
Role of a ZF-HD Transcription Factor in miR157-Mediated Feed-Forward Regulatory Module That Determines Plant Architecture in Arabidopsis


Abstract
:1. Introduction
2. Results
2.1. ZF-HD TFs Regulate Plant Architecture in Arabidopsis
2.2. ZF-HD TFs Regulate Shoot Architecture in a Dose-Dependent Manner
2.3. ZF-HD TFs Act as Central Regulators to Control Expression of Multiple TF Families
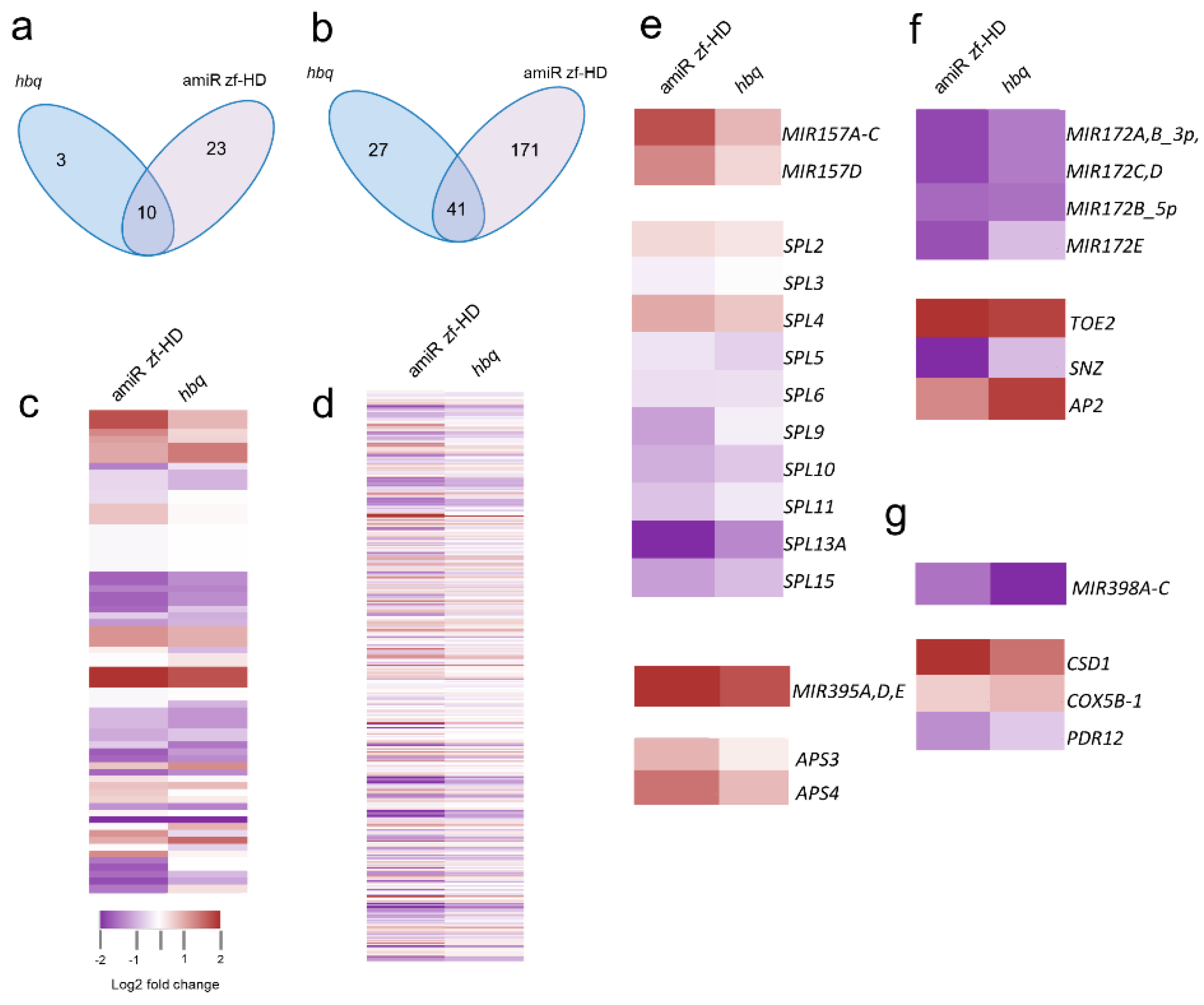
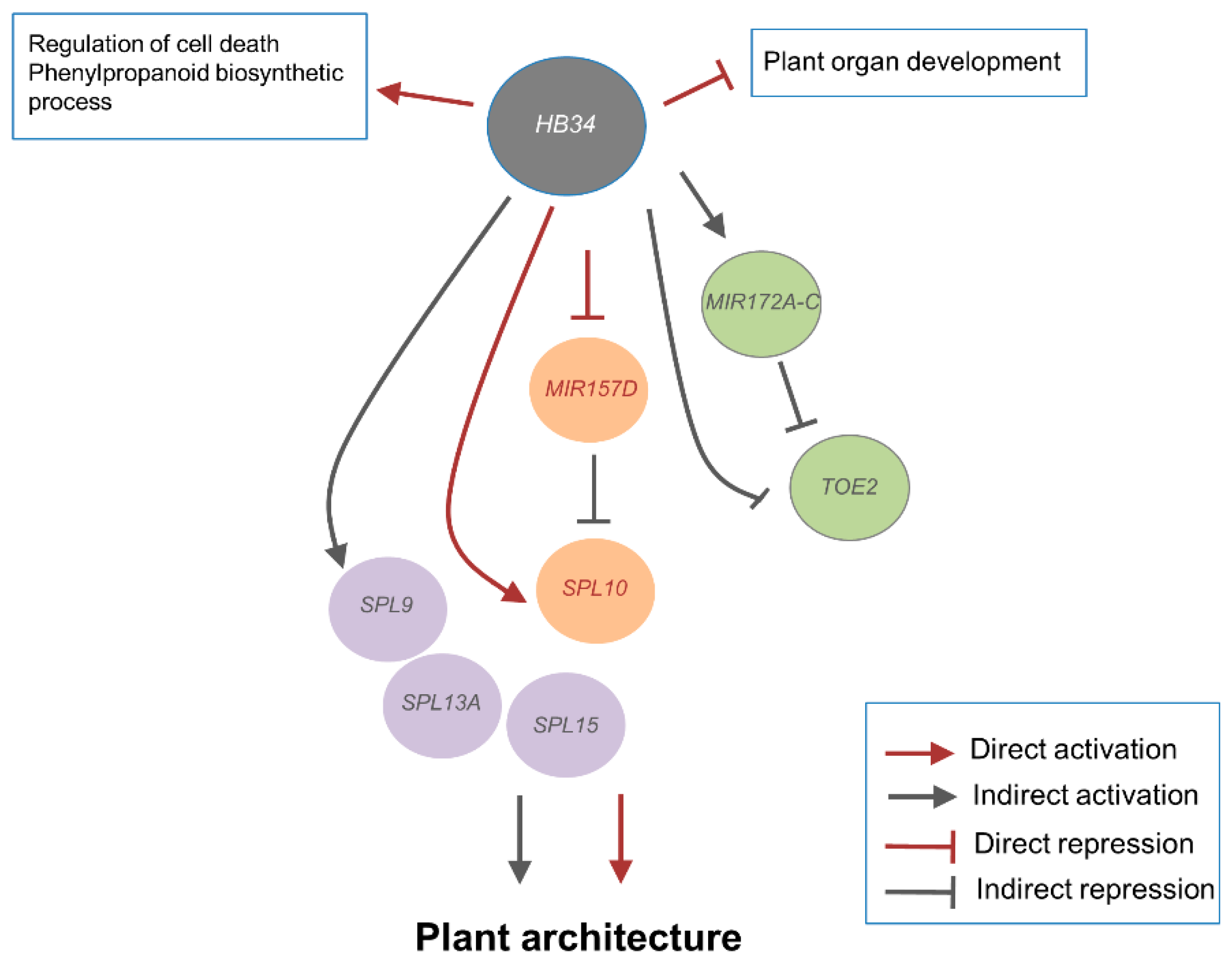
2.4. ZF-HD Gene Members Positively Regulate SPL Genes in a Feed-Forward Loop (FFL) via miR157
2.5. Genome-Wide Analysis of HB34 Binding
3. Discussion
4. Materials and Methods
4.1. Plant Growth and Mutant Collection
4.2. Localization of ZF-HD TF and Tobacco Transient Assay
4.3. Gene Expression and Sequence Analysis
4.4. miRNA Expression Analysis by TaqMan PCR
4.5. RNA-seq Data and smRNA Library Data Analysis
4.6. ChIP-seq Analysis
4.7. Motif Prediction Analysis
4.8. Network Motif Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Abbreviation | Definition |
| amiRNA | artificial miRNA |
| ChIP-seq | chromatin immunoprecipitation sequencing |
| CUC | CUP SHAPED COTYLEDONS |
| FFL | feed-forward loop |
| GRF | GROWTH-REGULATING FACTOR |
| GRNs | gene regulatory networks |
| qRT-PCR | quantitative Reverse Transcription polymerase chain reaction |
| REV | REVOLUTA |
| smRNA | small RNA |
| SPL | SQUAMOSA PROMOTER BINDING–LIKE |
| STM | SHOOT MERISTEMLESS |
| TFs | transcription factors |
| ZF-HD | Zinc finger homeodomain (ZF-HD) |
References
- Shimizu-Sato, S.; Mori, H. Control of Outgrowth and Dormancy in Axillary Buds. Plant Physiol. 2001, 127, 1405–1413. [Google Scholar] [CrossRef]
- McSteen, P.; Leyser, O. Shoot Branching. Annu. Rev. Plant Biol. 2005, 56, 353–374. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Barton, M.K. Initiation of Axillary and Floral Meristems in Arabidopsis. Dev. Biol. 2000, 218, 341–353. [Google Scholar] [CrossRef] [PubMed]
- González-Grandío, E.; Pajoro, A.; Franco-Zorrilla, J.M.; Tarancón, C.; Immink, R.G.H.; Cubas, P. Abscisic Acid Signaling Is Controlled by a BRANCHED1/HD-ZIP I Cascade in Arabidopsis Axillary Buds. Proc. Natl. Acad. Sci. USA 2017, 114, E245–E254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hibara, K.; Karim, R.; Takada, S.; Taoka, K.; Furutani, M.; Aida, M.; Tasaka, M. Arabidopsis CUP-SHAPED COTYLEDON3 Regulates Postembryonic Shoot Meristem and Organ Boundary Formation. Plant Cell 2006, 18, 2946–2957. [Google Scholar] [CrossRef] [Green Version]
