
Overexpression of AcEXPA23 Promotes Lateral Root Development in Kiwifruit

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Phylogenetic Tree Analysis of the AcExpansin Protein Family
2.2. Physicochemical Properties of AcExpansin Protein Family
2.3. AcExpansin Protein Family Chromosomal Location
2.4. Analysis of Conserved Domains and Gene Structure of the AcExpansin Protein Family
2.5. Intraspecies Collinearity Analysis of Expansion Genes in Kiwifruit
2.6. Screening of Candidate Expansin gene under BR and Brassinazole Treatment
2.7. Subcellular Localisation of AcEXPA23
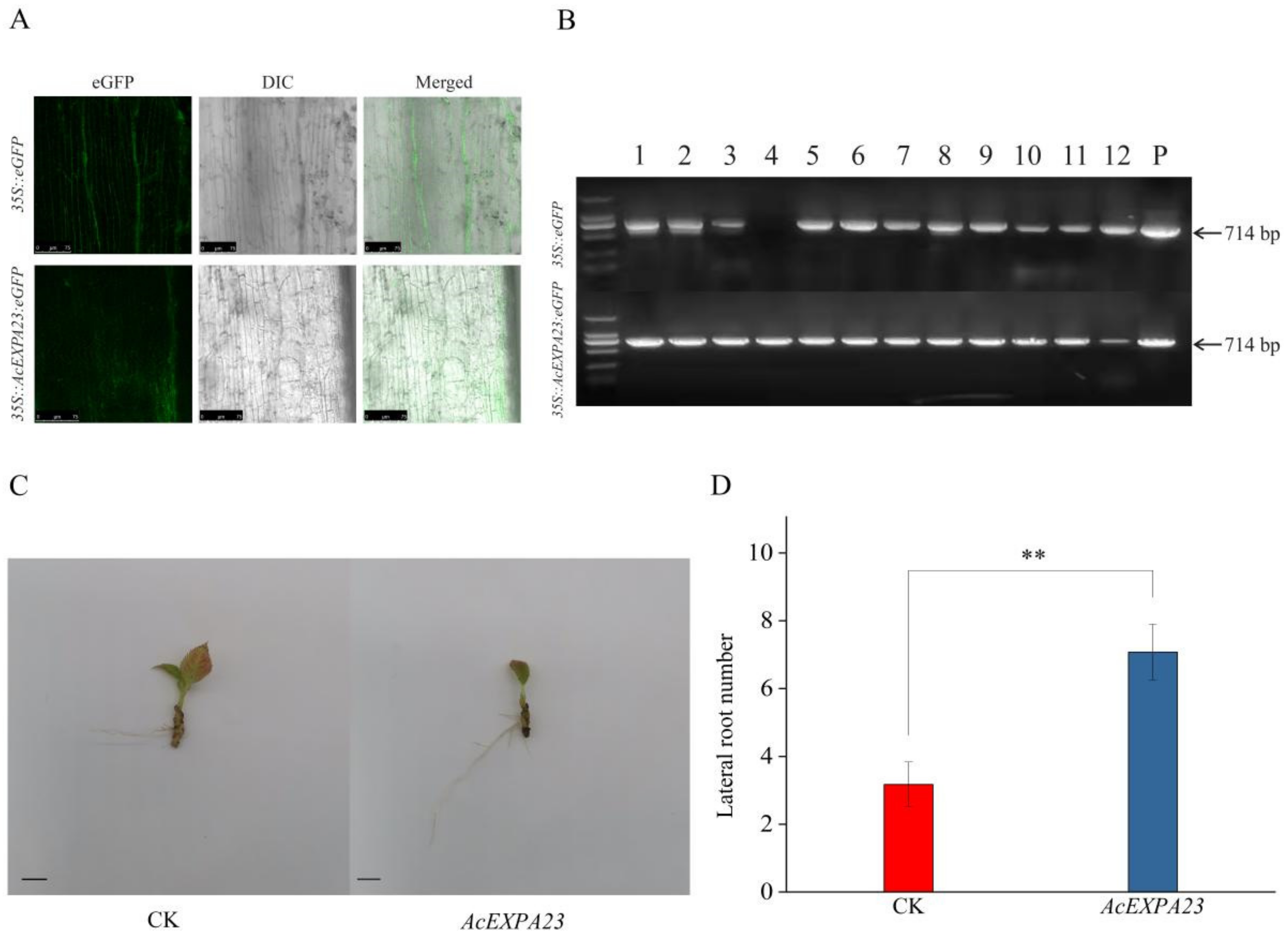
2.8. Transient Overexpression of AcEXPA23 in Kiwifruit
2.9. AcEXPA23 Overexpression in Kiwifruit Enhanced the Number of Lateral Roots
3. Discussion
4. Materials and Methods
4.1. Identification of Kiwifruit Expansin Genes
4.2. Analysis of AcExpansin Protein Family Characteristics
4.3. Transient Expression Hairy Root
4.4. Transformation of A. Chinensis Leaves
4.5. Subcellular Localisation
4.6. Quantitative Real-Time Polymerase Chain Reaction
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fang, J.B.; Zhong, C.H. Fruit scientific research in new China in the past 70 years: Kiwifruit. J. Fruit Sci. 2019, 36, 1352–1359. [Google Scholar]
- Xu, X.B.; Zhang, Q.M. Research and utilizations of germplasm resource of kiwifruit in china. Chin. Bull. Bot. 2003, 20, 648–655. [Google Scholar]
- Iwasawa, H.; Morita, E.; Yui, S.; Yamazaki, M. Anti-oxidant effects of kiwi fruit in vitro and in vivo. Biol. Pharm. Bull. 2011, 34, 128–134. [Google Scholar] [CrossRef] [Green Version]
- He, J.L.; Wu, D.T.; Zhang, Q.; Chen, H.; Li, H.Y.; Han, Q.H.; Lai, X.Y.; Wang, H.; Wu, Y.X.; Yuan, J.G.; et al. Efficacy and mechanism of cinnamon essential oil on inhibition of colletotrichum acutatum isolated from ‘hongyang’ kiwifruit. Front. Microbiol. 2018, 9, 1288. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.J.; Guo, D.D.; Wang, R.; Zhong, Y.P.; Fang, J.B. Development status and suggestions on Chinese kiwifruit industry. J. Fruit Sci. 2020, 37, 754–763. [Google Scholar]
- Chen, J.Y.; Fang, J.B.; Qi, X.J.; Gu, H.; Lin, M.M.; Zhang, W.Y.; Wei, C.G. Research progress on rootstock of kiwifruit. J. Fruit Sci. 2015, 32, 959–968. [Google Scholar]
- Lynch, J. Root architecture and plant productivity. Plant Physiol. 1995, 109, 7–13. [Google Scholar] [CrossRef]
- Malamy, J.E.; Benfey, P.N. Organization and cell differentiation in lateral roots of Arabidopsis thaliana. Development 1997, 124, 33–44. [Google Scholar] [CrossRef]
- Peret, B.; De Rybel, B.; Casimiro, I.; Benkova, E.; Swarup, R.; Laplaze, L.; Beeckman, T.; Bennett, M.J. Arabidopsis lateral root development: An emerging story. Trends Plant Sci. 2009, 14, 399–408. [Google Scholar] [CrossRef]
