
Long-Term Alcohol Consumption Caused a Significant Decrease in Serum High-Density Lipoprotein (HDL)-Cholesterol and Apolipoprotein A-I with the Atherogenic Changes of HDL in Middle-Aged Korean Women



Abstract
:1. Introduction
2. Results
2.1. Anthropometric Profiles and Blood Parameters
2.2. Characteristics of Lipoproteins Depend on Alcohol Intake
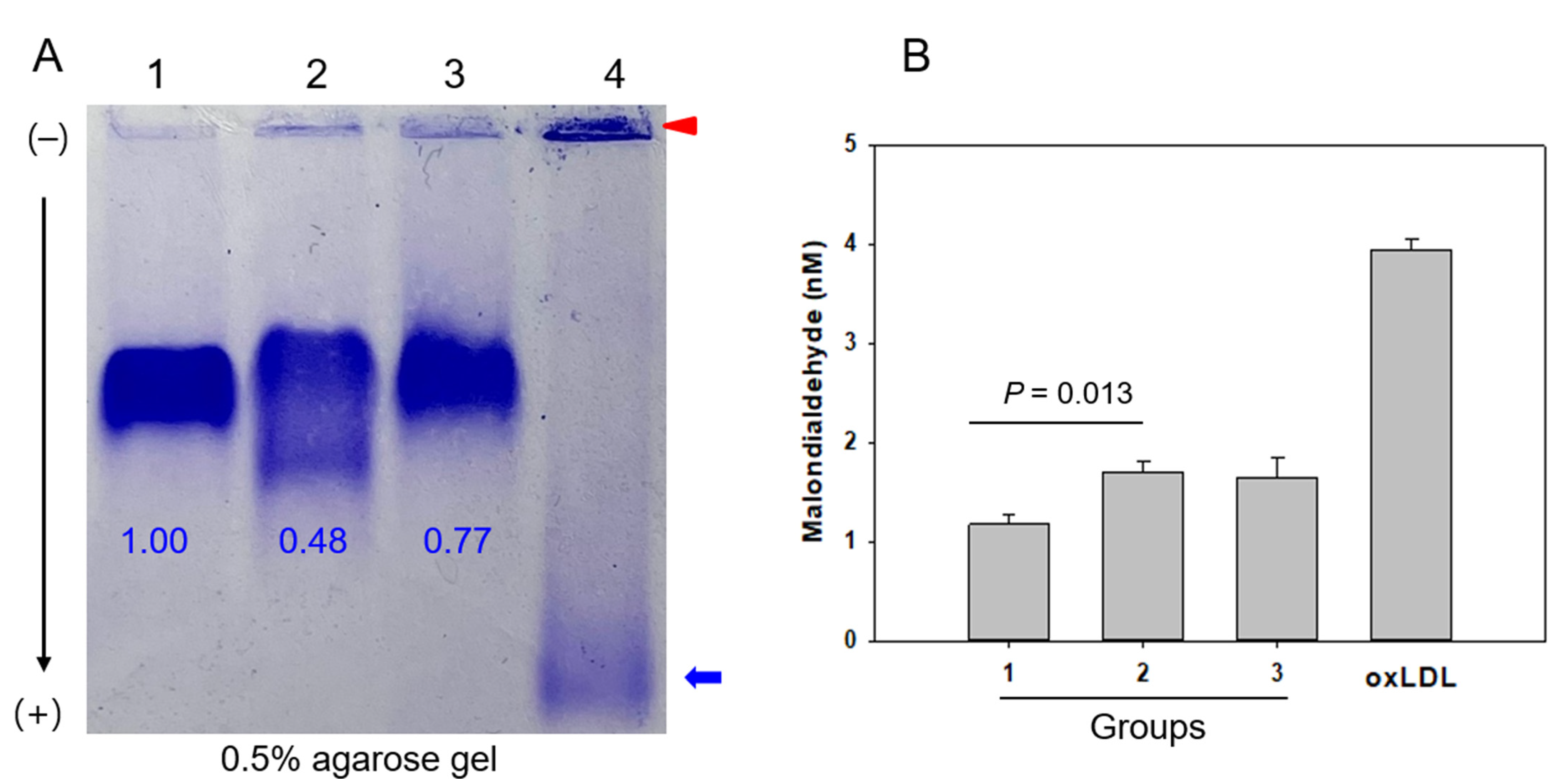
2.3. LDL Particle and Lipid Composition
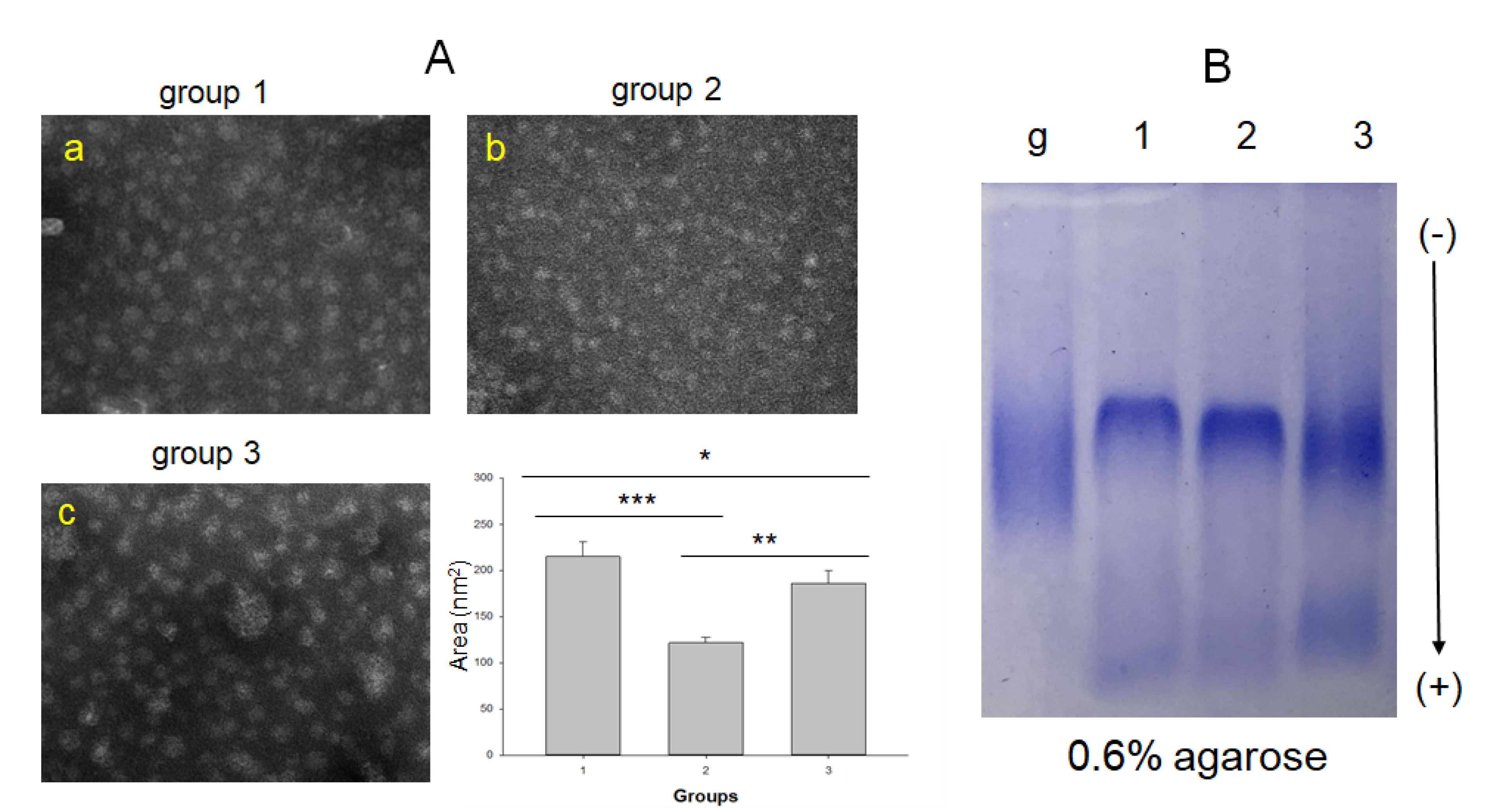
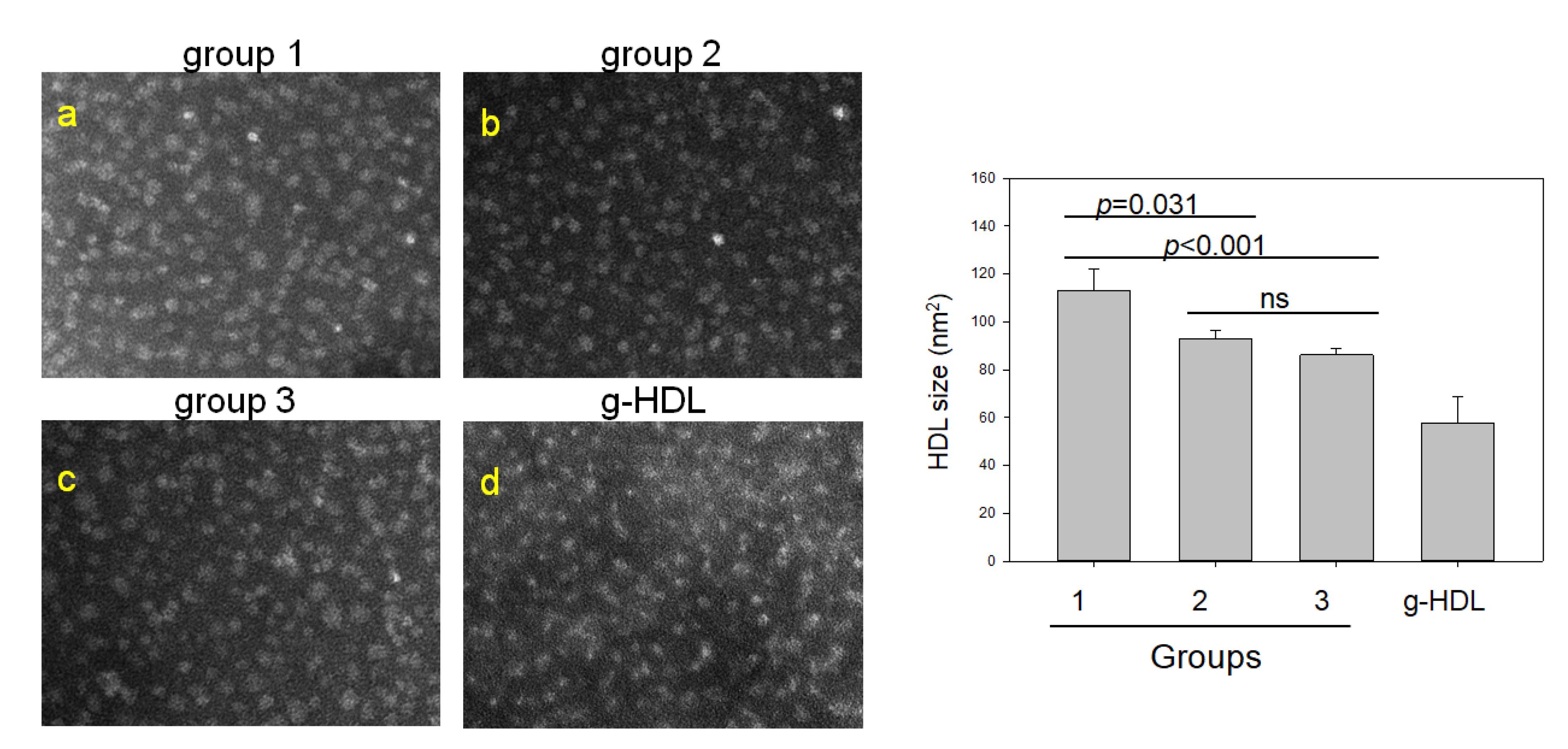
2.4. Comparison of HDL Particles and Lipid Composition
2.5. Spearman Correlation Analysis
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Anthropometric Analysis
4.3. Blood Analysis
4.4. Isolation of Lipoproteins
4.5. LDL Oxidation and Quantification
4.6. Paraoxonase Assay
4.7. Electromobility of Lipoproteins
4.8. Glycation of HDL
4.9. Electron Microscopy
4.10. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baraona, E.; Lieber, C.S. Effects of ethanol on lipid metabolism. J. Lipid Res. 1979, 20, 289. [Google Scholar] [CrossRef]
- Pathogenesis of alcohol-induced accumulation of protein in the liver. J. Clin. Investig. 1977, 60, 546–554. [CrossRef] [PubMed]
- Pflaum, T.; Hausler, T.; Baumung, C.; Ackermann, S.; Kuballa, T.; Rehm, J.; Lachenmeier, D.W. Carcinogenic compounds in alcoholic beverages: An update. Arch. Toxicol. 2016, 90, 2349–2367. [Google Scholar] [CrossRef] [PubMed]
- Kannel, W.B.; Ellison, R.C. Alcohol and coronary heart disease: The evidence for a protective effect. Clin. Chim. Acta 1996, 246, 59–76. [Google Scholar] [CrossRef]
- Rimm, E.B.; Williams, P.; Fosher, K.; Criqui, M.; Stampfer, M.J. Moderate alcohol intake and lower risk of coronary heart disease: Meta-analysis of effects on lipids and haemostatic factors. BMJ 1999, 319, 1523–1528. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.E. Relationship between heavy drinking, binge drinking, and metabolic syndrome in obese and non-obese Korean male adults. Nutr. Res. Pract. 2018, 12, 166–172. [Google Scholar] [CrossRef]
- Frohlich, J.J. Effects of alcohol on plasma lipoprotein metabolism. Clin. Chim. Acta 1996, 246, 39–49. [Google Scholar] [CrossRef]
- Rosales, C.; Gillard, B.K.; Gotto, A.M., Jr.; Pownall, H.J. The Alcohol–High-Density Lipoprotein Athero-Protective Axis. Biomolecules 2020, 10, 987. [Google Scholar] [CrossRef]
