activity of the Human TRPML2 Channel Induces Cell Degeneration. J. Biol. Chem. 2010, 285, 2771–2782. [Google Scholar] [CrossRef] [Green Version]
- Zeevi, D.A.; Lev, S.; Frumkin, A.; Minke, B.; Bach, G. Heteromultimeric TRPML Channel Assemblies Play a Crucial Role in the Regulation of Cell Viability Models and Starvation-Induced Autophagy. J. Cell Sci. 2010, 123, 3112–3124. [Google Scholar] [CrossRef] [Green Version]
- LaPlante, J.M.; Sun, M.; Falardeau, J.; Dai, D.; Brown, E.M.; Slaugenhaupt, S.A.; Vassilev, P.M. Lysosomal Exocytosis Is Impaired in Mucolipidosis Type IV. Mol. Genet. Metab. 2006, 89, 339–348. [Google Scholar] [CrossRef]
- Song, Y.; Dayalu, R.; Matthews, S.A.; Scharenberg, A.M. TRPML Cation Channels Regulate the Specialized Lysosomal Compartment of Vertebrate B-Lymphocytes. Eur. J. Cell Biol. 2006, 85, 1253–1264. [Google Scholar] [CrossRef]
- Venkatachalam, K.; Hofmann, T.; Montell, C. Lysosomal Localization of TRPML3 Depends on TRPML2 and the Mucolipidosis-Associated Protein TRPML1. J. Biol. Chem. 2006, 281, 17517–17527. [Google Scholar] [CrossRef] [Green Version]
- Zeevi, D.A.; Frumkin, A.; Bach, G. TRPML and Lysosomal Function. Biochim. Biophys. Acta 2007, 1772, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Samie, M.A.; Grimm, C.; Evans, J.A.; Curcio-Morelli, C.; Heller, S.; Slaugenhaupt, S.A.; Cuajungco, M.P. The Tissue-Specific Expression of TRPML2 (MCOLN-2) Gene Is Influenced by the Presence of TRPML1. Pflügers Arch.-Eur. J. Physiol. 2009, 459, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Grimm, C.; Butz, E.; Chen, C.-C.; Wahl-Schott, C.; Biel, M. From Mucolipidosis Type IV to Ebola: TRPML and Two-Pore Channels at the Crossroads of Endo-Lysosomal Trafficking and Disease. Cell Calcium 2017, 67, 148–155. [Google Scholar] [CrossRef]
- Santoni, G.; Morelli, M.B.; Marinelli, O.; Nabissi, M.; Santoni, M.; Amantini, C. Calcium Signaling and the Regulation of Chemosensitivity in Cancer Cells: Role of the Transient Receptor Potential Channels. In Advances in Experimental Medicine and Biology; SpringerNature: Berlin/Heidelberg, Germany, 2020; pp. 505–517. [Google Scholar]
- Thul, P.J.; Åkesson, L.; Wiking, M.; Mahdessian, D.; Geladaki, A.; Ait Blal, H.; Alm, T.; Asplund, A.; Björk, L.; Breckels, L.M.; et al. A Subcellular Map of the Human Proteome. Science 2017, 356, eaal3321. [Google Scholar] [CrossRef]
- Zeevi, D.A.; Frumkin, A.; Offen-Glasner, V.; Kogot-Levin, A.; Bach, G. A Potentially Dynamic Lysosomal Role for the Endogenous TRPML Proteins. J. Pathol. 2009, 219, 153–162. [Google Scholar] [CrossRef]
- Sardiello, M.; Palmieri, M.; di Ronza, A.; Medina, D.L.; Valenza, M.; Gennarino, V.A.; di Malta, C.; Donaudy, F.; Embrione, V.; Polishchuk, R.S.; et al. A Gene Network Regulating Lysosomal Biogenesis and Function. Science 2009, 325, 473–477. [Google Scholar] [CrossRef] [Green Version]
