
Protein Abundance of Drug Transporters in Human Hepatitis C Livers

 , , , ,
, , , ,
Abstract
:1. Introduction
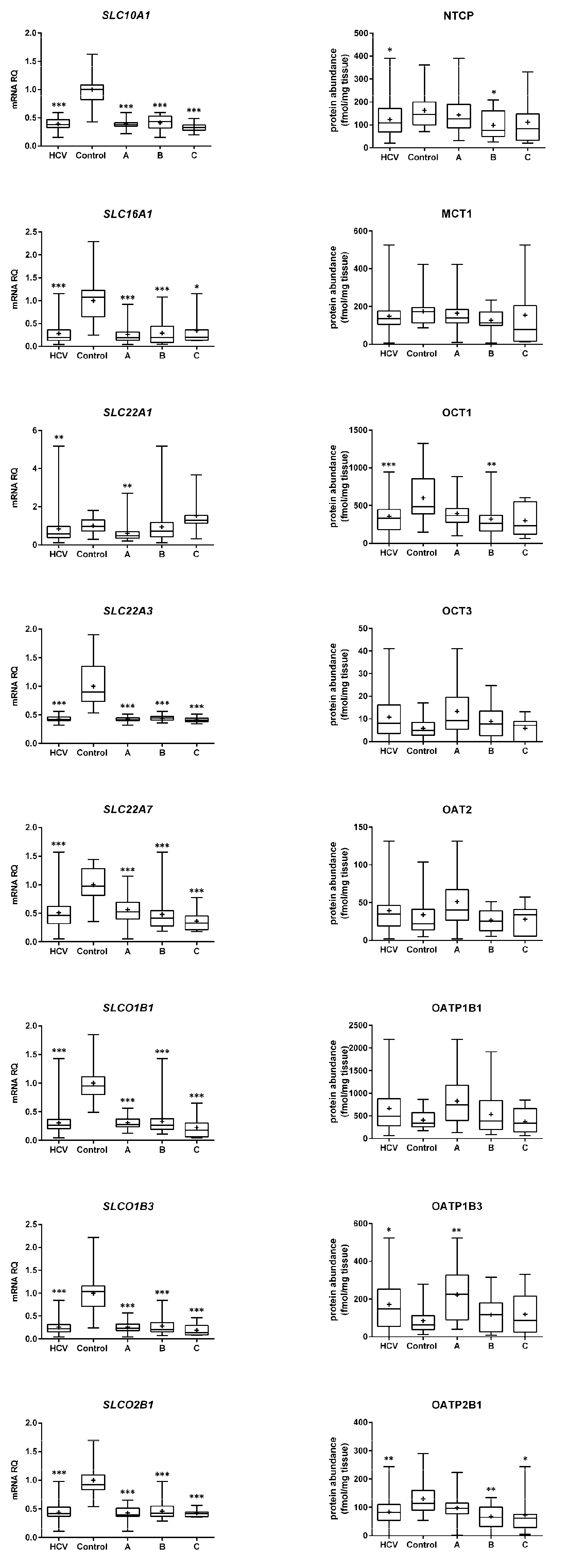
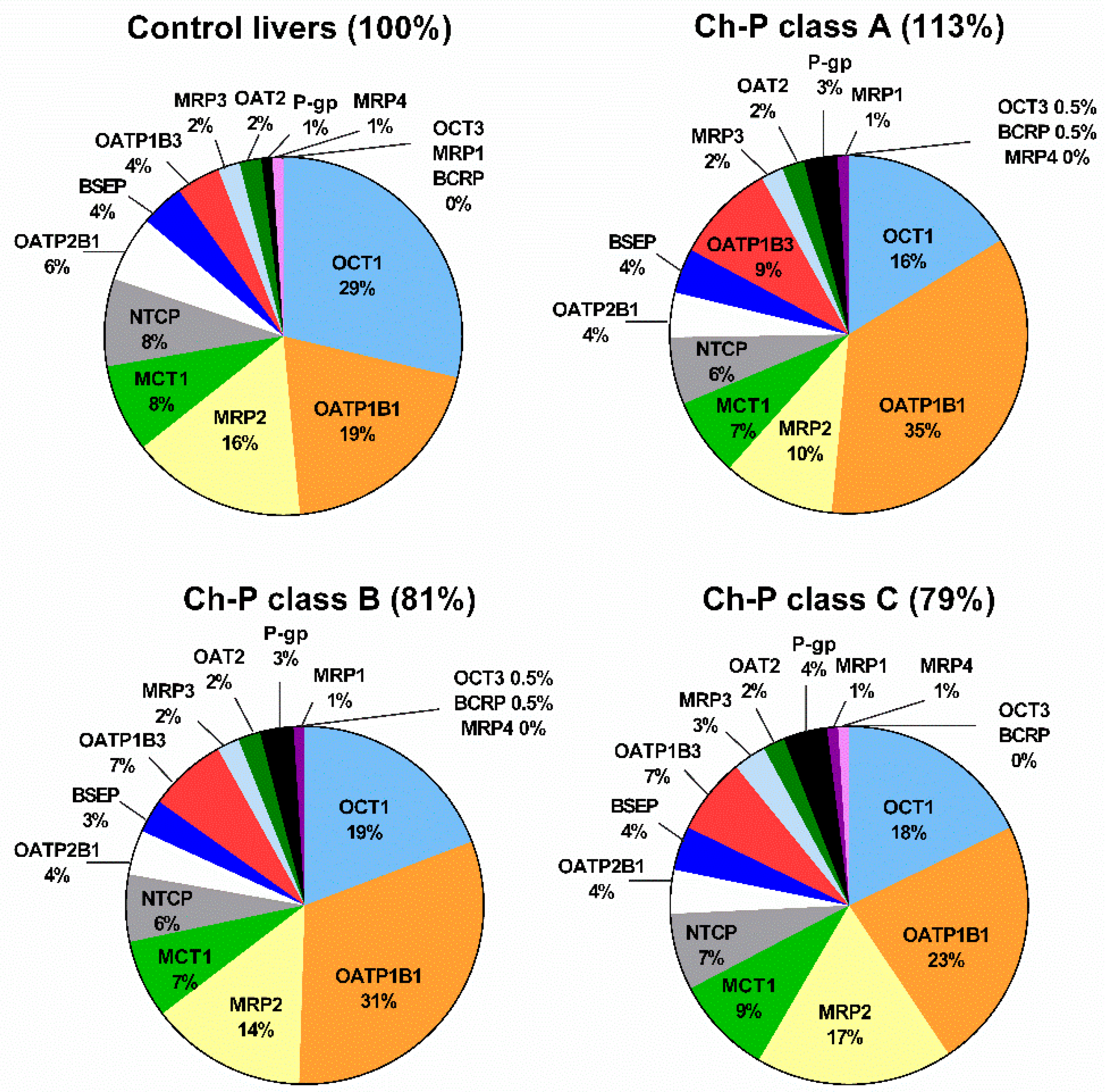
2. Results
2.1. mRNA Expression
2.2. Protein Abundance
3. Discussion
4. Materials and Methods
4.1. Liver Samples
4.2. mRNA Isolation and Quantitative Real-Time RT-PCR
4.3. Protein Quantification by Liquid Chromatography-Tandem Mass Spectrometry (LC−MC/MS)
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, D.K.; Chung, R.T. Overview of Direct-Acting Antiviral Drugs and Drug Resistance of Hepatitis C Virus. Methods Mol. Biol. 2019, 191, 3–32. [Google Scholar]
- Esposito, I.; Trinks, J.; Soriano, V. Hepatitis C virus resistance to the new direct-acting antivirals. Expert Opin. Drug Metab. Toxicol. 2016, 12, 1197–1209. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.; Chan, G.H.; Houle, R.; Lin, M.; Yabut, J.; Fandozzi, C. In Vitro Assessment of Transporter Mediated Perpetrator DDIs for Several Hepatitis C Virus Direct-Acting Antiviral Drugs and Prediction of DDIs with Statins Using Static Models. AAPS J. 2022, 24, 45. [Google Scholar] [CrossRef] [PubMed]
- Sjöstedt, N.; Adiwidjaja, J.; Murphy, W.A.; Brouwer, K.L.R. Drug transport in the liver. In Drug Transporters: Molecular Characterization and Role in Drug Disposition, 3rd ed.; You, G., Morris, M.E., Eds.; John Wiley & Sons: Boston, MA, USA, 2022. [Google Scholar]
- Smolders, E.J.; Jansen, A.; Ter Horst, P.; Rockstroh, J.; Back, D.J.; Burger, D.M. Viral Hepatitis C Therapy: Pharmacokinetic and Pharmacodynamic Considerations: A 2019 Update. Clin. Pharmacokinet. 2019, 58, 1237–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drozdzik, M.; Busch, D.; Lapczuk, J.; Müller, J.; Ostrowski, M.; Kurzawski, M.; Oswald, S. Protein Abundance of Clinically Relevant Drug Transporters in the Human Liver and Intestine: A Comparative Analysis in Paired Tissue Specimens. Clin. Pharmacol. Ther. 2019, 105, 1204–1212. [Google Scholar] [CrossRef]
