
Proteome-Wide Differential Effects of Peritoneal Dialysis Fluid Properties in an In Vitro Human Endothelial Cell Model

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Alterations Induced by Different PD Fluids in Endothelial Cells
2.2. PD Fluid Exposure Leads to Distinct Common and Fluid Specific Molecular Alterations in Endothelial Cells
2.3. Specific Protein and Biological Processes Regulated by Individual PD Fluids
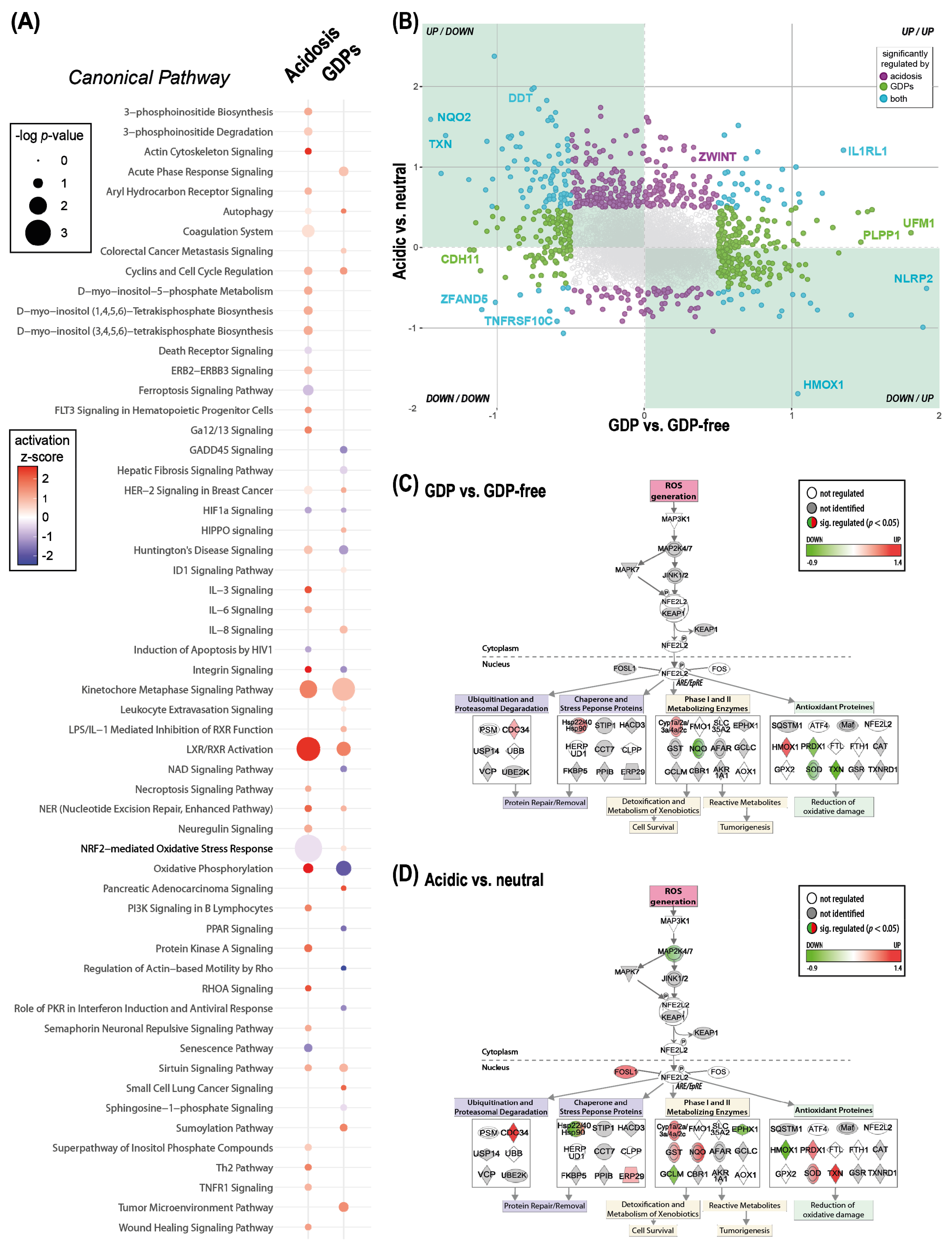
2.4. Major Properties of PD Fluids Perturb Common Proteins and Pathways but Not Always in the Same Direction
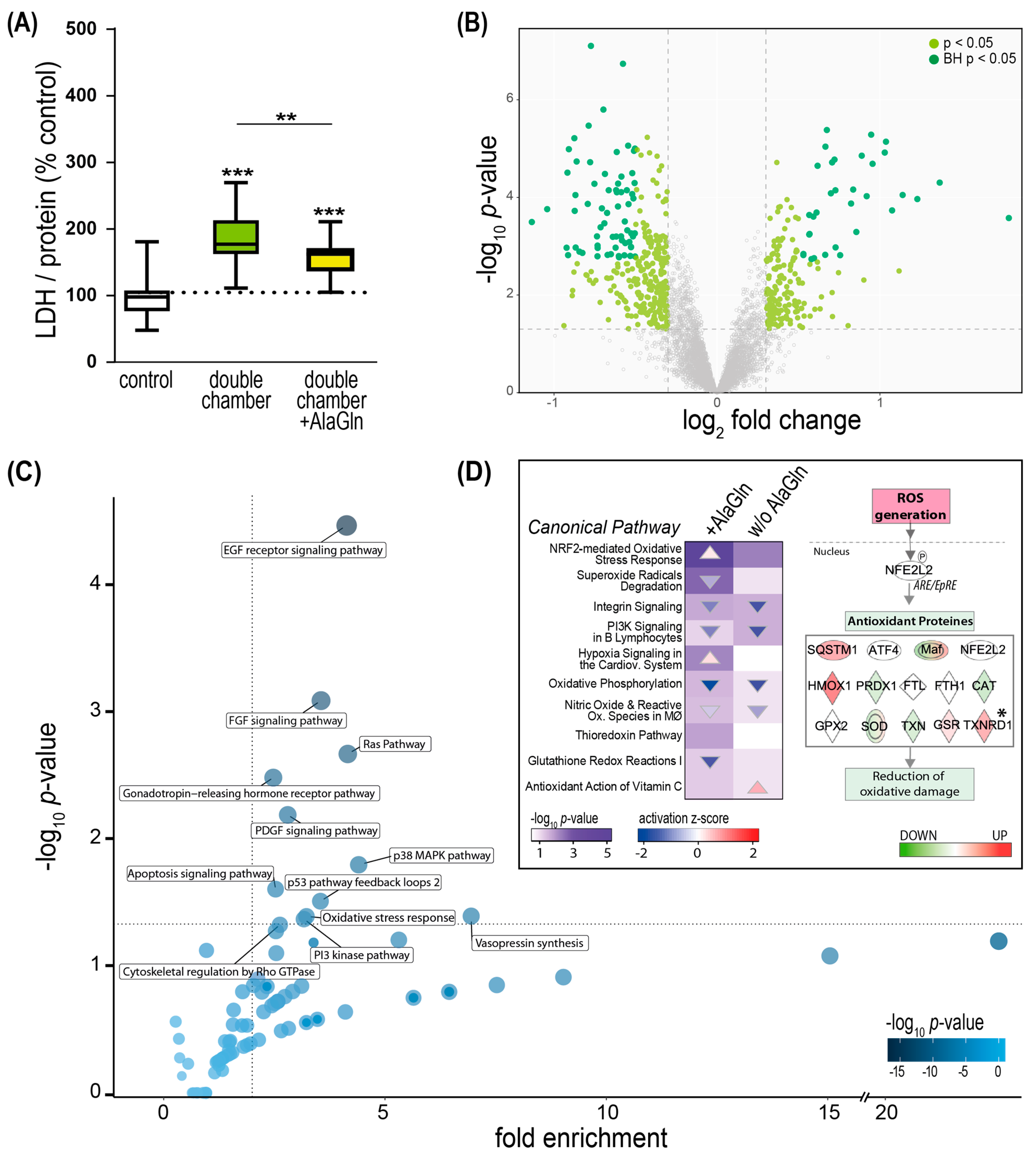
2.5. PD Fluid Induced Perturbation of the Redox Stress Signaling Can Be Counteracted by the Addition of Alanyl-Glutamine
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cell Culture
5.2. Experimental PD Fluid Exposure Setting
5.3. Cell Damage Assay
5.4. Protein Sample Preparation
5.5. SP3 and Tandem Mass Tag (TMT) Labeling
5.6. Mass Spectrometry Data Analysis
5.7. Enrichment Map Analysis
5.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Teitelbaum, I. Peritoneal Dialysis. N. Engl. J. Med. 2021, 385, 1786–1795. [Google Scholar] [CrossRef] [PubMed]
- Morelle, J.; Stachowska-Pietka, J.; Öberg, C.; Gadola, L.; La Milia, V.; Yu, Z.; Lambie, M.; Mehrotra, R.; de Arteaga, J.; Davies, S. ISPD recommendations for the evaluation of peritoneal membrane dysfunction in adults: Classification, measurement, interpretation and rationale for intervention. Perit. Dial. Int. 2021, 41, 352–372. [Google Scholar] [CrossRef] [PubMed]
- Mistry, C.D.; Gokal, R.; Peers, E.; Brown, C.; Smith, S.; Edwards, D.; Junor, B.; Gordon, A.; McMillan, M.; Robertson, M.; et al. A randomized multicenter clinical trial comparing isosmolar Icodextrin with hyperosmolar glucose solutions in CAPD. Kidney Int. 1994, 46, 496–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morelle, J.; Sow, A.; Fustin, C.-A.; Fillée, C.; Garcia-Lopez, E.; Lindholm, B.; Goffin, E.; Vandemaele, F.; Rippe, B.; Öberg, C.M.; et al. Mechanisms of Crystalloid versus Colloid Osmosis across the Peritoneal Membrane. J. Am. Soc. Nephrol. 2018, 29, 1875–1886. [Google Scholar] [CrossRef]
- Bartosova, M.; Schmitt, C.P. Biocompatible Peritoneal Dialysis: The Target Is Still Way Off. Front. Physiol. 2019, 9, 1853. [Google Scholar] [CrossRef]
- Goffin, E. Peritoneal Membrane Structural and Functional Changes during Peritoneal Dialysis. Semin. Dial. 2008, 21, 258–265. [Google Scholar] [CrossRef]
