
Aquatic Freshwater Vertebrate Models of Epilepsy Pathology: Past Discoveries and Future Directions for Therapeutic Discovery


{kind=link}
{kind=link}
Abstract
:1. Introduction
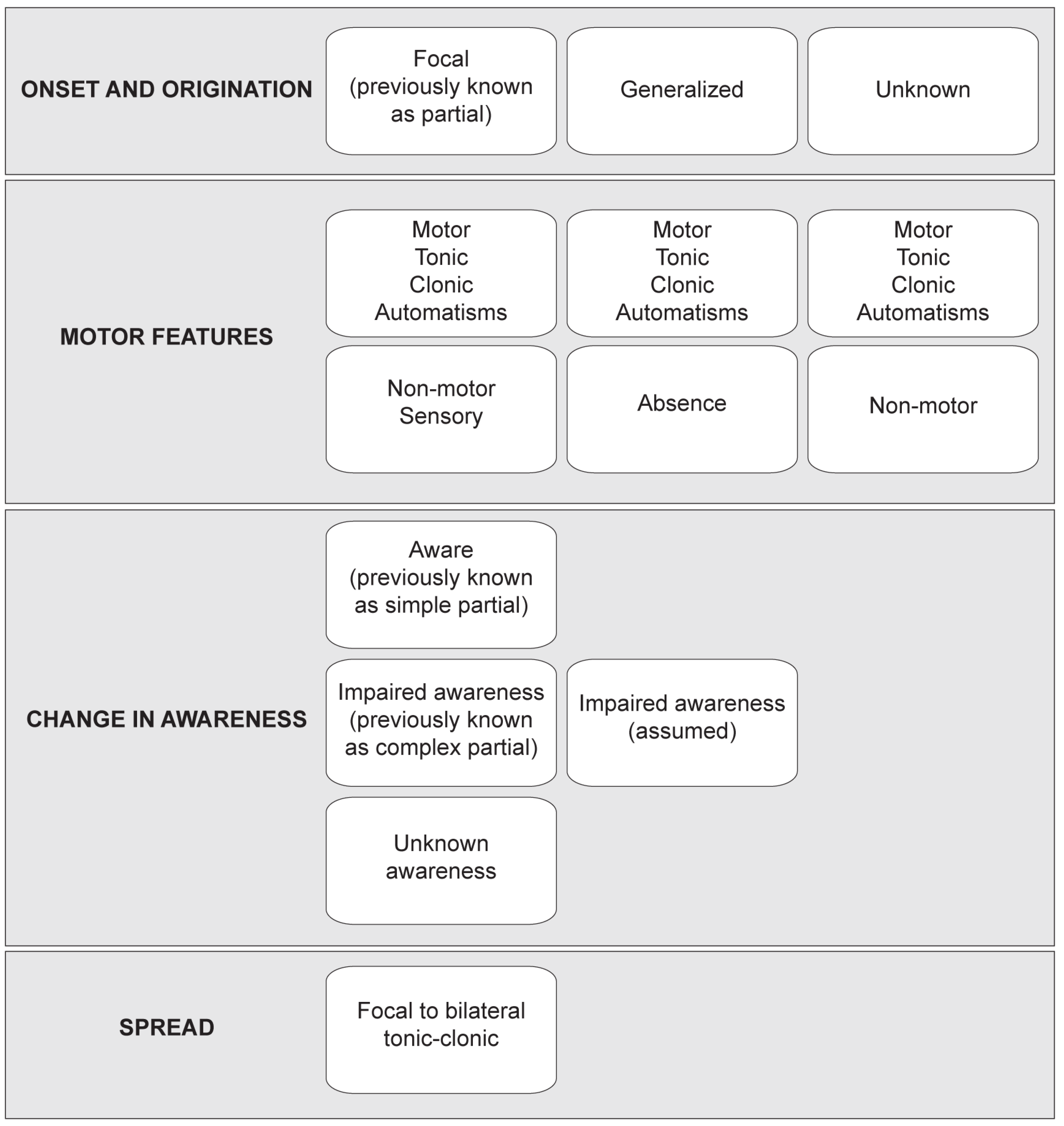
Classification and Etiology of Seizures
2. Aquatic Freshwater Vertebrate Animal Model Advantages
2.1. Aquatic Freshwater Vertebrates as Seizure Models
2.2. Advantages of Aquatic Freshwater Vertebrates as Seizure Models
2.3. Disadvantages of Aquatic Freshwater Vertebrates as Seizure Models
3. Current Aquatic Vertebrate Seizure Models
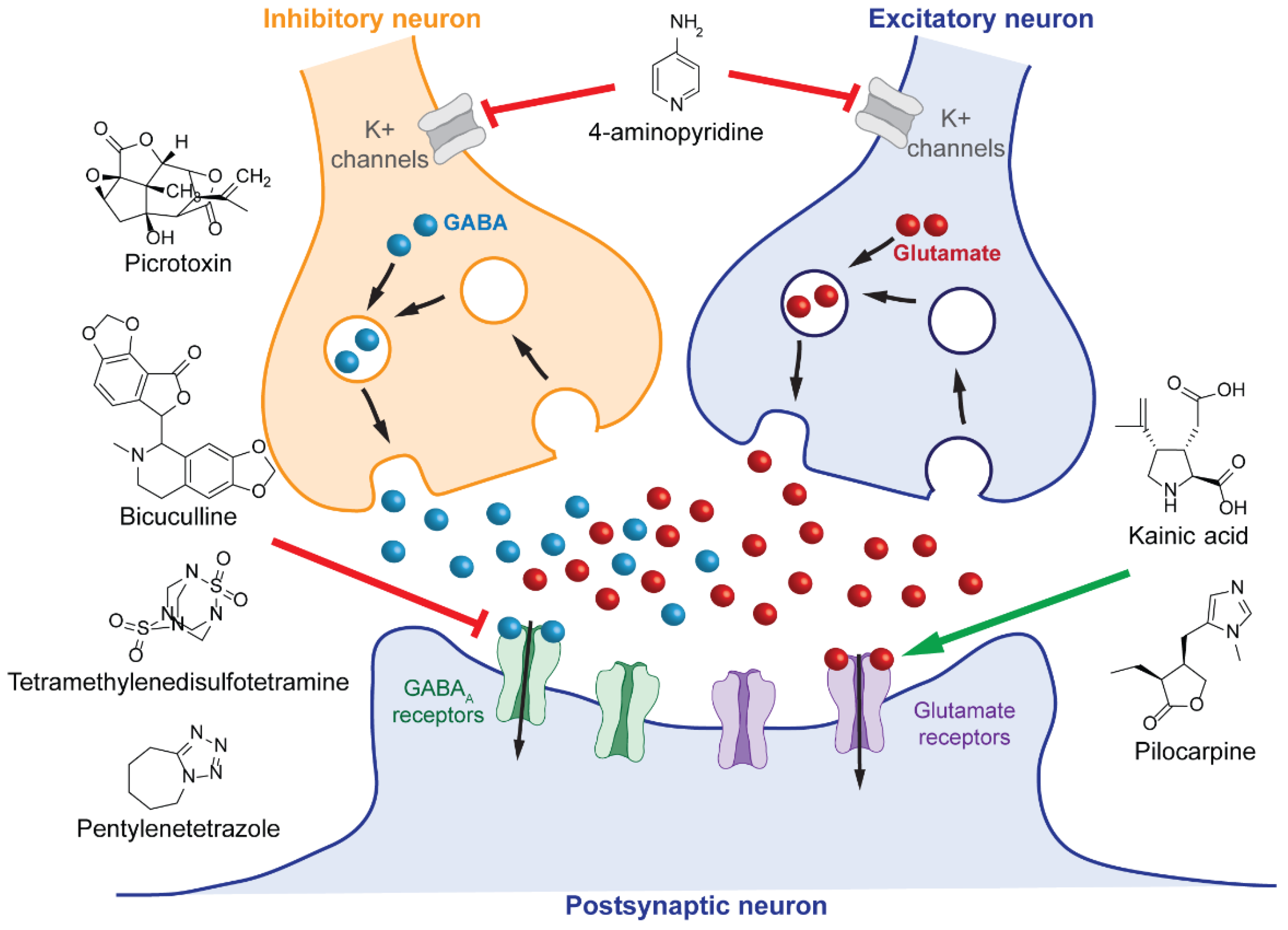
3.1. Chemical Induction of Seizures
3.2. Genetic Induction of Seizures
3.3. New but Less Established Aquatic Vertebrate Seizure Models
4. Looking to the Future for Aquatic Seizure Models
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Epilepsy. 2022. Available online: https://www.who.int/health-topics/epilepsy#tab=tab_1 (accessed on 27 April 2022).
