
MicroRNA in Acromegaly: Involvement in the Pathogenesis and in the Response to First-Generation Somatostatin Receptor Ligands

 , , and
, , and
Abstract
:1. Introduction
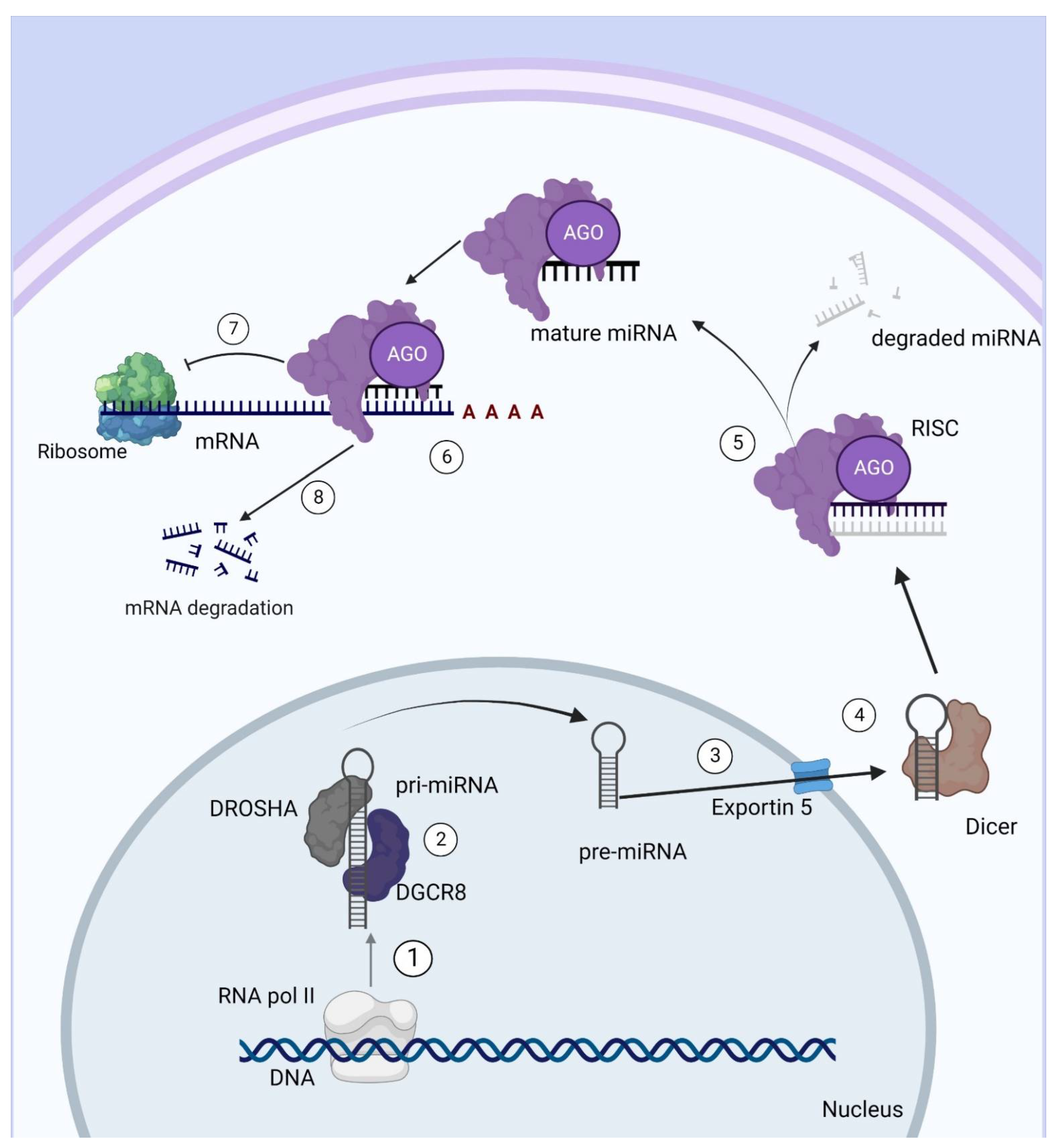
2. miRNAs: Definition and Mechanism of Action
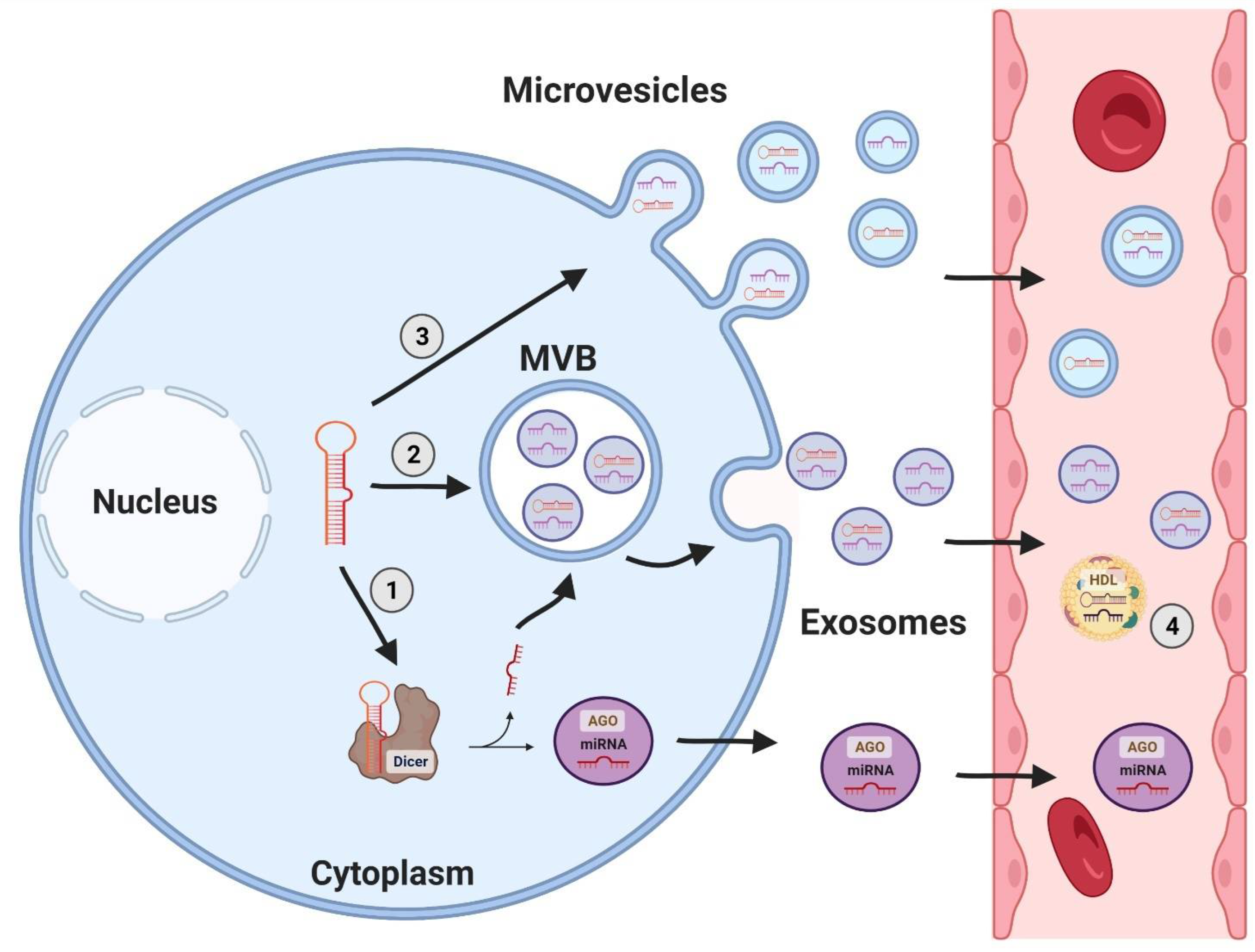
3. miRNA as Circulating Biomarkers of Disease
4. miRNAs in Acromegaly
4.1. miRNAs Involved in Tumorigenesis
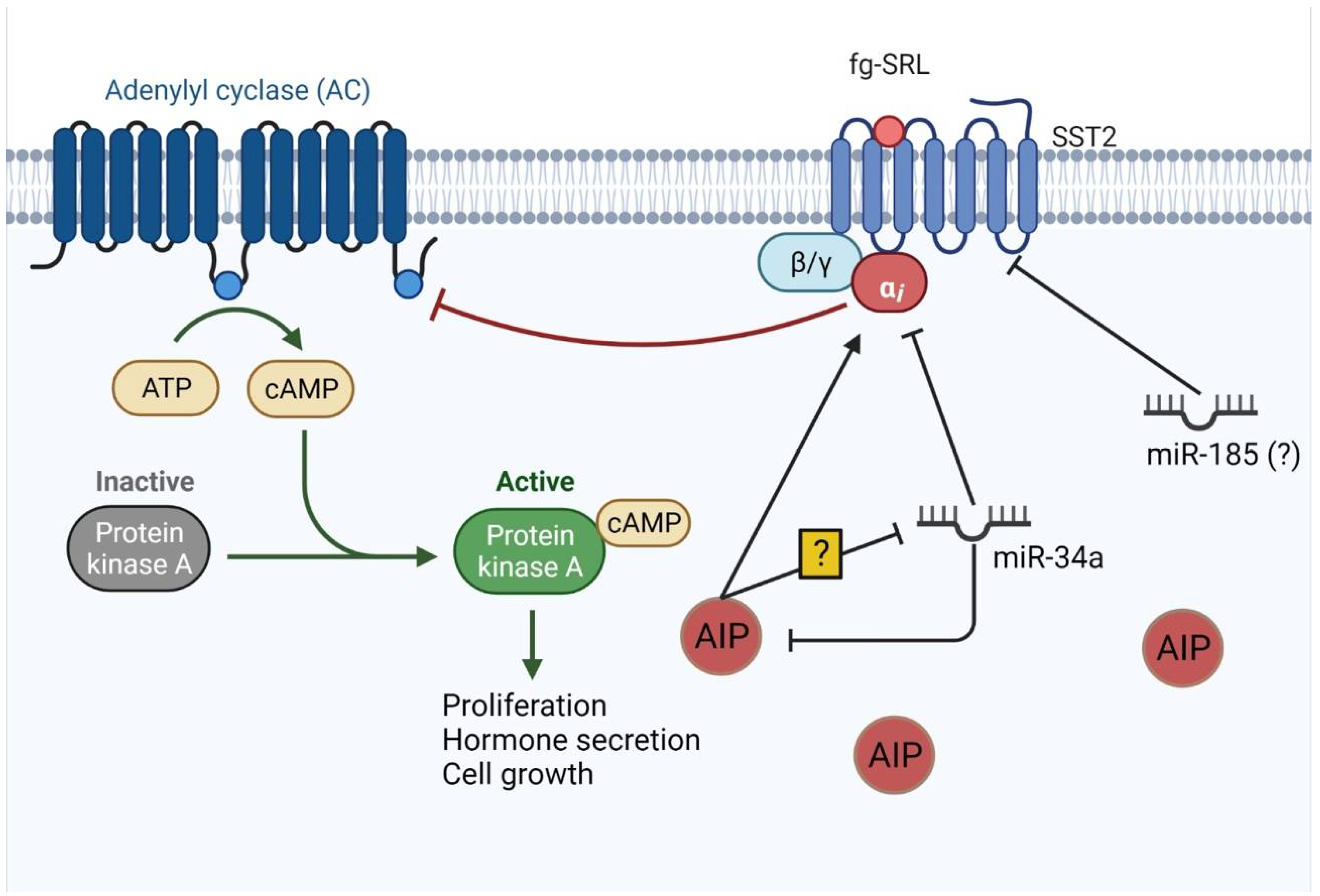
4.2. miRNAs Involved in Treatment Resistance
5. Circulating miRNAs in Acromegaly
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Etxabe, J.; Gaztambide, S.; Latorre, P.; Vazquez, J.A. Acromegaly: An epidemiological study. J. Endocrinol. Investig. 1993, 16, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Kasuki, L.; Rocha, P.D.S.; Lamback, E.B.; Gadelha, M.R. Determinants of morbidities and mortality in acromegaly. Arch. Endocrinol. Metab. 2019, 63, 630–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasuki, L.; Antunes, X.; Lamback, E.B.; Gadelha, M.R. Acromegaly: Update on Management and Long-Term Morbidities. Endocrinol. Metab. Clin. N. Am. 2020, 49, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Gadelha, M.R.; Kasuki, L.; Lim, D.S.T.; Fleseriu, M. Systemic Complications of Acromegaly and the Impact of the Current Treatment Landscape: An Update. Endocr. Rev. 2019, 40, 268–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleseriu, M.; Biller, B.M.K.; Freda, P.U.; Gadelha, M.R.; Giustina, A.; Katznelson, L.; Molitch, M.E.; Samson, S.L.; Strasburger, C.J.; van der Lely, A.J.; et al. A Pituitary Society update to acromegaly management guidelines. Pituitary 2021, 24, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Melmed, S.; Colao, A.; Barkan, A.; Molitch, M.; Grossman, A.B.; Kleinberg, D.; Clemmons, D.; Chanson, P.; Laws, E.; Schlechte, J.; et al. Guidelines for acromegaly management: An update. J. Clin. Endocrinol. Metab. 2009, 94, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Antunes, X.; Ventura, N.; Camilo, G.B.; Wildemberg, L.E.; Guasti, A.; Pereira, P.J.M.; Camacho, A.H.S.; Chimelli, L.; Niemeyer, P.; Gadelha, M.R.; et al. Predictors of surgical outcome and early criteria of remission in acromegaly. Endocrine 2018, 60, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Casar-Borota, O.; Heck, A.; Schulz, S.; Nesland, J.M.; Ramm-Pettersen, J.; Lekva, T.; Alafuzoff, I.; Bollerslev, J. Expression of SSTR2a, but not of SSTRs 1, 3, or 5 in somatotroph adenomas assessed by monoclonal antibodies was reduced by octreotide and correlated with the acute and long-term effects of octreotide. J. Clin. Endocrinol. Metab. 2013, 98, E1730–E1739. [Google Scholar] [CrossRef] [Green Version]
- Ferone, D.; de Herder, W.W.; Pivonello, R.; Kros, J.M.; van Koetsveld, P.M.; de Jong, T.; Minuto, F.; Colao, A.; Lamberts, S.W.; Hofland, L.J. Correlation of in vitro and in vivo somatotropic adenoma responsiveness to somatostatin analogs and dopamine agonists with immunohistochemical evaluation of somatostatin and dopamine receptors and electron microscopy. J. Clin. Endocrinol. Metab. 