- Balzer, M.S. Molecular pathways in peritoneal fibrosis. Cell. Signal. 2020, 75, 109778. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Lara-Pezzi, E.; Selgas, R.; Ramírez-Huesca, M.; Domínguez-Jiménez, C.; Jiménez-Heffernan, J.A.; Aguilera, A.; Sánchez-Tomero, J.A.; Bajo, M.A.; Álvarez, V.; et al. Peritoneal Dialysis and Epithelial-to-Mesenchymal Transition of Mesothelial Cells. N. Engl. J. Med. 2003, 348, 403–413. [Google Scholar] [CrossRef]
- Büchel, J.; Bartosova, M.; Eich, G.; Wittenberger, T.; Klein-Hitpass, L.; Steppan, S.; Hackert, T.; Schaefer, F.; Passlick-Deetjen, J.; Schmitt, C.P. Interference of Peritoneal Dialysis Fluids with Cell Cycle Mechanisms. Perit. Dial. Int. J. Int. Soc. Perit. Dial. 2015, 35, 259–274. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, C.P.; Aufricht, C. Is there such a thing as biocompatible peritoneal dialysis fluid? Pediatr. Nephrol. 2017, 32, 1835–1843. [Google Scholar] [CrossRef] [Green Version]
- Herzog, R.; Sacnun, J.M.; González-Mateo, G.; Bartosova, M.; Bialas, K.; Wagner, A.; Unterwurzacher, M.; Sobieszek, I.J.; Daniel-Fischer, L.; Rusai, K.; et al. Lithium preserves peritoneal membrane integrity by suppressing mesothelial cell αB-crystallin. Sci. Transl. Med. 2021, 13, eaaz9705. [Google Scholar] [CrossRef] [PubMed]
- Bender, T.O.; Kratochwill, K.; Herzog, R.; Ulbrich, A.; Böhm, M.; Jörres, A.; Aufricht, C. Injury-Induced Inflammation and Inadequate HSP Expression in Mesothelial Cells upon Repeat Exposure to Dual-Chamber Bag Peritoneal Dialysis Fluids. Int. J. Artif. Organs 2015, 38, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Bajo, M.A.; Pérez-Lozano, M.L.; Albar-Vizcaino, P.; del Peso, G.; Castro, M.-J.; Gonzalez-Mateo, G.; Fernández-Perpén, A.; Aguilera, A.; Sánchez-Villanueva, R.; Sánchez-Tomero, J.A.; et al. Low-GDP peritoneal dialysis fluid (’balance’) has less impact in vitro and ex vivo on epithelial-to-mesenchymal transition (EMT) of mesothelial cells than a standard fluid. Nephrol. Dial. Transplant. 2010, 26, 282–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eich, G.; Bartosova, M.; Tischer, C.; Wlodkowski, T.T.; Schaefer, B.; Pichl, S.; Kraewer, N.; Ranchin, B.; Vondrak, K.; Liebau, M.C.; et al. Bicarbonate buffered peritoneal dialysis fluid upregulates angiopoietin-1 and promotes vessel maturation. PLoS ONE 2017, 12, e0189903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonomini, M.; Di Silvestre, S.; Di Tomo, P.; Di Pietro, N.; Mandatori, D.; Di Liberato, L.; Sirolli, V.; Chiarelli, F.; Indiveri, C.; Pandolfi, A.; et al. Effect of peritoneal dialysis fluid containing osmo-metabolic agents on human endothelial cells. Drug Des. Dev. Ther. 2016, 10, 3925–3932. [Google Scholar] [CrossRef] [Green Version]