- Moshe, S.L.; Perucca, E.; Ryvlin, P.; Tomson, T. Epilepsy: New advances. Lancet 2015, 385, 884–898. [Google Scholar] [CrossRef]
- Go, C.; Snead, O.C., 3rd. Pharmacologically intractable epilepsy in children: Diagnosis and preoperative evaluation. Neurosurg Focus 2008, 25, E2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annegers, J.F.; Hauser, W.A.; Elveback, L.R. Remission of seizures and relapse in patients with epilepsy. Epilepsia 1979, 20, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Cockerell, O.C.; Johnson, A.L.; Sander, J.W.; Hart, Y.M.; Shorvon, S.D. Remission of epilepsy: Results from the National General Practice Study of Epilepsy. Lancet 1995, 346, 140–144. [Google Scholar] [CrossRef]
- Kwan, P.; Arzimanoglou, A.; Berg, A.T.; Brodie, M.J.; Allen Hauser, W.; Mathern, G.; Moshe, S.L.; Perucca, E.; Wiebe, S.; French, J. Definition of drug resistant epilepsy: Consensus proposal by the ad hoc Task Force of the ILAE Commission on Therapeutic Strategies. Epilepsia 2010, 51, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Kwan, P.; Brodie, M.J. Early identification of refractory epilepsy. N. Engl. J. Med. 2000, 342, 314–319. [Google Scholar] [CrossRef]
- Fisher, R.S.; Cross, J.H.; French, J.A.; Higurashi, N.; Hirsch, E.; Jansen, F.E.; Lagae, L.; Moshé, S.L.; Peltola, J.; Roulet Perez, E.; et al. Operational classification of seizure types by the International League Against Epilepsy: Position Paper of the ILAE Commission for Classification and Terminology. Epilepsia 2017, 58, 522–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanner, A.M.; Ashman, E.; Gloss, D.; Harden, C.; Bourgeois, B.; Bautista, J.F.; Abou-Khalil, B.; Burakgazi-Dalkilic, E.; Llanas Park, E.; Stern, J.; et al. Practice guideline update summary: Efficacy and tolerability of the new antiepileptic drugs I: Treatment of new-onset epilepsy: Report of the Guideline Development, Dissemination, and Implementation Subcommittee of the American Academy of Neurology and the American Epilepsy Society. Neurology 2018, 91, 74–81. [Google Scholar] [CrossRef] [Green Version]
- Kanner, A.M.; Ashman, E.; Gloss, D.; Harden, C.; Bourgeois, B.; Bautista, J.F.; Abou-Khalil, B.; Burakgazi-Dalkilic, E.; Park, E.L.; Stern, J.; et al. Practice guideline update summary: Efficacy and tolerability of the new antiepileptic drugs I: Treatment of new-onset epilepsy: Report of the American Epilepsy Society and the Guideline Development, Dissemination, and Implementation Subcommittee of the American Academy of Neurology. Epilepsy Curr. 2018, 18, 260–268. [Google Scholar] [CrossRef] [PubMed]
- French, J.A.; Perucca, E. Time to Start Calling Things by Their Own Names? The Case for Antiseizure Medicines. Epilepsy Curr. 2020, 20, 69–72. [Google Scholar] [CrossRef] [Green Version]
- Hakami, T. Efficacy and tolerability of antiseizure drugs. Ther. Adv. Neurol. Disord. 2021, 14, 17562864211037430. [Google Scholar] [CrossRef] [PubMed]
- Löscher, W.; Schmidt, D. Modern antiepileptic drug development has failed to deliver: Ways out of the current dilemma. Epilepsia 2011, 52, 657–678. [Google Scholar] [CrossRef] [PubMed]
- Shellhaas, R.A. Seizure classification, etiology, and management. Handb. Clin. Neurol. 2019, 162, 347–361. [Google Scholar] [CrossRef]
- Maljevic, S.; Vejzovic, S.; Bernhard, M.K.; Bertsche, A.; Weise, S.; Döcker, M.; Lerche, H.; Lemke, J.R.; Merkenschlager, A.; Syrbe, S. Novel KCNQ3 Mutation in a Large Family with Benign Familial Neonatal Epilepsy: A Rare Cause of Neonatal Seizures. Mol. Syndromol. 2016, 7, 189–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piro, E.; Nardello, R.; Gennaro, E.; Fontana, A.; Taglialatela, M.; Mangano, G.D.; Corsello, G.; Mangano, S. A novel mutation in KCNQ3-related benign familial neonatal epilepsy: Electroclinical features and neurodevelopmental outcome. Epileptic Disord. Int. Epilepsy J. Videotape 2019, 21, 87–91. [Google Scholar] [CrossRef]
- Li, H.; Li, N.; Shen, L.; Jiang, H.; Yang, Q.; Song, Y.; Guo, J.; Xia, K.; Pan, Q.; Tang, B. A novel mutation of KCNQ3 gene in a Chinese family with benign familial neonatal convulsions. Epilepsy Res. 2008, 79, 1–5. [Google Scholar] [CrossRef]
- Miceli, F.; Striano, P.; Soldovieri, M.V.; Fontana, A.; Nardello, R.; Robbiano, A.; Bellini, G.; Elia, M.; Zara, F.; Taglialatela, M.; et al. A novel KCNQ3 mutation in familial epilepsy with focal seizures and intellectual disability. Epilepsia 2015, 56, e15–e20. [Google Scholar] [CrossRef]
- Nardello, R.; Mangano, G.D.; Miceli, F.; Fontana, A.; Piro, E.; Salpietro, V. Benign familial infantile epilepsy associated with KCNQ3 mutation: A rare occurrence or an underestimated event? Epileptic Disord. Int. Epilepsy J. Videotape 2020, 22, 807–810. [Google Scholar] [CrossRef] [PubMed]
- Guerrini, R.; Barba, C. Focal cortical dysplasia: An update on diagnosis and treatment. Expert Rev. Neurother. 2021, 21, 1213–1224. [Google Scholar] [CrossRef]
- Koenig, M.; Dobyns, W.B.; Di Donato, N. Lissencephaly: Update on diagnostics and clinical management. Eur. J. Paediatr. Neurol. EJPN Off. J. Eur. Paediatr. Neurol. Soc. 2021, 35, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Hundallah, K.; Tabarki, B. Treatable inherited metabolic epilepsies. Neurosciences 2021, 26, 229–235. [Google Scholar] [CrossRef]
- Pearl, P.L.; Bennett, H.D.; Khademian, Z. Seizures and metabolic disease. Curr. Neurol. Neurosci. Rep. 2005, 5, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Choi, T.Y.; Choi, T.I.; Lee, Y.R.; Choe, S.K.; Kim, C.H. Zebrafish as an animal model for biomedical research. Exp. Mol. Med. 2021, 53, 310–317. [Google Scholar] [CrossRef]
- Ishibashi, S.; Saldanha, F.Y.L.; Amaya, E. Chapter 14—Xenopus as a Model Organism for Biomedical Research. In Basic Science Methods for Clinical Researchers; Jalali, M., Saldanha, F.Y.L., Jalali, M., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 263–290. [Google Scholar]
- Kwan, K.M.; Fujimoto, E.; Grabher, C.; Mangum, B.D.; Hardy, M.E.; Campbell, D.S.; Parant, J.M.; Yost, H.J.; Kanki, J.P.; Chien, C.B. The Tol2kit: A multisite gateway-based construction kit for Tol2 transposon transgenesis constructs. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2007, 236, 3088–3099. [Google Scholar] [CrossRef] [PubMed]
- Horb, M.; Wlizla, M.; Abu-Daya, A.; McNamara, S.; Gajdasik, D.; Igawa, T.; Suzuki, A.; Ogino, H.; Noble, A.; Centre de Ressource Biologique Xenope Team in France; et al. Xenopus Resources: Transgenic, Inbred and Mutant Animals, Training Opportunities, and Web-Based Support. Front. Physiol. 2019, 10, 387. [Google Scholar] [CrossRef]
- Thessen, A.E.; Marvel, S.; Achenbach, J.C.; Fischer, S.; Haendel, M.A.; Hayward, K.; Klüver, N.; Könemann, S.; Legradi, J.; Lein, P.; et al. Implementation of Zebrafish Ontologies for Toxicology Screening. Front. Toxicol. 2022, 4, 817999. [Google Scholar] [CrossRef]