2008, 93, 1412–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadelha, M.R.; Wildemberg, L.E.; Bronstein, M.D.; Gatto, F.; Ferone, D. Somatostatin receptor ligands in the treatment of acromegaly. Pituitary 2017, 20, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Wildemberg, L.E.; da Silva Camacho, A.H.; Miranda, R.L.; Elias, P.C.L.; de Castro Musolino, N.R.; Nazato, D.; Jallad, R.; Huayllas, M.K.P.; Mota, J.I.S.; Almeida, T.; et al. Machine Learning-based Prediction Model for Treatment of Acromegaly with First-generation Somatostatin Receptor Ligands. J. Clin. Endocrinol. Metab. 2021, 106, 2047–2056. [Google Scholar] [CrossRef] [PubMed]
- Wildemberg, L.E.; Gadelha, M.R. Pasireotide for the treatment of acromegaly. Expert Opin. Pharmacother. 2016, 17, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Iacovazzo, D.; Carlsen, E.; Lugli, F.; Chiloiro, S.; Piacentini, S.; Bianchi, A.; Giampietro, A.; Mormando, M.; Clear, A.J.; Doglietto, F.; et al. Factors predicting pasireotide responsiveness in somatotroph pituitary adenomas resistant to first-generation somatostatin analogues: An immunohistochemical study. Eur. J. Endocrinol. 2016, 174, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Gadelha, M.R.; Bronstein, M.D.; Brue, T.; Coculescu, M.; Fleseriu, M.; Guitelman, M.; Pronin, V.; Raverot, G.; Shimon, I.; Lievre, K.K.; et al. Pasireotide versus continued treatment with octreotide or lanreotide in patients with inadequately controlled acromegaly (PAOLA): A randomised, phase 3 trial. Lancet Diabetes Endocrinol. 2014, 2, 875–884. [Google Scholar] [CrossRef]
- Gadelha, M.R.; Wildemberg, L.E.; Kasuki, L. The Future of Somatostatin Receptor Ligands in Acromegaly. J. Clin. Endocrinol. Metab. 2022, 107, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Shimon, I.; Saeger, W.; Wildemberg, L.E.; Gadelha, M.R. Somatotropinomas inadequately controlled with octreotide may over-respond to pasireotide: The importance of dose adjustment to achieve long-term biochemical control. Hormones 2017, 16, 84–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Efstathiadou, Z.A.; Bargiota, A.; Chrisoulidou, A.; Kanakis, G.; Papanastasiou, L.; Theodoropoulou, A.; Tigas, S.K.; Vassiliadi, D.A.; Alevizaki, M.; Tsagarakis, S. Impact of gsp mutations in somatotroph pituitary adenomas on growth hormone response to somatostatin analogs: A meta-analysis. Pituitary 2015, 18, 861–867. [Google Scholar] [CrossRef]
- Wildemberg, L.E.; Henriques, D.; Elias, P.C.L.; Lima, C.H.A.; Musolino, N.R.C.; Camacho, A.H.S.; Faria, O.; Nazato, D.; Abucham, J.; Vilar, L.; et al. Mutation Is Not a Molecular Biomarker of Long-Term Response to First-Generation Somatostatin Receptor Ligands in Acromegaly. Cancers 2021, 13, 4857. [Google Scholar] [CrossRef] [PubMed]
- Caimari, F.; Hernández-Ramírez, L.C.; Dang, M.N.; Gabrovska, P.; Iacovazzo, D.; Stals, K.; Ellard, S.; Korbonits, M.; Consortium, I.F. Risk category system to identify pituitary adenoma patients with. J. Med. Genet. 2018, 55, 254–260. [Google Scholar] [CrossRef] [Green Version]
- Araujo, P.B.; Kasuki, L.; de Azeredo Lima, C.H.; Ogino, L.; Camacho, A.H.S.; Chimelli, L.; Korbonits, M.; Gadelha, M.R. Mutations in Brazilian patients with sporadic pituitary adenomas: A single-center evaluation. Endocr. Connect. 2017, 6, 914–925. [Google Scholar] [CrossRef] [Green Version]