- Misian, M.; Baum, E.; Bręborowicz, A. Secretory activity of the coronary artery endothelial cells in conditions of the peritoneal dialysis. Ren. Fail. 2022, 44, 54–61. [Google Scholar] [CrossRef]
- Bartosova, M.; Zarogiannis, S.G.; Schmitt, C.P.; Members of the International Pediatric Peritoneal Biobank. How peritoneal dialysis transforms the peritoneum and vasculature in children with chronic kidney disease-what can we learn for future treatment? Mol. Cell. Pediatr. 2022, 9, 9. [Google Scholar] [CrossRef]
- Herzog, R.; Bartosova, M.; Tarantino, S.; Wagner, A.; Unterwurzacher, M.; Sacnun, J.M.; Lichtenauer, A.M.; Kuster, L.; Schaefer, B.; Alper, S.L.; et al. Peritoneal Dialysis Fluid Supplementation with Alanyl-Glutamine Attenuates Conventional Dialysis Fluid-Mediated Endothelial Cell Injury by Restoring Perturbed Cytoprotective Responses. Biomolecules 2020, 10, 1678. [Google Scholar] [CrossRef]
- Dubin, R.F.; Rhee, E.P. Proteomics and Metabolomics in Kidney Disease, including Insights into Etiology, Treatment, and Prevention. Clin. J. Am. Soc. Nephrol. 2019, 15, 404–411. [Google Scholar] [CrossRef] [Green Version]
- Herzog, R.; Boehm, M.; Unterwurzacher, M.; Wagner, A.; Parapatics, K.; Májek, P.; Mueller, A.C.; Lichtenauer, A.; Bennett, K.L.; Alper, S.L.; et al. Effects of Alanyl-Glutamine Treatment on the Peritoneal Dialysis Effluent Proteome Reveal Pathomechanism-Associated Molecular Signatures. Mol. Cell. Proteom. 2018, 17, 516–532. [Google Scholar] [CrossRef] [Green Version]
- Bonomini, M.; Borras, F.E.; Troya-Saborido, M.; Carreras-Planella, L.; Di Liberato, L.; Arduini, A. Proteomic Research in Peritoneal Dialysis. Int. J. Mol. Sci. 2020, 21, 5489. [Google Scholar] [CrossRef] [PubMed]
- Kratochwill, K. The Extracorporeal Proteome-The Significance of Selective Protein Removal During Dialysis Therapy. Proteom.–Clin. Appl. 2018, 12, e1800078. [Google Scholar] [CrossRef] [PubMed]
- Bartosova, M.; Herzog, R.; Ridinger, D.; Levai, E.; Jenei, H.; Zhang, C.; Mateo, G.T.G.; Marinovic, I.; Hackert, T.; Bestvater, F.; et al. Alanyl-Glutamine Restores Tight Junction Organization after Disruption by a Conventional Peritoneal Dialysis Fluid. Biomolecules 2020, 10, 1178. [Google Scholar] [CrossRef] [PubMed]