- De Rybel, B.; Vassileva, V.; Parizot, B.; Demeulenaere, M.; Grunewald, W.; Audenaert, D.; Van Campenhout, J.; Overvoorde, P.; Jansen, L.; Vanneste, S.; et al. A novel AUX/IAA28 signaling cascade activates GATA23-dependent specification of lateral root founder cell identity. Curr. Biol. 2010, 20, 1697–1706. [Google Scholar] [CrossRef]
- Fukaki, H.; Tameda, S.; Masuda, H.; Tasaka, M. Lateral root formation is blocked by a gain-of-function mutation in the SOLITARY-ROOT/IAA14 gene of Arabidopsis. Plant J. 2002, 29, 153–168. [Google Scholar] [CrossRef]
- Fukaki, H.; Nakao, Y.; Okushima, Y.; Theologis, A.; Tasaka, M. Tissue-specific expression of stabilized SOLITARY-ROOT/IAA14 alters lateral root development in Arabidopsis. Plant J. 2005, 44, 382–395. [Google Scholar] [CrossRef]
- Okushima, Y.; Overvoorde, P.J.; Arima, K.; Alonso, J.M.; Chan, A.; Chang, C.; Ecker, J.R.; Hughes, B.; Lui, A.; Nguyen, D.; et al. Functional genomic analysis of the AUXIN RESPONSE FACTOR gene family members in Arabidopsis thaliana: Unique and overlapping functions of ARF7 and ARF19. Plant Cell 2005, 17, 444–463. [Google Scholar] [CrossRef] [Green Version]
- Okushima, Y.; Fukaki, H.; Onoda, M.; Theologis, A.; Tasaka, M. ARF7 and ARF19 regulate lateral root formation via direct activation of LBD/ASL genes in Arabidopsis. Plant Cell 2007, 19, 118–130. [Google Scholar] [CrossRef] [Green Version]
- De Smet, I.; Lau, S.; Voss, U.; Vanneste, S.; Benjamins, R.; Rademacher, E.H.; Schlereth, A.; De Rybel, B.; Vassileva, V.; Grunewald, W.; et al. Bimodular auxin response controls organogenesis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2010, 107, 2705–2710. [Google Scholar] [CrossRef] [Green Version]
- Smet, I.D. Multimodular auxin response controls lateral root development in Arabidopsis. Plant Signal. Behav. 2010, 5, 580–582. [Google Scholar] [CrossRef] [Green Version]
- Goh, T.; Kasahara, H.; Mimura, T.; Kamiya, Y.; Fukaki, H. Multiple AUX/IAA-ARF modules regulate lateral root formation: The role of Arabidopsis SHY2/IAA3-mediated auxin signalling. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1461–1468. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.W.; Kim, M.J.; Kim, N.Y.; Lee, S.H.; Kim, J. LBD18 acts as a transcriptional activator that directly binds to the EXPANSIN14 promoter in promoting lateral root emergence of Arabidopsis. Plant J. 2013, 73, 212–224. [Google Scholar] [CrossRef]
- Lee, D. Genome-wide analysis of the transcriptome downstream of iaa 1 during early auxin response and expression analysis of iaa1-regulated auxin-response genes. J. Exp. Bot. 2009, 60, 3935–3957. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.W.; Kim, N.Y.; Lee, D.J.; Kim, J. LBD18/ASL20 regulates lateral root formation in combination with LBD16/ASL18 downstream of ARF7 and ARF19 in Arabidopsis. Plant Physiol. 2009, 151, 1377–1389. [Google Scholar] [CrossRef] [Green Version]