- Cazabat, L.; Bouligand, J.; Salenave, S.; Bernier, M.; Gaillard, S.; Parker, F.; Young, J.; Guiochon-Mantel, A.; Chanson, P. Germline AIP mutations in apparently sporadic pituitary adenomas: Prevalence in a prospective single-center cohort of 443 patients. J. Clin. Endocrinol. Metab. 2012, 97, E663–E670. [Google Scholar] [CrossRef]
- Cuny, T.; Pertuit, M.; Sahnoun-Fathallah, M.; Daly, A.; Occhi, G.; Odou, M.F.; Tabarin, A.; Nunes, M.L.; Delemer, B.; Rohmer, V.; et al. Genetic analysis in young patients with sporadic pituitary macroadenomas: Besides AIP don’t forget MEN1 genetic analysis. Eur. J. Endocrinol. 2013, 168, 533–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadelha, M.R.; Trivellin, G.; Hernández Ramírez, L.C.; Korbonits, M. Genetics of pituitary adenomas. Front. Horm. Res. 2013, 41, 111–140. [Google Scholar] [CrossRef]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Vishnoi, A.; Rani, S. MiRNA Biogenesis and Regulation of Diseases: An Overview. Methods Mol. Biol. 2017, 1509, 1–10. [Google Scholar] [CrossRef]
- Bartel, D.P. Metazoan MicroRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Jeon, K.; Lee, J.T.; Kim, S.; Kim, V.N. MicroRNA maturation: Stepwise processing and subcellular localization. EMBO J. 2002, 21, 4663–4670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadelha, M.R.; Kasuki, L.; Denes, J.; Trivellin, G.; Korbonits, M. MicroRNAs: Suggested role in pituitary adenoma pathogenesis. J. Endocrinol. Investig. 2013, 36, 889–895. [Google Scholar] [CrossRef] [PubMed]
- Michlewski, G.; Cáceres, J.F. Post-transcriptional control of miRNA biogenesis. RNA 2019, 25, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.K.; Kim, B.; Kim, V.N. Re-evaluation of the roles of DROSHA, Export in 5, and DICER in microRNA biogenesis. Proc. Natl. Acad. Sci. USA 2016, 113, E1881–E1889. [Google Scholar] [CrossRef] [Green Version]
- Okamura, K.; Hagen, J.W.; Duan, H.; Tyler, D.M.; Lai, E.C. The mirtron pathway generates microRNA-class regulatory RNAs in Drosophila. Cell 2007, 130, 89–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, B.; Zhao, Z.; Cai, Q.; Zhang, Y.; Zhang, P.; Shi, S.; Xie, H.; Peng, X.; Yin, W.; Tao, Y.; et al. miRNA-based biomarkers, therapies, and resistance in Cancer. Int. J. Biol. Sci. 2020, 16, 2628–2647. [Google Scholar] [CrossRef] [PubMed]
- Correia de Sousa, M.; Gjorgjieva, M.; Dolicka, D.; Sobolewski, C.; Foti, M. Deciphering miRNAs’ Action through miRNA Editing. Int. J. Mol. Sci. 2019, 20, 6249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef]
- Lujambio, A.; Lowe, S.W. The microcosmos of cancer. Nature 2012, 482, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbach, H.; Nishida, N.; Calin, G.A.; Pantel, K. Clinical relevance of circulating cell-free microRNAs in cancer. Nat. Rev. Clin. Oncol. 2014, 11, 145–156. [Google Scholar] [CrossRef]
- Kubo, H. Extracellular Vesicles in Lung Disease. Chest 2018, 153, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Takousis, P.; Sadlon, A.; Schulz, J.; Wohlers, I.; Dobricic, V.; Middleton, L.; Lill, C.M.; Perneczky, R.; Bertram, L. Differential expression of microRNAs in Alzheimer’s disease brain, blood, and cerebrospinal fluid. Alzheimers Dement. 2019, 15, 1468–1477. [Google Scholar] [CrossRef] [PubMed]