- Padilla, S.; Corum, D.; Padnos, B.; Hunter, D.L.; Beam, A.; Houck, K.A.; Sipes, N.; Kleinstreuer, N.; Knudsen, T.; Dix, D.J.; et al. Zebrafish developmental screening of the ToxCast™ Phase I chemical library. Reprod. Toxicol. 2012, 33, 174–187. [Google Scholar] [CrossRef]
- Truong, L.; Reif, D.M.; St Mary, L.; Geier, M.C.; Truong, H.D.; Tanguay, R.L. Multidimensional in vivo hazard assessment using zebrafish. Toxicol. Sci. Off. J. Soc. Toxicol. 2014, 137, 212–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tice, R.R.; Austin, C.P.; Kavlock, R.J.; Bucher, J.R. Improving the human hazard characterization of chemicals: A Tox21 update. Environ. Health Perspect. 2013, 121, 756–765. [Google Scholar] [CrossRef] [Green Version]
- Mannioui, A.; Vauzanges, Q.; Fini, J.B.; Henriet, E.; Sekizar, S.; Azoyan, L.; Thomas, J.L.; Pasquier, D.D.; Giovannangeli, C.; Demeneix, B.; et al. The Xenopus tadpole: An in vivo model to screen drugs favoring remyelination. Mult. Scler. 2018, 24, 1421–1432. [Google Scholar] [CrossRef] [Green Version]
- Mughal, B.B.; Demeneix, B.A.; Fini, J.B. Evaluating Thyroid Disrupting Chemicals In Vivo Using Xenopus laevis. Methods Mol. Biol. 2018, 1801, 183–192. [Google Scholar] [CrossRef]
- Sullivan, K.G.; Levin, M. Inverse Drug Screening of Bioelectric Signaling and Neurotransmitter Roles: Illustrated Using a Xenopus Tail Regeneration Assay. Cold Spring Harb. Protoc. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Patton, E.E.; Zon, L.I.; Langenau, D.M. Zebrafish disease models in drug discovery: From preclinical modelling to clinical trials. Nat. Rev. Drug Discov. 2021, 20, 611–628. [Google Scholar] [CrossRef]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellsten, U.; Harland, R.M.; Gilchrist, M.J.; Hendrix, D.; Jurka, J.; Kapitonov, V.; Ovcharenko, I.; Putnam, N.H.; Shu, S.; Taher, L.; et al. The genome of the Western clawed frog Xenopus tropicalis. Science 2010, 328, 633–636. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Jiao, Y.; Pan, F.; Guan, Z.; Cheng, S.H.; Sun, D. Knock-in of a Large Reporter Gene via the High-Throughput Microinjection of the CRISPR/Cas9 System. IEEE Trans. Bio-Med. Eng. 2022. [Google Scholar] [CrossRef] [PubMed]
- Colijn, S.; Yin, Y.; Stratman, A.N. High-throughput methodology to identify CRISPR-generated Danio rerio mutants using fragment analysis with unmodified PCR products. Dev. Biol. 2022, 484, 22–29. [Google Scholar] [CrossRef]
- Guo, S.; Gao, G.; Zhang, C.; Peng, G. Multiplexed Genome Editing for Efficient Phenotypic Screening in Zebrafish. Vet. Sci. 2022, 9, 92. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Zhang, Y.; Zhou, Y.; Zhang, B.; Krueger, C.J.; Bi, X.; Zhu, Z.; Tong, X.; Zhang, B. ErCas12a and T5exo-ErCas12a Mediate Simple and Efficient Genome Editing in Zebrafish. Biology 2022, 11, 411. [Google Scholar] [CrossRef]
- Hernandez-Huertas, L.; Kushawah, G.; Diaz-Moscoso, A.; Tomas-Gallardo, L.; Moreno-Sanchez, I.; da Silva Pescador, G.; Bazzini, A.A.; Moreno-Mateos, M.A. Optimized CRISPR-RfxCas13d system for RNA targeting in zebrafish embryos. STAR Protoc. 2022, 3, 101058. [Google Scholar] [CrossRef]
- Thumberger, T.; Tavhelidse-Suck, T.; Gutierrez-Triana, J.A.; Cornean, A.; Medert, R.; Welz, B.; Freichel, M.; Wittbrodt, J. Boosting targeted genome editing using the hei-tag. eLife 2022, 11, e70558. [Google Scholar] [CrossRef]
- Blitz, I.L.; Nakayama, T. CRISPR-Cas9 Mutagenesis in Xenopus tropicalis for Phenotypic Analyses in the F(0) Generation and Beyond. Cold Spring Harb. Protoc. 2022, 2022. [Google Scholar] [CrossRef]
- Corkins, M.E.; DeLay, B.D.; Miller, R.K. Tissue-Targeted CRISPR-Cas9-Mediated Genome Editing of Multiple Homeologs in F(0)-Generation Xenopus laevis Embryos. Cold Spring Harb. Protoc. 2022, 2022. [Google Scholar] [CrossRef]
- Godden, A.M.; Antonaci, M.; Ward, N.J.; van der Lee, M.; Abu-Daya, A.; Guille, M.; Wheeler, G.N. An efficient miRNA knockout approach using CRISPR-Cas9 in Xenopus. Dev. Biol. 2022, 483, 66–75. [Google Scholar] [CrossRef]
- Parain, K.; Lourdel, S.; Donval, A.; Chesneau, A.; Borday, C.; Bronchain, O.; Locker, M.; Perron, M. CRISPR/Cas9-Mediated Models of Retinitis Pigmentosa Reveal Differential Proliferative Response of Müller Cells between Xenopus laevis and Xenopus tropicalis. Cells 2022, 11, 807. [Google Scholar] [CrossRef] [PubMed]
- Tanouchi, M.; Igawa, T.; Suzuki, N.; Suzuki, M.; Hossain, N.; Ochi, H.; Ogino, H. Optimization of CRISPR/Cas9-mediated gene disruption in Xenopus laevis using a phenotypic image analysis technique. Dev. Growth Differ. 2022. [Google Scholar] [CrossRef]
- Hamm, J.T.; Ceger, P.; Allen, D.; Stout, M.; Maull, E.A.; Baker, G.; Zmarowski, A.; Padilla, S.; Perkins, E.; Planchart, A.; et al. Characterizing sources of variability in zebrafish embryo screening protocols. Altex 2019, 36, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Alzualde, A.; Behl, M.; Sipes, N.S.; Hsieh, J.H.; Alday, A.; Tice, R.R.; Paules, R.S.; Muriana, A.; Quevedo, C. Toxicity profiling of flame retardants in zebrafish embryos using a battery of assays for developmental toxicity, neurotoxicity, cardiotoxicity and hepatotoxicity toward human relevance. Neurotoxicol. Teratol. 2018, 70, 40–50. [Google Scholar] [CrossRef]
- O’Connell, D.; Mruk, K.; Rocheleau, J.M.; Kobertz, W.R. Xenopus laevis oocytes infected with multi-drug-resistant bacteria: Implications for electrical recordings. J. Gen. Physiol. 2011, 138, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Gundersen, C.B.; Miledi, R.; Parker, I. Serotonin receptors induced by exogenous messenger RNA in Xenopus oocytes. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1983, 219, 103–109. [Google Scholar] [CrossRef]
- Gundersen, C.B.; Miledi, R.; Parker, I. Glutamate and kainate receptors induced by rat brain messenger RNA in Xenopus oocytes. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1984, 221, 127–143. [Google Scholar] [CrossRef]
- Houamed, K.M.; Bilbe, G.; Smart, T.G.; Constanti, A.; Brown, D.A.; Barnard, E.A.; Richards, B.M. Expression of functional GABA, glycine and glutamate receptors in Xenopus oocytes injected with rat brain mRNA. Nature 1984, 310, 318–321. [Google Scholar] [CrossRef] [PubMed]
- Sumikawa, K.; Parker, I.; Miledi, R. Partial purification and functional expression of brain mRNAs coding for neurotransmitter receptors and voltage-operated channels. Proc. Natl. Acad. Sci. USA 1984, 81, 7994–7998. [Google Scholar] [CrossRef] [Green Version]
- Sumikawa, K.; Parker, I.; Miledi, R. Messenger RNA from rat brain induces noradrenaline and dopamine receptors in Xenopus oocytes. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1984, 223, 255–260. [Google Scholar] [CrossRef]