- Billington, S.; Ray, A.S.; Salphati, L.; Xiao, G.; Chu, X.; Humphreys, W.G.; Liao, M.; Lee, C.A.; Mathias, A.; Hop, C.E.C.A.; et al. Transporter expression in noncancerous and cancerous liver tissue from donors with hepatocellular carcinoma and chronic Hepatitis C infection quantified by LC-MS/MS proteomics. Drug Metab. Dispos. 2018, 46, 189–196. [Google Scholar] [CrossRef]
- Wang, L.; Collins, C.; Kelly, E.J.; Chu, X.; Ray, A.S.; Salphati, L.; Xiao, G.; Lee, C.; Lai, Y.; Liao, M.; et al. Transporter expression in liver tissue from subjects with alcoholic or hepatitis C cirrhosis quantified by targeted quantitative proteomics. Drug Metab. Dispos. 2016, 44, 1752–1758. [Google Scholar] [CrossRef] [Green Version]
- El-Khateeb, E.; Achour, B.; Al-Majdoub, Z.M.; Barber, J.; Rostami-Hodjegan, A. Non-uniformity of Changes in Drug-Metabolizing Enzymes and Transporters in Liver Cirrhosis: Implications for Drug Dosage Adjustment. Mol. Pharm. 2021, 18, 3563–3577. [Google Scholar] [CrossRef]
- Drozdzik, M.; Szelag-Pieniek, S.; Post, M.; Zeair, S.; Wrzesinski, M.; Kurzawski, M.; Prieto, J.; Oswald, S. Protein Abundance of Hepatic Drug Transporters in Patients with Different Forms of Liver Damage. Clin. Pharmacol. Ther. 2020, 107, 1138–1148. [Google Scholar] [CrossRef]
- Droździk, M.; Szeląg-Pieniek, S.; Grzegółkowska, J.; Łapczuk-Romańska, J.; Post, M.; Domagała, P.; Miętkiewski, J.; Oswald, S.; Kurzawski, M. Monocarboxylate Transporter 1 (MCT1) in Liver Pathology. Int. J. Mol. Sci. 2020, 21, 1606. [Google Scholar] [CrossRef] [Green Version]
- Virgolini, I.; Müller, C.; Angelberger, P.; Höbart, J.; Bergmann, H.; Sinzinger, H. Functional liver imaging with 99Tcmgalactosylneoglycoalbumin (NGA) in alcoholic liver cirrhosis and liver fibrosis. Nucl. Med. Commun. 