- Cederbaum, A.I. Alcohol metabolism. Clin. Liver Dis. 2012, 16, 667–685. [Google Scholar] [CrossRef] [Green Version]
- Palmisano, B.T.; Zhu, L.; Eckel, R.H.; Stafford, J.M. Sex differences in lipid and lipoprotein metabolism. Mol. Metab. 2018, 15, 45–55. [Google Scholar] [CrossRef]
- Sun, K.; Ren, M.; Liu, D.; Wang, C.; Yang, C.; Yan, L. Alcohol consumption and risk of metabolic syndrome: A meta-analysis of prospective studies. Clin. Nutr. 2014, 33, 596–602. [Google Scholar] [CrossRef]
- Yoon, Y.S.; Oh, S.W.; Baik, H.W.; Park, H.S.; Kim, W.Y. Alcohol consumption and the metabolic syndrome in Korean adults: The 1998 Korean National Health and Nutrition Examination Survey. Am. J. Clin. Nutr. 2004, 80, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Minzer, S.; Losno, R.A.; Casas, R. The Effect of Alcohol on Cardiovascular Risk Factors: Is There New Information? Nutrients 2020, 12, 912. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.H. The Current Status of Research on High-Density Lipoproteins (HDL): A Paradigm Shift from HDL Quantity to HDL Quality and HDL Functionality. Int. J. Mol. Sci. 2022, 23, 3967. [Google Scholar] [CrossRef]
- Hannuksela, M.L.; Liisanantti, M.K.; Savolainen, M.J. Effect of Alcohol on Lipids and Lipoproteins in Relation to Atherosclerosis. Crit. Rev. Clin. Lab. Sci. 2002, 39, 225–283. [Google Scholar] [CrossRef]
- Lin, Y.; Ying, Y.Y.; Li, S.X.; Wang, S.J.; Gong, Q.H.; Li, H. Association between alcohol consumption and metabolic syndrome among Chinese adults. Public Health Nutr. 2021, 24, 4582–4590. [Google Scholar] [CrossRef]
- Cho, K.H.; Kim, J.R. Rapid Decrease in HDL-C in the Puberty Period of Boys Associated with an Elevation of Blood Pressure and Dyslipidemia in Korean Teenagers: An Explanation of Why and When Men Have Lower HDL-C Levels Than Women. Med. Sci. 2021, 9, 35. [Google Scholar] [CrossRef]
- Wang, X.; Magkos, F.; Mittendorfer, B. Sex Differences in Lipid and Lipoprotein Metabolism: It’s Not Just about Sex Hormones. J. Clin. Endocrinol. Metab. 2011, 96, 885–893. [Google Scholar] [CrossRef] [Green Version]