- McQueen-Mason, S.J.; Cosgrove, D.J. Expansin mode of action on cell walls (analysis of wall hydrolysis, stress relaxation, and binding). Plant Physiol. 1995, 107, 87–100. [Google Scholar] [CrossRef] [Green Version]
- McQueen-Mason, S.; Durachko, D.M.; Cosgrove, D.J. Two endogenous proteins that induce cell wall extension in plants. Plant Cell 1992, 4, 1425–1433. [Google Scholar]
- McQueen-Mason, S.; Cosgrove, D.J. Disruption of hydrogen bonding between plant cell wall polymers by proteins that induce wall extension. Proc. Natl. Acad. Sci. USA 1994, 91, 6574–6578. [Google Scholar] [CrossRef] [Green Version]
- Kende, H.; Bradford, K.; Brummell, D.A.; Cho, H.T.; Cosgrove, D.J.; Fleming, A.J.; Gehring, C.; Lee, Y.; McQueen-Mason, S.; Rose, J.K. Nomenclature for members of the expansin superfamily of genes and proteins. Plant Mol. Biol. 2004, 55, 311–314. [Google Scholar] [CrossRef] [Green Version]
- Che, J.; Yamaji, N.; Shen, R.F.; Ma, J.F. An Al-inducible expansin gene, OsEXPA10 is involved in root cell elongation of rice. Plant J. 2016, 88, 132–142. [Google Scholar] [CrossRef]
- Cho, H.T.; Cosgrove, D.J. Regulation of root hair initiation and expansin gene expression in Arabidopsis. Plant Cell 2002, 14, 3237–3253. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.T.; Kende, H. Expression of expansin genes is correlated with growth in deepwater rice. Plant Cell 1997, 9, 1661–1671. [Google Scholar]
- Goh, H.H.; Sloan, J.; Dorca-Fornell, C.; Fleming, A. Inducible repression of multiple expansin genes leads to growth suppression during leaf development. Plant Physiol. 2012, 159, 1759–1770. [Google Scholar] [CrossRef] [Green Version]
- Kuluev, B.R.; Knyazev, A.V.; Mikhaylova, E.V.; Chemeris, A.V. The role of expansin genes PtrEXPA3 and PnEXPA3 in the regulation of leaf growth in poplar. Russ. J. Genet. 2017, 53, 651–660. [Google Scholar] [CrossRef]
- Saito, T.; Pham Anh, T.; Katsumi-Horigane, A.; Bai, S.; Ito, A.; Sekiyama, Y.; Ono, H.; Moriguchi, T. Development of flower buds in the Japanese pear (Pyrus pyrifolia) from late autumn to early spring. Tree Physiol. 2015, 35, 653–662. [Google Scholar] [CrossRef] [Green Version]
- Civello, P.M.; Powell, A.L.T.; Sabehat, A.; Bennett, A.B. An expansin gene expressed in ripening strawberry fruit. Plant Physiol. 1999, 121, 1273–1279. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Han, Y.; Zhang, M.; Zhou, S.; Kong, X.; Wang, W. Overexpression of the wheat expansin gene TaEXPA2 improved seed production and drought tolerance in transgenic tobacco plants. PLoS ONE 2016, 11, e0153494. [Google Scholar] [CrossRef]