1991, 12, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Weiskirchen, R.; Weiskirchen, S.; Tacke, F. Recent advances in understanding liver fibrosis: Bridging basic science and individualized treatment concepts. F1000Research 2018, 7, 921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsia, C.Y.; Huo, T.I.; Chiang, S.Y.; Lu, M.F.; Sun, C.L.; Wu, J.C.; Lee, P.C.; Chi, C.W.; Lui, W.Y.; Lee, S.D. Evaluation of interleukin-6, interleukin-10 and human hepatocyte growth factor as tumor markers for hepatocellular carcinoma. Eur. J. Surg. Oncol. 2007, 33, 208–212. [Google Scholar] [CrossRef]
- Bertoletti, A.; D’Elios, M.M.; Boni, C.; De Carli, M.; Zignego, A.L.; Durazzo, M.; Missale, G.; Penna, A.; Fiaccadori, F.; Del Prete, G.; et al. Different cytokine profiles of intraphepatic T cells in chronic hepatitis B and hepatitis C virus infections. Gastroenterology 1997, 112, 193–199. [Google Scholar] [CrossRef]
- Sakurai, T.; He, G.; Matsuzawa, A.; Yu, G.Y.; Maeda, S.; Hardiman, G.; Karin, M. Hepatocyte necrosis induced by oxidative stress and IL-1α release mediate carcinogen-induced compensatory proliferation and liver tumorigenesis. Cancer Cell. 2008, 14, 156–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrivastava, S.; Mukherjee, A.; Ray, R.; Ray, R.B. Hepatitis C virus induces interleukin-1β (IL-1β)/IL-18 in circulatory and resident liver macrophages. J. Virol. 2013, 87, 12284–12290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, S.; Piquette-Miller, M. The involvement of the pregnane X receptor in hepatic gene regulation during inflammation in mice. J. Pharmacol. Exp. Ther. 2005, 312, 841–848. [Google Scholar] [CrossRef] [Green Version]
- Fardel, O.; Le Vee, M. Regulation of human hepatic drug transporter expression by pro-inflammatory cytokines. Exp. Opin. Drug Metab. Toxicol. 2009, 5, 1469–1481. [Google Scholar] [CrossRef]
- Xu, S.; Weerachayaphorn, J.; Cai, S.Y.; Soroka, C.J.; Boyer, J.L. Aryl hydrocarbon receptor and NF-E2-related factor 2 are key regulators of human MRP4 expression. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G126–G135. [Google Scholar] [CrossRef] [Green Version]