- Brien, S.E.; Ronksley, P.E.; Turner, B.J.; Mukamal, K.J.; Ghali, W.A. Effect of alcohol consumption on biological markers associated with risk of coronary heart disease: Systematic review and meta-analysis of interventional studies. BMJ 2011, 342, d636. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, E.; Silva, E.R.; Foster, D.; McGee Harper, M.; Seidman, C.E.; Smith, J.D.; Breslow, J.L.; Brinton, E.A. Alcohol Consumption Raises HDL Cholesterol Levels by Increasing the Transport Rate of Apolipoproteins A-I and A-II. Circulation 2000, 102, 2347–2352. [Google Scholar] [CrossRef]
- Huang, S.; Li, J.; Shearer, G.C.; Lichtenstein, A.H.; Zheng, X.; Wu, Y.; Jin, C.; Wu, S.; Gao, X. Longitudinal study of alcohol consumption and HDL concentrations: A community-based study. Am. J. Clin. Nutr. 2017, 105, 905–912. [Google Scholar] [CrossRef] [Green Version]
- Roerecke, M.; Rehm, J. Alcohol consumption, drinking patterns, and ischemic heart disease: A narrative review of meta-analyses and a systematic review and meta-analysis of the impact of heavy drinking occasions on risk for moderate drinkers. BMC Med. 2014, 12, 182. [Google Scholar] [CrossRef] [Green Version]
- Clevidence, B.A.; Reichman, M.E.; Judd, J.T.; Muesing, R.A.; Schatzkin, A.; Schaefer, E.J.; Li, Z.; Jenner, J.; Brown, C.C.; Sunkin, M.; et al. Effects of alcohol consumption on lipoproteins of premenopausal women. A controlled diet study. Arter. Thromb. Vasc. Biol. 1995, 15, 179–184. [Google Scholar] [CrossRef]
- Marmillot, P.; Munoz, J.; Patel, S.; Garige, M.; Rosse, R.B.; Lakshman, M.R. Long-term ethanol consumption impairs reverse cholesterol transport function of high-density lipoproteins by depleting high-density lipoprotein sphingomyelin both in rats and in humans. Metabolism 2007, 56, 947–953. [Google Scholar] [CrossRef] [Green Version]
- Kovář, J.; Zemánková, K. Moderate alcohol consumption and triglyceridemia. Physiol. Res. 2015, 64, S371–S375. [Google Scholar] [CrossRef]
- Van de Wiel, A. The effect of alcohol on postprandial and fasting triglycerides. Int. J. Vasc. Med. 2012, 2012, 862504. [Google Scholar] [CrossRef] [Green Version]
- Ottestad, I.O.; Halvorsen, B.; Balstad, T.R.; Otterdal, K.; Borge, G.I.; Brosstad, F.; Myhre, A.M.; Ose, L.; Nenseter, M.S.; Holven, K.B. Triglyceride-rich HDL3 from patients with familial hypercholesterolemia are less able to inhibit cytokine release or to promote cholesterol efflux. J. Nutr. 2006, 136, 877–881. [Google Scholar] [CrossRef] [Green Version]