- Boehm, M.; Herzog, R.; Klinglmüller, F.; Lichtenauer, A.M.; Wagner, A.; Unterwurzacher, M.; Beelen, R.H.J.; Alper, S.L.; Aufricht, C.; Kratochwill, K. The Peritoneal Surface Proteome in a Model of Chronic Peritoneal Dialysis Reveals Mechanisms of Membrane Damage and Preservation. Front. Physiol. 2019, 10, 472. [Google Scholar] [CrossRef]
- Vychytil, A.; Herzog, R.; Probst, P.; Ribitsch, W.; Lhotta, K.; Machold-Fabrizii, V.; Wiesholzer, M.; Kaufmann, M.; Salmhofer, H.; Windpessl, M.; et al. A randomized controlled trial of alanyl-glutamine supplementation in peritoneal dialysis fluid to assess impact on biomarkers of peritoneal health. Kidney Int. 2018, 94, 1227–1237. [Google Scholar] [CrossRef] [Green Version]
- Kratochwill, K.; Boehm, M.; Herzog, R.; Lichtenauer, A.M.; Salzer, E.; Lechner, M.; Kuster, L.; Bergmeister, K.; Rizzi, A.; Mayer, B.; et al. Alanyl–glutamine dipeptide restores the cytoprotective stress proteome of mesothelial cells exposed to peritoneal dialysis fluids. Nephrol. Dial. Transplant. 2011, 27, 937–946. [Google Scholar] [CrossRef]
- Basha, B.; Samuel, S.M.; Triggle, C.R.; Ding, H. Endothelial dysfunction in diabetes mellitus: Possible involvement of endoplasmic reticulum stress? Exp. Diabetes Res. 2012, 2012, 481840. [Google Scholar] [CrossRef] [Green Version]
- Bartosova, M.; Zhang, C.; Schaefer, B.; Herzog, R.; Ridinger, D.; Damgov, I.; Levai, E.; Marinovic, I.; Eckert, C.; Romero, P.; et al. Glucose Derivative Induced Vasculopathy in Children on Chronic Peritoneal Dialysis. Circ. Res. 2021, 129, e102–e118. [Google Scholar] [CrossRef]
- Bartosova, M.; Schaefer, B.; Bermejo, J.L.; Tarantino, S.; Lasitschka, F.; Macher-Goeppinger, S.; Sinn, H.-P.; Warady, B.A.; Zaloszyc, A.; Parapatics, K.; et al. Complement Activation in Peritoneal Dialysis–Induced Arteriolopathy. J. Am. Soc. Nephrol. 2017, 29, 268–282. [Google Scholar] [CrossRef]
- Bartosova, M.; Schaefer, B.; Vondrak, K.; Sallay, P.; Taylan, C.; Cerkauskiene, R.; Dzierzega, M.; Milosevski-Lomic, G.; Büscher, R.; Zaloszyc, A.; et al. Peritoneal Dialysis Vintage and Glucose Exposure but Not Peritonitis Episodes Drive Peritoneal Membrane Transformation During the First Years of PD. Front. Physiol. 2019, 10, 356. [Google Scholar] [CrossRef]