- Evers, R.; Piquette-Miller, M.; Polli, J.W.; Russel, F.; Sprowl, J.A.; Tohyama, K.; Ware, J.A.; de Wildt, S.N.; Xie, W.; Brouwer, K.; et al. Disease-Associated Changes in Drug Transporters May Impact the Pharmacokinetics and/or Toxicity of Drugs: A White Paper from the International Transporter Consortium. Clin. Pharmacol. Ther. 2018, 104, 900–915. [Google Scholar] [CrossRef]
- Kim, H.Y.; Choi, J.Y.; Park, C.H.; Song, M.; Song, D.S.; Kim, C.W.; Bae, S.H.; Yoon, S.K.; Lee, Y.J.; Rha, S.E. Clinical factors predictive of insufficient liver enhancement on the hepatocyte-phase of Gd-EOB-DTPA-enhanced magnetic resonance imaging in patients with liver cirrhosis. J. Gastroenterol. 2013, 48, 1180–1187. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Barton, H.A.; Maurer, T.S. A Mechanistic pharmacokinetic model for liver transporter substrates under liver cirrhosis conditions. CPT Pharmacomet. Syst. Pharmacol. 2015, 4, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Prasad, B.; Lai, Y.; Lin, Y.; Unadkat, J.D. Interindividual variability in the hepatic expression of the human breast cancer resistance protein (BCRP/ABCG2): Effect of age, sex, and genotype. J. Pharm. Sci. 2013, 102, 787–793. [Google Scholar] [CrossRef]
- Yang, Y.; Li, P.; Zhang, Z.; Wang, Z.; Liu, L.; Liu, X. Prediction of Cyclosporin-Mediated Drug Interaction Using Physiologically Based Pharmacokinetic Model Characterizing Interplay of Drug Transporters and Enzymes. Int. J. Mol. Sci. 2020, 21, 7023. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.; Prasad, B.; Neuhoff, S.; Yoshida, K.; Leeder, J.S.; Mukherjee, D.; Taskar, K.; Varma, M.V.S.; Zhang, X.; Yang, X.; et al. Clinical Implications of Altered Drug Transporter Abundance/Function and PBPK Modeling in Specific Populations: An ITC Perspective. Clin. Pharmacol. Ther. 2022. ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Néant, N.; Solas, C. Drug-Drug Interactions Potential of Direct-Acting Antivirals for the treatment of Chronic Hepatitis C Virus infection. Int. J. Antimicrob. Agents 2020, 56, 105571. [Google Scholar] [CrossRef]