- Street, J.M.; Koritzinsky, E.H.; Glispie, D.M.; Yuen, P.S.T. Urine Exosome Isolation and Characterization. Methods Mol. Biol. 2017, 1641, 413–423. [Google Scholar] [CrossRef]
- El-Hefnawy, T.; Raja, S.; Kelly, L.; Bigbee, W.L.; Kirkwood, J.M.; Luketich, J.D.; Godfrey, T.E. Characterization of amplifiable, circulating RNA in plasma and its potential as a tool for cancer diagnostics. Clin. Chem. 2004, 50, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Lawrie, C.H.; Gal, S.; Dunlop, H.M.; Pushkaran, B.; Liggins, A.P.; Pulford, K.; Banham, A.H.; Pezzella, F.; Boultwood, J.; Wainscoat, J.S.; et al. Detection of elevated levels of tumour-associated microRNAs in serum of patients with diffuse large B-cell lymphoma. Br. J. Haematol. 2008, 141, 672–675. [Google Scholar] [CrossRef] [PubMed]
- Rovas, A.; Puriene, A.; Snipaitiene, K.; Punceviciene, E.; Buragaite-Staponkiene, B.; Matuleviciute, R.; Butrimiene, I.; Jarmalaite, S. Analysis of periodontitis-associated miRNAs in gingival tissue, gingival crevicular fluid, saliva and blood plasma. Arch. Oral Biol. 2021, 126, 105125. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Butz, H.; Kinga, N.; Racz, K.; Patocs, A. Circulating miRNAs as biomarkers for endocrine disorders. J. Endocrinol. Investig. 2016, 39, 1–10. [Google Scholar] [CrossRef]
- Agbu, P.; Carthew, R.W. MicroRNA-mediated regulation of glucose and lipid metabolism. Nat. Rev. Mol. Cell Biol. 2021, 22, 425–438. [Google Scholar] [CrossRef] [PubMed]
- Chim, S.S.; Shing, T.K.; Hung, E.C.; Leung, T.Y.; Lau, T.K.; Chiu, R.W.; Lo, Y.M. Detection and characterization of placental microRNAs in maternal plasma. Clin. Chem. 2008, 54, 482–490. [Google Scholar] [CrossRef] [Green Version]
- Marzi, M.J.; Ghini, F.; Cerruti, B.; de Pretis, S.; Bonetti, P.; Giacomelli, C.; Gorski, M.M.; Kress, T.; Pelizzola, M.; Muller, H.; et al. Degradation dynamics of microRNAs revealed by a novel pulse-chase approach. Genome Res. 2016, 26, 554–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Li, Z.; Zhou, S.; Wen, J.; Geng, B.; Yang, J.; Cui, Q. Genome-wide analysis of human microRNA stability. Biomed. Res. Int. 2013, 2013, 368975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; et al. Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahadi, A. A systematic review of microRNAs as potential biomarkers for diagnosis and prognosis of gastric cancer. Immunogenetics 2021, 73, 155–161. [Google Scholar] [CrossRef]
- Ganju, A.; Khan, S.; Hafeez, B.B.; Behrman, S.W.; Yallapu, M.M.; Chauhan, S.C.; Jaggi, M. miRNA nanotherapeutics for cancer. Drug Discov. Today 2017, 22, 424–432. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.J.; Cho, J.M.; Moon, J.H.; Ku, C.R.; Kim, J.; Kim, S.H.; Lee, E.J. Increased miR-338-3p expression correlates with invasiveness of GH-producing pituitary adenomas. Endocrine 2017, 58, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.G.; He, D.S.; Zhou, J.; Yao, B.; Xiao, W.W.; Chen, C.H.; Zhu, Y.H.; Wang, H.J. Differential expression of microRNAs in GH-secreting pituitary adenomas. Diagn. Pathol. 2010, 5, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palumbo, T.; Faucz, F.R.; Azevedo, M.; Xekouki, P.; Iliopoulos, D.; Stratakis, C.A. Functional screen analysis reveals miR-26b and miR-128 as central regulators of pituitary somatomammotrophic tumor growth through activation of the PTEN-AKT pathway. Oncogene 2013, 32, 1651–1659. [Google Scholar] [CrossRef] [Green Version]