- Greene, D.J.; Skeggs, J.W.; Morton, R.E. Elevated triglyceride content diminishes the capacity of high density lipoprotein to deliver cholesteryl esters via the scavenger receptor class B type I (SR-BI). J. Biol. Chem. 2001, 276, 4804–4811. [Google Scholar] [CrossRef] [Green Version]
- Skeggs, J.W.; Morton, R.E. LDL and HDL enriched in triglyceride promote abnormal cholesterol transport. J. Lipid Res. 2002, 43, 1264–1274. [Google Scholar] [CrossRef]
- Ginsberg, H.N.; Packard, C.J.; Chapman, M.J.; Borén, J.; Aguilar-Salinas, C.A.; Averna, M.; Ference, B.A.; Gaudet, D.; Hegele, R.A.; Kersten, S.; et al. Triglyceride-rich lipoproteins and their remnants: Metabolic insights, role in atherosclerotic cardiovascular disease, and emerging therapeutic strategies-a consensus statement from the European Atherosclerosis Society. Eur. Heart J. 2021, 42, 4791–4806. [Google Scholar] [CrossRef]
- Hernandez-Hernandez, A.; Gea, A.; Ruiz-Canela, M.; Toledo, E.; Beunza, J.J.; Bes-Rastrollo, M.; Martinez-Gonzalez, M.A. Mediterranean Alcohol-Drinking Pattern and the Incidence of Cardiovascular Disease and Cardiovascular Mortality: The SUN Project. Nutrients 2015, 7, 9116–9126. [Google Scholar] [CrossRef] [Green Version]
- Institute of Medicine (US) Committee on Health and Behavior. Research, Practice, and Policy. In Health and Behavior: The Interplay of Biological, Behavioral, and Societal Influences; National Academies Press: Washington, DC, USA, 2001; ISBN 0-309-07030-9. [Google Scholar]
- Li, Y.; Pan, A.; Wang, D.D.; Liu, X.; Dhana, K.; Franco, O.H.; Kaptoge, S.; Di Angelantonio, E.; Stampfer, M.; Willett, W.C.; et al. Impact of Healthy Lifestyle Factors on Life Expectancies in the US Population. Circulation 2018, 138, 345–355. [Google Scholar] [CrossRef]
- Roberts, H.C.; Denison, H.J.; Martin, H.J.; Patel, H.P.; Syddall, H.; Cooper, C.; Sayer, A.A. A review of the measurement of grip strength in clinical and epidemiological studies:Towards a standardised approach. Age Ageing 2011, 40, 423–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havel, R.J.; Eder, H.A.; Bragdon, J.H. The distribution and chemical composition of ultracentrifugally separated lipoproteins in human serum. J. Clin. Investig. 1955, 34, 1345–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markwell, M.A.; Haas, S.M.; Bieber, L.L.; Tolbert, N.E. A modification of the lowry procedure to simplify protein determination in membrane and lipoprotein samples. Anal. Biochem. 