- Rippe, B. Peritoneal Angiogenesis in Response to Dialysis Fluid. Perit. Dial. Basic Concepts Clin. Excell. 2009, 163, 60–66. [Google Scholar] [CrossRef]
- Cho, Y.; Johnson, D.W.; Craig, J.C.; Strippoli, G.F.; Badve, S.V.; Wiggins, K.J. Biocompatible dialysis fluids for peritoneal dialysis. Cochrane Database Syst. Rev. 2014, 10, CD007554. [Google Scholar] [CrossRef] [PubMed]
- Retana, C.; Sanchez, E.; Perez-Lopez, A.; Cruz, A.; Lagunas, J.; Cruz, C.; Vital, S.; Reyes, J.L. Alterations of Intercellular Junctions in Peritoneal Mesothelial Cells from Patients Undergoing Dialysis: Effect of Retinoic Acid. Perit. Dial. Int. J. Int. Soc. Perit. Dial. 2015, 35, 275–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corciulo, S.; Nicoletti, M.C.; Mastrofrancesco, L.; Milano, S.; Mastrodonato, M.; Carmosino, M.; Gerbino, A.; Corciulo, R.; Russo, R.; Svelto, M.; et al. AQP1-Containing Exosomes in Peritoneal Dialysis Effluent As Biomarker of Dialysis Efficiency. Cells 2019, 8, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stachowska-Pietka, J.; Poleszczuk, J.; Flessner, M.F.; Lindholm, B.; Waniewski, J. Alterations of peritoneal transport characteristics in dialysis patients with ultrafiltration failure: Tissue and capillary components. Nephrol. Dial. Transplant. 2018, 34, 864–870. [Google Scholar] [CrossRef]
- Morelle, J.; Marechal, C.; Yu, Z.; Debaix, H.; Corre, T.; Lambie, M.; Verduijn, M.; Dekker, F.; Bovy, P.; Evenepoel, P.; et al. AQP1 Promoter Variant, Water Transport, and Outcomes in Peritoneal Dialysis. N. Engl. J. Med. 2021, 385, 1570–1580. [Google Scholar] [CrossRef]
- Lopez-Anton, M.; Rudolf, A.; Baird, D.M.; Roger, L.; Jones, R.E.; Witowski, J.; Fraser, D.J.; Bowen, T. Telomere length profiles in primary human peritoneal mesothelial cells are consistent with senescence. Mech. Ageing Dev. 2017, 164, 37–40. [Google Scholar] [CrossRef] [Green Version]
- Ksiazek, K.; Korybalska, K.; Jorres, A.; Witowski, J. Accelerated senescence of human peritoneal mesothelial cells exposed to high glucose: The role of TGF-beta1. Lab. Investig. 2007, 87, 345–356. [Google Scholar] [CrossRef] [Green Version]
- Masola, V.; Bonomini, M.; Borrelli, S.; Di Liberato, L.; Vecchi, L.; Onisto, M.; Gambaro, G.; Palumbo, R.; Arduini, A. Fibrosis of Peritoneal Membrane as Target of New Therapies in Peritoneal Dialysis. Int. J. Mol. Sci. 2022, 23, 4831. [Google Scholar] [CrossRef]