- Raman, S.; Greb, T.; Peaucelle, A.; Blein, T.; Laufs, P.; Theres, K. Interplay of MiR164, CUP-SHAPED COTYLEDON Genes and LATERAL SUPPRESSOR Controls Axillary Meristem Formation in Arabidopsis thaliana. Plant J. 2008, 55, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-K.; Geisler, M.; Springer, P.S. LATERAL ORGAN FUSION1 and LATERAL ORGAN FUSION2 Function in Lateral Organ Separation and Axillary Meristem Formation in Arabidopsis. Development 2009, 136, 2423–2432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, J.; Tian, F.; Yang, D.-C.; Meng, Y.-Q.; Kong, L.; Luo, J.; Gao, G. PlantTFDB 4.0: Toward a Central Hub for Transcription Factors and Regulatory Interactions in Plants. Nucleic Acids Res. 2017, 45, D1040–D1045. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Ma, H. Characterization of a Novel Putative Zinc Finger Gene MIF1: Involvement in Multiple Hormonal Regulation of Arabidopsis Development. Plant J. 2006, 45, 399–422. [Google Scholar] [CrossRef]
- Wang, H.; Yin, X.; Li, X.; Wang, L.; Zheng, Y.; Xu, X.; Zhang, Y.; Wang, X. Genome-Wide Identification, Evolution and Expression Analysis of the Grape (Vitis vinifera L.) Zinc Finger-Homeodomain Gene Family. Int. J. Mol. Sci. 2014, 15, 5730–5748. [Google Scholar] [CrossRef] [Green Version]
- Tran, L.-S.P.; Nakashima, K.; Sakuma, Y.; Osakabe, Y.; Qin, F.; Simpson, S.D.; Maruyama, K.; Fujita, Y.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Co-Expression of the Stress-Inducible Zinc Finger Homeodomain ZFHD1 and NAC Transcription Factors Enhances Expression of the ERD1 Gene in Arabidopsis. Plant J. 2007, 49, 46–63. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, D.D.; Barros, P.M.; Cordeiro, A.M.; Serra, T.S.; Lourenço, T.; Chander, S.; Oliveira, M.M.; Saibo, N.J.M. Seven Zinc-Finger Transcription Factors Are Novel Regulators of the Stress Responsive Gene OsDREB1B. J. Exp. Bot. 2012, 63, 3643–3656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Q.K.-G.; Irish, V.F. The Arabidopsis Zinc Finger-Homeodomain Genes Encode Proteins with Unique Biochemical Properties That Are Coordinately Expressed during Floral Development. Plant Physiol. 2006, 140, 1095–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.-Y.; Kim, O.-K.; Kim, S.-G.; Yang, M.-S.; Park, C.-M. Nuclear Import and DNA Binding of the ZHD5 Transcription Factor Is Modulated by a Competitive Peptide Inhibitor in Arabidopsis. J. Biol. Chem. 2011, 286, 1659–1668. [Google Scholar] [CrossRef] [Green Version]
- Windhövel, A.; Hein, I.; Dabrowa, R.; Stockhaus, J. Characterization of a Novel Class of Plant Homeodomain Proteins That Bind to the C4 Phosphoenolpyruvate Carboxylase Gene of Flaveria trinervia. Plant Mol. Biol. 2001, 45, 201–214. [Google Scholar] [CrossRef]