1978, 87, 206–210. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Blatter Garin, M.C.; Moren, X.; James, R.W. Paraoxonase-1 and serum concentrations of HDL-cholesterol and apoA-I. J. Lipid Res. 2006, 47, 515–520. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.H.; Kim, J.R.; Lee, I.C.; Kwon, H.J. Native High-Density Lipoproteins (HDL) with Higher Paraoxonase Exerts a Potent Antiviral Effect against SARS-CoV-2 (COVID-19), While Glycated HDL Lost the Antiviral Activity. Antioxidants 2021, 10, 209. [Google Scholar] [CrossRef]
- Noble, R.P. Electrophoretic separation of plasma lipoproteins in agarose gel. J. Lipid Res. 1968, 9, 693–700. [Google Scholar] [CrossRef]
- Cho, K.H.; Kang, D.J.; Nam, H.S.; Kim, J.H.; Kim, S.Y.; Lee, J.O.; Kim, B.J. Ozonated Sunflower Oil Exerted Protective Effect for Embryo and Cell Survival via Potent Reduction Power and Antioxidant Activity in HDL with Strong Antimicrobial Activity. Antioxidants 2021, 10, 1651. [Google Scholar] [CrossRef]
- Park, K.H.; Kim, J.Y.; Choi, I.; Kim, J.R.; Won, K.C.; Cho, K.H. Fructated apolipoprotein A-I exacerbates cellular senescence in human umbilical vein endothelial cells accompanied by impaired insulin secretion activity and embryo toxicity. Biochem. Cell Biol. Biochim. et Biol. Cell. 2016, 94, 337–345. [Google Scholar] [CrossRef]
- McPherson, J.D.; Shilton, B.H.; Walton, D.J. Role of fructose in glycation and cross-linking of proteins. Biochemistry 1988, 27, 1901–1907. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group 1 (0 g) 1 n = 18 | Group 2 (21~105 g) 1 n = 20 | Group 3 (168~1680 g) 1 n = 16 | p | 1 vs. 2 | 1 vs. 3 | 2 vs. 3 | |
|---|---|---|---|---|---|---|---|
| Age (year-old), (min, max) | 53.0 (45.0; 58.0) (38, 64) | 52.0 (49.3; 55.8) (35, 62) | 49.5 (41.3; 51.8) (36, 58) | 0.163 | 1.000 | 0.208 | 0.427 |
| Menopausal (n) | 10 | 12 | 4 | ||||
| Alcohol intake amount (g/month) | 0 | 63 (21; 63) 1 | 315 (210; 560) 2 | <0.001 | 1.000 | <0.001 | <0.001 |
| Height (cm) | 157.5 (156.0; 162.3) | 160.5 (157.3; 162.8) | 162.0 (157.3; 166.8) | 0.147 | 1.000 | 0.174 | 0.468 |
| Weight (kg) | 60.3 (51.6; 62.6) | 55.5 (50.7; 60.6) | 58.6 (54.1; 61.1) | 0.611 | 1.000 | 1.000 | 1.000 |