- Jardemark, K.; Nystrom, B.; Rydenhag, B.; Hamberger, A.; Jacobson, I. Expression of Ca(2+)-ion permeable alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionate (AMPA) receptors in Xenopus oocytes injected with total RNA from human epileptic temporal lobe. Neurosci. Lett. 1995, 194, 93–96. [Google Scholar] [CrossRef]
- Spauschus, A.; Eunson, L.; Hanna, M.G.; Kullmann, D.M. Functional characterization of a novel mutation in KCNA1 in episodic ataxia type 1 associated with epilepsy. Ann. N. Y. Acad. Sci. 1999, 868, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.P.; Levesque, P.C.; Little, W.A.; Conder, M.L.; Ramakrishnan, P.; Neubauer, M.G.; Blanar, M.A. Functional expression of two KvLQT1-related potassium channels responsible for an inherited idiopathic epilepsy. J. Biol. Chem. 1998, 273, 19419–19423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madeja, M.; Stocker, M.; Musshoff, U.; Pongs, O.; Speckmann, E.J. Potassium currents in epilepsy: Effects of the epileptogenic agent pentylenetetrazol on a cloned potassium channel. Brain Res. 1994, 656, 287–294. [Google Scholar] [CrossRef]
- Musshoff, U.; Madeja, M.; Bloms-Funke, P.; Speckmann, E.J. Effects of the epileptogenic agent bicuculline methiodide on membrane currents induced by N-methyl-D-aspartate and kainate (oocyte; Xenopus laevis). Brain Res. 1994, 639, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Airaksinen, M.S.; Panula, P. Comparative neuroanatomy of the histaminergic system in the brain of the frog Xenopus laevis. J. Comp. Neurol. 1990, 292, 412–423. [Google Scholar] [CrossRef]
- Gupta, T.; Marquart, G.D.; Horstick, E.J.; Tabor, K.M.; Pajevic, S.; Burgess, H.A. Morphometric analysis and neuroanatomical mapping of the zebrafish brain. Methods 2018, 150, 49–62. [Google Scholar] [CrossRef]
- Herget, U.; Wolf, A.; Wullimann, M.F.; Ryu, S. Molecular neuroanatomy and chemoarchitecture of the neurosecretory preoptic-hypothalamic area in zebrafish larvae. J. Comp. Neurol. 2014, 522, 1542–1564. [Google Scholar] [CrossRef]
- Kunst, M.; Laurell, E.; Mokayes, N.; Kramer, A.; Kubo, F.; Fernandes, A.M.; Förster, D.; Dal Maschio, M.; Baier, H. A Cellular-Resolution Atlas of the Larval Zebrafish Brain. Neuron 2019, 103, 21–38 e25. [Google Scholar] [CrossRef] [PubMed]
- Panula, P.; Chen, Y.C.; Priyadarshini, M.; Kudo, H.; Semenova, S.; Sundvik, M.; Sallinen, V. The comparative neuroanatomy and neurochemistry of zebrafish CNS systems of relevance to human neuropsychiatric diseases. Neurobiol. Dis. 2010, 40, 46–57. [Google Scholar] [CrossRef]
- Ridd, K. Neuroanatomy: From fin to forelimb. Nature 2010, 466, 701. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.; Alford, S.T. Descending projections and excitation during fictive swimming in Xenopus embryos: Neuroanatomy and lesion experiments. J. Comp. Neurol. 1986, 250, 253–261. [Google Scholar] [CrossRef]
- Roberts, A.; Clarke, J.D. The neuroanatomy of an amphibian embryo spinal cord. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1982, 296, 195–212. [Google Scholar] [CrossRef]
- Zhu, X.Y.; Guo, S.Y.; Xia, B.; Li, C.Q.; Wang, L.; Wang, Y.H. Development of zebrafish demyelination model for evaluation of remyelination compounds and RORgammat inhibitors. J. Pharmacol. Toxicol. Methods 2019, 98, 106585. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Lei, X.; Li, X.; Chen, Y.; Xu, F.; Feng, X.; Wei, S.; Li, Y. A novel model of demyelination and remyelination in a GFP-transgenic zebrafish. Biol. Open 2014, 4, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Chung, A.Y.; Kim, P.S.; Kim, S.; Kim, E.; Kim, D.; Jeong, I.; Kim, H.K.; Ryu, J.H.; Kim, C.H.; Choi, J.; et al. Generation of demyelination models by targeted ablation of oligodendrocytes in the zebrafish CNS. Mol. Cells 2013, 36, 82–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaya, F.; Mannioui, A.; Chesneau, A.; Sekizar, S.; Maillard, E.; Ballagny, C.; Houel-Renault, L.; Dupasquier, D.; Bronchain, O.; Holtzmann, I.; et al. Live imaging of targeted cell ablation in Xenopus: A new model to study demyelination and repair. J. Neurosci. Off. J. Soc. Neurosci. 2012, 32, 12885–12895. [Google Scholar] [CrossRef] [Green Version]
- Sekizar, S.; Mannioui, A.; Azoyan, L.; Colin, C.; Thomas, J.L.; Du Pasquier, D.; Mallat, M.; Zalc, B. Remyelination by Resident Oligodendrocyte Precursor Cells in a Xenopus laevis Inducible Model of Demyelination. Dev. Neurosci. 2015, 37, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Mannioui, A.; Zalc, B. Conditional Demyelination and Remyelination in a Transgenic Xenopus laevis. Methods Mol. Biol. 2019, 1936, 239–248. [Google Scholar] [CrossRef]
- Drenthen, G.S.; Fonseca Wald, E.L.A.; Backes, W.H.; Debeij-Van Hall, M.; Hendriksen, J.G.M.; Aldenkamp, A.P.; Vermeulen, R.J.; Klinkenberg, S.; Jansen, J.F.A. Lower myelin-water content of the frontal lobe in childhood absence epilepsy. Epilepsia 2019, 60, 1689–1696. [Google Scholar] [CrossRef] [Green Version]
- Bencurova, P.; Laakso, H.; Salo, R.A.; Paasonen, E.; Manninen, E.; Paasonen, J.; Michaeli, S.; Mangia, S.; Bares, M.; Brazdil, M.; et al. Infantile status epilepticus disrupts myelin development. Neurobiol. Dis. 2022, 162, 105566. [Google Scholar] [CrossRef] [PubMed]
- de Curtis, M.; Garbelli, R.; Uva, L. A hypothesis for the role of axon demyelination in seizure generation. Epilepsia 2021, 62, 583–595. [Google Scholar] [CrossRef] [PubMed]
- Reith, C.A.; Sillar, K.T. Development and role of GABA(A) receptor-mediated synaptic potentials during swimming in postembryonic Xenopus laevis tadpoles. J. Neurophysiol. 1999, 82, 3175–3187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, C.; Hollmann, M. Molecular and functional characterization of Xenopus laevis N-methyl-d-aspartate receptors. Mol. Cell. Neurosci. 2009, 42, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.; Klein, C.; Hollmann, M. Xenopus laevis oocytes endogenously express all subunits of the ionotropic glutamate receptor family. J. Mol. Biol. 2009, 390, 182–195. [Google Scholar] [CrossRef]
- Cox, J.A.; Kucenas, S.; Voigt, M.M. Molecular characterization and embryonic expression of the family of N-methyl-D-aspartate receptor subunit genes in the zebrafish. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2005, 234, 756–766. [Google Scholar] [CrossRef] [PubMed]
- Wahlstrom-Helgren, S.; Montgomery, J.E.; Vanpelt, K.T.; Biltz, S.L.; Peck, J.H.; Masino, M.A. Glutamate receptor subtypes differentially contribute to optogenetically activated swimming in spinally transected zebrafish larvae. J. Neurophysiol. 2019, 122, 2414–2426. [Google Scholar] [CrossRef]
- Turrini, L.; Fornetto, C.; Marchetto, G.; Müllenbroich, M.C.; Tiso, N.; Vettori, A.; Resta, F.; Masi, A.; Mannaioni, G.; Pavone, F.S.; et al. Optical mapping of neuronal activity during seizures in zebrafish. Sci. Rep. 2017, 7, 3025. [Google Scholar] [CrossRef] [Green Version]