- D’Angelo, D.; Palmieri, D.; Mussnich, P.; Roche, M.; Wierinckx, A.; Raverot, G.; Fedele, M.; Croce, C.M.; Trouillas, J.; Fusco, A. Altered microRNA expression profile in human pituitary GH adenomas: Down-regulation of miRNA targeting HMGA1, HMGA2, and E2F1. J. Clin. Endocrinol. Metab. 2012, 97, E1128–E1138. [Google Scholar] [CrossRef]
- Bottoni, A.; Piccin, D.; Tagliati, F.; Luchin, A.; Zatelli, M.C.; degli Uberti, E.C. miR-15a and miR-16-1 down-regulation in pituitary adenomas. J. Cell. Physiol. 2005, 204, 280–285. [Google Scholar] [CrossRef]
- Monsalves, E.; Juraschka, K.; Tateno, T.; Agnihotri, S.; Asa, S.L.; Ezzat, S.; Zadeh, G. The PI3K/AKT/mTOR pathway in the pathophysiology and treatment of pituitary adenomas. Endocr. Relat. Cancer 2014, 21, R331–R344. [Google Scholar] [CrossRef] [Green Version]
- Salehi, F.; Kovacs, K.; Scheithauer, B.W.; Lloyd, R.V.; Cusimano, M. Pituitary tumor-transforming gene in endocrine and other neoplasms: A review and update. Endocr. Relat. Cancer 2008, 15, 721–743. [Google Scholar] [CrossRef]
- Zhao, S.; Li, J.; Feng, J.; Li, Z.; Liu, Q.; Lv, P.; Wang, F.; Gao, H.; Zhang, Y. Identification of Serum miRNA-423-5p Expression Signature in Somatotroph Adenomas. Int. J. Endocrinol. 2019, 2019, 8516858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammadi-Yeganeh, S.; Hosseini, V.; Paryan, M. Wnt pathway targeting reduces triple-negative breast cancer aggressiveness through miRNA regulation in vitro and in vivo. J. Cell. Physiol. 2019, 234, 18317–18328. [Google Scholar] [CrossRef] [PubMed]
- Xuan, Y.W.; Liao, M.; Zhai, W.L.; Peng, L.J.; Tang, Y. MicroRNA-381 inhibits lung adenocarcinoma cell biological progression by directly targeting LMO3 through regulation of the PI3K/Akt signaling pathway and epithelial-to-mesenchymal transition. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8411–8421. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Cao, Z.; Luo, W.; Zheng, L.; Zhang, T. MicroRNA-381-A Key Transcriptional Regulator: Its Biological Function and Clinical Application Prospects in Cancer. Front. Oncol. 2020, 10, 535665. [Google Scholar] [CrossRef] [PubMed]
- Denduluri, S.K.; Idowu, O.; Wang, Z.; Liao, Z.; Yan, Z.; Mohammed, M.K.; Ye, J.; Wei, Q.; Wang, J.; Zhao, L.; et al. Insulin-like growth factor (IGF) signaling in tumorigenesis and the development of cancer drug resistance. Genes Dis. 2015, 2, 13–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, Z.R.; Asa, S.L.; Siomi, H.; Siomi, M.C.; Yoshimoto, K.; Yamada, S.; Wang, E.L.; Rahman, M.M.; Inoue, H.; Itakura, M.; et al. Overexpression of HMGA2 relates to reduction of the let-7 and its relationship to clinicopathological features in pituitary adenomas. Mod. Pathol. 2009, 22, 431–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Mao, Z.G.; Wang, X.; Du, Q.; Jian, M.; Zhu, D.; Xiao, Z.; Wang, H.J.; Zhu, Y.H. MicroRNAs and Target Genes in Pituitary Adenomas. Horm. Metab. Res. 2018, 50, e3. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wang, J.; Xiang, H.; Ding, P.; Wu, T.; Ji, G. The biochemical and clinical implications of phosphatase and tensin homolog deleted on chromosome ten in different cancers. Am. J. Cancer Res. 2021, 11, 5833–5855. [Google Scholar]