- Wei, P.; Chen, S.; Zhang, X.; Zhao, P.; Xiong, Y.; Wang, W.; Chen, J.; Wang, X. An alpha-expansin, VfEXPA1, is involved in regulation of stomatal movement in Vicia faba L. Chin. Sci. Bull. 2011, 56, 3531–3537. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.T.; Cosgrove, D.J. Altered expression of expansin modulates leaf growth and pedicel abscission in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2000, 97, 9783–9788. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Zhao, J.; Li, X.; Qin, L.; Yan, X.; Liao, H. A soybean β-expansin gene GmEXPB2 intrinsically involved in root system architecture responses to abiotic stresses. Plant J. 2011, 66, 541–552. [Google Scholar] [CrossRef]
- Wu, Y.J.; Thorne, E.T.; Sharp, R.E.; Cosgrove, D.J. Modification of expansin transcript levels in the maize primary root at low water potentials. Plant Physiol. 2001, 126, 1471–1479. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Feng, X.H.; Nakahara, H.; Irshad, M.; Eneji, A.E.; Zheng, Y.; Ping, A. Apical-root apoplastic acidification affects cell wall extensibility in wheat under salinity stress. Physiol. Plant. 2021, 173, 1850–1861. [Google Scholar] [CrossRef]
- Chen, Y.; Han, Y.; Kong, X.; Kang, H.; Ren, Y.; Wang, W. Ectopic expression of wheat expansin gene TaEXPA2 improved the salt tolerance of transgenic tobacco by regulating Na+/K+ and antioxidant competence. Physiol. Plant. 2017, 159, 161–177. [Google Scholar] [CrossRef]
- Wu, Z.; Gu, S.; Gu, H.; Cheng, D.; Li, L.; Guo, X.; Wang, M.; He, S.; Li, M.; Chen, J. Physiological and transcriptomic analyses of brassinosteroid function in kiwifruit root. Environ. Exp. Bot. 2022, 194, 104685. [Google Scholar] [CrossRef]
- Sun, W.; Yu, H.; Liu, M.; Ma, Z.; Chen, H. Evolutionary research on the expansin protein family during the plant transition to land provides new insights into the development of Tartary buckwheat fruit. BMC Genom. 2021, 22, 252. [Google Scholar] [CrossRef]
- Bordoloi, K.S.; Dihingia, P.; Krishnatreya, D.B.; Agarwala, N. Genome-wide identification, characterization and expression analysis of the expansin gene family under drought stress in tea (Camellia sinensis L.). Plant Sci. Today 2021, 8, 32–44. [Google Scholar] [CrossRef]
- Han, Z.; Liu, Y.; Deng, X.; Liu, D.; Liu, Y.; Hu, Y.; Yan, Y. Genome-wide identification and expression analysis of expansin gene family in common wheat (Triticum aestivum L.). BMC Genom. 2019, 20, 101. [Google Scholar] [CrossRef]
- Lee, Y.; Choi, D.; Kende, H. Expansins: Ever-expanding numbers and functions. Curr. Opin. Plant Biol. 2001, 4, 527–532. [Google Scholar] [CrossRef]
- Zhang, W.; Yan, H.; Chen, W.; Liu, J.; Jiang, C.; Jiang, H.; Zhu, S.; Cheng, B. Genome-wide identification and characterization of maize expansin genes expressed in endosperm. Mol. Genet. Genom. 2014, 289, 1061–1074. [Google Scholar] [CrossRef]
- Li, N.; Pu, Y.; Gong, Y.; Yu, Y.; Ding, H. Genomic location and expression analysis of expansin gene family reveals the evolutionary and functional significance in Triticum aestivum. Genes Genom. 2016, 38, 1021–1030. [Google Scholar] [CrossRef]