- Rosch, R.E.; Hunter, P.R.; Baldeweg, T.; Friston, K.J.; Meyer, M.P. Calcium imaging and dynamic causal modelling reveal brain-wide changes in effective connectivity and synaptic dynamics during epileptic seizures. PLoS Comput. Biol. 2018, 14, e1006375. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Baraban, S.C. Network Properties Revealed during Multi-Scale Calcium Imaging of Seizure Activity in Zebrafish. eNeuro 2019, 6. [Google Scholar] [CrossRef] [Green Version]
- Burrows, D.R.W.; Samarut, É.; Liu, J.; Baraban, S.C.; Richardson, M.P.; Meyer, M.P.; Rosch, R.E. Imaging epilepsy in larval zebrafish. Eur. J. Paediatr. Neurol. EJPN Off. J. Eur. Paediatr. Neurol. Soc. 2020, 24, 70–80. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Hsieh, F.; Ge, L. A Data-Driven Approach to Predict and Classify Epileptic Seizures from Brain-Wide Calcium Imaging Video Data. IEEE/ACM Trans. Comput. Biol. Bioinform. 2020, 17, 1858–1870. [Google Scholar] [CrossRef]
- Niemeyer, J.E.; Gadamsetty, P.; Chun, C.; Sylvester, S.; Lucas, J.P.; Ma, H.; Schwartz, T.H.; Aksay, E.R.F. Seizures initiate in zones of relative hyperexcitation in a zebrafish epilepsy model. Brain A J. Neurol. 2022. [Google Scholar] [CrossRef]
- Cline, H.T. Imaging Structural and Functional Dynamics in Xenopus Neurons. Cold Spring Harb. Protoc. 2022, 2022, top106773. [Google Scholar] [CrossRef] [PubMed]
- Li, V.J.; Schohl, A.; Ruthazer, E.S. Topographic map formation and the effects of NMDA receptor blockade in the developing visual system. Proc. Natl. Acad. Sci. USA 2022, 119, e2107899119. [Google Scholar] [CrossRef]
- Banote, R.K.; Koutarapu, S.; Chennubhotla, K.S.; Chatti, K.; Kulkarni, P. Oral gabapentin suppresses pentylenetetrazole-induced seizure-like behavior and cephalic field potential in adult zebrafish. Epilepsy Behav. EB 2013, 27, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.; Dhamne, S.C.; LaCoursiere, C.M.; Tambunan, D.; Poduri, A.; Rotenberg, A. Microarray Noninvasive Neuronal Seizure Recordings from Intact Larval Zebrafish. PLoS ONE 2016, 11, e0156498. [Google Scholar] [CrossRef] [Green Version]
- Bandara, S.B.; Carty, D.R.; Singh, V.; Harvey, D.J.; Vasylieva, N.; Pressly, B.; Wulff, H.; Lein, P.J. Susceptibility of larval zebrafish to the seizurogenic activity of GABA type A receptor antagonists. Neurotoxicology 2020, 76, 220–234. [Google Scholar] [CrossRef] [PubMed]
- Cozzolino, O.; Sicca, F.; Paoli, E.; Trovato, F.; Santorelli, F.M.; Ratto, G.M.; Marchese, M. Evolution of Epileptiform Activity in Zebrafish by Statistical-Based Integration of Electrophysiology and 2-Photon Ca(2+) Imaging. Cells 2020, 9, 769. [Google Scholar] [CrossRef] [Green Version]
- Hadjiabadi, D.; Lovett-Barron, M.; Raikov, I.G.; Sparks, F.T.; Liao, Z.; Baraban, S.C.; Leskovec, J.; Losonczy, A.; Deisseroth, K.; Soltesz, I. Maximally selective single-cell target for circuit control in epilepsy models. Neuron 2021, 109, 2556–2572.e2556. [Google Scholar] [CrossRef]
- Baraban, S.C.; Taylor, M.R.; Castro, P.A.; Baier, H. Pentylenetetrazole induced changes in zebrafish behavior, neural activity and c-fos expression. Neuroscience 2005, 131, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Hewapathirane, D.S.; Dunfield, D.; Yen, W.; Chen, S.; Haas, K. In vivo imaging of seizure activity in a novel developmental seizure model. Exp Neurol 2008, 211, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lin, Z.J.; Liu, L.; Xu, H.Q.; Shi, Y.W.; Yi, Y.H.; He, N.; Liao, W.P. Epilepsy-associated genes. Seizure 2017, 44, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Cárdenas-Rodríguez, N.; Carmona-Aparicio, L.; Pérez-Lozano, D.L.; Ortega-Cuellar, D.; Gómez-Manzo, S.; Ignacio-Mejía, I. Genetic variations associated with pharmacoresistant epilepsy (Review). Mol. Med. Rep. 2020, 21, 1685–1701. [Google Scholar] [CrossRef] [PubMed]
- Pauli, A.; Montague, T.G.; Lennox, K.A.; Behlke, M.A.; Schier, A.F. Antisense Oligonucleotide-Mediated Transcript Knockdown in Zebrafish. PLoS ONE 2015, 10, e0139504. [Google Scholar] [CrossRef] [Green Version]
- Dubińska-Magiera, M.; Chmielewska, M.; Kozioł, K.; Machowska, M.; Hutchison, C.J.; Goldberg, M.W.; Rzepecki, R. Xenopus LAP2β protein knockdown affects location of lamin B and nucleoporins and has effect on assembly of cell nucleus and cell viability. Protoplasma 2016, 253, 943–956. [Google Scholar] [CrossRef] [Green Version]
- Crossley, M.P.; Krude, T. Targeting Functional Noncoding RNAs. Methods Mol. Biol. 2017, 1565, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Stainier, D.Y.R.; Raz, E.; Lawson, N.D.; Ekker, S.C.; Burdine, R.D.; Eisen, J.S.; Ingham, P.W.; Schulte-Merker, S.; Yelon, D.; Weinstein, B.M.; et al. Guidelines for morpholino use in zebrafish. PLoS Genet. 2017, 13, e1007000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Clark, K.; Barton, C.; Tanguay, R.; Moulton, H. A Novel Zebrafish Model for Assessing In Vivo Delivery of Morpholino Oligomers. Methods Mol. Biol. 2018, 1828, 293–306. [Google Scholar] [CrossRef]
- Paraiso, K.D.; Blitz, I.L.; Zhou, J.J.; Cho, K.W.Y. Morpholinos Do Not Elicit an Innate Immune Response during Early Xenopus Embryogenesis. Dev. Cell 2019, 49, 643–650 e643. [Google Scholar] [CrossRef] [PubMed]
- Van Gils, M.; Vanakker, O.M. Morpholino-Mediated Gene Knockdown in Zebrafish: It Is All About Dosage and Validation. J. Investig. Dermatol. 2019, 139, 1599–1600. [Google Scholar] [CrossRef]
- Iida, M.; Suzuki, M.; Sakane, Y.; Nishide, H.; Uchiyama, I.; Yamamoto, T.; Suzuki, K.T.; Fujii, S. A simple and practical workflow for genotyping of CRISPR-Cas9-based knockout phenotypes using multiplexed amplicon sequencing. Genes Cells Devoted Mol. Cell. Mech. 2020, 25, 498–509. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.H.; Zhang, G. Generating Stable Knockout Zebrafish Lines by Deleting Large Chromosomal Fragments Using Multiple gRNAs. G3 (Bethesda Md.) 2020, 10, 1029–1037. [Google Scholar] [CrossRef] [Green Version]
- Kroll, F.; Powell, G.T.; Ghosh, M.; Gestri, G.; Antinucci, P.; Hearn, T.J.; Tunbak, H.; Lim, S.; Dennis, H.W.; Fernandez, J.M.; et al. A simple and effective F0 knockout method for rapid screening of behaviour and other complex phenotypes. eLife 2021, 10, e59683. [Google Scholar] [CrossRef]
- Zhang, C.; Li, J.; Tarique, I.; Zhang, Y.; Lu, T.; Wang, J.; Chen, A.; Wen, F.; Zhang, Z.; Zhang, Y.; et al. A Time-Saving Strategy to Generate Double Maternal Mutants by an Oocyte-Specific Conditional Knockout System in Zebrafish. Biology 2021, 10, 777. [Google Scholar] [CrossRef] [PubMed]
- Aksoy, Y.A.; Yang, B.; Chen, W.; Hung, T.; Kuchel, R.P.; Zammit, N.W.; Grey, S.T.; Goldys, E.M.; Deng, W. Spatial and Temporal Control of CRISPR-Cas9-Mediated Gene Editing Delivered via a Light-Triggered Liposome System. ACS Appl. Mater. Interfaces 2020, 12, 52433–52444. [Google Scholar] [CrossRef]