- Liu, M.; Wang, X.; Sun, W.; Ma, Z.; Zheng, T.; Huang, L.; Wu, Q.; Tang, Z.; Bu, T.; Li, C.; et al. Genome-Wide Investigation of the ZF-HD Gene Family in Tartary Buckwheat (Fagopyrum tataricum). BMC Plant Biol. 2019, 19, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sicard, A.; Hernould, M.; Chevalier, C. The Inhibitor of Meristem Activity (IMA) Protein: The Nexus between Cell Division, Differentiation and Hormonal Control of Development. Plant Signal Behav. 2008, 3, 908–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arabidopsis Interactome Mapping Consortium Evidence for Network Evolution in an Arabidopsis Interactome Map. Science 2011, 333, 601–607. [CrossRef] [Green Version]
- Rhoades, M.W.; Reinhart, B.J.; Lim, L.P.; Burge, C.B.; Bartel, B.; Bartel, D.P. Prediction of Plant MicroRNA Targets. Cell 2002, 110, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Sieber, P.; Wellmer, F.; Gheyselinck, J.; Riechmann, J.L.; Meyerowitz, E.M. Redundancy and Specialization among Plant MicroRNAs: Role of the MIR164 Family in Developmental Robustness. Development 2007, 134, 1051–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Poethig, R.S. Temporal Regulation of Shoot Development in Arabidopsis thaliana by MiR156 and Its Target SPL3. Development 2006, 133, 3539–3547. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Cheng, X.; Liu, P.; Sun, J. MiR156-Targeted SBP-Box Transcription Factors Interact with DWARF53 to Regulate TEOSINTE BRANCHED1 and BARREN STALK1 Expression in Bread Wheat. Plant Physiol. 2017, 174, 1931–1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, C.; Ye, M.; Sang, M.; Wu, R. A Regulatory Network for MiR156-SPL Module in Arabidopsis thaliana. Int. J. Mol. Sci. 2019, 20, 6166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.; Shi, J.; Li, Z.; Zhang, S.; Zhang, S.; Zhang, J.; Bao, M.; Liu, G. MiR156/157 Targets SPLs to Regulate Flowering Transition, Plant Architecture and Flower Organ Size in Petunia. Plant Cell Physiol. 2021, 62, 839–857. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Wang, J.; Yang, X.; Lu, H.; Miao, X.; Shi, Z. Modulation of Plant Architecture by the MiR156f–OsSPL7–OsGH3.8 Pathway in Rice. J. Exp. Bot. 2018, 69, 5117–5130. [Google Scholar] [CrossRef] [Green Version]
- Chuck, G.; Cigan, A.M.; Saeteurn, K.; Hake, S. The Heterochronic Maize Mutant Corngrass1 Results from Overexpression of a Tandem MicroRNA. Nat. Genet. 2007, 39, 544–549. [Google Scholar] [CrossRef]
- Preston, J.C.; Jorgensen, S.A.; Orozco, R.; Hileman, L.C. Paralogous SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) Genes Differentially Regulate Leaf Initiation and Reproductive Phase Change in Petunia. Planta 2016, 243, 429–440. [Google Scholar] [CrossRef] [Green Version]