| BMI (kg/m2), (min., max.) | 23.7 (20.5; 24.9) (19.2, 26.4) | 21.5 (20.2; 23.1) (18.7, 30.9) | 21.5 (21.0; 23.7) (17.0, 29.3) | 0.295 | 0.360 | 1.000 | 1.000 |
| Heart rate (BPM) | 76.0 (70.5; 86.3) | 73.5 (70.8; 80.5) | 72.0 (69.3; 82.5) | 0.518 | 1.000 | 0.856 | 1.000 |
| SBP (mmHg) | 123.5 (116.8; 132.0) | 119.0 (108.5; 123.3) | 123.5 (114.5; 135.8) | 0.044 | 0.090 | 1.000 | 0.103 |
| DBP (mmHg) | 68.5 (65.0; 77.0) | 70.0 (65.5; 79.5) | 77.0 (66.3; 84.0) | 0.433 | 1.000 | 0.985 | 0.650 |
| Muscle mass (kg) | 37.7 (34.9; 40.2) | 37.6 (34.7; 40.1) | 38.4 (36.3; 40.5) | 0.760 | 1.000 | 1.000 | 1.000 |
| Fat mass_subcutaneous (kg) | 16.6 (12.5; 17.2) | 13.6 (11.2; 16.5) | 14.3 (12.8; 16.2) | 0.584 | 0.967 | 1.000 | 1.000 |
| Fat mass_visceral (kg) | 2.0 (1.3; 2.2) | 1.5 (1.2; 1.9) | 1.6 (1.4; 2.1) | 0.341 | 0.430 | 1.000 | 1.000 |
| handgrip strength (HS, kg) (min., max.) | 27.5 (26.3; 29.3) (13, 33) | 26.5 (23.0; 28.8) (19, 32) | 26.0 (23.0; 29.0) (21, 31) | 0.374 | 0.572 | 0.828 | 1.000 |
| Body water (kg) | 29.6 (27.8; 31.5) | 29.4 (27.1; 31.3) | 30.2 (28.4; 31.6) | 0.650 | 1.000 | 1.000 | 1.000 |
| Group 1 (0 g) n = 18 | Group 2 (21~105 g) 1 n = 20 | Group 3 (168~1680 g) 2 n = 16 | p | 1 vs. 2 | 1 vs. 3 | 2 vs. 3 | |
|---|---|---|---|---|---|---|---|
| TC (mg/dL) | 219.5 (200.5; 239.2) | 232.1 (185.6; 245.9) | 216.4 (197.6; 246.8) | 0.947 | 1.000 | 1.000 | 1.000 |
| HDL-C (mg/dL) | 56.3 (47.6; 61.6) | 52.8 (47.7; 59.2) | 45.2 (38.5; 55.5) | 0.012 | 0.864 | 0.011 | 0.123 |
| HDL-C/TC (ratio) | 25.5 (21.8; 29.4) | 24.1 (21.3; 28.6) | 22.1 (17.6; 26.3) | 0.096 | 1.000 | 0.106 | 0.366 |
| LDL-C (mg/dL) | 138.0 (126.5; 173.5) | 158.5 (121.5; 174.5) | 146.5 (132.5; 176.3) | 0.864 | 1.000 | 1.000 | 1.000 |
| TG (mg/dL) | 84.5 (54.2; 104.6) | 67.0 (56.4; 95.0) | 108.2 (62.2; 167.4) | 0.170 | 1.000 | 0.667 | 0.186 |
| TG/HDL-C (ratio) | 1.6 (0.8; 2.3) | 1.2 (1.0; 1.7) | 2.4 (1.3; 3.6) | 0.088 | 1.000 | 0.273 | 0.105 |
| LDL-C/HDL-C (ratio) | 2.7 (2.1; 3.3) | 2.9 (2.3; 3.5) | 3.1 (2.7; 4.1) | 0.123 | 1.000 | 0.127 | 0.646 |
| ApoA-I (mg/dL) | 181.5 (169.5; 197.5) | 169.0 (156.0; 187.8) | 165.5 (145.3; 182.3) | 0.029 | 0.375 | 0.024 | 0.663 |
| Apo-B (mg/dL) | 101.5 (81.5; 126.3) | 100.0 (81.5; 112.5) | 107.5 (92.5; 130.5) | 0.249 | 0.933 | 1.000 | 0.307 |
| Apo-B/ApoA-I (ratio) | 0.6 (0.5; 0.6) | 0.6 (0.5; 0.7) | 0.7 (0.5; 0.9) | 0.081 | 1.000 | 0.110 | 0.207 |