- Książek, K.; Bręborowicz, A.; Jörres, A.; Witowski, J. Oxidative stress contributes to accelerated development of the senescent phenotype in human peritoneal mesothelial cells exposed to high glucose. Free Radic. Biol. Med. 2007, 42, 636–641. [Google Scholar] [CrossRef]
- Raby, A.-C.; Colmont, C.S.; Kift-Morgan, A.; Koehl, J.; Eberl, M.; Fraser, D.; Topley, N.; Labéta, M.O. Toll-Like Receptors 2 and 4 Are Potential Therapeutic Targets in Peritoneal Dialysis–Associated Fibrosis. J. Am. Soc. Nephrol. 2016, 28, 461–478. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.B.; Yu, M.R.; Song, J.S.; Ha, H. Reactive oxygen species amplify protein kinase C signaling in high glucose-induced fibronectin expression by human peritoneal mesothelial cells. Kidney Int. 2004, 65, 1170–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ksiazek, K.; Mikuła-Pietrasik, J.; Jörres, A.; Witowski, J. Oxidative stress-mediated early senescence contributes to the short replicative life span of human peritoneal mesothelial cells. Free Radic. Biol. Med. 2008, 45, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Hung, K.-Y.; Liu, S.-Y.; Yang, T.-C.; Liao, T.-L.; Kao, S.-H. High-Dialysate-Glucose-Induced Oxidative Stress and Mitochondrial-Mediated Apoptosis in Human Peritoneal Mesothelial Cells. Oxid. Med. Cell. Longev. 2014, 2014, 642793. [Google Scholar] [CrossRef] [Green Version]
- Dou, L.; Jourde-Chiche, N.; Faure, V.; Cerini, C.; Berland, Y.; Dignat-George, F.; Brunet, P. The uremic solute indoxyl sulfate induces oxidative stress in endothelial cells. J. Thromb. Haemost. 2007, 5, 1302–1308. [Google Scholar] [CrossRef]
- Roumeliotis, S.; Dounousi, E.; Salmas, M.; Eleftheriadis, T.; Liakopoulos, V. Unfavorable Effects of Peritoneal Dialysis Solutions on the Peritoneal Membrane: The Role of Oxidative Stress. Biomolecules 2020, 10, 768. [Google Scholar] [CrossRef] [PubMed]
- Liakopoulos, V.; Roumeliotis, S.; Gorny, X.; Eleftheriadis, T.; Mertens, P.R. Oxidative Stress in Patients Undergoing Peritoneal Dialysis: A Current Review of the Literature. Oxidative Med. Cell. Longev. 2017, 2017, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kratochwill, K.; Boehm, M.; Herzog, R.; Gruber, K.; Lichtenauer, A.M.; Kuster, L.; Csaicsich, D.; Gleiss, A.; Alper, S.L.; Aufricht, C.; et al. Addition of Alanyl-Glutamine to Dialysis Fluid Restores Peritoneal Cellular Stress Responses—A First-In-Man Trial. PLoS ONE 2016, 11, e0165045. [Google Scholar] [CrossRef] [Green Version]