- Zhu, Y.; Wu, N.; Song, W.; Yin, G.; Qin, Y.; Yan, Y.; Hu, Y. Soybean (Glycine max) expansin gene superfamily origins: Segmental and tandem duplication events followed by divergent selection among subfamilies. BMC Plant Biol. 2014, 14, 93. [Google Scholar] [CrossRef] [Green Version]
- Ding, A.; Marowa, P.; Kong, Y. Genome-wide identification of the expansin gene family in tobacco (Nicotiana tabacum). Mol. Genet. Genom. 2016, 291, 1891–1907. [Google Scholar] [CrossRef]
- Lu, Y.; Liu, L.; Wang, X.; Han, Z.; Ouyang, B.; Zhang, J.; Li, H. Genome-wide identification and expression analysis of the expansin gene family in tomato. Mol. Genet. Genom. 2016, 291, 597–608. [Google Scholar] [CrossRef]
- Zhang, S.; Xu, R.; Gao, Z.; Chen, C.; Jiang, Z.; Shu, H. A genome-wide analysis of the expansin genes in Malus x Domestica. Mol. Genet. Genom. 2014, 289, 225–236. [Google Scholar] [CrossRef]
- Dal Santo, S.; Vannozzi, A.; Tornielli, G.B.; Fasoli, M.; Venturini, L.; Pezzotti, M.; Zenoni, S. Genome-wide analysis of the expansin gene superfamily reveals grapevine-specific structural and functional characteristics. PLoS ONE 2013, 8, e62206. [Google Scholar] [CrossRef]
- Qiao, X.; Li, Q.; Yin, H.; Qi, K.; Li, L.; Wang, R.; Zhang, S.; Paterson, A.H. Gene duplication and evolution in recurring polyploidization-diploidization cycles in plants. Genome Biol. 2019, 20, 38. [Google Scholar] [CrossRef] [Green Version]
- Nan, Q.; Qian, D.; Niu, Y.; He, Y.; Tong, S.; Niu, Z.; Ma, J.; Yang, Y.; An, L.; Wan, D.; et al. Plant actin-depolymerizing factors possess opposing biochemical properties arising from key amino acid changes throughout evolution. Plant Cell 2017, 29, 395–408. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, M.A.; Mateos, I.; Labrador, E.; Dopico, B. Brassinolides and IAA induce the transcription of four a-expansin genes related to development in Cicer arietinum. Plant Physiol. Biochem. 2004, 42, 709–716. [Google Scholar] [CrossRef]
- Park, C.H.; Kim, T.W.; Son, S.H.; Hwang, J.Y.; Lee, S.C.; Chang, S.C.; Kim, S.H.; Kim, S.W.; Kim, S.K. Brassinosteroids control AtEXPA5 gene expression in Arabidopsis thaliana. Phytochemistry 2010, 71, 380–387. [Google Scholar] [CrossRef]
- Bae, J.M.; Kwak, M.S.; Noh, S.A.; Oh, M.J.; Kim, Y.S.; Shin, J.S. Overexpression of sweetpotato expansin cDNA (IbEXP1) increases seed yield in Arabidopsis. Transgenic Res. 2014, 23, 657–667. [Google Scholar] [CrossRef]
- Zhao, F.; Li, G.; Hu, P.; Zhao, X.; Li, L.; Wei, W.; Feng, J.; Zhou, H. Identification of basic/helix-loop-helix transcription factors reveals candidate genes involved in anthocyanin biosynthesis from the strawberry white-flesh mutant. Sci. Rep. 2018, 8, 2721. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.W.; Kim, J. EXPANSINA17 up-regulated by LBD18/ASL20 promotes lateral root formation during the auxin response. Plant Cell Physiol. 2013, 54, 1600–1611. [Google Scholar] [CrossRef] [Green Version]