| Glucose (mg/dL) | 93.0 (84.0; 97.3) | 90.0 (83.0; 100.8) | 103.0 (84.3; 100.5) | 0.999 | 1.000 | 1.000 | 1.000 |
| hs-CRP (mg/L) | 0.5 (0.3; 0.6) | 0.3 (0.2; 0.5) | 0.4 (0.2; 0.9) | 0.972 | 1.000 | 1.000 | 1.000 |
| AST (Unit/L) | 17.5 (13.0; 19.5) | 17.0 (14.0; 20.8) | 17.0 (13.3; 21.0) | 0.861 | 1.000 | 1.000 | 1.000 |
| ALT (Unit/L) | 13.5 (9.0; 17.0) | 12.0 (10.0; 16.0) | 12.5 (11.0; 16.0) | 0.828 | 1.000 | 1.000 | 1.000 |
| γ-GTP (Unit/L) | 11.0 (9.0; 14.0) | 11.5 (8.3; 14.8) | 15.0 (10.0; 33.5) | 0.105 | 1.000 | 0.161 | 0.221 |
| Group 1 (0 g) n = 18 | Group 2 (21~105 g) 1 n = 20 | Group 3 (168~1680 g) 2 n = 16 | p | 1 vs. 2 | 1 vs. 3 | 2 vs. 3 | |
|---|---|---|---|---|---|---|---|
| LDL-size (nm2) | 627 (487; 779) | 494 (445; 571) | 460 (390; 508) | <0.001 | 0.008 | <0.001 | 1.000 |
| LDL-MDA (nM) | 1.2 (0.9; 1.5) | 1.8 (1.3; 2.0) | 1.5 (1.3; 1.9) | 0.011 | 0.013 | 0.086 | 1.000 |
| LDL-glycation (FI) | 4236 (3825; 5007) | 4489 (4116; 5434) | 4568 (3632; 5547) | 0.710 | 1.000 | 1.000 | 1.000 |
| LDL-TC (mg/mg of protein) | 19.7 (15.3; 21.8) | 18.0 (15.1; 22.2) | 17.4 (15.1; 22.9) | 0.926 | 1.000 | 1.000 | 1.000 |
| LDL = TG (mg/mg of protein) | 2.5 (1.9; 3.4) | 3.4 (2.9; 4.8) | 3.8 (3.1; 4.3) | 0.004 | 0.027 | 0.005 | 1.000 |
| HDL2 -size (nm2) | 214 (160; 241) | 125 (98; 138) | 183 (144; 200) | <0.001 | <0.001 | 0.309 | 0.001 |
| HDL2-glycation (FI) | 1568 (1283; 1757) | 1512 (1415; 1709) | 1608 (1421; 1832) | 0.510 | 1.000 | 0.834 | 1.000 |
| HDL2-TC (mg/mg of protein) | 2.8 (1.8; 3.3) | 3.3 (2.1; 3.7) | 3.5 (2.7; 3.6) | 0.052 | 0.076 | 0.020 | 0.511 |
| HDL2-TG (mg/mg of protein) | 0.6 (0.4; 0.8) | 0.5 (0.3; 0.7) | 0.6 (0.5; 0.9) | 0.328 | 0.530 | 0.388 | 0.136 |
| HDL2-PON (Absorbance at 415 nm) | 0.247 (0.175; 0.412) | 0.183 (0.113; 0.261) | 0.237 (0.128; 0.326) | 0.021 | 0.007 | 0.048 | 0.582 |
| HDL3-size (nm2) | 111 (81; 142) | 92 (79; 102) | 81 (63; 100) | <0.001 | 0.031 | <0.001 | 0.822 |
| HDL3-TC (mg/mg of protein) | 2.8 (2.4; 4.0) | 3.1 (2.9; 4.0) | 3.1 (2.9; 3.5) | 0.463 | 0.741 | 1.000 | 1.000 |
| HDL3-TG (mg/mg of protein) | 0.3 (0.3; 0.4) | 0.5 (0.4; 0.6) | 0.6 (0.5; 0.9) | 0.002 | 0.032 | <0.001 | 0.118 |
| HDL3-PON (Absorbance at 415 nm) | 0.797 (0.661; 1.226) | 0.941 (0.598; 1.070) | 0.465 (0.274; 0.811) | 0.010 | 1.000 | 0.055 | 0.012 |
| r | p | ||