- Pol, S.; Haour, G.; Fontaine, H.; Dorival, C.; Petrov-Sanchez, V.; Bourliere, M.; Capeau, J.; Carrieri, P.; Larrey, D.; Larsen, C.; et al. The negative impact of HBV/HCV coinfection on cirrhosis and its consequences. Aliment. Pharmacol. Ther. 2017, 46, 1054–1060. [Google Scholar] [CrossRef] [Green Version]
- Sabariegos, R.; Picazo, F.; Domingo, B.; Franco, S.; Martinez, M.A.; Llopis, J. Fluorescence resonance energy transfer-based assay for characterization of hepatitis C virus NS3-4A protease in live cells. Antimicrob. Agents Chemother. 2009, 53, 728–734. [Google Scholar] [CrossRef] [Green Version]
- Arias-Arias, J.L.; MacPherson, D.J.; Hill, M.E.; Hardy, J.A.; Mora-Rodríguez, R. A fluorescence-activatable reporter of flavivirus NS2B-NS3 protease activity enables live imaging of infection in single cells and viral plaques. J. Bio. Chem. 2020, 295, 2212–2226. [Google Scholar] [CrossRef]
- Marcuello, C.; Frempong, G.A.; Balsera, M.; Medina, M.; Lostao, A. Atomic Force Microscopy to Elicit Conformational Transitions of Ferredoxin-Dependent Flavin Thioredoxin Reductases. Antioxidants 2021, 90, 1437. [Google Scholar] [CrossRef]
- Vega, S.; Neira, J.L.; Marcuello, C.; Lostao, A.; Abian, O.; Velazquez-Campoy, A. NS3 protease from hepatitis C virus: Biophysical studies on an intrinsically disordered protein domain. Int. J. Mol. Sci. 2013, 14, 13282–13306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frimayanti, N.; Chee, C.F.; Zain, S.M.; Rahman, N. A Design of new competitive dengue NS2B/NS3 protease inhibitors-a computational approach. Int. J. Mol. Sci. 2011, 12, 1089–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Costa, R.A.; Rocha, J.A.P.D.; Pinheiro, A.S.; Costa, A.D.S.S.D.; Rocha, E.C.M.D.; Silva, R.C.; Gonçalves, A.D.S.; Santos, C.B.R.; Brasil, D.D.S.B. A Computational Approach Applied to the Study of Potential Allosteric Inhibitors Protease NS2B/NS3 from Dengue Virus. Molecules 2022, 27, 4118. [Google Scholar] [CrossRef] [PubMed]