- Li, A.X.; Han, Y.Y.; Wang, X.; Chen, Y.H.; Zhao, M.R.; Zhou, S.-M.; Wang, W. Root-specific expression of wheat expansin gene TaEXPB23 enhances root growth and water stress tolerance in tobacco. Environ. Exp. Bot. 2015, 110, 73–84. [Google Scholar] [CrossRef]
- Kong, Y.; Wang, B.; Du, H.; Li, W.; Li, X.; Zhang, C. GmEXLB1, a soybean expansin-like B gene, alters root architecture to improve phosphorus acquisition in Arabidopsis. Front. Plant Sci. 2019, 10, 808. [Google Scholar] [CrossRef]
- Yang, Z.; Gao, Z.; Zhou, H.; He, Y.; Liu, Y.; Lai, Y.; Zheng, J.; Li, X.; Liao, H. GmPTF1 modifies root architecture responses to phosphate starvation primarily through regulating GmEXPB2 expression in soybean. Plant J. 2021, 107, 525–543. [Google Scholar] [CrossRef]
- Cortes, A.J.; Blair, M.W. Genotyping by sequencing and genome-environment associations in wild common bean predict widespread divergent adaptation to drought. Front. Plant Sci. 2018, 9, 128. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Islam, A.S.M.F.; Limpot, N.; Mackasmiel, L.; Mierzwa, J.; Cortes, A.J.; Blair, M.W. Genome-wide SNP identification and association mapping for seed mineral concentration in mung bean (Vigna radiata L.). Front. Genet. 2020, 11, 656. [Google Scholar] [CrossRef]
- Buitrago-Bitar, M.A.; Cortes, A.J.; Lopez-Hernandez, F.; Londono-Caicedo, J.M.; Munoz-Florez, J.E.; Munoz, L.C.; Blair, M.W. Allelic diversity at abiotic stress responsive genes in relationship to ecological drought indices for cultivated tepary bean, Phaseolus acutifolius A. gray, and its wild relatives. Genes 2021, 12, 556. [Google Scholar] [CrossRef]
- Rizzo, G.; Monzon, J.P.; Tenorio, F.A.; Howard, R.; Cassman, K.G.; Grassini, P. Climate and agronomy, not genetics, underpin recent maize yield gains in favorable environments. Proc. Natl. Acad. Sci. USA 2022, 119, e2113629119. [Google Scholar] [CrossRef]
- Zou, H.; Wenwen, Y.; Zang, G.; Kang, Z.; Zhang, Z.; Huang, J.; Wang, G. OsEXPB2, a β-expansin gene, is involved in rice root system architecture. Mol. Breed. 2015, 35, 1–14. [Google Scholar] [CrossRef]
- Liu, J.; Peng, H.; Cui, J.; Huang, W.; Kong, L.; Clarke, J.L.; Jian, H.; Wang, G.L.; Peng, D. Molecular characterization of a novel effector expansin-like protein from heterodera avenae that induces cell death in Nicotiana benthamiana. Sci. Rep. 2016, 6, 35677. [Google Scholar] [CrossRef] [Green Version]
- Ren, H.; Wen, L.Z.; Guo, Y.H.; Yu, Y.Y.; Sun, C.H.; Fan, H.M.; Ma, F.F.; Zheng, C.S. Expressional and functional verification of the involvement of CmEXPA4 in chrysanthemum root development. J. Plant Growth Regul. 2019, 38, 1375–1386. [Google Scholar] [CrossRef]