|---|---|---|---|
| Anthropometric profiles | BMI (kg/m2) | −0.135 | 0.329 |
| Fat mass_subcutaneous (kg) | −0.064 | 0.648 | |
| Fat mass_visceral (kg) | −0.097 | 0.485 | |
| SBP (mmHg) | −0.007 | 0.962 | |
| DBP (mmHg) | 0.152 | 0.272 | |
| HS (kg) | −0.156 | 0.261 | |
| Blood lipid andinflammatory parameters | HDL-C (mg/dL) | −0.363 | 0.007 |
| ApoA-I (mg/dL) | −0.365 | 0.007 | |
| ApoB (mg/dL) | 0.107 | 0.440 | |
| ApoB/ApoA-I (ratio) | 0.281 | 0.040 | |
| LDL-C (mg/dL) | 0.087 | 0.533 | |
| LDL-MDA (nM) | 0.315 | 0.020 | |
| TC (mg/dL) | 0.025 | 0.860 | |
| TG (mg/dL) | 0.157 | 0.257 | |
| TG/HDL-C (ratio) | 0.222 | 0.107 | |
| AST (Unit/L) | 0.045 | 0.745 | |
| ALT (Unit/L) | 0.018 | 0.896 | |
| γ-GTP (Unit/L) | 0.237 | 0.084 | |
| hs-CRP (mg/L) | −0.032 | 0.819 | |
| Lipoprotein profiles | LDL-TC (mg/mg of protein) | 0.438 | <0.001 |
| LDL-TG (mg/mg of protein) | 0.016 | 0.907 | |
| HDL2-TC (mg/mg of protein) | 0.325 | 0.017 | |
| HDL2-TG (mg/mg of protein) | 0.153 | 0.270 | |
| HDL3-TC (mg/mg of protein) | 0.205 | 0.153 | |
| HDL3-TG (mg/mg of protein) | 0.500 | <0.001 | |
| HDL3-PON (AU) | −0.346 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, K.-H.; Nam, H.-S.; Kang, D.-J.; Park, M.-H.; Kim, J.-H. Long-Term Alcohol Consumption Caused a Significant Decrease in Serum High-Density Lipoprotein (HDL)-Cholesterol and Apolipoprotein A-I with the Atherogenic Changes of HDL in Middle-Aged Korean Women. Int. J. Mol. Sci. 2022, 23, 8623. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23158623
Cho K-H, Nam H-S, Kang D-J, Park M-H, Kim J-H. Long-Term Alcohol Consumption Caused a Significant Decrease in Serum High-Density Lipoprotein (HDL)-Cholesterol and Apolipoprotein A-I with the Atherogenic Changes of HDL in Middle-Aged Korean Women. International Journal of Molecular Sciences. 2022; 23(15):8623. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23158623
Chicago/Turabian StyleCho, Kyung-Hyun, Hyo-Seon Nam, Dae-Jin Kang, Min-Hee Park, and Ju-Hyun Kim. 2022. "Long-Term Alcohol Consumption Caused a Significant Decrease in Serum High-Density Lipoprotein (HDL)-Cholesterol and Apolipoprotein A-I with the Atherogenic Changes of HDL in Middle-Aged Korean Women" International Journal of Molecular Sciences 23, no. 15: 8623. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23158623