- Wang, R.; Fan, H.; Sun, M.; Lv, Z.; Yi, W. Roles of BMI1 in the Initiation, Progression, and Treatment of Hepatocellular Carcinoma. Technol. Cancer Res. Treat. 2022, 21, 15330338211070689. [Google Scholar] [CrossRef]
- Tanabe, S.; Quader, S.; Cabral, H.; Ono, R. Interplay of EMT and CSC in Cancer and the Potential Therapeutic Strategies. Front. Pharmacol. 2020, 11, 904. [Google Scholar] [CrossRef]
- Jiang, N.; Dai, Q.; Su, X.; Fu, J.; Feng, X.; Peng, J. Role of PI3K/AKT pathway in cancer: The framework of malignant behavior. Mol. Biol. Rep. 2020, 47, 4587–4629. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.; Chellappan, D.K.; de Jesus Andreoli Pinto, T.; Hansbro, P.M.; Bebawy, M.; Dua, K. Tumor suppressor role of miR-503. Panminerva Med. 2018, 60, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Xie, P.; Han, Q.; Liu, D.; Yao, D.; Lu, X.; Wang, Z.; Zuo, X. miR-525-5p Modulates Proliferation and Epithelial-Mesenchymal Transition of Glioma by Targeting Stat-1. OncoTargets Ther. 2020, 13, 9957–9966. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Liu, L.X. MiR-525-5p Repressed Metastasis and Anoikis Resistance in Cervical Cancer via Blocking UBE2C/ZEB1/2 Signal Axis. Dig. Dis. Sci. 2020, 65, 2442–2451. [Google Scholar] [CrossRef] [PubMed]
- Zatelli, M.C.; degli Uberti, E.C. MicroRNAs and possible role in pituitary adenoma. Semin. Reprod. Med. 2008, 26, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, W.; Yan, W.; Han, L.; Zhang, K.; Shi, Z.; Zhang, J.; Wang, Y.; Li, Y.; Yu, S.; et al. The putative tumor suppressor miR-524-5p directly targets Jagged-1 and Hes-1 in glioma. Carcinogenesis 2012, 33, 2276–2282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yavropoulou, M.P.; Maladaki, A.; Yovos, J.G. The role of Notch and Hedgehog signaling pathways in pituitary development and pathogenesis of pituitary adenomas. Hormones 2015, 14, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Zhang, Y.; Zhang, T. MiR-524-5p Suppresses Migration, Invasion, and EMT Progression in Breast Cancer Cells through Targeting. Cancer Biother. Radiopharm. 2020, 35, 789–801. [Google Scholar] [CrossRef]
- Li, X.; Li, Z.; Zhu, Y.; Yao, L.; Zhang, L.; Yuan, L.; Shang, Y.; Liu, J.; Li, C. miR-524-5p inhibits angiogenesis through targeting WNK1 in colon cancer cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 318, G827–G839. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Shoorei, H.; Bahroudi, Z.; Abak, A.; Majidpoor, J.; Taheri, M. An update on the role of miR-124 in the pathogenesis of human disorders. Biomed. Pharmacother. 2021, 135, 111198. [Google Scholar] [CrossRef]