- Gao, R.; Wang, Y.; Gruber, M.Y.; Hannoufa, A. MiR156/SPL10 Modulates Lateral Root Development, Branching and Leaf Morphology in Arabidopsis by Silencing AGAMOUS-LIKE 79. Front. Plant Sci. 2018, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Allen, E.; Fahlgren, N.; Calamar, A.; Givan, S.A.; Carrington, J.C. Expression of Arabidopsis MIRNA Genes. Plant Physiol. 2005, 138, 2145–2154. [Google Scholar] [CrossRef] [Green Version]
- Cuperus, J.T.; Fahlgren, N.; Carrington, J.C. Evolution and Functional Diversification of MIRNA Genes. Plant Cell 2011, 23, 431–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, K.; Chen, X. Biogenesis, Turnover, and Mode of Action of Plant MicroRNAs. Plant Cell 2013, 25, 2383–2399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, M.; Kim, V.N. Regulation of MicroRNA Biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Hauser, F.; Chen, W.; Deinlein, U.; Chang, K.; Ossowski, S.; Fitz, J.; Hannon, G.J.; Schroeder, J.I. A Genomic-Scale Artificial MicroRNA Library as a Tool to Investigate the Functionally Redundant Gene Space in Arabidopsis. Plant Cell 2013, 25, 2848–2863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, P.; Hirsch-Hoffmann, M.; Hennig, L.; Gruissem, W. GENEVESTIGATOR. Arabidopsis Microarray Database and Analysis Toolbox. Plant Physiol. 2004, 136, 2621–2632. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Brandizzi, F.; Benning, C.; Larkin, R.M. A Membrane-Tethered Transcription Factor Defines a Branch of the Heat Stress Response in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2008, 105, 16398–16403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, S.-F.; Lai, H.-C.; Jinn, T.-L. Cytosol-Localized Heat Shock Factor-Binding Protein, AtHSBP, Functions as a Negative Regulator of Heat Shock Response by Translocation to the Nucleus and Is Required for Seed Development in Arabidopsis. Plant Physiol. 2010, 153, 773–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riechmann, J.L.; Krizek, B.A.; Meyerowitz, E.M. Dimerization Specificity of Arabidopsis MADS Domain Homeotic Proteins APETALA1, APETALA3, PISTILLATA, and AGAMOUS. Proc. Natl. Acad. Sci. USA 1996, 93, 4793–4798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, X.; Vega-Léon, R.; Hugouvieux, V.; Blanc-Mathieu, R.; van der Wal, F.; Lucas, J.; Silva, C.S.; Jourdain, A.; Muino, J.M.; Nanao, M.H.; et al. The Intervening Domain Is Required for DNA-Binding and Functional Identity of Plant MADS Transcription Factors. Nat. Commun. 2021, 12, 4760. [Google Scholar] [CrossRef] [PubMed]