- Quek, J.-P.; Ser, Z.; Chew, B.L.A.; Li, X.; Wang, L.; Sobota, R.M.; Luo, D.; Phoo, W.W. Dynamic Interactions of Post Cleaved NS2B Cofactor and NS3 Protease Identified by Integrative Structural Approaches. Viruses 2022, 14, 1440. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Gröer, C.; Brück, S.; Lai, Y.; Paulick, A.; Busemann, A.; Heidecke, C.D.; Siegmund, W.; Oswald, S. LC-MS/MS-based quantification of clinically relevant intestinal uptake and efflux transporter proteins. J. Pharm. Biomed. Anal. 2013, 85, 253–261. [Google Scholar] [CrossRef]




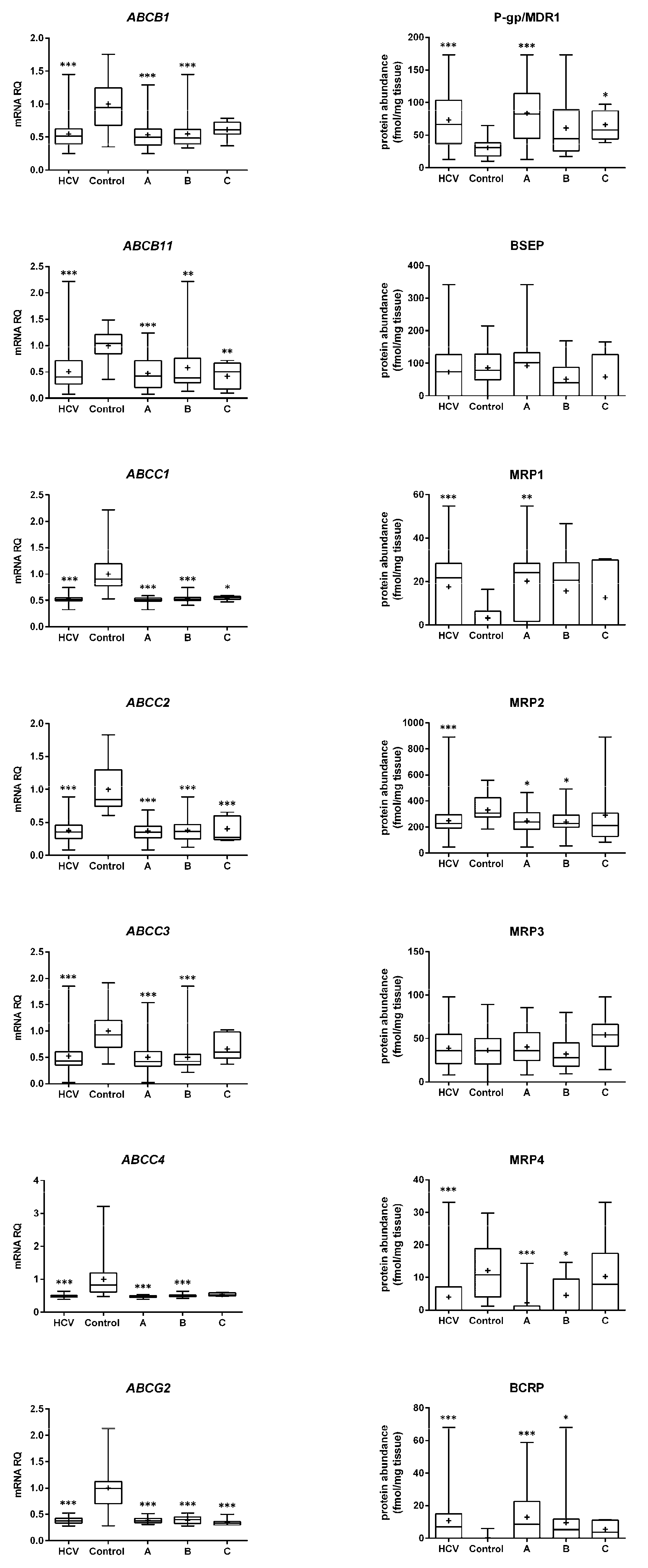{kind=link}
{kind=link}
{kind=link}
| Gene/Protein | Control n = 20 | HCV n = 58 | Child–Pugh Class A n = 30 | Child–Pugh Class B n = 21 | Child–Pugh Class C n = 7 |
|---|---|---|---|---|---|
| ABCB1/P-gp | 0.432 | −0.251 | −0.174 | −0.270 | −0.393 |
| ABCC1/MRP1 | −0.481 * | 0.168 | 0.157 | 0.291 | −0.079 |
| ABCC2/MRP2 | 0.206 | 0.375 ** | 0.326 | 0.331 | 0.607 |
| ABCC3/MRP3 | 0.029 | 0.264 * | 0.304 | 0.092 | 0.571 |
| ABCC4/MRP4 | −0.220 | 0.304 * | 0.430 * | 0.041 | −0.185 |
| ABCG2/BCRP | LLOQ | −0.322* | 0.012 | −0.578 ** | −0.811 * |
| ABCB11/BSEP | 0.021 | 0.011 | 0.063 | −0.069 | 0.197 |
| SLC10A1/NTCP | 0.066 | 0.268 * | 0.189 | 0.357 | 0.321 |
| SLC22A1/OCT1 | 0.424 | 0.265 * | 0.195 | 0.104 | 0.714 |
| SLC22A3/OCT3 | 0.296 | −0.228 | −0.289 | −0.281 | 0.126 |
| SLC22A7/OAT2 | 0.302 | 0.412 ** | 0.270 | 0.325 | 0.214 |
| SLCO1B1/OATP1B1 | 0.295 | −0.030 | −0.071 | −0.392 | 0.179 |
| SLCO1B3/OATP1B3 | 0.519 * | 0.025 | −0.174 | −0.234 | 0.893 ** |
| SLCO2B1/OATP2B1 | −0.077 | 0.191 | −0.086 | 0.436 * | 0.214 |
| This Study n = 58 | Billington et al. [7] ^ n = 30 | Wang et al. [8] n = 30 | El-Khateeb et al. [9] # n = 9 | Drozdzik et al. [10] n = 21 | |
|---|---|---|---|---|---|
| P-gp | ↑ | ↔ | ↓ | ↔ | |