- Zhang, E.; Liu, S.; Xu, Z.; Huang, S.; Tan, X.; Sun, C.; Lu, L. Pituitary tumor-transforming gene 1 (PTTG1) is overexpressed in oral squamous cell carcinoma (OSCC) and promotes migration, invasion and epithelial-mesenchymal transition (EMT) in SCC15 cells. Tumour Biol. 2014, 35, 8801–8811. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhao, Q.; Yu, S.; Lin, R.; Yi, X. Pttg1 inhibits TGFβ signaling in breast cancer cells to promote their growth. Tumour Biol. 2015, 36, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Horwitz, G.A.; Heaney, A.P.; Nakashima, M.; Prezant, T.R.; Bronstein, M.D.; Melmed, S. Pituitary tumor transforming gene (PTTG) expression in pituitary adenomas. J. Clin. Endocrinol. Metab. 1999, 84, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Trivellin, G.; Butz, H.; Delhove, J.; Igreja, S.; Chahal, H.S.; Zivkovic, V.; McKay, T.; Patócs, A.; Grossman, A.B.; Korbonits, M. MicroRNA miR-107 is overexpressed in pituitary adenomas and inhibits the expression of aryl hydrocarbon receptor-interacting protein in vitro. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E708–E719. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez-Costa, A.; Korbonits, M. AIP and the somatostatin system in pituitary tumours. J. Endocrinol. 2017, 235, R101–R116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denes, J.; Kasuki, L.; Trivellin, G.; Colli, L.M.; Takiya, C.M.; Stiles, C.E.; Barry, S.; de Castro, M.; Gadelha, M.R.; Korbonits, M. Regulation of aryl hydrocarbon receptor interacting protein (AIP) protein expression by MiR-34a in sporadic somatotropinomas. PLoS ONE 2015, 10, e0117107. [Google Scholar] [CrossRef]
- Bogner, E.M.; Daly, A.F.; Gulde, S.; Karhu, A.; Irmler, M.; Beckers, J.; Mohr, H.; Beckers, A.; Pellegata, N.S. miR-34a is upregulated in AIP-mutated somatotropinomas and promotes octreotide resistance. Int. J. Cancer 2020, 147, 3523–3538. [Google Scholar] [CrossRef]
- Wildemberg, L.E.; Neto, L.V.; Costa, D.F.; Nasciuti, L.E.; Takiya, C.M.; Alves, L.M.; Rebora, A.; Minuto, F.; Ferone, D.; Gadelha, M.R. Low somatostatin receptor subtype 2, but not dopamine receptor subtype 2 expression predicts the lack of biochemical response of somatotropinomas to treatment with somatostatin analogs. J. Endocrinol. Investig. 2013, 36, 38–43. [Google Scholar] [CrossRef]
- Fan, X.; Mao, Z.; He, D.; Liao, C.; Jiang, X.; Lei, N.; Hu, B.; Wang, X.; Li, Z.; Lin, Y.; et al. Expression of somatostatin receptor subtype 2 in growth hormone-secreting pituitary adenoma and the regulation of miR-185. J. Endocrinol. Investig. 2015, 38, 1117–1128. [Google Scholar] [CrossRef] [PubMed]
- Khoshnevisan, A.; Parvin, M.; Ghorbanmehr, N.; Hatefi, N.; Galehdari, H.; Ziaee, S.A.; Mowla, S.J. A Significant Upregulation of miR-886-5p in High Grade and Invasive Bladder Tumors. Urol. J. 2015, 12, 2160–2164. [Google Scholar]
- Li, J.H.; Xiao, X.; Zhang, Y.N.; Wang, Y.M.; Feng, L.M.; Wu, Y.M.; Zhang, Y.X. MicroRNA miR-886-5p inhibits apoptosis by down-regulating Bax expression in human cervical carcinoma cells. Gynecol. Oncol. 2011, 120, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Xiang, P.; Liu, Y.; Liu, L.; Lin, Q.; Liu, X.; Zhang, H.; Xu, J.; Fang, B. The Biological Function and Clinical Significance of miR-886-5p in Multiple Myeloma. Acta Haematol. 2019, 142, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Kasuki, L.; Wildemberg, L.E.; Gadelha, M.R. Management of Endocrine Disease: Personalized medicine in the treatment of acromegaly. Eur. J. Endocrinol. 2018, 178, R89–R100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korkmaz, H.; Hekimler Öztürk, K.; Torus, B. Circulating miR-29c-3p is downregulated in patients with acromegaly. Turk. J. Med. Sci. 2021, 51, 2081–2086. [Google Scholar] [CrossRef] [PubMed]