- Mantegazza, O.; Gregis, V.; Chiara, M.; Selva, C.; Leo, G.; Horner, D.S.; Kater, M.M. Gene Coexpression Patterns during Early Development of the Native Arabidopsis Reproductive Meristem: Novel Candidate Developmental Regulators and Patterns of Functional Redundancy. Plant J. 2014, 79, 861–877. [Google Scholar] [CrossRef]
- Kozomara, A.; Griffiths-Jones, S. MiRBase: Annotating High Confidence MicroRNAs Using Deep Sequencing Data. Nucleic Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef] [Green Version]
- Alon, U. Network Motifs: Theory and Experimental Approaches. Nat. Rev. Genet. 2007, 8, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Mangan, S.; Zaslaver, A.; Alon, U. The Coherent Feedforward Loop Serves as a Sign-Sensitive Delay Element in Transcription Networks. J. Mol. Biol. 2003, 334, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Hamant, O.; Pautot, V. Plant Development: A TALE Story. Comptes Rendus Biol. 2010, 333, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Onghena, P. Randomization Tests or Permutation Tests? A Historical and Terminological Clarification. In Randomization, Masking, and Allocation Concealment; Chapman and Hall: London, UK; CRC: Boca Raton, FL, USA, 2017; ISBN 978-1-315-30511-0. [Google Scholar]
- Riechmann, J.L.; Meyerowitz, E.M. Determination of Floral Organ Identity by Arabidopsis MADS Domain Homeotic Proteins AP1, AP3, PI, and AG Is Independent of Their DNA-Binding Specificity. Mol. Biol. Cell 1997, 8, 1243–1259. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Yasuno, N.; Sato, Y.; Yoda, M.; Yamazaki, R.; Kimizu, M.; Yoshida, H.; Nagamura, Y.; Kyozuka, J. Inflorescence Meristem Identity in Rice Is Specified by Overlapping Functions of Three AP1/FUL-like MADS Box Genes and PAP2, a SEPALLATA MADS Box Gene. Plant Cell 2012, 24, 1848–1859. [Google Scholar] [CrossRef] [Green Version]
- Uno, Y.; Furihata, T.; Abe, H.; Yoshida, R.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis Basic Leucine Zipper Transcription Factors Involved in an Abscisic Acid-Dependent Signal Transduction Pathway under Drought and High-Salinity Conditions. Proc. Natl. Acad. Sci. USA 2000, 97, 11632–11637. [Google Scholar] [CrossRef] [Green Version]
- Garcia, M.E.; Lynch, T.; Peeters, J.; Snowden, C.; Finkelstein, R. A Small Plant-Specific Protein Family of ABI Five Binding Proteins (AFPs) Regulates Stress Response in Germinating Arabidopsis Seeds and Seedlings. Plant Mol. Biol. 2008, 67, 643–658. [Google Scholar] [CrossRef]
- Yang, J.; Worley, E.; Udvardi, M. A NAP-AAO3 Regulatory Module Promotes Chlorophyll Degradation via ABA Biosynthesis in Arabidopsis Leaves. Plant Cell 2014, 26, 4862–4874. [Google Scholar] [CrossRef] [Green Version]