- Quick, R.E.; Buck, L.D.; Parab, S.; Tolbert, Z.R.; Matsuoka, R.L. Highly Efficient Synthetic CRISPR RNA/Cas9-Based Mutagenesis for Rapid Cardiovascular Phenotypic Screening in F0 Zebrafish. Front. Cell Dev. Biol. 2021, 9, 735598. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Sharma, B.S.; Verma, R.J. CRISPR-based genome editing of zebrafish. Prog. Mol. Biol. Transl. Sci. 2021, 180, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Uribe-Salazar, J.M.; Kaya, G.; Sekar, A.; Weyenberg, K.; Ingamells, C.; Dennis, M.Y. Evaluation of CRISPR gene-editing tools in zebrafish. BMC Genom. 2022, 23, 12. [Google Scholar] [CrossRef]
- Zhang, C.; Lu, T.; Zhang, Y.; Li, J.; Tarique, I.; Wen, F.; Chen, A.; Wang, J.; Zhang, Z.; Zhang, Y.; et al. Rapid generation of maternal mutants via oocyte transgenic expression of CRISPR-Cas9 and sgRNAs in zebrafish. Sci. Adv. 2021, 7, eabg4243. [Google Scholar] [CrossRef] [PubMed]
- Abdelrahman, D.; Hasan, W.; Da’as, S.I. Microinjection quality control in zebrafish model for genetic manipulations. MethodsX 2021, 8, 101418. [Google Scholar] [CrossRef]
- Lane, M.; Mis, E.K.; Khokha, M.K. Microinjection of Xenopus tropicalis Embryos. Cold Spring Harb. Protoc. 2022, 2022, prot107644. [Google Scholar] [CrossRef]
- Moody, S.A. Microinjection of mRNAs and Oligonucleotides. Cold Spring Harb. Protoc. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Szaro, B.G. A method for using direct injection of plasmid DNA to study cis-regulatory element activity in F0 Xenopus embryos and tadpoles. Dev. Biol. 2015, 398, 11–23. [Google Scholar] [CrossRef] [Green Version]
- Yasuoka, Y.; Taira, M. Microinjection of DNA Constructs into Xenopus Embryos for Gene Misexpression and cis-Regulatory Module Analysis. Cold Spring Harb. Protoc. 2019, 2019. [Google Scholar] [CrossRef]
- Cho, S.J.; Park, E.; Baker, A.; Reid, A.Y. Post-Traumatic Epilepsy in Zebrafish Is Drug-Resistant and Impairs Cognitive Function. J. Neurotrauma 2021, 38, 3174–3183. [Google Scholar] [CrossRef]
- Kumari, S.; Sharma, P.; Mazumder, A.G.; Rana, A.K.; Sharma, S.; Singh, D. Development and validation of chemical kindling in adult zebrafish: A simple and improved chronic model for screening of antiepileptic agents. J. Neurosci. Methods 2020, 346, 108916. [Google Scholar] [CrossRef]
- Mussulini, B.H.; Leite, C.E.; Zenki, K.C.; Moro, L.; Baggio, S.; Rico, E.P.; Rosemberg, D.B.; Dias, R.D.; Souza, T.M.; Calcagnotto, M.E.; et al. Seizures induced by pentylenetetrazole in the adult zebrafish: A detailed behavioral characterization. PLoS ONE 2013, 8, e54515. [Google Scholar] [CrossRef]
- Samarut, E.; Swaminathan, A.; Riche, R.; Liao, M.; Hassan-Abdi, R.; Renault, S.; Allard, M.; Dufour, L.; Cossette, P.; Soussi-Yanicostas, N.; et al. gamma-Aminobutyric acid receptor alpha 1 subunit loss of function causes genetic generalized epilepsy by impairing inhibitory network neurodevelopment. Epilepsia 2018, 59, 2061–2074. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Lee, K.J.; Jang, J.W.; Lee, S.I.; Kim, S. An EEG system to detect brain signals from multiple adult zebrafish. Biosens. Bioelectron. 2020, 164, 112315. [Google Scholar] [CrossRef]
- Cassar, S.; Breidenbach, L.; Olson, A.; Huang, X.; Britton, H.; Woody, C.; Sancheti, P.; Stolarik, D.; Wicke, K.; Hempel, K.; et al. Measuring drug absorption improves interpretation of behavioral responses in a larval zebrafish locomotor assay for predicting seizure liability. J. Pharmacol. Toxicol. Methods 2017, 88, 56–63. [Google Scholar] [CrossRef]
- Van Wijk, R.C.; Krekels, E.H.J.; Kantae, V.; Ordas, A.; Kreling, T.; Harms, A.C.; Hankemeier, T.; Spaink, H.P.; van der Graaf, P.H. Mechanistic and Quantitative Understanding of Pharmacokinetics in Zebrafish Larvae through Nanoscale Blood Sampling and Metabolite Modeling of Paracetamol. J. Pharmacol. Exp. Ther. 2019, 371, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Howard, A.M.; Papich, M.G.; Felt, S.A.; Long, C.T.; McKeon, G.P.; Bond, E.S.; Torreilles, S.L.; Luong, R.H.; Green, S.L. The pharmacokinetics of enrofloxacin in adult African clawed frogs (Xenopus laevis). J. Am. Assoc. Lab. Anim. Sci. JAALAS 2010, 49, 800–804. [Google Scholar]
- Fleming, A.; Diekmann, H.; Goldsmith, P. Functional characterisation of the maturation of the blood-brain barrier in larval zebrafish. PLoS ONE 2013, 8, e77548. [Google Scholar] [CrossRef]
- Johnston, G.A. Advantages of an antagonist: Bicuculline and other GABA antagonists. Br. J. Pharmacol. 2013, 169, 328–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishek, B.J.; Moss, S.J.; Smart, T.G. A functional comparison of the antagonists bicuculline and picrotoxin at recombinant GABAA receptors. Neuropharmacology 1996, 35, 1289–1298. [Google Scholar] [CrossRef]
- Simmonds, M.A. Evidence that bicuculline and picrotoxin act at separate sites to antagonize gamma-aminobutyric acid in rat cuneate nucleus. Neuropharmacology 1980, 19, 39–45. [Google Scholar] [CrossRef]
- de Feo, M.R.; Mecarelli, O.; Ricci, G.F. Bicuculline- and allylglycine-induced epilepsy in developing rats. Exp. Neurol. 1985, 90, 411–421. [Google Scholar] [CrossRef]
- Jones, A.; Barker-Haliski, M.; Ilie, A.S.; Herd, M.B.; Baxendale, S.; Holdsworth, C.J.; Ashton, J.P.; Placzek, M.; Jayasekera, B.A.P.; Cowie, C.J.A.; et al. A multiorganism pipeline for antiseizure drug discovery: Identification of chlorothymol as a novel γ-aminobutyric acidergic anticonvulsant. Epilepsia 2020, 61, 2106–2118. [Google Scholar] [CrossRef] [PubMed]
- Murao, K.; Shingu, K.; Miyamoto, E.; Ikeda, S.; Nakao, S.; Masuzawa, M.; Yamada, M. Anticonvulsant effects of sevoflurane on amygdaloid kindling and bicuculline-induced seizures in cats: Comparison with isoflurane and halothane. J. Anesth. 2002, 16, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Soares-Silva, B.; Beserra-Filho, J.I.A.; Morera, P.M.A.; Custódio-Silva, A.C.; Maria-Macêdo, A.; Silva-Martins, S.; Alexandre-Silva, V.; Silva, S.P.; Silva, R.H.; Ribeiro, A.M. The bee venom active compound melittin protects against bicuculline-induced seizures and hippocampal astrocyte activation in rats. Neuropeptides 2022, 91, 102209. [Google Scholar] [CrossRef]
- Vicente-Silva, W.; Silva-Freitas, F.R.; Beserra-Filho, J.I.A.; Cardoso, G.N.; Silva-Martins, S.; Sarno, T.A.; Silva, S.P.; Soares-Silva, B.; Dos Santos, J.R.; da Silva, R.H.; et al. Sakuranetin exerts anticonvulsant effect in bicuculline-induced seizures. Fundam. Clin. Pharmacol. 2022. [Google Scholar] [CrossRef]