- Valadez, J.A.; Cuajungco, M.P. PAX5 Is the Transcriptional Activator of Mucolipin-2 (MCOLN2) Gene. Gene 2015, 555, 194–202. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Wei, K.; Zhang, H.; Tang, K.; Li, F.; Zhang, T.; Liu, J.; Xu, P.; Yu, Y.; Sun, W.; et al. Mechanisms by Which Dendritic Cells Present Tumor Microparticle Antigens to CD8(+) T Cells. Cancer Immunol. Res. 2018, 6, 1057–1068. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Yu, L.; Xu, H. Lysosome Calcium in ROS Regulation of Autophagy. Autophagy 2016, 12, 1954–1955. [Google Scholar] [CrossRef] [Green Version]
- Shih, Y.-T.; Wang, M.-C.; Peng, H.-H.; Chen, T.-F.; Chen, L.; Chang, J.-Y.; Chiu, J.-J. Modulation of Chemotactic and Pro-Inflammatory Activities of Endothelial Progenitor Cells by Hepatocellular Carcinoma. Cell Signal. 2012, 24, 779–793. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Genes | WT T98 | DN T98 | WT U251 | DN U251 | ||||
|---|---|---|---|---|---|---|---|---|
| cDNA Copies/μL | Gene/ACTB | cDNA Copies/μL | Gene/ACTB | cDNA Copies/μL | Gene/ACTB | cDNA Copies/μL | Gene/ACTB | |
| ALCAM | 826 | 0.11 | 487 * | 0.06 | 1153 | 0.10 | 653 * | 0.06 |
| CD44 | 4510 | 0.61 | 2875 * | 0.38 | 10,200 | 0.90 | 12,161 * | 1.16 |
| EPCAM | 5 | - | 7 | - | 0 | - | 36 | - |
| VIMENTIN | 6421 | 0.87 | 3885 * | 0.52 | 6740 | 0.60 | 8013 * | 0.76 |
| SHH | 0 | - | 0 | - | 5 | - | 1 | - |
| DHH | 7 | - | 1 | - | 3 | - | 1 | - |
| IHH | 0 | - | 0 | - | 4 | - | 0 | - |
| PTCH1 | 83 | - | 85 | 0.01 | 27 | - | 25 | - |
| PTCH2 | 2 | - | 1 | - | 102 | 0.01 | 55 | 0.00 |
| ZEB1 | 566 | 0.08 | 297 * | 0.04 | 110 | 0.01 | 78 | 0.01 |
| ZEB2 | 44 | - | 27 | - | 167 | 0.01 | 49 | 0.00 |
| SMO | 257 | 0.03 | 207 | 0.03 | 37 | - | 45 | - |
| VEGFA | 1506 | 0.20 | 1048 * | 0.14 | 3440 | 0.30 | 2055 * | 0.20 |
| VEGFB | 1492 | 0.20 | 1120 * | 0.15 | 537 | 0.05 | 440 | 0.04 |
| NOTCH1 | 34 | - | 26 | - | 381 | 0.03 | 173 * | 0.01 |
| NOTCH2 | 1492 | 0.20 | 484 * | 0.06 | 2020 | 0.18 | 645 * | 0.06 |
| STAT3 | 1090 | 0.15 | 778 * | 0.10 | 984 | 0.09 | 1099 * | 0.10 |
| POU5F1B | 0 | - | 0 | - | 1 | - | 20 | - |
| SPARC | 1835 | 0.25 | 574 * | 0.08 | 1964 | 0.17 | 1871 | 0.18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santoni, G.; Maggi, F.; Amantini, C.; Arcella, A.; Marinelli, O.; Nabissi, M.; Santoni, M.; Morelli, M.B. Coexpression of TRPML1 and TRPML2 Mucolipin Channels Affects the Survival of Glioblastoma Patients. Int. J. Mol. Sci. 2022, 23, 7741. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23147741
Santoni G, Maggi F, Amantini C, Arcella A, Marinelli O, Nabissi M, Santoni M, Morelli MB. Coexpression of TRPML1 and TRPML2 Mucolipin Channels Affects the Survival of Glioblastoma Patients. International Journal of Molecular Sciences. 2022; 23(14):7741. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23147741
Chicago/Turabian StyleSantoni, Giorgio, Federica Maggi, Consuelo Amantini, Antonietta Arcella, Oliviero Marinelli, Massimo Nabissi, Matteo Santoni, and Maria Beatrice Morelli. 2022. "Coexpression of TRPML1 and TRPML2 Mucolipin Channels Affects the Survival of Glioblastoma Patients" International Journal of Molecular Sciences 23, no. 14: 7741. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23147741