- Nowak, K.; Morończyk, J.; Grzyb, M.; Szczygieł-Sommer, A.; Gaj, M.D. MiR172 Regulates WUS during Somatic Embryogenesis in Arabidopsis via AP2. Cells 2022, 11, 718. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Woo, H.R.; Kim, J.; Lim, P.O.; Lee, I.C.; Choi, S.H.; Hwang, D.; Nam, H.G. Trifurcate Feed-Forward Regulation of Age-Dependent Cell Death Involving MiR164 in Arabidopsis. Science 2009, 323, 1053–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serivichyaswat, P.; Ryu, H.-S.; Kim, W.; Kim, S.; Chung, K.S.; Kim, J.J.; Ahn, J.H. Expression of the Floral Repressor MiRNA156 Is Positively Regulated by the AGAMOUS-like Proteins AGL15 and AGL18. Mol. Cells 2015, 38, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Gehring, W.J.; Affolter, M.; Bürglin, T. Homeodomain Proteins. Annu. Rev. Biochem. 1994, 63, 487–526. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, R.C.; Huang, S.-S.C.; Song, L.; Lewsey, M.G.; Bartlett, A.; Nery, J.R.; Galli, M.; Gallavotti, A.; Ecker, J.R. Cistrome and Epicistrome Features Shape the Regulatory DNA Landscape. Cell 2016, 165, 1280–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Karimi, M.; Depicker, A.; Hilson, P. Recombinational Cloning with Plant Gateway Vectors. Plant Physiol. 2007, 145, 1144–1154. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Naithani, S.; Tello-Ruiz, M.K.; Chougule, K.; D’Eustachio, P.; Fabregat, A.; Jiao, Y.; Keays, M.; Lee, Y.K.; Kumari, S.; et al. Gramene Database: Navigating Plant Comparative Genomics Resources. Curr. Plant Biol. 2016, 7, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Varkonyi-Gasic, E.; Wu, R.; Wood, M.; Walton, E.F.; Hellens, R.P. Protocol: A Highly Sensitive RT-PCR Method for Detection and Quantification of MicroRNAs. Plant Methods 2007, 3, 12. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering Splice Junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript Assembly and Quantification by RNA-Seq Reveals Unannotated Transcripts and Isoform Switching during Cell Differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Huber, W. Differential Expression Analysis for Sequence Count Data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and Memory-Efficient Alignment of Short DNA Sequences to the Human Genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Z.; Zhou, X.; Ling, Y.; Zhang, Z.; Su, Z. AgriGO: A GO Analysis Toolkit for the Agricultural Community. Nucleic Acids Res. 2010, 38, W64–W70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowler, C.; Benvenuto, G.; Laflamme, P.; Molino, D.; Probst, A.V.; Tariq, M.; Paszkowski, J. Chromatin Techniques for Plant Cells. Plant J. 2004, 39, 776–789. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, T.; Meyer, C.A.; Eeckhoute, J.; Johnson, D.S.; Bernstein, B.E.; Nusbaum, C.; Myers, R.M.; Brown, M.; Li, W.; et al. Model-Based Analysis of ChIP-Seq (MACS). Genome Biol. 2008, 9, R137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machanick, P.; Bailey, T.L. MEME-ChIP: Motif Analysis of Large DNA Datasets. Bioinformatics 2011, 27, 1696–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.D.; Sumazin, P.; Xuan, Z.; Zhang, M.Q. DNA Motifs in Human and Mouse Proximal Promoters Predict Tissue-Specific Expression. Proc. Natl. Acad. Sci. USA 2006, 103, 6275–6280. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Zhao, P.X. PsRNATarget: A Plant Small RNA Target Analysis Server. Nucleic Acids Res. 2011, 39, W155–W159. [Google Scholar] [CrossRef] [Green Version]
- Tello-Ruiz, M.K.; Naithani, S.; Gupta, P.; Olson, A.; Wei, S.; Preece, J.; Jiao, Y.; Wang, B.; Chougule, K.; Garg, P.; et al. Gramene 2021: Harnessing the Power of Comparative Genomics and Pathways for Plant Research. Nucleic Acids Res. 2021, 49, D1452–D1463. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA–Target Feed-Forward Motif | Number of Loops in Real Network (Average Number of Loops in Randomized Network) | Number of Loops in Real Network in p-Value | ||
|---|---|---|---|---|
| mutant | amiR zf-HD | hbq | amiR zf-HD | hbq |
| Incoherent type I | 20 (13.54) | 3 (4.0769) | NS (0.1182) | NS (0.7195) |
| Incoherent type II | 21 (15.9115) | 0 (2.3681) | NS (0.1930) | NS (1.0000) |
| Coherent type III | 20 (15.8256) | 5 (3.8105) | NS (0.2298) | NS (0.3346) |
| Coherent type IV | 41 (13.6795) | 14 (2.5448) | *** (0.0001) | *** (0.0002) |
| Total | 102 (58.9525) | 22 (12.8003) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.K.; Kumari, S.; Olson, A.; Hauser, F.; Ware, D. Role of a ZF-HD Transcription Factor in miR157-Mediated Feed-Forward Regulatory Module That Determines Plant Architecture in Arabidopsis. Int. J. Mol. Sci. 2022, 23, 8665. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23158665
Lee YK, Kumari S, Olson A, Hauser F, Ware D. Role of a ZF-HD Transcription Factor in miR157-Mediated Feed-Forward Regulatory Module That Determines Plant Architecture in Arabidopsis. International Journal of Molecular Sciences. 2022; 23(15):8665. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23158665
Chicago/Turabian StyleLee, Young Koung, Sunita Kumari, Andrew Olson, Felix Hauser, and Doreen Ware. 2022. "Role of a ZF-HD Transcription Factor in miR157-Mediated Feed-Forward Regulatory Module That Determines Plant Architecture in Arabidopsis" International Journal of Molecular Sciences 23, no. 15: 8665. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23158665