- Herzog, R.; Bender, T.O.; Vychytil, A.; Bialas, K.; Aufricht, C.; Kratochwill, K. DynamicO-LinkedN-Acetylglucosamine Modification of Proteins Affects Stress Responses and Survival of Mesothelial Cells Exposed to Peritoneal Dialysis Fluids. J. Am. Soc. Nephrol. 2014, 25, 2778–2788. [Google Scholar] [CrossRef] [Green Version]
- Ferrantelli, E.; Liappas, G.; Vila Cuenca, M.; Keuning, E.D.; Foster, T.L.; Vervloet, M.G.; Lopez-Cabrera, M.; Beelen, R.H. The dipeptide alanyl-glutamine ameliorates peritoneal fibrosis and attenuates IL-17 dependent pathways during peritoneal dialysis. Kidney Int. 2016, 89, 625–635. [Google Scholar] [CrossRef]
- Curi, R.; Newsholme, P.; Procopio, J.; Lagranha, C.; Gorjao, R.; Pithon-Curi, T.C. Glutamine, gene expression, and cell function. Front. Biosci. 2007, 12, 344–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, E. Nonnutritive Effects of Glutamine. J. Nutr. 2008, 138, 2025S–2031S. [Google Scholar] [CrossRef]
- Wiesenhofer, F.M.; Herzog, R.; Boehm, M.; Wagner, A.; Unterwurzacher, M.; Kasper, D.C.; Alper, S.L.; Vychytil, A.; Aufricht, C.; Kratochwill, K. Targeted Metabolomic Profiling of Peritoneal Dialysis Effluents Shows Anti-oxidative Capacity of Alanyl-Glutamine. Front. Physiol. 2019, 9, 1961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordberg, J.; Arnér, E.S. Reactive oxygen species, antioxidants, and the mammalian thioredoxin system. Free Radic. Biol. Med. 2001, 31, 1287–1312. [Google Scholar] [CrossRef]
- Vanholder, R.; Baurmeister, U.; Brunet, P.; Cohen, G.; Glorieux, G.; Jankowski, J. A Bench to Bedside View of Uremic Toxins. J. Am. Soc. Nephrol. 2008, 19, 863–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haynes, W.A.; Tomczak, A.; Khatri, P. Gene annotation bias impedes biomedical research. Sci. Rep. 2018, 8, 1362. [Google Scholar] [CrossRef] [Green Version]
- Oba, T.; Tatsunami, R.; Sato, K.; Takahashi, K.; Hao, Z.; Tampo, Y. Methylglyoxal has deleterious effects on thioredoxin in human aortic endothelial cells. Environ. Toxicol. Pharmacol. 2012, 34, 117–126. [Google Scholar] [CrossRef]
- Schmitz, A.E.; de Souza, L.; Santos, B.; Maher, P.; Lopes, F.M.; Londero, G.F.; Klamt, F.; Dafre, A.L. Methylglyoxal-Induced Protection Response and Toxicity: Role of Glutathione Reductase and Thioredoxin Systems. Neurotox. Res. 2017, 32, 340–350. [Google Scholar] [CrossRef]