- Wong, K.; Stewart, A.; Gilder, T.; Wu, N.; Frank, K.; Gaikwad, S.; Suciu, C.; Dileo, J.; Utterback, E.; Chang, K.; et al. Modeling seizure-related behavioral and endocrine phenotypes in adult zebrafish. Brain Res. 2010, 1348, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Stankevicius, D.; Rodrigues-Costa, E.C.; Camilo Florio, J.; Palermo-Neto, J. Neuroendocrine, behavioral and macrophage activity changes induced by picrotoxin effects in mice. Neuropharmacology 2008, 54, 300–308. [Google Scholar] [CrossRef]
- Acharya, M.M.; Khamesra, S.H.; Katyare, S.S. Picrotoxin-induced convulsions and lysosomal function in the rat brain. Indian J. Clin. Biochem. IJCB 2005, 20, 56–60. [Google Scholar] [CrossRef] [Green Version]
- Acharya, M.M.; Khamesra, S.H.; Katyare, S.S. Effect of repeated intraperitoneal exposure to picrotoxin on rat liver lysosomal function. Indian J. Exp. Biol. 2004, 42, 808–811. [Google Scholar]
- Li, J.M.; Gan, J.; Zeng, T.F.; Sander, J.W.; Zhou, D. Tetramethylenedisulfotetramine intoxication presenting with de novo Status Epilepticus: A case series. Neurotoxicology 2012, 33, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Mundy, P.C.; Pressly, B.; Carty, D.R.; Yaghoobi, B.; Wulff, H.; Lein, P.J. The efficacy of gamma-aminobutyric acid type A receptor (GABA AR) subtype-selective positive allosteric modulators in blocking tetramethylenedisulfotetramine (TETS)-induced seizure-like behavior in larval zebrafish with minimal sedation. Toxicol. Appl. Pharmacol. 2021, 426, 115643. [Google Scholar] [CrossRef]
- Maroso, M.; Balosso, S.; Ravizza, T.; Liu, J.; Aronica, E.; Iyer, A.M.; Rossetti, C.; Molteni, M.; Casalgrandi, M.; Manfredi, A.A.; et al. Toll-like receptor 4 and high-mobility group box-1 are involved in ictogenesis and can be targeted to reduce seizures. Nat. Med. 2010, 16, 413–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bragin, A.; Azizyan, A.; Almajano, J.; Wilson, C.L.; Engel, J., Jr. Analysis of chronic seizure onsets after intrahippocampal kainic acid injection in freely moving rats. Epilepsia 2005, 46, 1592–1598. [Google Scholar] [CrossRef]
- Bragin, A.; Wilson, C.L.; Engel, J., Jr. Chronic epileptogenesis requires development of a network of pathologically interconnected neuron clusters: A hypothesis. Epilepsia 2000, 41 (Suppl. S6), S144–S152. [Google Scholar] [CrossRef] [Green Version]
- Zupanc, G.K.; Hinsch, K.; Gage, F.H. Proliferation, migration, neuronal differentiation, and long-term survival of new cells in the adult zebrafish brain. J. Comp. Neurol. 2005, 488, 290–319. [Google Scholar] [CrossRef] [PubMed]
- Heylen, L.; Pham, D.H.; De Meulemeester, A.S.; Samarut, E.; Skiba, A.; Copmans, D.; Kazwiny, Y.; Vanden Berghe, P.; de Witte, P.A.M.; Siekierska, A. Pericardial Injection of Kainic Acid Induces a Chronic Epileptic State in Larval Zebrafish. Front. Mol. Neurosci. 2021, 14, 753936. [Google Scholar] [CrossRef]
- Paudel, Y.N.; Kumari, Y.; Abidin, S.A.Z.; Othman, I.; Shaikh, M.F. Pilocarpine Induced Behavioral and Biochemical Alterations in Chronic Seizure-Like Condition in Adult Zebrafish. Int J. Mol. Sci 2020, 21, 2492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paudel, Y.N.; Othman, I.; Shaikh, M.F. Anti-High Mobility Group Box-1 Monoclonal Antibody Attenuates Seizure-Induced Cognitive Decline by Suppressing Neuroinflammation in an Adult Zebrafish Model. Front. Pharm. 2020, 11, 613009. [Google Scholar] [CrossRef] [PubMed]
- Heuzeroth, H.; Wawra, M.; Fidzinski, P.; Dag, R.; Holtkamp, M. The 4-Aminopyridine Model of Acute Seizures in vitro Elucidates Efficacy of New Antiepileptic Drugs. Front. Neurosci. 2019, 13, 677. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R. Encyclopedia of Toxicology; Academic Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Blight, A.R.; Henney, H.R., 3rd; Cohen, R. Development of dalfampridine, a novel pharmacologic approach for treating walking impairment in multiple sclerosis. Ann. N. Y. Acad. Sci. 2014, 1329, 33–44. [Google Scholar] [CrossRef]
- DeForge, D.; Nymark, J.; Lemaire, E.; Gardner, S.; Hunt, M.; Martel, L.; Curran, D.; Barbeau, H. Effect of 4-aminopyridine on gait in ambulatory spinal cord injuries: A double-blind, placebo-controlled, crossover trial. Spinal Cord 2004, 42, 674–685. [Google Scholar] [CrossRef] [Green Version]
- Hayes, K.C. The use of 4-aminopyridine (fampridine) in demyelinating disorders. CNS Drug Rev. 2004, 10, 295–316. [Google Scholar] [CrossRef]
- Rahimi, N.; Modabberi, S.; Faghir-Ghanesefat, H.; Shayan, M.; Farzad Maroufi, S.; Asgari Dafe, E.; Reza Dehpour, A. The possible role of nitric oxide signaling and NMDA receptors in allopurinol effect on maximal electroshock- and pentylenetetrazol-induced seizures in mice. Neurosci. Lett. 2022, 778, 136620. [Google Scholar] [CrossRef]
- van der Linde, H.; Kreir, M.; Teisman, A.; Gallacher, D.J. Seizure-induced Torsades de pointes: In a canine drug-induced long-QT1 model. J. Pharmacol. Toxicol. Methods 2021, 111, 107086. [Google Scholar] [CrossRef]
- Nagata, S.; Fujiwara, K.; Kuga, K.; Ozaki, H. Prediction of GABA receptor antagonist-induced convulsion in cynomolgus monkeys by combining machine learning and heart rate variability analysis. J. Pharmacol. Toxicol. Methods 2021, 112, 107127. [Google Scholar] [CrossRef] [PubMed]
- Loscher, W. Critical review of current animal models of seizures and epilepsy used in the discovery and development of new antiepileptic drugs. Seizure 2011, 20, 359–368. [Google Scholar] [CrossRef] [Green Version]
- Alachkar, A.; Ojha, S.K.; Sadeq, A.; Adem, A.; Frank, A.; Stark, H.; Sadek, B. Experimental Models for the Discovery of Novel Anticonvulsant Drugs: Focus on Pentylenetetrazole-Induced Seizures and Associated Memory Deficits. Curr. Pharm. Des. 2020, 26, 1693–1711. [Google Scholar] [CrossRef]
- Chipiti, T.; Viljoen, A.M.; Cordero-Maldonado, M.L.; Veale, C.G.L.; Van Heerden, F.R.; Sandasi, M.; Chen, W.; Crawford, A.D.; Enslin, G.M. Anti-seizure activity of African medicinal plants: The identification of bioactive alkaloids from the stem bark of Rauvolfia caffra using an in vivo zebrafish model. J. Ethnopharmacol. 2021, 279, 114282. [Google Scholar] [CrossRef]
- Copmans, D.; Orellana-Paucar, A.M.; Steurs, G.; Zhang, Y.; Ny, A.; Foubert, K.; Exarchou, V.; Siekierska, A.; Kim, Y.; De Borggraeve, W.; et al. Methylated flavonoids as anti-seizure agents: Naringenin 4′,7-dimethyl ether attenuates epileptic seizures in zebrafish and mouse models. Neurochem. Int. 2018, 112, 124–133. [Google Scholar] [CrossRef] [Green Version]
- Copmans, D.; Rateb, M.; Tabudravu, J.N.; Perez-Bonilla, M.; Dirkx, N.; Vallorani, R.; Diaz, C.; Perez Del Palacio, J.; Smith, A.J.; Ebel, R.; et al. Zebrafish-Based Discovery of Antiseizure Compounds from the Red Sea: Pseurotin A2 and Azaspirofuran A. ACS Chem. Neurosci. 2018, 9, 1652–1662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koziol, E.; Jozwiak, K.; Budzynska, B.; de Witte, P.A.M.; Copmans, D.; Skalicka-Wozniak, K. Comparative Antiseizure Analysis of Diverse Natural Coumarin Derivatives in Zebrafish. Int. J. Mol. Sci. 2021, 22, 1420. [Google Scholar] [CrossRef]