- Lutsenko, A.; Belaya, Z.; Nikitin, A.; Solodovnikov, A.; Lapshina, A.; Koshkin, P.; Vorontsova, M.; Rozhinskaya, L.; Melnichenko, G.; Dedov, I. Circulating Plasma MicroRNA in Patients with Active Acromegaly. J. Clin. Endocrinol. Metab. 2022, 107, 500–511. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Wei, R.; Li, H.; Dou, Y.; Wang, J. miR-452-5p and miR-215-5p expression levels in colorectal cancer tissues and their relationship with clinicopathological features. Oncol. Lett. 2020, 20, 2955–2961. [Google Scholar] [CrossRef]
- Kasuki, L.; Maia, B.; Gadelha, M.R. Acromegaly and Colorectal Neoplasm: An Update. Front. Endocrinol. 2022, 13, 924952. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Pathways/Proteins | miRNAs | miRNA Expression | Findings | References |
|---|---|---|---|---|
| PTTG | miR-126 miR-381 miR-338-3p | Downregulated Downregulated Upregulated | Proliferation Invasion | [54,55] |
| PTEN/PI3K/AKT/mTOR | miR-26b miR-128 | Downregulated Upregulated | Tumor size Invasion | [55,56] |
| EMT | miR-503 miR-525-5p | Downregulated Upregulated | Differentiation, proliferation Proliferation, invasion, migration | [55] |
| HMGA1/2 | miR-34b miR-548c-3p miR-34b miR-326 miR-432 miR-548c-3p miR-570 | Downregulated | Proliferation | [57] |
| E2F1 | miR-326 miR-603 | Downregulated | Proliferation | [57] |
| RARS | miR-16-1 | Downregulated | Inverse correlation with tumor size | [58] |
| FGF2 | miR-503 | Downregulated | Differentiation, proliferation | [55] |
| miRNA | Target | miRNA Expression | Finding | Reference |
|---|---|---|---|---|
| miR-34a | AIP | Upregulated | Poor response to fg-SRL | [86,87] |
| miR-185 | SST2 | Upregulated | Poor response to fg-SRL | [89] |
| miR-125a-5p miR-524-5p | IGFBP-3 IGFALS chain precursor MMP-9 | Downregulated | Better response to fg-SRL | [55] |
| miR-886-5p | P53/Bax pathway | Upregulated | Better response to fg-SRL | [55] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henriques, D.G.; Lamback, E.B.; Dezonne, R.S.; Kasuki, L.; Gadelha, M.R. MicroRNA in Acromegaly: Involvement in the Pathogenesis and in the Response to First-Generation Somatostatin Receptor Ligands. Int. J. Mol. Sci. 2022, 23, 8653. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23158653
Henriques DG, Lamback EB, Dezonne RS, Kasuki L, Gadelha MR. MicroRNA in Acromegaly: Involvement in the Pathogenesis and in the Response to First-Generation Somatostatin Receptor Ligands. International Journal of Molecular Sciences. 2022; 23(15):8653. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23158653
Chicago/Turabian StyleHenriques, Daniel G., Elisa B. Lamback, Romulo S. Dezonne, Leandro Kasuki, and Monica R. Gadelha. 2022. "MicroRNA in Acromegaly: Involvement in the Pathogenesis and in the Response to First-Generation Somatostatin Receptor Ligands" International Journal of Molecular Sciences 23, no. 15: 8653. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23158653