| MRP1 | ↑ | ↔ | |||
| MRP2 | ↓ | ↓ | ↓ | ↓ | ↔ |
| MRP3 | ↔ | ↔ | ↔ | ↔ | ↔ |
| MRP4 | ↓ | ↔ | |||
| BCRP | ↑ | ↔ | ↔ | ↔ | |
| BSEP | ↔ | ↓ | ↓ | ↓ | |
| NTCP | ↓ | ↓ | ↓ | ↔ | |
| MCT1 | ↔ | ↔ * | |||
| OCT1 | ↓ | ↓ | ↓ | ↓ | |
| OCT3 | ↔ | ↔ | |||
| OAT2 | ↔ | ↔ | ↔ | ||
| OATP1B1 | ↔ | ↔ | ↔ | ↓ | |
| OATP1B3 | ↑ | ↓ | ↓ | ↔ | |
| OATP2B1 | ↓ | ↔ | ↔ | ↓ | ↓ |
| Parameter/Disease | Population Normal | Control n = 20 | HCV n = 58 | Ch-P A n = 30 | Ch-P B n = 21 | Ch-P C n = 7 |
|---|---|---|---|---|---|---|
| Sex (male/female) | 11/9 | 30/28 | 16/14 | 11/10 | 3/4 | |
| Age (years) | 63 ± 10 | 56 ± 7 | 57 ± 7 | 55 ± 8 | 52 ± 9 | |
| Total bilirubin (mg/dL) | 0.1–1.2 | 0.59 ± 0.25 | 1.75 ± 1.26 | 1.03 ± 0.57 | 2.05 ± 0.84 | 3.62 ± 1.78 |
| Albumin (g/dL) | 3.4–5.4 | 3.89 ± 0.38 | 3.38 ± 0.57 | 3.67 ± 0.49 | 3.23 ± 0.45 | 2.71 ± 0.40 |
| INR | 0.9–1.1 | 1.14 ± 0.21 | 1.30 ± 0.28 | 1.20 ± 0.22 | 1.29 ± 0.17 | 1.71 ± 0.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Droździk, M.; Lapczuk-Romanska, J.; Wenzel, C.; Skalski, Ł.; Szeląg-Pieniek, S.; Post, M.; Syczewska, M.; Kurzawski, M.; Oswald, S. Protein Abundance of Drug Transporters in Human Hepatitis C Livers. Int. J. Mol. Sci. 2022, 23, 7947. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23147947
Droździk M, Lapczuk-Romanska J, Wenzel C, Skalski Ł, Szeląg-Pieniek S, Post M, Syczewska M, Kurzawski M, Oswald S. Protein Abundance of Drug Transporters in Human Hepatitis C Livers. International Journal of Molecular Sciences. 2022; 23(14):7947. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23147947
Chicago/Turabian StyleDroździk, Marek, Joanna Lapczuk-Romanska, Christoph Wenzel, Łukasz Skalski, Sylwia Szeląg-Pieniek, Mariola Post, Marta Syczewska, Mateusz Kurzawski, and Stefan Oswald. 2022. "Protein Abundance of Drug Transporters in Human Hepatitis C Livers" International Journal of Molecular Sciences 23, no. 14: 7947. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23147947