- Choo, B.K.M.; Kundap, U.P.; Kumari, Y.; Hue, S.M.; Othman, I.; Shaikh, M.F. Orthosiphon stamineus Leaf Extract Affects TNF-alpha and Seizures in a Zebrafish Model. Front. Pharm. 2018, 9, 139. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.; Vishwanath, M.; Zhang, J.; Sarafan, S.; Trigo Torres, R.S.; Le, T.; Lau, M.P.H.; Nguyen, A.H.; Cao, H. Microelectrode array membranes to simultaneously assess cardiac and neurological signals of xenopus laevis under chemical exposures and environmental changes. Biosens. Bioelectron. 2022, 210, 114292. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Xie, X.; Walker, S.; Saxena, M.; Kozlowski, D.J.; Mumm, J.S.; Cowell, J.K. Loss of zebrafish lgi1b leads to hydrocephalus and sensitization to pentylenetetrazol induced seizure-like behavior. PLoS ONE 2011, 6, e24596. [Google Scholar] [CrossRef]
- Johnson, M.R.; Behmoaras, J.; Bottolo, L.; Krishnan, M.L.; Pernhorst, K.; Santoscoy, P.L.M.; Rossetti, T.; Speed, D.; Srivastava, P.K.; Chadeau-Hyam, M.; et al. Systems genetics identifies Sestrin 3 as a regulator of a proconvulsant gene network in human epileptic hippocampus. Nat. Commun. 2015, 6, 6031. [Google Scholar] [CrossRef] [Green Version]
- Mei, X.; Wu, S.; Bassuk, A.G.; Slusarski, D.C. Mechanisms of prickle1a function in zebrafish epilepsy and retinal neurogenesis. Dis. Models Mech. 2013, 6, 679–688. [Google Scholar] [CrossRef] [Green Version]
- Baraban, S.C. A zebrafish-centric approach to antiepileptic drug development. Dis. Models Mech. 2021, 14. [Google Scholar] [CrossRef]
- Baraban, S.C.; Dinday, M.T.; Hortopan, G.A. Drug screening in Scn1a zebrafish mutant identifies clemizole as a potential Dravet syndrome treatment. Nat. Commun. 2013, 4, 2410. [Google Scholar] [CrossRef] [Green Version]
- Moog, M.; Baraban, S.C. Clemizole and Trazodone are Effective Antiseizure Treatments in a Zebrafish Model of STXBP1 Disorder. Epilepsia Open 2022. [Google Scholar] [CrossRef]
- Xu, Y.; Song, R.; Chen, W.; Strong, K.; Shrey, D.; Gedela, S.; Traynelis, S.F.; Zhang, G.; Yuan, H. Recurrent seizure-related GRIN1 variant: Molecular mechanism and targeted therapy. Ann. Clin. Transl. Neurol. 2021, 8, 1480–1494. [Google Scholar] [CrossRef] [PubMed]
- Sega, A.G.; Mis, E.K.; Lindstrom, K.; Mercimek-Andrews, S.; Ji, W.; Cho, M.T.; Juusola, J.; Konstantino, M.; Jeffries, L.; Khokha, M.K.; et al. De novo pathogenic variants in neuronal differentiation factor 2 (NEUROD2) cause a form of early infantile epileptic encephalopathy. J. Med. Genet. 2019, 56, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Hirai, H.; Iino, M.; Fuse, I.; Mitsumura, K.; Washiyama, K.; Kasai, S.; Ikeda, K. Inhibitory effects of the antiepileptic drug ethosuximide on G protein-activated inwardly rectifying K+ channels. Neuropharmacology 2009, 56, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Ottosson, N.E.; Silvera Ejneby, M.; Wu, X.; Estrada-Mondragon, A.; Nilsson, M.; Karlsson, U.; Schupp, M.; Rognant, S.; Jepps, T.A.; Konradsson, P.; et al. Synthetic resin acid derivatives selectively open the hKV 7.2/7.3 channel and prevent epileptic seizures. Epilepsia 2021, 62, 1744–1758. [Google Scholar] [CrossRef] [PubMed]
- Stadler, M.; Monticelli, S.; Seidel, T.; Luger, D.; Salzer, I.; Boehm, S.; Holzer, W.; Schwarzer, C.; Urban, E.; Khom, S.; et al. Design, Synthesis, and Pharmacological Evaluation of Novel beta2/3 Subunit-Selective gamma-Aminobutyric Acid Type A (GABAA) Receptor Modulators. J. Med. Chem. 2019, 62, 317–341. [Google Scholar] [CrossRef]
- Kobayashi, T.; Ikeda, K.; Kumanishi, T. Inhibition by various antipsychotic drugs of the G-protein-activated inwardly rectifying K(+) (GIRK) channels expressed in xenopus oocytes. Br. J. Pharmacol. 2000, 129, 1716–1722. [Google Scholar] [CrossRef] [Green Version]
- Anderson, L.L.; Absalom, N.L.; Abelev, S.V.; Low, I.K.; Doohan, P.T.; Martin, L.J.; Chebib, M.; McGregor, I.S.; Arnold, J.C. Coadministered cannabidiol and clobazam: Preclinical evidence for both pharmacodynamic and pharmacokinetic interactions. Epilepsia 2019, 60, 2224–2234. [Google Scholar] [CrossRef] [Green Version]
- Bell, M.R.; Belarde, J.A.; Johnson, H.F.; Aizenman, C.D. A neuroprotective role for polyamines in a Xenopus tadpole model of epilepsy. Nat. Neurosci. 2011, 14, 505–512. [Google Scholar] [CrossRef]
- Najm, I.; el-Skaf, G.; Tocco, G.; Vanderklish, P.; Lynch, G.; Baudry, M. Seizure activity-induced changes in polyamine metabolism and neuronal pathology during the postnatal period in rat brain. Brain Res. Dev. Brain Res. 1992, 69, 11–21. [Google Scholar] [CrossRef]
- Ibhazehiebo, K.; Gavrilovici, C.; de la Hoz, C.L.; Ma, S.C.; Rehak, R.; Kaushik, G.; Meza Santoscoy, P.L.; Scott, L.; Nath, N.; Kim, D.Y.; et al. A novel metabolism-based phenotypic drug discovery platform in zebrafish uncovers HDACs 1 and 3 as a potential combined anti-seizure drug target. Brain J. Neurol. 2018, 141, 744–761. [Google Scholar] [CrossRef] [Green Version]
- Hentig, J.; Campbell, L.J.; Cloghessy, K.; Lee, M.; Boggess, W.; Hyde, D.R. Prophylactic Activation of Shh Signaling Attenuates TBI-Induced Seizures in Zebrafish by Modulating Glutamate Excitotoxicity through Eaat2a. Biomedicines 2021, 10, 32. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, Y.; Kawasaki, T. Stab Wound Injury Model of the Adult Optic Tectum using Zebrafish and Medaka for the Comparative Analysis of Regenerative Capacity. J. Vis. Exp. JoVE 2022. [Google Scholar] [CrossRef] [PubMed]
- Uemura, N.; Koike, M.; Ansai, S.; Kinoshita, M.; Ishikawa-Fujiwara, T.; Matsui, H.; Naruse, K.; Sakamoto, N.; Uchiyama, Y.; Todo, T.; et al. Viable neuronopathic Gaucher disease model in Medaka (Oryzias latipes) displays axonal accumulation of alpha-synuclein. PLoS Genet. 2015, 11, e1005065. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, R.E.; Mruk, K. Aquatic Freshwater Vertebrate Models of Epilepsy Pathology: Past Discoveries and Future Directions for Therapeutic Discovery. Int. J. Mol. Sci. 2022, 23, 8608. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23158608
Williams RE, Mruk K. Aquatic Freshwater Vertebrate Models of Epilepsy Pathology: Past Discoveries and Future Directions for Therapeutic Discovery. International Journal of Molecular Sciences. 2022; 23(15):8608. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23158608
Chicago/Turabian StyleWilliams, Rachel E., and Karen Mruk. 2022. "Aquatic Freshwater Vertebrate Models of Epilepsy Pathology: Past Discoveries and Future Directions for Therapeutic Discovery" International Journal of Molecular Sciences 23, no. 15: 8608. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23158608