- He, X.; Zeng, J.; Cao, F.; Ahmed, I.M.; Zhang, G.; Vincze, E.; Wu, F. HvEXPB7, a novel β-expansin gene revealed by the root hair transcriptome of Tibetan wild barley, improves root hair growth under drought stress. J. Exp. Bot. 2015, 66, 7405–7419. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Feng, X.; Chen, Z.H.; Zhang, G.; Wu, F. Transient silencing of an expansin HvEXPA1 inhibits root cell elongation and reduces Al accumulation in root cell wall of Tibetan wild barley. Environ. Exp. Bot. 2019, 165, 120–128. [Google Scholar] [CrossRef]
- Li, J.; Liu, Z.; Gao, C.; Miao, Y.; Cui, K. Overexpression of DsEXLA2 gene from Dendrocalamus sinicus accelerates the plant growth rate of Arabidopsis. Phytochemistry 2022, 199, 113178. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11 molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Wang, T.; Atkinson, R.; Janssen, B. The choice of agrobacterium strain for transformation of kiwifruit. Acta Hortic. 2007, 753, 227–232. [Google Scholar] [CrossRef]
- Wang, N.; Zhang, W.; Qin, M.; Li, S.; Qiao, M.; Liu, Z.; Xiang, F. Drought tolerance conferred in soybean (Glycine max. L) by GmMYB84, a novel R2R3-MYB transcription factor. Plant Cell Physiol. 2017, 58, 1764–1776. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Ma, F.; Liang, D.; Li, J.; Wang, Y. Ascorbate biosynthesis during early fruit development is the main reason for its accumulation in kiwi. PLoS ONE 2010, 5, e14281. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subfamily | Gene Sequence Number | Gene Name | Number of Amino Acids | Molecular Weight | Theoretical pI | Instability Index | Aliphatic Index | GRAVY |
|---|---|---|---|---|---|---|---|---|
| AcEXPA | Actinidia00998 | AcEXPA1 | 249 | 26,626.76 | 7.53 | 26.28 | 67.03 | −0.116 |
| Actinidia02062 | AcEXPA2 | 344 | 37,499.78 | 9.31 | 33.13 | 66.40 | −0.074 | |
| Actinidia02869 | AcEXPA3 | 210 | 22,935.07 | 9.20 | 24.52 | 57.67 | −0.136 | |
| Actinidia04838 | AcEXPA4 | 228 | 25,123.66 | 8.59 | 24.98 | 73.90 | −0.154 | |
| Actinidia04880 | AcEXPA5 | 247 | 26,267.65 | 9.23 | 32.29 | 71.09 | −0.023 | |
| Actinidia11513 | AcEXPA6 | 236 | 26,088.61 | 8.12 | 22.97 | 81.48 | −0.078 | |
| Actinidia12294 | AcEXPA7 | 210 | 22,278.80 | 9.72 | 31.12 | 67.81 | −0.100 | |
| Actinidia12816 | AcEXPA8 | 247 | 27,400.70 | 10.16 | 40.00 | 63.20 | −0.431 | |
| Actinidia13883 | AcEXPA9 | 259 | 27,589.34 | 8.94 | 41.09 | 70.42 | −0.060 | |
| Actinidia19034 | AcEXPA10 | 249 | 26,683.88 | 8.88 | 34.04 | 64.22 | −0.157 | |
| Actinidia19880 | AcEXPA11 | 321 | 34,930.65 | 9.31 | 38.12 | 75.67 | 0.029 | |
| Actinidia20584 | AcEXPA12 | 255 | 27,582.32 | 8.01 | 29.89 | 73.49 | 0.014 | |
| Actinidia23935 | AcEXPA13 | 259 | 28,024.18 | 9.74 | 35.36 | 76.45 | 0.026 | |
| Actinidia26471 | AcEXPA14 | 259 | 28,122.98 | 9.47 | 36.04 | 67.88 | −0.066 | |
| Actinidia28781 | AcEXPA15 | 358 | 38,953.08 | 9.45 | 46.03 | 65.98 | −0.375 | |
| Actinidia29037 | AcEXPA16 | 255 | 27,616.01 | 8.89 | 27.10 | 64.27 | −0.101 | |
| Actinidia29231 | AcEXPA17 | 258 | 27,701.47 | 9.47 | 38.78 | 69.92 | −0.006 | |