- Fujino, G.; Noguchi, T.; Matsuzawa, A.; Yamauchi, S.; Saitoh, M.; Takeda, K.; Ichijo, H. Thioredoxin and TRAF Family Proteins Regulate Reactive Oxygen Species-Dependent Activation of ASK1 through Reciprocal Modulation of the N-Terminal Homophilic Interaction of ASK1. Mol. Cell. Biol. 2007, 27, 8152–8163. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, M.E.; Belinda, P.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, G.; Chaerkady, R.; Yu, W.; Hizal, D.B.; Hess, S.; Zhao, W.; Lekstrom, K.; Guo, X.; White, W.I.; Roskos, L.; et al. Statistical Models for the Analysis of Isobaric Tags Multiplexed Quantitative Proteomics. J. Proteome Res. 2017, 16, 3124–3136. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PD Fluid | Osmolality (mOsm/L) | GDPs | pH | Osmotic Agent | Number of Proteins * (BH p < 0.05) | Number of Proteins * (BH p < 0.01) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| All | Up | Down | All | Up | Down | |||||
| Single-chamber bag (“Conventional”) | 483 | ++++ | ~5.5 | 3.86% glucose | 2468 | 1342 | 1126 | 1778 | 955 | 823 |
| Double-chamber bag (“Biocompatible”) | 483 | ++ | ~7.4 | 3.86% glucose | 1677 | 787 | 890 | 946 | 406 | 540 |
| Icodextrin (“Polyglucose”) | 280–300 | + | ~5.5 | 7.5% icodextrin | 726 | 274 | 452 | 360 | 126 | 234 |
| Filter (“Lab made”) | 483 | - | ~5.5 | 3.86% glucose | 3232 | 1387 | 1845 | 2510 | 1026 | 1484 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sacnun, J.M.; Hoogenboom, R.; Eibensteiner, F.; Sobieszek, I.J.; Unterwurzacher, M.; Wagner, A.; Herzog, R.; Kratochwill, K. Proteome-Wide Differential Effects of Peritoneal Dialysis Fluid Properties in an In Vitro Human Endothelial Cell Model. Int. J. Mol. Sci. 2022, 23, 8010. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23148010
Sacnun JM, Hoogenboom R, Eibensteiner F, Sobieszek IJ, Unterwurzacher M, Wagner A, Herzog R, Kratochwill K. Proteome-Wide Differential Effects of Peritoneal Dialysis Fluid Properties in an In Vitro Human Endothelial Cell Model. International Journal of Molecular Sciences. 2022; 23(14):8010. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23148010
Chicago/Turabian StyleSacnun, Juan Manuel, Robin Hoogenboom, Fabian Eibensteiner, Isabel J. Sobieszek, Markus Unterwurzacher, Anja Wagner, Rebecca Herzog, and Klaus Kratochwill. 2022. "Proteome-Wide Differential Effects of Peritoneal Dialysis Fluid Properties in an In Vitro Human Endothelial Cell Model" International Journal of Molecular Sciences 23, no. 14: 8010. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23148010