| Actinidia29762 | AcEXPA18 | 255 | 27,408.12 | 8.96 | 26.64 | 70.78 | −0.008 | |
| Actinidia31623 | AcEXPA19 | 193 | 20,731.32 | 9.30 | 30.18 | 64.20 | −0.202 | |
| Actinidia32475 | AcEXPA20 | 281 | 31,079.42 | 9.40 | 32.99 | 68.01 | −0.441 | |
| Actinidia33207 | AcEXPA21 | 250 | 26,443.57 | 8.98 | 33.57 | 65.56 | −0.096 | |
| Actinidia35005 | AcEXPA22 | 248 | 26,471.56 | 8.38 | 36.38 | 61.73 | −0.157 | |
| Actinidia35709 | AcEXPA23 | 222 | 23,767.37 | 8.69 | 35.10 | 83.11 | 0.131 | |
| Actinidia37198 | AcEXPA24 | 259 | 27,821.58 | 9.39 | 44.33 | 70.85 | −0.008 | |
| Actinidia37944 | AcEXPA25 | 255 | 27,415.04 | 9.06 | 26.35 | 65.41 | −0.074 | |
| Actinidia38621 | AcEXPA26 | 247 | 26,284.66 | 8.76 | 28.93 | 70.32 | 0.029 | |
| Actinidia38761 | AcEXPA27 | 240 | 25,567.64 | 8.64 | 38.18 | 67.08 | −0.049 | |
| Actinidia38779 | AcEXPA28 | 240 | 25,567.64 | 8.64 | 38.18 | 67.08 | −0.049 | |
| AcEXPB | Actinidia09861 | AcEXPB1 | 266 | 28,632.73 | 8.79 | 37.55 | 77.74 | −0.021 |
| Actinidia11045 | AcEXPB2 | 266 | 28,861.98 | 8.86 | 38.09 | 75.53 | −0.064 | |
| Actinidia13599 | AcEXPB3 | 197 | 21,192.05 | 8.97 | 37.32 | 72.79 | −0.307 | |
| Actinidia22767 | AcEXPB4 | 231 | 23,757.54 | 6.21 | 32.77 | 57.92 | −0.082 | |
| Actinidia33485 | AcEXPB5 | 248 | 25,863.81 | 4.82 | 35.78 | 67.62 | −0.119 | |
| Actinidia39310 | AcEXPB6 | 265 | 28,448.37 | 8.46 | 33.87 | 76.87 | −0.070 | |
| AcEXLA | Actinidia18015 | AcEXLA1 | 457 | 50,895.06 | 7.04 | 32.07 | 81.93 | −0.082 |
| Actinidia20552 | AcEXLA2 | 259 | 28,282.42 | 8.81 | 25.72 | 81.70 | 0.026 | |
| Actinidia24612 | AcEXLA3 | 783 | 86,023.25 | 6.88 | 43.40 | 82.95 | −0.258 | |
| AcEXLB | Actinidia10878 | AcEXLB1 | 256 | 27,956.06 | 4.73 | 32.56 | 72.34 | −0.200 |
| Actinidia25907 | AcEXLB2 | 236 | 25,757.25 | 7.52 | 30.21 | 79.28 | −0.072 | |
| Actinidia25941 | AcEXLB3 | 254 | 27,840.00 | 4.81 | 30.87 | 67.56 | −0.254 | |
| Actinidia31474 | AcEXLB4 | 256 | 28,035.36 | 5.13 | 36.77 | 71.99 | −0.186 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Z.; Li, M.; Zhong, Y.; Li, L.; Cheng, D.; Gu, H.; Guo, X.; Qi, X.; Chen, J. Overexpression of AcEXPA23 Promotes Lateral Root Development in Kiwifruit. Int. J. Mol. Sci. 2022, 23, 8026. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23148026
Wu Z, Li M, Zhong Y, Li L, Cheng D, Gu H, Guo X, Qi X, Chen J. Overexpression of AcEXPA23 Promotes Lateral Root Development in Kiwifruit. International Journal of Molecular Sciences. 2022; 23(14):8026. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23148026
Chicago/Turabian StyleWu, Zhiyong, Ming Li, Yunpeng Zhong, Lan Li, Dawei Cheng, Hong Gu, Xizhi Guo, Xiujuan Qi, and Jinyong Chen. 2022. "Overexpression of AcEXPA23 Promotes Lateral Root Development in Kiwifruit" International Journal of Molecular Sciences 23, no. 14: 8026. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23148026